
ЃлеЊвЊЃн ПЙОњыФЪЧвЛжжаЁЗжзгЛюадыФЃЌРДдДЙуЗКЃЌдкЩњЮяЬхУтвпМЄЛюКЭЕїНкжаЗЂЛгживЊзїгУЃЌЧвОпгаПЙОњЛюадЃЌетЮЊНтОіПЙЩњЫиФЭвЉадЮЪЬтЬсЙЉСЫПЩФмЁЃLLЉ\37ЪЧзщжЏЕААзУИвжжЦЫиМвзхжаЮЈвЛдкШЫЬхФкЕФГЩдБЃЌЖдИяРМвѕадОњКЭИяРМбєадОњОљгавжжЦзїгУЁЃИУЮФДгLLЉ\37ЛюадНсЙЙгђНјааЗжЮіЃЌВћЪіСЫLLЉ\37дкПЙОњЁЂПЙФкЖОЫиЁЂДДЩЫгњКЯЁЂУтвпЕїНкЕШЗНУцЕФзїгУЛњжЦМАСйДВжЮСЦЧБСІЃЌеЙЭћСЫЦфгІгУЧАОАЃЌЮЊаТвЉбаЗЂЁЂЪГЦЗЗРИЏКЭЖЏЮяЫЧСЯЬэМгЬсЙЉЫМТЗЁЃ
ПЙЩњЫидјдкЪРНчЩЯеМгаживЊЕФЕиЮЛЃЌеќОШЙ§ЮоЪ§ШЫЕФЩњУќЃЌдкХЉвЉЁЂЪГЦЗЬэМгМСЁЂаѓФСРрбјжГжаЗЂЛгСЫживЊзїгУЃЌГЩЮЊШШУХЕФПЙОњвЉЁЃШЛЖјЃЌдк 20 ЪРМЭ70ФъДњКѓЃЌбаЗЂГіИпаЇПЙЩњЫиШДГЩЮЊФбЬтЁЃгЩгкПЙЩњЫиЕФГЄЦкЪЙгУЃЌдНРДдНЖрЕФЯИОњБэЯжГіФЭвЉадЃЌЩѕжСВњЩњСЫЭчЧПЕФЁАГЌМЖЯИОњЁБЃл1~4ЃнЁЃЯИОњПЙвЉадЕФдіЧПЪЙжюЖрПЙЩњЫиЪЇШЅаЇСІЃЌвђДЫЃЌПЊЗЂФмЙЛЬцДњПЙЩњЫиЕФаТаЭгааЇВњЦЗПЬВЛШнЛКЃл5ЃЌ6ЃнЁЃ
ПЙОњыФРДдДЙуЗКЃЌзюдчЗЂЯжгкРЅГцЕФУтвпЯЕЭГжаЁЃЫцКѓЃЌДгЯИОњЁЂецОњЁЂСНЦмЖЏЮяЁЂИпЕШжВЮяКЭВИШщЖЏЮяжавВЬсШЁСЫРрЫЦЕФЖрыФЁЃетжжЛюадыФОпгаЬьШЛУтвпЙІФмЃЌЪЧЫожїЕжПЙВЁдЮЂЩњЮяШыЧжЕФживЊЗжзгЦСеЯЃЌвђДЫБЛГЦЮЊПЙОњыФЃл7ЃЌ8ЃнЁЃПЙОњыФОпгаПЙОњЦзЙуЁЂЯрЖдЗжзгжЪСПаЁЁЂШШЮШЖЈадИпЁЂЫЎШмадКУЕШЬиЕуЁЃФПЧАбаОПвбЩцМАЕНЗжзгНсЙЙЁЂЗжзгИФСМЁЂзїгУЛњжЦЕШЖрЗНУцЃл9ЃнЁЃДгПЙОњЛњжЦЗНУцРДПДЃЌПЙЩњЫизїгУгкЬиЪтЪмЬхЃЌЯИОњПЩЭЈЙ§БфвьРДЖдПЙЫќЕФзїгУЃЌзюжеЕМжТФЭвЉОњЕФВњЩњЁЃЖјПЙОњыФПЩОЙ§ЮяРэзїгУЪЙЯИАћФЄДЉПзЃЌзюжеДяЕНЙуЦзПЙОњЕФаЇЙћЃЌвђДЫВЛвзВњЩњФЭвЉадЁЃЫљвдЃЌПЙОњыФМЋЦфПЩФмГЩЮЊПЙЩњЫиЕФЬцДњЦЗЃл10ЃнЁЃ
ЦфжаЃЌШЫдДадПЙОњыФЯИАћЖОадЕЭЧвХХвьЗДгІаЁЃЌЯдЪОГівЛЖЈЕФСйДВжЮСЦгХЪЦЃл11ЃнЁЃФПЧАШЫдДПЙОњыФгазщжЏЕААзУИвжжЦЫиЃЈcathelicidinЃЉМвзхЁЂЗРгљЫиЃЈdefensinЃЉМвзхЁЂИЛКЌзщАБЫсЕААзМвзхЕШЃл12ЃнЁЃЗРгљЫиМвзхЪЧвЛжжФкдДадПЙОњыФЃЌПЙОњЛюадНЯЧПЃЌНі 1~10 µgМДЯдЪОГівжОњЛюадЁЃзщжЏЕААзУИвжжЦЫиМвзхЪЧВИШщРрЖЏЮяЬигаЕФПЙОњыФЃЌLLЉ\37 ЪЧЦфЮЈвЛдкШЫЬхФкЕФПЙОњыФЃЌОпгаЖржжзїгУЁЃзщжЏЫиЃЈhistatinЃЉЪЧвЛРрИЛКЌзщАБЫсЕФПЙОњыФЃЌгаНЯЧППЙЯИОњКЭПЙецОњЛюадЃл13ЃнЁЃЫцзХвНбЇММЪѕОЋНјЃЌЖржжШЫдДПЙОњыФвбНјШыСйДВЪдбщЃл14ЃнЃЌШчШЫШщЬњЕААз1Љ\11ЃЈhuman lactoferrin1Љ\11ЃЌhLF1Љ\11ЃЉвбНјШыСйДВЂђЦкНзЖЮЃЌгУгкЙЧЫшвЦжВЙ§ГЬПЙИаШОжЮСЦЃЛЬьВЯЫи BбмЩњЮяЃЈHBЉ\107ЃЉНјШыСйДВЧАЦкЃЌгУгкДДЩЫаоИДЃЛРДдДгкжэПЙОњыФЕФРрЫЦЮяЃЈprotegrin analogueЃЌIBЉ\367ЃЉНјШыЂѓЦкСйДВЪдбщЃЌгУжЮСЦ№ЄФЄбзЃл15~17ЃнЁЃЫљвдЃЌШЫдДПЙОњыФМЋгаПЩФмГЩЮЊбаОПШШЕуЁЃ
