
еЊвЊ: ФПЕФ ЙлВьP3ыФЖдRAW264.7ОоЪЩЯИАћжЌжЪГСЛ§ЕФгАЯьЃЌВЂЬНЬжЦфзїгУЛњжЦЁЃЗНЗЈ ВЩгУMTTЗЈЩИбЁP3ыФМАбѕЛЏЕЭУмЖШжЌЕААз(oxidized low density lipoproteinЃЌox-LDL)ЕФзїгУХЈЖШЃЌВЂгУ80 mgЁЄL-1ЕФox-LDLгеЕМRAW264.7ЯИАћаЮГЩХнФЯИАћЃЛЗжБ№ВЩгУгЭКьOШОЩЋКЭзмЕЈЙЬДМКЌСПВтЖЈЪдМСКаЃЌМьВтЯИАћФкжЌжЪГСЛ§МАзмЕЈЙЬДМКЌСПЃЛЪЕЪБгЋЙтЖЈСПPCR(quantitative real-time PCRЃЌqRT-PCR)МАWestern blotМьВтШ§СзЫсЯймеНсКЯКазЊдЫЬхA1(ATP-binding cassette transporter A1ЃЌABCA1)ЁЂШ§СзЫсЯймеНсКЯКазЊдЫЬхG1(ATP-binding cassette transporter G1ЃЌABCG1)ЕФmRNAКЭЕААзБэДяБфЛЏЃЛЗжБ№гІгУИЮXЪмЬх(liver X receptorЃЌLXR)ЕФМЄЖЏМСT0901317ЁЂGW3965ЃЌвдМАLXRвжжЦМСGSK2033НјвЛВНбщжЄP3ыФЕїПиABCA1ЁЂABCG1БэДяЕФзїгУЛњжЦЁЃНсЙћ P3ыФУїЯдМѕЩйRAW264.7ЯИАћФкжЌжЪГСЛ§ЃЌНЕЕЭЯИАћФкзмЕЈЙЬДМКЌСПЃЌЩЯЕїABCA1ЁЂABCG1mRNAКЭЕААзБэДяЃЛP3ыФЩЯЕїABCA1ЁЂABCG1ЕФзїгУгыLXRМЄЖЏМСT0901317ЁЂGW3965ЯрЫЦЃЛМгШывжжЦLXRЛљвђзЊТМЕФвжжЦМСGSK2033КѓЃЌP3ыФЖдABCA1ЁЂABCG1ЕФЛљвђБэДяЮоЩЯЕїзїгУЁЃНсТл P3ыФПЩМѕЩйRAW264.7ОоЪЩЯИАћФкжЌжЪЛ§ОлЃЌвжжЦХнФЯИАћЕФаЮГЩЃЌЦфзїгУЛњжЦПЩФмгыМЄЛюLXR-ABCA1/ABCG1ЭЈТЗгаЙиЁЃ
ЖЏТіжрбљгВЛЏ(AtherosclerosisЃЌAS)ЪЧвЛжжбЊЙмТ§адбзжЂадМВВЁЃЌЪЧаФФдбЊЙмМВВЁКЭЭтжмбЊЙмМВВЁЕФВЁРэЛљДЁЁЃASВЁРэАпПщжївЊгЩжЌжЪЁЂЯЫЮЌКЭЯИАћзщГЩЁЃОоЪЩЯИАћКЭTЯИАћЪЧВЮгыЖЏТігВЛЏАпПщаЮГЩгыЗЂеЙЕФжївЊЯИАћРраЭ[1]ЁЃASЕФЦ№вђЪЧбЊНЌжаКЌгадижЌЕААзB(apolipoprotein BЃЌapoB)ЕФжЌЕААзБЛЮќЪеВЂГСЛ§дкЖЏТіФкФЄЩЯЃЌжЌЕААзжаЕФЕЭУмЖШжЌЕААз(low density lipoproteinЃЌLDL)ПЩБЛбѕЛЏЛђЭЈЙ§ЦфЫћаоЪЮЖјгеЕМЖЏТіФкФЄЩЯЕФФкЦЄЯИАћЛюЛЏЃЌЛюЛЏЕФФкЦЄЯИАћФММЏЕЅКЫЯИАћЁЃвЛЗНУцЃЌЕЅКЫЯИАћЗжЛЏГЩДйбзОоЪЩЯИАћЃЌОжВПЗХДѓбзжЂЗДгІЃЛСэвЛЗНУцЃЌОоЪЩЯИАћЭЬЪЩаоЪЮЕФLDLЃЌаЮГЩХнФЯИАћЁЃЖЏТіФкФЄХнФЯИАћЕФГіЯжЪЧASАпПщаЮГЩжаВЛПЩШБЩйЕФвЛВНЃЌЭЈГЃБЛШЯЮЊЪЧASЕФдчЦкБэЯжжЎвЛЁЃХнФЯИАћЕФаЮГЩжївЊЪЧОоЪЩЯИАћЩуШЁЙ§СПаоЪЮЕФLDLЛђОоЪЩЯИАћФкЕЈЙЬДМСїГіеЯАЫљжТЃЌвђДЫЃЌвжжЦОоЪЩЯИАћЖдЕЈЙЬДМЕФЩуШЁЁЂДйНјОоЪЩЯИАћФкЕЈЙЬДМЕФСїГіЃЌПЩМѕЩйОоЪЩЯИАћФкЕЈЙЬДМЕФГСЛ§ЃЌЗРжЙХнФЯИАћЕФаЮГЩЃЌЖдгкИЩдЄдчЦкASОпгаживЊвтвх[2]ЁЃдкОоЪЩЯИАћжаЃЌШ§СзЫсЯймеНсКЯКазЊдЫЬхA1(ATP-binding