LLЉ\37 жївЊКЯГЩВПЮЛЮЊжаадСЃЯИАћЃЌОпгаСНЧзадІСТна§НсЙЙЃЌПЩгЩОоЪЩЯИАћЁЂЕЅКЫЯИАћЁЂНЧЕААзЯИАћЁЂжаадСЃЯИАћЕШНјааЗжУкЃл18ЃнЁЃLLЉ\37ОпгавжжЦИяРМбєадЃЈG+ЃЉМАИяРМвѕадЃЈG-ЃЉОњЛюадЬиЕуЃЌВЂгаПЙФкЖОЫиЁЂЧїЛЏзїгУЁЂДйНјбЊЙмЩњГЩзїгУЕШЙІФмЃЌетаЉЖМЯдЪОГіЦфДњЬцПЙЩњЫиЕФПЩФмадЁЃЕЋПЙОњыФгЩгкКЌСПНЯЩйЃЌЧвЗжРыДПЛЏВНжшИДдгЃЌВЛвзЬсШЁЃЌЗбгУАКЙѓЃЌДЋЭГЗжРыЗНЗЈвбВЛЪЪКЯДѓСПЩњВњЃл19ЃнЁЃЯжНёЖрВЩгУЛљвђЙЄГЬЛЏЩњВњЃЌЩИбЁГіБэДяСПИпЁЂИпПЙОњЛюадЁЂЕЭЖОадПЙОњыФЃЌВЂНјааЯТвЛВНЗжЮіЃл20ЃнЁЃ
ШЛЖјЃЌЯрБШгкПЙЩњЫиЃЌLLЉ\37ПЙОњЛюСІШдШЛДІгкСгЪЦЃЌШчКЮЬсИпПЙОњЛюадВЂНЕЕЭШмбЊЛюадКЭЯИАћЖОадЪЧЙиМќЕуЃл21ЃнЁЃФПЧАЃЌНЋФПЕФЛљвђЕМШыдиЬхФкНјааИпаЇБэДяЪЧНЯЮЊРэЯыЕФЪжЖЮЁЃдкНјаааоЪЮИФдьЧАЃЌШЫУЧашвЊдкЛљБОВЛИФБфПЙОњыФгХЪЦНсЙЙЧАЬсЯТЃЌЭЈЙ§АБЛљЫсжУЛЛЁЂВПЗжађСаНиШЁКЭзщКЯЛЏбЇПтЕШЗНЗЈЛёЕУФПЕФПЙОњыФЃЌетНЋЪЧЮДРДЙЙНЈгХаЇПЙОњыФЕФживЊЗНЗЈЃл22ЃЌ23ЃнЁЃИФБфПЙОњыФЖдЯИОњЯИАћФЄЕФНсКЯзїгУЃЌЖдПЙОњыФвжОњЛюадЕїПигаЙиМќзїгУЁЃбюбоРіЕШЃл24ЃнЗжБ№гУ Gln16ЁЂAsn26 КЭ Gln36 ШЁДњ Glu16ЁЂAsp26 КЭGlu36ЃЌЪЙОВЕчДг+5.8 діМгЕН+9.0ЁЃетжжаоЪЮЬсИпСЫLLЉ\37 е§ЕчКЩЫЎЦНЃЌВЂЮДИФБф LLЉ\37 ПеМфНсЙЙЁЃGaЉ\gnon ЕШЃл25ЃнЗЂЯжыФСДГЄЖШвВгыПЙОњЛюадгаЙиЁЃдкыФСДГЄЖШЯрЭЌЧщПіЯТЃЌГЄЖШдНГЄЃЌе§ЕчКЩдНЖрЕФПЙОњыФПЙОњаЇЙћдНКУЁЃ
вђДЫЃЌБОЮФЖд LLЉ\37 ПЙОњыФЕФЩњЮяНсЙЙНјааНтЮіЃЌЗжЮіВЛЭЌЧјгђЕФЙиМќзїгУЃЌВЂЖдЩњЮябЇЬиадЃЈПЙОњЁЂПЙФкЖОЫиЁЂУтвпЕїНкЁЂДйНјбЊЙмЩњГЩЃЉЕШЗНУцНјааВћЪіЃЌНЋЮЊШЫРрПЙОњыФЕФПЊЗЂЬсЙЉРэТлвРОнЃЌВЂдкLLЉ\37ЧБдкгІгУЗНУцЬсЙЉЫМТЗЁЃ
1 LL⁃37ЩњЮяНсЙЙМАЩњЮяЙІФм
1.1 ЩњЮяНсЙЙ
ШЫбєРызгПЙОњЕААз18ЃЈhuman cationic antimicrobialprotein 18ЃЌhCAP18ЃЉЪЧзщжЏЕААзвжжЦЫиМвзхГЩдБЃЌвджївЊЕААзаЮЪНДцдкгкжаадСЃЯИАћжаЃЌгЩиКЭшЁЂЙЧЫшЁЂЗЮЩЯЦЄЯИАћЁЂбзадЩЯЦЄНЧжЪаЮГЩЯИАћЁЂЪГЙмЁЂЙЌОБЕШВњЩњЃЌжївЊЭЈЙ§ОпгаПЙОњЛюадЕФ C ЖЫгыжЌЕААзСЌНгЃл26ЃнЁЃhCAP18вдЗЧЛюадЧАЬхЕААзаЮЪНДЂДцдкПХСЃжаЃЌдквЛЖЈЬѕМўЯТЗжСбГіЦфжаЕФCЖЫЛюадыФЃЌМДLL Љ\37ыФЃл27ЃнЁЃдкЬхФк hCAP18 ГіАћКѓЃЌЕААзУИ 3 ЖдЦфНјааСбНтЁЃгаЪЕбщБэУїЃЌжЛвЊЕААзУИ 3 ХЈЖШзуЙЛИпЃЌhCAP18ОЭФмШЋВПЩњГЩLLЉ\37Ѓл28ЃнЁЃ