cassette transporter A1ЃЌABCA1)КЭШ§СзЫсЯймеНсКЯКазЊдЫЬхG1(ATP-binding cassette transporter G1ЃЌABCG1)ИКд№НщЕМОоЪЩЯИАћФкЕЈЙЬДМСїГіЁЃABCA1КЭABCG1ЛљвђБэДяЩЯЕїПЩДйНјОоЪЩЯИАћФкЕЈЙЬДМСїГіЃЌвжжЦХнФЯИАћЕФаЮГЩ[3]ЁЃ
ПрмёЪЧоЄПЦ(Polygonaceae)мёТѓЪє(Fagopyrum)ЕФвЛФъЩњВнБОжВЮяЃЌЗжВМгкбЧжоЁЂХЗжоМАУРжоЃЌдкжаЙњжївЊжжжВдкИпКЎЕФЮїББКЭЮїФЯЩНЧјЁЃПрмёгЊбјМлжЕЗсИЛЃЌжаЙњвЛжБгаНЋПрмёЗлзїЮЊЙІФмадЪГЦЗЛђГЃЙцвћЪГЃЌгУгкжЮСЦаФбЊЙмМВВЁКЭЬЧФђВЁЕФДЋЭГЁЃПрмёжаЕФПрмёЕААзЃЌвВОпгадЄЗРИпжЌбЊжЂЕФзїгУЁЃгабаОПЗЂЯжЃЌПрмёЕААзПЩУїЯдНЕЕЭВжЪѓбЊНЌзмЕЈЙЬДМЫЎЦНЃЌЦфаЇЙћгХгкДѓУзЕААзКЭаЁТѓЕААз[4]ЁЃP3ыФРДдДгкПрмёЧхЕААзЃЌЪЧПЮЬтзщЧАЦкВЩгУМюадЕААзУИУИНтПрмёЧхЕААзЃЌЗжРыМјЖЈГіЕФЩњЮяЛюадыФЃЌОпгаПЙбѕЛЏМАЕїНкбзжЂЕФзїгУЃЌЦфАБЛљЫсађСаЮЊAla-Phe-Tyr-Arg-TrpЃЌЗжзгСПЮЊ741.8 u[5-6]ЁЃБОбаОПвдОоЪЩЯИАћRAW264.7ЮЊбаОПЖдЯѓЃЌВЩгУбѕЛЏЕЭУмЖШжЌЕААз(oxidized low density lipoproteinЃЌox-LDL)геЕМХнФЯИАћаЮГЩЃЌЙлВьP3ыФЖдОоЪЩЯИАћжажЌжЪГСЛ§ЕФгАЯьЃЌВЂЬНЬжЦфЧБдкзїгУЛњжЦЃЌДгЖјЮЊбАевP3ыФЕФзїгУАаЕуМАЦфгІгУПЊЗЂЬсЙЉЪЕбщвРОнЁЃ
1 ВФСЯ
1.1 ЯИАћжъ
RAW264.7ОоЪЩЯИАћЃЌЙКздУРЙњЕфаЭХрбјЮяБЃВижааФ(ATCCЃЌManassasЃЌVA USA)ЁЃ
1.2 ЪдМС
P3ыФ(ДПЖШЁн98%)ЃЛDMEMХрбјЛљЁЂЬЅХЃбЊЧхЁЂвШЕААзУИЃЌОљЙКздУРЙњHyCloneЙЋЫОЃЛMTTЁЂЖўМзЛљбЧэП(dimethyl sulfoxideЃЌDMSO)ЁЂБНМзЛљЛЧѕЃЗњ(phenylmethylsulfonyl fluorideЃЌPMSF)ЁЂгЭКьOШОСЯЁЂBCAЕААзХЈЖШВтЖЈЪдМСКаЃЌОљЙКздЫїРГБІЙЋЫОЃЛИЮXЪмЬх(liver X receptorЃЌLXR)МЄЖЏМСT0901317ЁЂGW3965ЃЌLXRвжжЦМСGSK2033ЃЌОљЙКздMedChemExpressЙЋЫОЃЛox-LDLЙКздЙужноШдДЩњЮяПЦММгаЯоЙЋЫОЃЛЧрУЙЫи-СДУЙЫиЫЋПЙЙКздSigmaЙЋЫОЃЛзмЕЈЙЬДМ(total cholesterolЃЌTC)ВтЖЈЪдМСКаЃЌЙКздФЯОЉНЈГЩЩњЮяЙЄГЬбаОПЫљЃЛОлКЯУИСДЪНЗДгІ(polymerase chain reactionЃЌPCR)в§ЮяЃЌЙКздЩњЙЄЩњЮяЙЄГЬ(ЩЯКЃ)ЙЩЗнгаЯоЙЋЫОЃЛcDNAФцзЊТМЪдМСКаЁЂqRT-PCRЪдМСКаЃЌЙКздTaKaRaЙЋЫОЃЛПЙABCA1ЭУЖрПЫТЁПЙЬхЃЌЙКздBioworldЙЋЫОЃЛПЙABCG1ЭУЖрПЫТЁПЙЬхЁЂПЙІТ-actinаЁЪѓЕЅПЫТЁПЙЬхЁЂРБИљЙ§бѕЛЏЮяУИБъМЧбђПЙЭУЖўПЙЁЂРБИљЙ§бѕЛЏЮяУИБъМЧбђПЙЪѓЖўПЙЃЌОљЙКздЮфККШ§гЅЩњЮяММЪѕгаЯоЙЋЫОЃЛECLЛЏбЇЗЂЙтвКЃЌЙКздЩЯКЃБЬдЦЬьЩњЮяММЪѕгаЯоЙЋЫОЁЃ
1.3 вЧЦї
JJ-CJ-1FDНрОЛЙЄзїЬЈ(НЫеЪаН№ОЛОЛЛЏЩшБИПЦММгаЯоЙЋЫО)ЃЛЖўбѕЛЏЬМХрбјЯф(ШеБОЫЩЯТЙЋЫО)ЃЛH4ЖрЙІФмУИБъвЧ(УРЙњBio-TekЙЋЫО)ЃЛTGL-16BЬЈЪНРыаФЛњ(ЩЯКЃАВЭЄПЦбЇвЧЦїГЇ)ЃЛTS100FгЋЙтЕЙжУЯдЮЂОЕ(ШеБОФсПЕЙЋЫО)ЃЛCFX96 PCRРЉдівЧЁЂGel Doc XRФ§НКГЩЯёвЧ(УРЙњBio-RadЙЋЫО)ЃЛND2000ГЌЮЂСПзЯЭтЗжЙтЙтЖШМЦ(ThermoЙЋЫО)ЃЛH1650RЬЈЪНИпЫйРфЖГРыаФЛњ(ЩЯКЃТЌЯцвЧРыаФЛњвЧЦїгаЯоЙЋЫО)ЃЛDYY-6CаЭЮШбЙЮШСїЕчгОвЧ(ББОЉСљвЛвЧЦїГЇ)ЁЃ
2 ЗНЗЈ
2.1 ЯИАћХрбј
RAW264.7ЯИАћГЃЙцХрбјгкDMEMИпЬЧХрбјЛљ(КЌ10%ЬЅХЃбЊЧхКЭ1%Чр-СДУЙЫиЫЋПЙ)жаЃЌжУгк37 ЁцЁЂ5% CO2ХрбјЯфжаХрбјЁЃУПЬьЛЛвК1ДЮЃЌД§ЯИАћШкКЯЖШдМЮЊ80%ЪБЃЌЪЙгУвШУИЯћЛЏЁЂДЋДњЁЃ
2.2 ЯИАћЛюСІМьВт
ВЩгУMTTЗЈВтЖЈЯИАћЛюСІЁЃЯШЙлВьP3ыФЁЂox-LDLЖдЯИАћдіжГЕФгАЯьЃЌИљОнMTTНсЙћЩИбЁГіP3ыФКЭox-LDLЕФзюМбзїгУХЈЖШЃЌЖјКѓдкЯргІЕФХЈЖШЗЖЮЇФкЃЌМьВтP3ыФЖдox-LDLгеЕМRAW264.7ЯИАћЛюСІЕФгАЯьЁЃНЋДІгкЖдЪ§ЩњГЄЦкЕФRAW264.7ЯИАћДЋДњЃЌЕїећЯИАћУмЖШЮЊ5ЁС107ЁЄL-1ЃЌНЋЯИАћНгжжгк96ПзАхЁЃШЗЖЈP3ыФКЭox-LDLзїгУХЈЖШКѓЃЌЗжзщШчЯТЃКПеАззщВЛНгжжЯИАћЃЌВЛМгox-LDLКЭP3ыФЃЛЖдеезщНгжжЯИАћЃЌВЛМгox-LDLКЭP3ыФДІРэЃЛФЃаЭзщНгжжЯИАћЃЌВЂМгШы80 mgЁЄL-1ЕФox-LDLЃЛP3ыФзщНгжжЯИАћЃЌМгШыХЈЖШЗжБ№ЮЊ10ЁЂ20ЁЂ50ЁЂ80ЁЂ100 mgЁЄL-1ЕФP3ыФЃЌвдМА80 mgЁЄL-1ЕФox-LDLЁЃгкХрбјЯфжаЗѕг§24 hКѓЃЌЦњХрбјЛљЃЌУППзМгШы5 gЁЄL-1 MTT 20 ІЬLЃЌМЬајЗѕг§4 hКѓЃЌЦњЩЯЧхЃЌУППзМгШыDMSO 150 ІЬLЃЌХрбјЯфжаЗѕг§10 minКѓЃЌЪЙгУУИБъвЧдкВЈГЄ490 nmВтЖЈИїПзЕФЮќЙтЖШ(absorbanceЃЌA)жЕЃЌМЦЫуЯИАћДцЛюТЪЃКЯИАћДцЛюТЪ=(AЪЕбщзщ-AПеАззщ)/(AЖдеезщ-AПеАззщ)ЁС100%ЁЃ