LLЉ\37ЕФ NЖЫЧАКЌгаЕФСНИіРЕАБЫсВаЛљЃЈLЃЉЃЌЧвгЩ 37 ИіАБЛљЫсзщГЩЃЌвђДЫЕУУћ LLЉ\37Ѓл29ЃнЁЃLLЉ\37 ПЩдкШЫЬхИїзщжЏЦїЙйжаБэДяЃЌЬиБ№ЪЧдкЯћЛЏЕРЁЂЦЄЗєЁЂУкФђЕРКЭПкЧЛЩЯЦЄЯИАћжаЁЃДЫЭтЃЌЫќвВБэДядквЛаЉЩЫПкЩјГівКЁЂКЙвККЭбЊвКжаЃл30ЃнЁЃLLЉ\37 ЛљвђЦєЖЏзгЧјгђФкАќКЌ 1Иі TAAA ађСаЁЂ1ИіАзНщЫи 6КЫзЊТМвђзгЃЈnuclear factor interleukinЉ\6ЃЌNFЉ\ILЉ\6ЃЉНсКЯЮЛЕуЁЂ3Иі МБ ад Цк ЗД гІ вђ згЃЈacute phase response factorЃЌAPRFЃЉЃЌПЩгыЬхФкЩњГЩЕФбзжЂвђзгНсКЯЃЌДгЖјдіМгLLЉ\37ЛљвђБэДяЫЎЦНЃл31ЃнЁЃ
дкДПЫЎжаЃЌLLЉ\37аЮГЩВЛЙцдђЕФТна§НсЙЙЃЌЕБжмЮЇЛЗОГИФБф ІС Тна§НсЙЙЪБЃЌЛсаЮГЩСНЧзадбєРызгНсЙЙЁЃLLЉ\37ДцдкЕФCЖЫКЭNЖЫІСЉ\Тна§ЃЌСНЖЫЕФТна§ЯпЭЈЙ§зЊелЛђЭфЧњЗжПЊЁЃЕк 5ЁЂ6ЁЂ17КЭ 27ЮЛБНБћАБЫсВрСДЯђЭЌвЛЗНЯђбгЩьЃЌаЮГЩЪшЫЎБэУцЃЌе§ЕчКЩВаЛљКЭЪшЫЎБэУцПЩгыИКЕчКЩЗжзгНсЙЙжБНгЯрСЌЃЌШчG+жЌЬЧЁЂG-СзБкЫсЕШЃл32ЃнЁЃдкШЫЬхКЙвКжаЃЌhCAP18аоГі LLЉ\37ЦЌЖЮВњЮяЃЈKSЉ\30КЭ RKЉ\31ЃЉЃЌЖўепЕФ NЖЫАБЛљЫсКЌСПБШ LLЉ\37 Щй 6/7ЃЌОпгаНЯЧПЕФПЙОњЛюадЃЌЕЋВЛОпгаУтвпЕїНкФмСІЃл33ЃнЁЃСэЭтЃЌLLЉ\37дкШБЗІNЖЫ4жжАБЛљЫсЪБЃЌШдФмБЃГжПЙОњЛюадЃЌЕЋДѓВПЗжШмбЊЛюадЩЅЪЇЁЃетаЉНсЙћБэУїЃЌNЖЫТна§ЖдЖрыФЕФЙбОлЁЂЧїЛЏЁЂШмбЊКЭПЙЕААзУИЫЎНтгаживЊзїгУЁЃаэЖрЪЕбщаЁзщбаОПСЫНиЖЬыФЕФЛюадЃЌвдСЫНт LLЉ\37 зюаЁПЙОњЧјгђЃл34ЃнЁЃНсЙћБэУїЃЌLLЉ\37ЕФзюаЁЧјгђдкАБЛљЫс17~29ЮЛЛђ 18~29 ЮЛжЎМфЃЌБэУї LLЉ\37 ЕФ C ЖЫТна§ОпгаПЙОњЁЂПЙВЁЖОКЭПЙжзСіЕШзїгУЁЃ
баОПЯдЪОЃЌКуКгКяЕФЭЌдДЕААз RLЉ\37 ОЛе§ЕчКЩЮЊ+8ЃЌДѓгк LLЉ\37 ЕФ+6ЃЌПЙОњЛюадвЊИпгк LLЉ\37Ѓл35ЃнЁЃвђДЫдіМгLLЉ\37жаЕФМюадАБЛљЫсЪ§ФПЖјВЛИФБфЦфІСЉ\Тна§ЙЙЯѓПЩЬсИпЦфПЙОњЛюадЃл36ЃнЁЃыФСДжаДје§ЕчКЩКЭЪшЫЎадАБЛљЫсЕФЪ§ФПвВгыЦфЫћЛюадгаЙиЃЌВЂОпгаЙЙЯѓвРРЕадЬиЕуЃЌвђДЫПЩЭЈЙ§вдЯТЗНЪНИФдьЃКЂйдкНсЙЙЩЯгІЮЊЙмзДЛђТна§зДЃЌдкаЮзДЩЯБЃГжЗжзгЕФЧзЫЎадКЭЪшЫЎадЃЛЂкДюХфе§ЕчадгыИКЕчадАБЛљЫсЁЂЧзЫЎадгыЪшЫЎадАБЛљЫсЃЌИЈвдвзаЮГЩТна§зДНсЙЙАБЛљЫсЃЌЬсИпПЙОњыФЛюадЃЛЂлдіМге§ЕчадКЭЪшЫЎАБЛљЫсЕФЪ§СПРДЬсИпЩБОњСІКЭПЙФкЖОЫиФмСІЁЃ
1.2 ЩњЮяЙІФм
1.2.1 ПЙОњзїгУ LLЉ\37Жд G+КЭ G-ЖМгаПЙОњзїгУЃЌдкОВЕчзїгУЯТЃЌДје§ЕчКЩ LLЉ\37 гыДјИКЕчКЩЯИОњНгЃЌВхШыЯИОњЯИАћФЄЃЌЕМжТЦфФкШнЮяСїГіЫРЭіЃл37ЃнЁЃФПЧАЃЌгаСНжжМйЫЕжЇГжLLЉ\37ШмНтЯИАћФЄЕФЛњжЦЃКЕиЬКФЃаЭЃЈЭМ 1ЃЉЪЧЛљгк LLЉ\37 БэУцКЌгабєРызгАБЛљЫсгыЯИАћФЄСзжЌЭЗВПНсКЯЕФРэТлЃЌПЩЦЬдкЯИАћФЄБэУцаЮГЩЕиЬКНсЙЙЃЌВЂЦЦЛЕЯИАћФЄЭъећадЁЃЕБ LLЉ\37 Тна§НсЙЙЁЂЕчКЩКЭЪшЫЎадЗЂЩњБфЛЏЪБЃЌЦфПЙОњЛюадЪмЕНгАЯьЃЛЛЗаЮПзФЃаЭЃЈЭМ 2ЃЉжївЊИљОнФЄБэУцеХСІКЭЧњТЪЃЌЪЙЯИОњФЄаЮГЩЛЗаЮПзЃЌДгЖјаЙТЖЯИОњФкШнЮяЃЌжТЪЙЯИОњЫРЭіЃл38ЃнЁЃЖдгкВИШщЖЏЮяРДЫЕЃЌЦфЯИАћФЄЛљБОДІгкжаадзДЬЌЃЌЫљвдLLЉ\37ЖдФЄзїгУНЯШѕЃЌЯИАћВЛвзЪмЕНЫ№ЩЫЁЃДЫЭтЃЌLLЉ\37 ФмдіЧПЩЯЦЄЯИАћФЄгВЖШВЂМѕШѕЦфЭЈЭИадЃЌДгЖјМѕЩйЯИОњЙЅЛїЃл39ЃнЁЃЦфДЮЃЌLLЉ\37ОпгаЫЋЧзадТна§НсЙЙЃЌЪшЫЎзїгУЯТвзДйНјЙбОлЬхаЮГЩЃЌЛњЬхПЙЬхЛюадгыЙбОлЬхаЮЬЌЕФ ІС Тна§НсЙЙГЩе§БШЃЌЕБІСТна§НЯЖрЪБЃЌПЙОњЛюадНЯЧПЁЃ
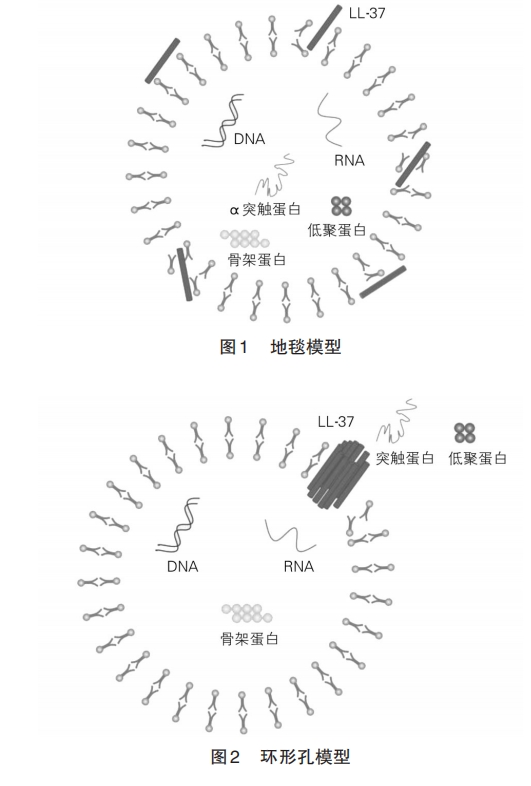
1.2.2 ПЙФкЖОЫизїгУ жЌЖрЬЧЃЈlipopolysaccharideЃЌLPSЃЉжївЊЮЛгк GЉ\ЯИАћЭтФЄЩЯЁЃВЮгыЙ§ГЬжївЊгаЃКЂйдк LPS геЕМЕФ Toll бљЪмЬхЃЈTollЉ\like receptorЃЌTLRЃЉЉ\КЫвђзг ІЪBЃЈnuclear factor kappaЉ\BЃЌNFЉ\ІЪBЃЉаХКХЭЈТЗжаЃЌLLЉ\37 Ц№ЕїНкзїгУЁЃДЫЭтЃЌLLЉ\37 вжжЦ LPS геЕМЕФ NFЉ\ІЪBбЧЕЅЮЛ p50МА p65ЕФзЊТМЃл41ЃнЁЃЂкЫПСбдЛюЛЏЕААзМЄУИЃЈmitogen Љ\activated protein kinaseЃЌMAPKЃЉЭЈЕРБЛМЄЛюЃЌДгЖјдіМгДйбзЗДгІЭЈЕРЛюадЃл42ЃнЁЃЂлбЁдёадЕиЕїНкЛљвђзЊТМЃЌНЕЕЭДйбзЗДгІЛљвђ p50МАжзСіЛЕЫРвђзгЉ\ІСЃЈtumor necrosis factorЉ\ІСЃЌTNFЉ\ІСЃЉЕФБэДяЁЃЂмвжжЦ LPS гы LPS НсКЯЕААзЃЈLPSЉ\binding proЉ\teinЃЌLBPЃЉЁЂЫшбљЗжЛЏЕААз 2ЃЈmyeloid differentiationprotein 2ЃЌMDЉ\2ЃЉЛђ TLR ИДКЯЮяЯрНсКЯЃЌНЕЕЭЯТгЮЭЈТЗЕФМЄЛюЁЃОЙ§етаЉЛњжЦЕїНкЃЌгЩLPSв§Ц№ЕФВЛСМЗДгІПЩБЛLLЉ\37гааЇвжжЦЁЃ
1.2.3 УтвпЕїНкзїгУ дкЮШЖЈзДЬЌЬѕМўЯТЃЌжаадСЃЯИАћЗЂЩњНсЙЙадЕђЭіЃЌвдНсЪјЦфЖЬднЪйУќЁЃжаадСЃЯИАћЛЕЫРЛђЕђЭіКѓЃЌдкбзжЂВПЮЛаЮГЩжаадСЃЯИАћАћЭтЩБОњЭјТчЃЈneutrophil extracellular trapsЃЌNETЃЉЃЌПЩЬсЙЉИпХЈЖШПЙОњЗжзгЃЌПьЫйПижЦЬхФкЯИОњИаШОЃл43ЃЌ44ЃнЁЃВЮгыЖржжТ§адбзжЂЕФ NET ПЩдкЬхФкЭтЪЭЗХ LLЉ\37ЃЌЭЌЪБLLЉ\37вВДйНјЭтжмжаадСЃЯИАћвдМССПвРРЕЗНЪНаЮГЩ NETЁЃЖј NETЭЈЙ§ C1qЃЈбЊЧхВЙЬхЃЉ/LLЉ\37ЬивьадвжжЦLPSМЄЛюЕФОоЪЩЯИАћЗжУкILЉ\6Ѓл45ЃЌ46ЃнЁЃдчЦкбаОПБэУїЃЌLLЉ\37 РДдДЕФГЩЪьЪїЭЛзДЯИАћЃЈmaturationof dendritic cellЃЌmDCЃЉгыLPSВњЩњСЫвЛИіЕфаЭЕФИЈжњадTЯИАћЃЈhelper T