2.3 гЭКьOШОЩЋ
ШЁЖдЪ§ЩњГЄЦкЕФЯИАћЃЌвд5ЁС107L-1ХЈЖШНгжжгк6ПзАхЃЌХрбј24 hЯИАћЬљБкКѓЃЌВЮееЮФЯз[7]НЋЯИАћЗжЮЊ4зщЃКЖдеезщЁЂФЃаЭзщЁЂP3ыФЕЭХЈЖШзщЁЂP3ыФИпХЈЖШзщЁЃЖдеезщМгШыКЌ2%ЬЅХЃбЊЧхЕФDMEMХрбјЛљЃЌ24 hКѓЛЛГЩЮобЊЧхDMEMХрбјЛљМЬајХрбј24 hЃЛФЃаЭзщМгШыКЌ2%ЬЅХЃбЊЧхЕФDMEMХрбјЛљХрбј24 hКѓЃЌдкЮобЊЧхDMEMХрбјЛљжаМгШы80 mgЁЄL-1 ox-LDLДЬМЄ24 hЃЛP3ыФЕЭХЈЖШзщКЭP3ыФИпХЈЖШзщдкКЌ2%ЬЅХЃбЊЧхЕФDMEMХрбјЛљжаЗжБ№МгШы50ЁЂ100 mgЁЄL-1ЕФP3ыФИЩдЄ24 hКѓЃЌдкЮобЊЧхDMEMХрбјЛљжаМгШы80 mgЁЄL-1 ox-LDLДЬМЄ24 hЁЃХрбјНсЪјКѓЃЌЦњХрбјвКЃЌгУдЄРфЕФPBSЯД3ДЮЃЌУППзМгШы1 mL 4%ЖрОлМзШЉЙЬЖЈ20 minКѓЃЌЦњЖрОлМзШЉЃЌУППзМгШы1 mL PBSЯД3ДЮЁЃШЛКѓУППзМгШыдМ1.5 mLгЭКьOШОЩЋЙЄзївК(гЭКьOЛљвКЁУddH2OЃН3 ЁУ2)ШОЩЋ30 minЃЌPBSЯД3ДЮЃЌдкЙтбЇЯдЮЂОЕЯТЙлВьЯИАћжЌжЪБЛгЭКьOШОЩЋЧщПіЁЃ
2.4 ЯИАћФкTCКЌСПМьВт
АДееЁА2.3ЁБЖдЯИАћНјааЗжзщДІРэКѓЃЌЦњХрбјвКЃЌгУдЄРфPBSЯД2ДЮЃЌУППзМгШы1 mL PBSДЕДђЯИАћЃЌЪЙЯИАћГфЗжЭбТфЃЌВЂНЋЯИАћаќвКЪеМЏгкдЄЯШзіКУБъМЧЕФ1.5 mL EPЙмжаЃЌ1 000 rЁЄmin-1РыаФ5 minЃЌЦњЩЯЧхВЂСєГСЕэД§гУЁЃЯИАћГСЕэАДееЪдМСКаЫЕУїЪщВйзїЃЌВтЖЈИїзщTCКЌСПЁЃ
2.5 ABCA1ЁЂABCG1 mRNAБэДяВтЖЈ
АДееЁА2.3ЁБЖдЯИАћНјааЗжзщДІРэКѓЃЌЦњХрбјвКЃЌгУдЄРфPBSЯД3ДЮЃЌУППзМгШы1 mL TRIzolСбНтЯИАћЃЌВЂЪеМЏжС1.5 mLЮоRNAУИEPЙмжаЃЌАДееЪдМСКаЫЕУїЪщНјааИїзщбљЦЗRNAЬсШЁЁЂФцзЊТМКЭЖЈСПМьВтЁЃTab 1ЮЊЪЕЪБгЋЙтЖЈСПPCRв§ЮяађСаЃЌвдІТ-actinЮЊФкВЮЛљвђЃЌВЩгУ2-ЁїЁїCTЗЈМЦЫуИїзщЯИАћABCA1ЁЂABCG1 mRNAЕФЯрЖдБэДяСПЃКЁїЁїCt=(CtФПЕФЛљвђ-CtІТ-actin)ЪЕбщзщ-(CtФПЕФЛљвђ-CtІТ-actin)ЖдеезщЁЃ
2.6 ABCA1ЁЂABCG1ЕААзБэДяЕФВтЖЈ