cellЃЌTh1ЃЉгеЕМЯИАћвђзгЦзЃЈILЉ\12ЁЂILЉ\6 КЭ TNFЉ\ІС ЯджјдіМгЃЌILЉ\4 ЯджјНЕЕЭЃЉЁЃДЫЭтЃЌдкmDCДЬМЄЯТЃЌTЯИАћЖдУтвпЛюадЮяжЪІУИЩШХЫиЃЈinterferЉ\on gammaЃЌIFNЉ\ІУЃЉЕФКЯГЩЯджјдіМгЁЃвђДЫЃЌLLЉ\37ЫЦКѕЪЧЯШЬьУтвпЯЕЭГКЭЪЪгІадУтвпЯЕЭГжЎМфЕФЧХСКЁЃЭЈЙ§ЦРЙР LL Љ\37 дкЭтжмбЊЕЅИіКЫЯИАћЃЈperipheralblood mononuclear cellsЃЌPBMCЃЉжаЕФзїгУЃЌбаОПШЫдБЗЂЯжLLЉ\37вВДйНјЕїНкадTЯИАћВњЩњЃЌЖјВЛгАЯьTЯИАћЛюЛЏЃЛдкбзжЂЃлжВЮябЊФ§ЫиЃЈphytohemagglutininЃЌPhaЃЉМЄЛюЃнБГОАЯТЃЌLLЉ\37ыФПЩгеЕМОВЯЂTЯИАћдіжГЃЌЯджјНЕЕЭДй PBMC ДйбзвђзгБэДяЃЈIFNЉ\ІУЃЌTNFЉ\ІСЃЉЃЛБэУїЕБыФЗЂЛгздЩэПЙИаШОЬиадЪБЃЌИУЗДгІЪМжеПижЦДйбзЗДгІЃЌвдБЃЛЄЛњЬхУтЪмбЯжибзжЂЗДгІЃл47ЃнЁЃЖјЧвЃЌLLЉ\37 ЕїНк TLR 3 ЕФБэДяЃЌДйНјЗжУкИќИпЫЎЦНПЙбзвђзгЃлпХпсАЗ2ЃЌ3Љ\ЫЋМгбѕУИЃЈindoleamine2ЃЌ3Љ\dioxyЉ\genaseЃЌIDOЃЉЃнЁЂILЉ\10КЭзЊЛЏЩњГЄвђзгІТЃЈtransforminggrowth factorЉ\ІТЃЌTGFЉ\ІТЃЉЃЌВЂЬсИпМфГфжЪИЩЯИАћЖдЙ§ЖШДЬМЄЕФTЯИАћЕФвжжЦЙІФмЁЃвђДЫЃЌLLЉ\37ПЩФмЬсЙЉЖдЛњ Ьх ЕФ БЃ ЛЄ ЃЌВЂ ЭЈ Й§ Жр жж ЭО ОЖ ЗЂ Лг Ут вп Еї НкзїгУЃл48ЃнЁЃ
1.2.4 ДДЩЫгњКЯ бЊЙмЩњГЩЁЂЩЯЦЄЯИАћЧЈвЦКЭдіжГЁЂГЩЯЫЮЌЯИАћвжжЦНКдЩњГЩЪЧЩЯЦЄДДЩЫгњКЯЕФ 3ИіЙиМќвђЫиЃЌДѓЖрЪ§зїгУЪЧЭЈЙ§МЄЛюБэЦЄЩњГЄвђзгЪмЬхЃЈepidermal growth factor receptorЃЌEGFRЃЉНщЕМЁЃLLЉ\37ЕФДцдкПЩМгЫйGЕААзХМСЊЪмЬхЃЈG proteinЉ\coupledreceptorsЃЌGPCRЃЉЖд EGFR ЗДЪНМЄЛюЃЌДйГЩНЧЛЏЯИАћЧЈвЦЃЌДйНјДДУцаоИДЁЃLLЉ\37ЛЙПЩгеЕМбЊЙмЩњГЩЃЌПЩНЋФкЦЄЧАЬхЯИАћОлМЏЕНЫ№ЩЫашвЊаоИДВПЮЛЃЌМЄЛюФкЦЄЯИАћЃЌДйНјФкЦЄЯИАћдіжГЃЌаЮГЩбЊЙмбљНсЙЙЃл49ЃнЁЃ
Г§ЩЯЪіЙІФмЭтЃЌLLЉ\37ЛЙОпгаПЙВЁЖОЁЂПЙецОњЕШзїгУЃЌетНЋЮЊLLЉ\37ЭЖШыЕНСйДВЪдбщЬсЙЉвРОнЁЃ
2 LL⁃37СйДВжЮСЦЧБСІ
2.1 ВЁЖОадМВВЁ
ЪШЩёОЕЅДП№хеюВЁЖО 1ЃЈherpes simplex virus 1ЃЌHSVЉ\1ЃЉЪЧШЫРр№хеюВЁЖОПЦЕФвЛдБЃЌПЩзїгУгкПкЧЛКЭЩњжГЦї№ЄФЄЃл50ЃнЁЃгабаОПЯдЪО LLЉ\37 Жд HSV ИДжЦгаЯджјгАЯьЁЃИЩШХЫигаПЙВЁЖОзїгУЃЌдкО LLЉ\37 ДІРэЕФИаШО HSVЉ\1 НЧжЪаЮГЩЯИАћжаЃЌЯдЪО 5 ИіживЊИЩШХЫиДЬМЄЛљвђЃлИЩШХЫигеЕМЕФЫФыФжиИДЕААз1ЃЈinterferonЉ\induced protein with tetratricopeptide repeats 1ЃЌIFIT1ЃЉЁЂИЩШХЫиДЬМЄЭтЧаУИЛљвђ 20 kDaЃЈinterferon stimulatedexonuclease gene 20 kDaЃЌISG20ЃЉЕШЃнБэДядіМгЃл51ЃнЁЃдкЬхЭтЯИАћХрбјЪЕбщжаЃЌvero ЯИАћЃЈЗЧжоТЬКяЩіЯИАћЃЉгУ LLЉ\37 дЄДІРэЃЌШЛКѓгУ HSVЉ\1 ИаШОЃЌвВМьВтЕНHSVЉ\1ИДжЦЯджјМѕЩйЃл52ЃнЁЃгабЇепгУжЌжЪЬхLLЉ\37ДІРэ HSVЉ\1ИаШОЕФ HaCaT ЯИАћЃЈШЫгРЩњЛЏНЧжЪаЮГЩЯИАћЃЉЃЌНсЙћЗЂЯжЃЌгыгЮРыLLЉ\37ЖрыФЯрБШЃЌФЩУзжЌжЪЬхLLЉ\37 ЖОадИќаЁЃЌБЃжЪЦкИќГЄЃЌИќвзБЛЮќЪеЁЃвђДЫЃЌжЌжЪЬх LLЉ\37 ЮЊдЄЗР HSVЉ\1 ИаШОЬсЙЉСЫЫМТЗЃЌКмгаПЩФмГЩЮЊЧБСІвЉЮяЃл53ЃнЁЃ
бЯжиМБадКєЮќЯЕЭГзлКЯеїЙкзДВЁЖО2аЭЃЈsevereacute respiratory syndrome coronavirus 2ЃЌSARSЉ\CoVЉ\2ЃЉЪЧв§Ц№ЕБНёСїааЕФаТЙкЗЮбзЕФжївЊВЁдЬхЃЌетжжАќФЄЕЅСДRNAВЁЖОПЩЕМжТМБадКєЮќЯЕЭГМВВЁЃЌбЯжиЪБПЩжТЫРЭіЃл54~56ЃнЁЃЪ§ОнНсЙћБэУїЃЌЮЌЩњЫи D ЩЯЕї LLЉ\37БэДяЃЌЫќПЩжБНгЛђМфНгзїгУгк SARSЉ\CoVЉ\2ЃЌзшжЙЦфИДжЦЃЌдіМгПЙбзЯИАћвђзгЕФВњЩњЃЌМѕЩйДйбзУтвпЗД гІЃл57Ѓн ЁЃВЩ гУ Мф Гф жЪ ИЩ ЯИ АћЃЈmesenchymal stemcellsЃЌMSCЃЉМАЦфбмЩњЮяжЮСЦ SARSЉ\CoVЉ\2 ИаШОЕФСйДВЪдбще§дкНјаажаЁЃMSCВњЩњЖржжПЩШмадвђзгЃЌАќРЈ ПЙ Оњ ыФ LL Љ\ 37ЃЌПЩ гы ВЁ ЖО Нс КЯ ВЂ ЪЙ Цф ИД жЦ ЪЇаЇЃл58ЃЌ59ЃнЁЃШЛЖјЃЌLLЉ\37ПЙ SARSЉ\CoVЉ\2ЕФЛњжЦЩаВЛУїШЗЃЌашНјвЛВНЬНЫїЁЃ
АЃВЉРВЁЖОЃЈEbola virusЃЌEVЃЉЪЧвЛжжРДздЫПзДВЁЖОПЦЕФЕЅСДRNAВЁЖОЃЌгаНЯЧПИаШОадКЭНЯИпЫРЭіТЪЃл60ЃнЁЃзюНќЗЂЯжЃЌLL Љ\37 КЭ LL Љ\37 БфЬхЃЈGI Љ\20 КЭ17BIЃЉдквжжЦжизщаЭЫЎХнадПкбзВЁЖОЃлПЩБэДяEVЉ\ЬЧЕААзЃЈglycoproteinЃЌGPЃЉЃнКЭвАЩњаЭ EV ИаШОжаЦ№ЕНживЊзїгУЁЃетаЉLLЉ\37вЉЮяЭЈЙ§Ы№КІHeLaЯИАћЯЕЃЈШЫЙЌОБАЉЯИАћЃЉжаЕФзщжЏЕААзУИbНщЕМЕФEVЬЧЕААзМгЙЄЃЌФмвжжЦ EVЯИАћНјШыЃЌЕЋВЛФмПижЦВЁЖОИДжЦЁЃИќживЊЕФЪЧЃЌСНжжКЌгаDЉ\АБЛљЫсИФСМПЙОњыФЖдАћФкУИНтОпгаЕжПЙСІЃЌБШ LаЭПЙОњыФИќгааЇЃл61ЃнЁЃетаЉгаЭћНјвЛВНЬНЫїЫќУЧдкЖЏЮяФЃаЭжадЄЗР EV ИаШОзїгУЃЌвдМАгыПЙВЁЖОвЉЮяСЊгУЕФПЩФмадЁЃ
2.2 жзСіадМВВЁ
НсжБГІАЉЃЈcolorectal cancerЃЌCRCЃЉЪЧЕкЖўДѓзюжТУќЕФАЉжЂЃЌеМАЉжЂЫРЭіТЪ 9.2%Ѓл62ЃнЁЃLLЉ\37дке§НсГІ№ЄФЄжаБэДяЧПСвЃЌЕЋдкНсГІжБАЉзщжЏжаБэДяЯТЕїЁЃЕЭLLЉ\37ЫЎЦНвбБЛШЯЮЊЪЧНсжБГІАЉЕФЩњЮяБъжОЮяЁЃЭЌЪБЃЌLLЉ\37ПЩНЕЕЭвжжЦЕђЭіBЯИАћСмАЭСіЉ\2ЛљвђЃЈbЉ\cell lymphomaЉ\2ЃЌBclЉ\2ЃЉЫЎЦНЃЌЬсИпДйЕђЭі Bax/BakЃЈBclЉ\2ЛљвђбЧМвзхГЩдБЃЉЫЎЦНЁЃLLЉ\37вВдіМгСЫP53 ЩЯЕїЕђЭіЕїНквђзгЃЈp53 upregulated modulatorof apoptosisЃЌPUMAЃЉКЭжзСівжжЦЛљвђ p53 ЕФБэДяЃЌPUMA ЪЧ p53 