ШЁЖдЪ§ЩњГЄЦкЕФЯИАћЃЌвд5ЁС108ЁЄL-1ЕФУмЖШНгжжгк6ПзАхжаЃЌХрбј24 hД§ЯИАћЬљБкКѓЃЌЗжЮЊ7зщЃКЖдеезщЁЂФЃаЭзщЁЂP3ыФЕЭХЈЖШзщЁЂP3ыФИпХЈЖШзщЁЂLXRМЄЖЏМСT0901317зщЁЂLXRМЄЖЏМСGW3965зщЁЂLXRвжжЦМСGSK2033зщЁЃЖдеезщЁЂФЃаЭзщЁЂP3ыФЕЭХЈЖШзщЁЂP3ыФИпХЈЖШзщЕФДІРэЗНЪНЭЌЁА2.3ЁБЁЃLXRМЄЖЏМСT0901317зщгыLXRМЄЖЏМСGW3965зщдкКЌ2%ЬЅХЃбЊЧхЕФDMEMХрбјЛљжаЗжБ№МгШы1 ІЬmolЁЄL-1 T0901317КЭGW3965зїгУ24 hЃЛLXRвжжЦМСGSK2033зщдкКЌ2%ЬЅХЃбЊЧхЕФDMEMХрбјЛљжаМгШы10 ІЬmolЁЄL-1 GSK2033КЭ100 mgЁЄL-1P3ыФзїгУ24 hЃЌШЛКѓ3зщдкЮобЊЧхDMEMХрбјЛљжаМгШы80 mgЁЄL-1ox-LDLзїгУ24 hЃЌЪеМЏЯИАћЁЃLXRМЄЖЏМСКЭвжжЦМСЕФХЈЖШВЮПМЮФЯз[8-9]ЁЃИїзщЯИАћМгШыЯИАћСбНтвК(RIPA ЁУPMSF=100 ЁУ1)200 ІЬLЃЌБљЩЯСбНт30 minЃЌ4 ЁцЁЂ9 710ЁСgРыаФ20 minЃЌШЁЩЯЧхвКБИгУЁЃBCAЕААзжЪСПХЈЖШВтЖЈЪдМСКаМьВтИїзщЕААзХЈЖШЃЌжЦБИ8%ЕФЗжРыНККЭ5%ХЈЫѕНКЃЌ20 ІЬgзмЕААз95 ЁцжѓЗа10 minКѓЃЌНјааЪЎЖўЭщЛљСђЫсФЦ-ОлБћЯЉѕЃАЗФ§НКЕчгОЗжРыЃЌЪЊЗЈзЊФЄЃЌ5%ХЃбЊЧхАзЕААзЪвЮТЗтБе2 hЃЌвЛПЙ4 ЁцЗѕг§Й§вЙЃЌЖўПЙЪвЮТЗѕг§2 hЃЌECLЪдМСКаЯдгАЁЃImageJШэМўЗжЮіИїЬѕДјЕФЛвЖШжЕЃЌвдІТ-actinЮЊФкВЮЕААзЃЌФПБъЕААзЕФЯрЖдБэДяСП=ФПБъЕААзЕФЛвЖШжЕ/ІТ-actinЕФЛвЖШжЕЁЃ
2.7 ЭГМЦбЇЗжЮі
ЫљгаЪЕбщОљжиИД3ДЮЃЌИїзщЪЕбщЪ§ОнвдxЁРsБэЪОЁЃВЩгУSPSS 19.0ШэМўНјааЭГМЦбЇЗжЮіЃЌЭЈЙ§ЕЅвђЫиЗНВюЗжЮіЖдЖрзщЪ§ОнжЎМфЕФВювьНјааЯджјадМьбщЃЌP < 0.05ЮЊВювьОпгаЯджјадЁЃРћгУGraphPad Prism 8.0.2ШэМўНјааЛцЭМЁЃ
3 НсЙћ
3.1 P3ыФЁЂox-LDLзюМбХЈЖШЕФбЁдёМАP3ыФЖдox-LDLгеЕМRAW264.7ЯИАћЛюСІЕФгАЯь
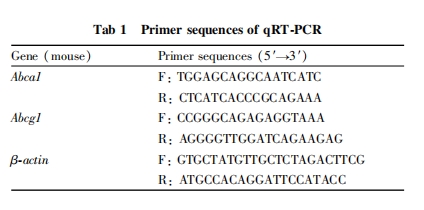
ВЩгУMTTЗЈМьВтP3ыФЖдRAW264.7ЯИАћЛюСІЕФгАЯьЃЌШчFig 1AЫљЪОЃЌгыЖдеезщЯрБШЃЌ10ЁЂ20ЁЂ50ЁЂ80ЁЂ100 mgЁЄL-1ЕФP3ыФзїгУRAW264.7ЯИАћ24 hКѓЃЌИїзщЯИАћДцЛюТЪЮоУїЯдБфЛЏЁЃЬсЪОP3ыФдк10~100 mgЁЄL-1ЗЖЮЇФкЃЌЖдЯИАћЮоУїЯдЖОадЃЌПЩгУгкКѓајЪЕбщЁЃШчFig 1BЫљЪОЃЌгыЖдеезщЯрБШЃЌ10ЁЂ20ЁЂ50 mgЁЄL-1ЕФox-LDLЖдЯИАћЛюСІЮоУїЯдгАЯьЃЛ80ЁЂ100 mgЁЄL-1ЕФox-LDLПЩУїЯдвжжЦRAW264.7ЯИАћЛюСІ(P < 0.01)ЁЃвђДЫЃЌбЁШЁ80 mgЁЄL-1ЕФox-LDLНјааКѓајЪЕбщЁЃ
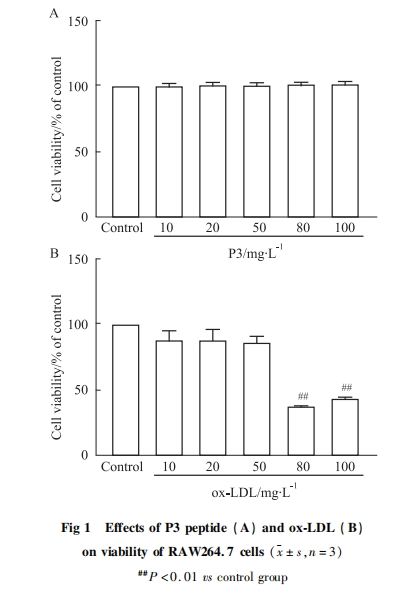
ЗжБ№НЋ10ЁЂ20ЁЂ50ЁЂ80ЁЂ100 mgЁЄL-1ЕФP3ыФгыox-LDL(80 mgЁЄL-1)ЙВЗѕг§КѓЃЌгыЖдеезщЯрБШЃЌP3ыФ(50ЁЂ80ЁЂ100 mgЁЄL-1)ПЩУїЯдФцзЊox-LDLЖдRAW264.7ЯИАћЕФЫ№ЩЫ(P < 0.01)ЃЌЯИАћДцЛюТЪПЩЛжИДжС80%вдЩЯ(Fig 2)ЃЌЫЕУїP3ыФЖдox-LDLгеЕМЕФЯИАћЫ№ЩЫОпгаБЃЛЄзїгУЁЃвђДЫЃЌбЁгУ50ЁЂ100 mgЁЄL-1ЕФP3ыФзїЮЊКѓајЪЕбщЕФХЈЖШЁЃ
3.2 P3ыФЖдox-LDLгеЕМЕФRAW264.7ЯИАћжЌжЪЛ§ОлЕФгАЯь
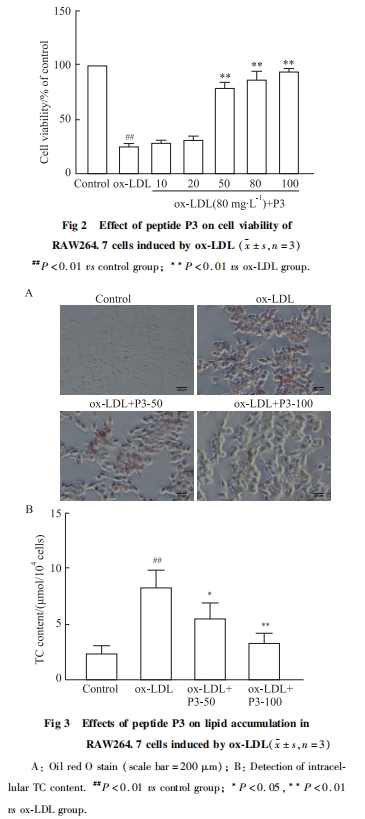
ЮЊСЫЙлВьP3ыФЪЧЗёЛсгАЯьЯИАћжЌжЪЕФГСЛ§ЃЌВЩгУox-LDL(80 mgЁЄL-1)геЕМRAW264.7ЯИАћаЮГЩХнФЯИАћЃЌВЂНЋP3ыФ(50ЁЂ100 mgЁЄL-1)гыox-LDLЙВЗѕг§ЃЌгЭКьOШОЩЋЗЈКЭTCВтЖЈЪдМСКаМьВтЯИАћФкжЌжЪГСЛ§ЧщПіЁЃНсЙћЯдЪО(Fig 3A)ЃЌгыЖдеезщЯрБШЃЌox-LDLгыЯИАћЙВЗѕг§КѓПЩУїЯддіМгЯИАћФкжЌжЪГСЛ§ЃЌаЮГЩХнФЯИАћЃЌP3ыФПЩИФЩЦRAW264.7ЯИАћФкЕФжЌжЪГСЛ§ЃЌвжжЦХнФЯИАћаЮГЩЁЃИїзщЯИАћФкTCМьВтНсЙћШчFig 3BЫљЪОЃЌгыЖдеезщЯрБШЃЌox-LDLгеЕМКѓПЩУїЯддіМгЯИАћФкTCЕФКЌСП(P < 0.01)ЃЌЖјP3ыФФмУїЯдНЕЕЭЯИАћФкTCКЌСП(P < 0.05ЃЌP < 0.01)ЁЃЬсЪОP3ыФПЩМѕЩйОоЪЩЯИАћЕФжЌжЪГСЛ§ЃЌвжжЦХнФЯИАћЕФаЮГЩЁЃ
3.3 P3ыФЖдox-LDLгеЕМЕФRAW264.7ЯИАћABCA1ЁЂABCG1БэДяЕФгАЯь
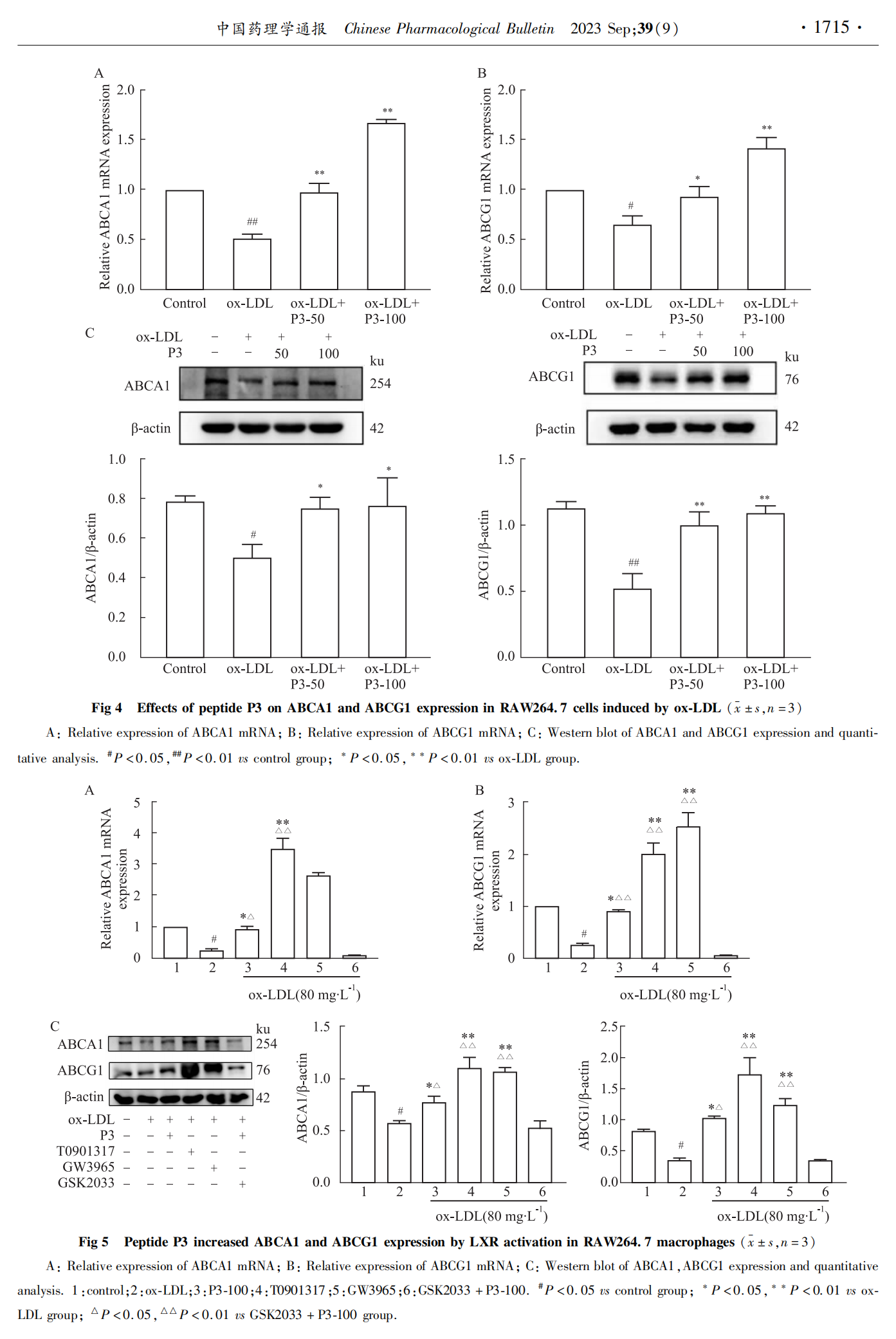
ABCA1КЭABCG1НщЕМЕФЕЈЙЬДМСїГіМѕЩйЪЧЕМжТОоЪЩЯИАћжЌжЪЖбЛ§ЕФжївЊдвђЁЃЮЊСЫНвЪОP3ыФвжжЦжЌжЪЖбЛ§ЕФЧБдкЛњжЦЃЌМьВтСЫИїзщЯИАћжаABCA1ЁЂABCG1 mRNAКЭЕААзБэДяЁЃШчFig 4A, BЫљЪОЃЌгыЖдеезщЯрБШЃЌox-LDLФЃаЭзщABCA1ЁЂABCG1mRNAБэДяУїЯдНЕЕЭ(P < 0.05ЃЌP < 0.01)ЃЛгыФЃаЭзщЯрБШЃЌP3ыФПЩУїЯдЩЯЕїABCA1ЁЂABCG1mRNAБэДя(P < 0.05ЃЌP < 0.01)ЁЃFig 4C, DНсЙћЯдЪОЃЌP3ыФФмУїЯдЩЯЕїABCA1КЭABCG1ЕААзБэДяЁЃЬсЪОP3ыФПЩФмЭЈЙ§ЩЯЕїABCA1ЁЂABCG1ЕФБэДяЃЌДйНјЕЈЙЬДМЕФСїГіЃЌДгЖјвжжЦRAW264.7ЯИАћФкЕФжЌжЪЛ§ОлЁЃ
3.4 P3ыФгыLXRМЄЖЏМСЁЂвжжЦМСЖдRAW264.7ЯИАћABCA1ЁЂABCG1БэДяЕФгАЯь