ЕФжБНгАаЕуЃЌвВЪЧНсжБГІАЉЯИАћЕђЭіЕФЕїНквђзгЁЃдкНсГІАЉЯИАћжаЃЌLLЉ\37 ЯджјгеЕМЕђЭігеЕМвђзгЃЈapoptosis inducing factorЃЌAIFЃЉКЭФкЧаУИGЃЈendonuclease GЃЌEndoGЃЉЃЌЕМжТызЬьЕААзУИЃЈcysteinylaspartate specific proteinaseЃЌcaspaseЃЉЗЧвРРЕадЕђЭіЁЃдкНсГІАЉЯИАћжаЃЌвЛжжЛљгкLLЉ\37ЕФЛњжЦМЄЛюGPCRЉ\p53Љ\Bax/Bak/BclЉ\2аХКХМЖСЊЃЌДЅЗЂ AIF/endogНщЕМЕФЕђЭіЃл63ЃЌ64ЃнЁЃЕБLLЉ\37ИНзХдкДХадФЩУзПХСЃЃЈmagneticnanoparticlesЃЌMNPЃЉБэУцЪБЃЌЦфПЙАЉЛюадгаЫљЬсИпЃЌЯдЪОГівЛЖЈЧБСІЃл65ЃнЁЃ
дкЮИАЉЯИАћжаЃЌLLЉ\37 КЌСПЯджјЕЭгке§ГЃЮИзщжЏЁЃгабаОПЯдЪО LLЉ\37 ЭЈЙ§ЙЧаЮЬЌЗЂЩњЕААзЃЈbonemorphogenetic proteinЃЌBMPЃЉаХКХЭЈТЗЕїНкЕААзУИЬхМЄЛюЃл66ЃнЁЃBMPаХКХДЅЗЂSmadЃЈSmaЉ\ and MadЉ\relatedproteinЃЉ1/5/8ЕФФММЏКЭСзЫсЛЏЃЌгыSmad4аЮГЩвьдДЖўОлЬхЃЌИУ Smads ИДКЯЮяЫцКѓЩЯЕїЯИАћжмЦкЕїПиЛљвђp21Waf1ЕФБэДяЫЎЦНЁЃдкИУЭООЖжаЃЌLLЉ\37вВПЩгеЕМЕЭЫЎЦНЕФЯИАћжмЦкЕААз E2ЁЃp21Waf1КЭЯИАћжмЦкЕААзE2 БэДяЫЎЦНЕФИФБфПЩв§Ц№ G0/G1ЦкЯИАћжмЦкзшжЭЁЃЫљвдЃЌLLЉ\37 ЭЈЙ§ЕААзУИЬхвРРЕЕФЛњжЦМЄЛю BMP аХКХЭЈТЗЃЌвжжЦЮИАЉЯИАћдіжГЁЃ
ПкЧЛСлАЉЃЈoral squamous cell carcinomaЃЌOSCCЃЉЪєгкЭЗОБВПГЃМћЖёаджзСіЃЌЦфзщЛЏЗжЮіЯдЪОЃЌгые§ГЃПкЧЛ№ЄФЄзщжЏЯрБШЃЌLLЉ\37 ЕЭБэДягызщжЏбЇЗжЛЏКЭСмАЭНсзЊвЦгаЙиЁЃетЫЕУї LLЉ\37 ПЩФмдк OSCC жаЪЧвЛжжжзСівжжЦвђзгЁЃhCAP18дквЛаЉПкЧЛАЉЯИАћЯЕжаЕФБэДяЕїПиЩцМАЕНШЫРр CAMP ЦєЖЏзгЧјгђЕФDNA МзЛљЛЏзДЬЌЃл68ЃнЁЃвВгабаОПБэУїЃЌhCAP18 дкOSCCSAS Љ\H1 ЯИАћжагеЕМызЬьЕААзУИЗЧвРРЕадЕђЭі ЃЌЕЋдкШЫбРіИГЩЯЫЮЌЯИАћЛђ HaCaT ЯИАћжадђЮоЃл69ЃнЁЃзюНќБЈЕРБэУїЃЌвЛаЉАЉЯИАћОпгаИЛКЌЕЈЙЬДМЕФжЌЗЄЃЌетвВПЩФмЪЧПижЦ LLЉ\37 ЖдВЛЭЌАЉЯИАћбЁдёадЕФЙиМќвђЫиЃЌДгЖјЮЊ LLЉ\37 ПЙАЉЛњжЦЬсЙЉСЫвЛжжаТЕФПЩФмадЃл70ЃЌ71ЃнЁЃ
ДЫЭтЃЌLLЉ\37 дкбЊвКЖёаджзСіжавВЗЂЛгвЛЖЈвжжЦзїгУЃЌЛњжЦПЩФмгыв§Ц№ЯИАћЕђЭіЭЈТЗгаЙиЁЃетаЉНсЙћЮЊжзСіСйДВжЮСЦЬсЙЉСЫЗНЯђЁЃ
3 LL⁃37гІгУЧАОА
3.1 аТвЉ
ЬигІадЦЄбзЃЈatopic dermatitisЃЌADЃЉЛМЖљTLR2КЭLLЉ\37ЕФЕЭБэДяМАбЊЧхLLЉ\37ЕФЕЭЫЎЦНЬсЪОADДцдкЯШЬьУтвпКЭЦЄЗєЦСеЯЙІФмШБЯнЕШЮЪЬтЁЃЮЌЩњЫиDПЩФмМЄЛюЮЌЩњЫиDЪмЬхЃЌЭЈЙ§TLR2ЭООЖДйНјLLЉ\37БэДяЃЌДгЖјЖд AD Ц№ЕНБЃЛЄзїгУЃл72ЃнЁЃЛђаэЮДРДПЩЩшМЦLLЉ\37гаЙивЉЮяИЈжњЮЌЩњЫиDЖдПЙЬигІадЦЄбзЁЃ