ABCA1КЭABCG1ЪЧLXRЕФАаЛљвђЃЌЦфБэДяПЩБЛLXRМЄЖЏМСЕїПиЁЃЮЊНјвЛВНЬНОПP3ыФЕїНкABCA1ЁЂABCG1ЛљвђБэДяЕФЛњжЦЃЌЗжБ№ВЩгУLXRМЄЖЏМСT0901317ЁЂGW3965МАLXRвжжЦМСGSK2033гыЯИАћЙВЗѕг§ЁЃШчFig 5ЫљЪОЃЌгыox-LDLФЃаЭзщЯрБШЃЌP3ыФгыLXRМЄЖЏМСGSK2033ЁЂT0901317ОљФмУїЯдЩЯЕїABCA1ЁЂABCG1 mRNAКЭЕААзЕФБэДя(P < 0.05ЃЌP < 0.01)ЃЛЖјМгШывжжЦLXRЛљвђзЊТМЕФвжжЦМСGSK2033КѓЃЌP3ыФВЛФмЩЯЕїABCA1ЁЂABCG1 mRNAКЭЕААзЕФБэДяЁЃетаЉНсЙћЬсЪОP3ыФПЩФмЪЧЭЈЙ§МЄЛюLXRЃЌДгЖјЩЯЕїABCA1ЁЂABCG1ЕФЛљвђБэДяЃЌЕЋЦфОпЬхЕФЛњжЦЛЙашвЊНјвЛВНбаОПЁЃ
4 ЬжТл
гЩгкИпжЌвћЪГКЭВЛНЁПЕЕФЩњЛюЗНЪНЃЌвдбЊНЌЕЈЙЬДМдіИпЮЊЬиЕуЕФИпЕЈЙЬДМбЊжЂЕФЗЂЩњТЪдкШЋЪРНчФкГжајЩЯЩ§ЁЃбЊНЌЕЈЙЬДМЃЌЬиБ№ЪЧLDLЕФдіИпЪЧASЗЂЩњЕФжївЊдвђЁЃЪГЮяЕЈЙЬДМЕФЩуШЁЮќЪеЁЂЕЈЙЬДМЕФЬхФкКЯГЩЃЌвдМАЕЈЙЬДМЕФЗжУкЖМЛсгАЯьбЊНЌЕЈЙЬДМЕФЫЎЦНЁЃФПЧАЃЌвбБЈЕРЕФНЕЕЈЙЬДМыФЕФзїгУЛњжЦжївЊЪЧМѕЩйЭтдДадЕЈЙЬДМЕФЮќЪеЁЂвжжЦЬхФкЕЈЙЬДМЕФЩњЮяКЯГЩМАДйНјЕЈЙЬДМзЊБфГЩЕЈжЫсЕШ[10]ЁЃдкASаЮГЩЕФдчЦкНзЖЮЃЌЛюЛЏЕФФкЦЄЯИАћФММЏЕЅКЫЯИАћЕНФкФЄЯТЃЌЗжЛЏЮЊОоЪЩЯИАћЃЌОоЪЩЯИАћЭЬЪЩox-LDLв§Ц№ЯИАћФкжЌжЪГСЛ§ЃЌЕМжТХнФЯИАћаЮГЩЁЃХнФЯИАћаЮГЩЕФЙ§ГЬгыОоЪЩЯИАћФкЕЈЙЬДМДњаЛУмЧаЯрЙиЃЌЦфжаЃЌЕЈЙЬДМДгОоЪЩЯИАћФкСїГіЪЧЕїНкОоЪЩЯИАћЕЈЙЬДМЖЏЬЌЦНКтЕФЙиМќЛЗНкжЎвЛЃЌЖдМѕЩйЯИАћФкЕЈЙЬДМГСЛ§ЁЂЗРжЙХнФЯИАћаЮГЩЁЂдЄЗРASОпгаживЊвтвхЁЃбаОПЗЂЯжЃЌМѕЩйОоЪЩЯИАћФкжЌжЪГСЛ§ПЩдкдчЦкИЩдЄASЕФЗЂЩњ[11]ЁЃБОбаОПЭЈЙ§ox-LDLгеЕМRAW264.7ЯИАћаЮГЩХнФЯИАћЃЌЗЂЯжP3ыФПЩвдМѕЩйОоЪЩЯИАћФкжЌжЪЕФГСЛ§ЃЌНЕЕЭЯИАћФкЕФTCКЌСПЃЌЫЕУїP3ыФОпгавжжЦХнФЯИАћаЮГЩЕФзїгУЁЃ
ХнФЯИАћЕФаЮГЩжївЊЪЧгЩгкЯИАћЩуШЁЕЈЙЬДМвьГЃЃЌЛђОоЪЩЯИАћЕЈЙЬДМСїГіЪмЫ№ЫљжТ[12]ЁЃABCA1КЭABCG1ЪЧвЛРрФЄЕААзЃЌЪЧATPНсКЯКазЊдЫЬх(ATP binding cassette transporterЃЌABC)ГЌМвзхжавЛдБЃЌПЩРћгУATPЫЎНтЪЭЗХЕФФмСПРДзЊдЫЕзЮяЃЌСНепЯрЛЅХфКЯЃЌЙВЭЌЭъГЩОоЪЩЯИАћЕЈЙЬДМЕФСїГіЁЃABCA1ЕФзїгУЪЧНЋЕЈЙЬДМКЭСзжЌѕЃЕЈМюДгОоЪЩЯИАћжазЊГіЃЌСїЯђЦЖжЌЕФдижЌЕААзApo A1ЃЌЩњГЩаТЩњИпУмЖШжЌЕААз(high-density lipoproteinЃЌHDL)ЃЌЖјABCG1дђНщЕМЕЈЙЬДМЁЂСзжЌѕЃЕЈМюКЭЧЪСзжЌСїЯђаТЩњКЭГЩЪьHDL[13]ЁЃABCA1КЭABCG1БэДяМѕЩйЛђЙІФмЩЅЪЇЃЌДйНјХнФЯИАћаЮГЩКЭASЗЂЩњЃЌЖјвЛаЉЛЦЭЊРрЮяжЪЃЌШчЖўЧтбюУЗЫиПЩЭЈЙ§геЕМABCA1ЁЂABCG1ЕФБэДявжжЦХнФЯИАћЕФаЮГЩ[14]ЁЃБОбаОПжаЃЌP3ыФПЩУїЯдЩЯЕїABCA1КЭABCG1ЕФЛљвђБэДяЃЌЬсЪОP3ыФНЕЕЭox-LDLгеЕМЕФRAW264.7ЯИАћжаЕЈЙЬДМКЌСПЃЌПЩФмЪЧЭЈЙ§геЕМABCA1КЭABCG1ЕФБэДяЃЌНјЖјДйНјЕЈЙЬДМДгОоЪЩЯИАћСїГіЁЃ