аТЩњЖљАмбЊжЂжївЊгЩИаШОв§Ц№ЃЌЫцзХФЭвЉОњЕФВњЩњЃЌПЙЩњЫибЁдёБфЕУдНРДдНМЌЪжЃЌвђДЫЬсИпздЩэЕжПЙСІгШЮЊживЊЁЃгабаОПЯдЪОЮЌЩњЫиDПЩЬсИпЛњЬхПЙОњыФ LLЉ\37 ЕФБэДяЃЌДгЖјЬсИпЛњЬхЗРгљИаШОЕФФмСІЃЌЮЊПЙЩњЫижЮСЦЬсЙЉгаСІБЃеЯЁЃетНЋЮЊЩшМЦПЙЩњЫиЬцДњЦЗЕФвЉЮяЬсЙЉЫМТЗЃл73ЃнЁЃ
3.2 ЖЏЮяЫЧСЯЬэМгМС
LLЉ\37 ПЙОњЦзЙуЧвПЙОњЛюадИпЃЌЯИОњВЛвзВњЩњФЭвЉадЃЌЛЙФмжаКЭФкЖОЫиЃЌПЩвдЪгЮЊПЙЩњЫиЕФаТаЭЬцДњЦЗЁЃвђДЫбЁдёКЯЪЪЕФБэДяФЃАхРДБэДяLLЉ\37ЕФРрЫЦЮяЃЌЖдгкЩњВњОпгаИпаЇПЙОњЁЂИпгЊбјЕФЖЏЮяЫЧСЯЬэМгМСгаДйНјзїгУЃЌЪЙЦфгЕгавЛЖЈЕФОМУаЇвцЁЃ
3.3 ЗРИЏМС
LLЉ\37 КЭ CP1 ЪЧЭЈЙ§ЛљвђЙЄГЬДѓЙцФЃЩњВњЕФЃЌЦфПЙОњЛюадЧПЃЌЯрЖдЗжзгжЪСПаЁЃЌ121ЁцМгШШ30 minПЙОњЛюадИпЃЌЫЎШмадКУЃЌАВШЋЮоЖОЃЌВЛвзВњЩњФЭвЉадЁЃдкХЃФЬжаЬэМгЪЪСППЙОњыФЖджќВигавЛЖЈБЃЯЪзїгУЁЃЯИАћЖОадЪЕбщЫфБэУїЫќУЧЖдДѓЪѓЗЮЯИАћЮоЖОЃЌЕЋжЛФмЮЊПЙОњыФЕФАВШЋадЬсЙЉвРОнЃЌНјвЛВНАВШЋадгІгУгаД§НјвЛВНбаОПЃЈЭМ3ЃЉЃл74ЃнЁЃ
4 еЙЭћ
ПЙОњыФLLЉ\37ЯрЖдЗжзгжЪСПаЁЃЌНсЙЙЬиЪтЃЌЩњЮябЇЙІФмЖрбљЃЌЦфЙуЦзПЙОњЬиадЪмЕНШЫУЧЧрэљЃЌЖјВЛвзВњЩњФЭвЉадЕФаджЪЃЌЪЙЦфКмгаПЩФмГЩЮЊПЙЩњЫиЬцДњЦЗЁЃДЫЭтЃЌПЙОњыФдкЪГЦЗжаЕФЮоЖОадБэЯжЃЌвВгаПЩФмЪЙЦфГЩЮЊаТвЛДњЪГЦЗЬэМгМСЁЃПЙОњыФЕФЪЙгУЃЌПЩИќгааЇЕиЗРжЙЖЏЮявпВЁСїааЃЌМѕЩйаѓФСвЕЫ№ЪЇЃЌетНЋМЋДѓДйНјЮвЙњаѓЧнбјжГвЕНЁПЕЗЂеЙЁЃЪЙгУПЙОњыФПЩдіЧПЖЏЮяЖдМВВЁЕжПЙСІЃЌМѕЩйМВВЁДЋВЅКЭПЙЩњЫиЪЙгУЃЌДгЖјМѕЩйвЉЮядкЖЏЮяЬхФкМАЦфВњЦЗжаЕФВаСєЃЌЮЊЮвЙњЙуДѓЯћЗбепЬсЙЉСЫИќАВШЋЕФШтРрЪГЦЗЁЃгЩгкПЙОњыФОпгаЙуЦзПЙОњадЃЌЖдG+КЭG-ОљгаКмКУвжЩБзїгУЃЌвђДЫПЩЙуЗКгІгУгкаѓЧнбјжГвЕЁЃПЙОњыФОпгаЙуРЋгІгУЧАОАЃЌРћгУЛљвђЙЄГЬММЪѕЩњВњжизщПЙОњыФВњЦЗОпгаКмКУЕФСйДВаЇЙћЁЃЙњФквВгаВПЗжЦѓвЕПЊЗЂГіСЫПЙОњыФВњЦЗЃЈШчЬьВЯЫиПЙОњыФЃЉЃЌдкЖЏЮяЩњВњжаЕУЕНСЫНЯКУгІгУаЇЙћЁЃПЙОњыФЕФбаОПШШГБЪЦБиДјЖЏвЛаЉВњвЕЕФТЬЩЋИяУќЃЌдквНвЉЗНУцЃЌШЫУЧдкХЌСІПЊЗЂТЬЩЋЁЂИпаЇЁЂЕЭЖОПЙИаШОвЉЮяЁЃдкХЉФСвЕЗНУцЃЌПЙОњыФзїЮЊаТаЭЫЧСЯЬэМгМСвВвбЭЖШыЩњВњЃЌЖјЦфдкЪГЦЗЙЄвЕЗНУцвВБэЯжГіОоДѓПЊЗЂЧБСІЁЃЫцзХбаОПЩюШыЃЌПЙОњыФзїЮЊЬэМгМСдкЫЧСЯжаЕФЙуЗКгІгУжеНЋБфГЩПЩФмЁЃПЙОњыФдкЮДРДЗЂеЙжаОпгаЮоЯоЧБСІЃЌгІгшвджиЪгЃЌвдЭкОђЦфЧБдкЕФЩчЛсгІгУМлжЕЁЃ
Утд№ЩљУїЃКБОЮФЮЊаавЕНЛСїбЇЯАЃЌАцШЈЙщдзїепМАддгжОЫљгаЃЌШчгаЧжШЈЃЌПЩСЊЯЕЩОГ§ЁЃЮФеТБъзЂгазїепМАЮФеТГіДІЃЌШчашдФЖСдЮФМАВЮПМЮФЯзЃЌПЩдФЖСддгжО