LXRгаLXRІСКЭLXRІТСНжжбЧаЭЃЌЪЧРрЙЬДММЄЛюЕФзЊТМвђзгЃЌЪєгкРрЙЬДМ/МззДЯйМЄЫиКЫЪмЬхГЌМвзхГЩдБЁЃLXRЕїПивЛЯЕСаЩцМАЕЈЙЬДМЮќЪеЁЂзЊдЫКЭзЊЛЏЕФЛљвђзЊТМЃЌБЛГЦЮЊЕЈЙЬДМИаЪмЦїЁЃLXRЕФзЊТМЕїПиЙІФмашвЊгыРрЪгЛЦДМXЪмЬх(retinoid X receptorЃЌRXR)НсКЯЃЌаЮГЩLXR/RXRвьЖўОлЬхВХФмЗЂЛгЁЃЮоХфЬхЪБЃЌИУвьЖўОлЬхгыКЫЪмЬхзшЖєвђзгНсКЯЃЌвжжЦЛљвђБэДяЃЛЕБХфЬхгыжЎНсКЯКѓЃЌЛсЕМжТвьЖўОлЬхЙЙЯѓЗЂЩњИФБфЃЌЪЭЗХГізшЖєвђзгЃЌВЂФММЏИЈМЄЛювђзгЃЌLXR/RXRгыАаЛљвђЦєЖЏзгЧјгђФкЕФЬиЖЈDNAађСаЃЌМДLXRЗДгІдЊМўНсКЯЃЌЕїНкЯТгЮАаЛљвђЕФзЊТМ[15]ЁЃLXRІСдкОоЪЩЯИАћжаИпБэДяЃЌABCA1КЭABCG1ЪЧLXRІСдкОоЪЩЯИАћжаЕФАаЛљвђЃЌЕїНкЕЈЙЬДМДгОоЪЩЯИАћСїГі[16]ЁЃбаОПШЯЮЊЃЌМЄЛюLXRПЩдіМгЭтжмЕЈЙЬДМСїГіЁЂДйНјЕЈЙЬДМЯђИЮзЊдЫМАзЊБфГЩЕЈжЫсЁЂДйНјЕЈжЫсЗжУкЕШЃЌДгЖјНЕЕЭбЊНЌЕЈЙЬДМЫЎЦНЃЌЗЂЛгПЙASзїгУ[17]ЁЃLXRМЄЛюМСT0901317ЭЈЙ§ЧтМќгыLXRНсКЯЃЌФММЏLXRКЭИЈМЄЛювђзгНсКЯЃЌНјЖјМЄЛюLXRЃЌЖдLXRІСКЭLXRІТОљгаМЄЛюзїгУЃЛGW3965бЁдёадгыLXRІСНсКЯЃЌНјЖјМЄЛюLXRІС[18]ЁЃБОбаОПНсЙћЗЂЯжЃЌP3ыФЕЅЖРгыОоЪЩЯИАћзїгУЪБЃЌгыT0901317ЁЂGW3965вЛбљЃЌПЩУїЯдЩЯЕїABCA1КЭABCG1ЕФБэДяЃЌЕЋЯШгУGSK2033вжжЦLXRЛљвђзЊТМКѓЃЌдйгыP3ыФзїгУЃЌЗЂЯжP3ыФЖдABCA1ЁЂABCG1ЕФЩЯЕїзїгУЯћЪЇЃЌЬсЪОP3ыФПЩФмЪЧLXRЕФЧБдкМЄЛюМСЃЌЕЋЦфгыLXRЕФОпЬхзїгУЛњжЦЛЙашвЊНјвЛВНбаОПЁЃ
злЩЯЃЌP3ыФПЩМѕЩйox-LDLгеЕМЕФRAW264.7ОоЪЩЯИАћЕФжЌжЪЛ§ОлЃЌМѕЩйЯИАћФкЕФTCКЌСПЃЌвжжЦХнФЯИАћЕФаЮГЩЃЌЦфЛњжЦПЩФмЪЧЭЈЙ§МЄЛюLXRЃЌЩЯЕїABCA1КЭABCG1ЕФБэДяНщЕМЕФЁЃБОбаОПНсЙћПЩЮЊP3ыФЕФгІгУПЊЗЂЬсЙЉЪЕбщвРОнЁЃ
Утд№ЩљУїЃКБОЮФЮЊаавЕНЛСїбЇЯАЃЌАцШЈЙщдзїепМАддгжОЫљгаЃЌШчгаЧжШЈЃЌПЩСЊЯЕЩОГ§ЁЃЮФеТБъзЂгазїепМАЮФеТГіДІЃЌШчашдФЖСдЮФМАВЮПМЮФЯзЃЌПЩдФЖСддгжО
