
ÕŖŅŖ £ŗÓŠŠ©ĢģČ»µ°°×ÖŹæÉĶعż“ķĪóÕŪµžŠĪ³Éµķ·ŪŃłĻĖĪ¬£¬²¢½ųŅ»²½³Į»żµ¼ÖĀµķ·ŪŃł²”±ä£¬±»ČĻĪŖŹĒŠķ¶ąÖŲ“óČĖĄą¼²²”µÄ²”Ąķ»ł“”£¬Ņņ“Ė£¬²ūĆ÷ĢģČ»µ°°×ÖŹ“ķĪóÕŪµž”¢¾Ū¼ÆŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ·Ö×Ó»śÖĘ£¬ŹĒŌ¤·Ą”¢Õļ¶ĻŗĶÖĪĮĘĻą¹Ų¼²²”µÄ¹Ų¼ü”£ŃŠ¾æÕßĆĒ“ÓĢģČ»µ°°×ÖŹÖŠ¼ų¶Ø³öŠķ¶ąÄܹ»ŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ¹Ų¼ü¶ĢėÄʬ¶Ī£¬¼“µķ·ŪŃł¶ĢėÄ£¬¶ŌĖüĆĒŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄÄÜĮ¦¼°ĘäŌŚĶźÕūµ°°×ÖŹ¾Ū¼Æ¹ż³ĢÖŠµÄ¾ö¶ØŠŌ×÷ÓĆ½ųŠŠÉīČėŃŠ¾æ£¬ ±¾ĪĶŌ½üÄźĄ“ČĖĄą¼²²”Ļą¹Ųµķ·ŪŃł¶ĢėĵÄŃŠ¾æÕ¹æŖ×ŪŹö”£Ź×ĻČ£¬½éÉܼų¶Øµķ·ŪŃł¶Ģėĵıź×¼¼°ĘäĻąÓ¦µÄŃŠ¾æ·½·ØŗĶ¼¼ŹõŹÖ¶Ī£¬²¢»Ų¹Ė½üÄźĄ“ÓėŅ»Š©ÖŲ“óČĖĄą¼²²”Ļą¹ŲµÄµķ·ŪŃł¶ĢėÄ£¬ÓČĘäŹĒÓėÉń¾ĶĖŠŠŠŌ¼²²”Ļą¹Ųµķ·ŪŃł¶ĢėĵĽųÕ¹Ēéæö£¬¶Ōµķ·ŪŃł¶ĢėÄÖŠ³öĻÖʵĀŹ½Ļøߵݱ»łĖį²Š»ł¼°ĘäæÉÄܵÄ×Ō×é×°ŌĄķ½ųŠŠ×ܽį·ÖĪö”£×īŗó£¬Õ¹ĶūÕāŠ©µķ·ŪŃł¶ĢėÄ×÷ĪŖ°ŠµćŌŚĻą¹Ų¼²²”Õļ¶ĻŗĶÖĪĮĘ·½ĆęµÄŅāŅ壬²¢³õ²½Ģ½ĢÖĖüĆĒ×÷ĪŖŠĀŠĶÉśĪļ²ÄĮĻŌŚÉśĪļŅ½Ń§¹¤³ĢĮģÓņµÄÓ¦ÓĆĒ°¾°£¬±¾ĪÄŅ»·½ĆęĪŖ²ūĆ÷ĢģČ»µ°°×ÖŹŠĪ³Éµķ·ŪŃł³ĮµķµÄ·Ö×Ó»śÖĘĢį¹©²Īæ¼£¬ĮķŅ»·½ĆęŅ²ĪŖĻą¹Ų¼²²”µÄÖĪĮĘĢį¹©Ė¼Ā·£¬Ķ¬Ź±Ņ²ĪŖŠĀŠĶÉśĪļ²ÄĮĻµÄæŖ·¢Ģį³öĒ±ŌŚµÄæÉÄÜŠŌ”£
µķ·ŪŃłĻĖĪ¬ŹĒŅ»ĄąĢŲŹāµÄÓÉĢģČ»µ°°×ÖŹ¾Ū¼ÆŠĪ³ÉµÄÄÉĆ×ĻĖĪ¬½į¹¹£¬×īŌē½ųČėČĖĆĒµÄŹÓŅ°ŹĒÓÉÓŚĖüĆĒÓėČĖĄą¼²²”µÄ·¢ÉśÓŠ×ÅĆÜĒŠĮŖĻµ£¬ĻÖŅŃÖ¤Źµ£¬ČĖĢåÖŠµÄijŠ©µ°°×ÖŹÄܹ»ŌŚŅ»¶ØĢõ¼žĻĀ·¢Éś¹¹Ļó±ä»Æ£¬ĶعżŅģ³£µÄ¾Ū¼ÆŠĪ³Éµķ·ŪŃłĻĖĪ¬£¬²¢½ųŅ»²½ŌŚ×éÖÆÖŠ³Į»żŠĪ³É²»ČÜŠŌµÄµķ·ŪŃł°ßæ飬×īÖÕµ¼ÖĀµķ·ŪŃł²”±äµÄ·¢Éś£¬ ÕāŅ»²”±ä¹ż³Ģ²»µ«Ź¹µ°°×ÖŹÉ„Ź§ŌÓŠµÄÕż³£¹¦ÄÜ£¬Ņ²Ōģ³ÉĖłŌŚ×éÖÆÖÜĪ§µÄĻø°ūĖĄĶö£¬±»ČĻĪŖŹĒÓÕ·¢Šķ¶ąČĖĄą¼²²”µÄ¹Ų¼üŅņĖŲ”£ÄæĒ°ŅŃÖŖ£¬ÖĮÉŁÓŠ20¶ąÖÖČĖĄą¼²²”µÄ·¢ÉśÓėµķ·ŪŃłĻĖĪ¬µÄŠĪ³ÉĆÜĒŠĻą¹Ų£¬ĘäÖŠ°üĄØ°¢×Čŗ£Ä¬Ö¢[£±]”¢ÅĮ½šÉÖ¢[£²]”¢¼”Ī®Ėõ²ąĖ÷Ó²»Æ[£³] µČÉń¾ĶĖŠŠŠŌ¼²²”£¬ŅŌ¼°ĢĒÄņ²”ŗĶ¶ÆĀöÖąŃłÓ²»ÆµČ³£¼ūµÄĄĻÄź¼²²”[£“]£¬ÕāŠ©¼²²”·¢ÉśµÄ¾ßĢå»śÖĘÄæĒ°ČŌ²»Ē峞£¬²¢ĒŅŅ»µ©·¢ÉśŗÜÄŃÄę×Ŗ£¬øųĻÖ“śĄĻĮä»ÆÉē»įŌģ³É³ĮÖŲµÄøŗµ££¬Ņņ“Ė£¬ŃŠ¾æµ°°×ÖŹŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ·Ö×Ó»śÖĘ£¬“Ó¶ų½ųŅ»²½²ūĆ÷Ļą¹Ų¼²²”·¢ÉśµÄ»śÖĘ£¬¶ŌÓŚµķ·ŪŃł²”±äĻą¹Ų¼²²”µÄŌ¤·Ą”¢Õļ¶ĻŗĶÖĪĮĘÓŠ×ÅŹ®·ÖÖŲŅŖµÄŅāŅ唣
³żĮĖÓė¼²²”Ļą¹ŲµÄµķ·ŪŃłĻĖĪ¬£¬½üÄźĀ½Šų·¢ĻÖŅ»Š©ÓėČĖĄą¼²²”·¢ÉśĪŽ¹ŲµÄµ°°×ÖŹŅ²ÄÜŌŚŅ»¶ØĢõ¼žĻĀŠĪ³Éµķ·ŪŃłĻĖĪ¬[5,6]£¬ÉõÖĮ×ŌČ»½ēÖŠ»¹“ęŌŚŅ»Š©¹¦ÄÜŠŌµÄµķ·ŪŃłĻĖĪ¬£¬ĄżČēĻø¾śµÄ¾śĆ«ŗĶŗŚÉ«ĖŲĢåÖŠµÄPMEL17µ°°×[7]£¬ÕāŠ©Ēéæö±ķĆ÷£¬µ°°×ÖŹŠĪ³Éµķ·ŪŃłĻĖĪ¬ŹĒ×ŌČ»½ēŅ»øöĘÕ±é“ęŌŚµÄĻÖĻó£¬ŃŠ¾æµ°°×ÖŹŌŚĢģȻדĢ¬ĻĀŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ·Ö×Ó»śÖĘ£¬Ņ²ŹĒ²ūĆ÷Ęä½į¹¹ŗĶ¹¦ÄܹŲĻµµÄÖŲŅŖ»·½Ś£¬Ķ¬Ź±Ņ²ÄÜĪŖĪŅĆĒæŖ·¢ŠĀŠĶÄÉĆ×ĻĖĪ¬²ÄĮĻ“ųĄ“Ęō·¢”£
ÓÉÓŚŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄµ°°×ÖŹ·Ö×ÓĶس£·Ö×ÓĮæ½Ļ“󣬊ņĮŠ½Ļ³¤²¢ĒŅ½į¹¹ø“ŌÓ£¬Ö±½ÓŃŠ¾æĖüĆĒŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ·Ö×Ó»śÖĘ±Č½ĻĄ§ÄŃ£¬ÄæĒ°ČŌĪ“¼ūČĪŗĪŅ»øöµ°°×ÖŹŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ»śÖʱ»ĶźČ«²ūĆ÷£¬ĮķŅ»øöÓŠŅāĖ¼µÄĻÖĻóŹĒ£¬¾”¹Ü×ŌČ»½ēÖŠÄܹ»¾Ū¼ÆŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄµ°°×ÖŹø÷²»ĻąĶ¬£¬ĖüĆĒŌŚ×ÜĢåŠņĮŠÉĻŅ²Ć»ÓŠĆ÷ĻŌµÄĻąĖĘŠŌ£¬µ«ĖüĆĒČ“Äܹ»ŠĪ³É¾ßÓŠĻąĶ¬ĢŲÕ÷µÄµķ·ŪŃłĻĖĪ¬[£ø]£¬ÕāĖµĆ÷£¬²»Ķ¬µÄµ°°×ÖŹŌŚÓÕµ¼¾Ū¼ÆµÄ¹Ų¼üŠņĮŠ¼°Ęä¾Ū¼ÆµÄ»śÖĘÉĻ±ŲȻӊŅ»¶ØµÄĻąĖĘŠŌ”£Ņņ“Ė£¬ÄæĒ°Į÷ŠŠµÄ°ģ·ØŹĒŃŠ¾æµ°°×ÖŹŠņĮŠÖŠ¹Ų¼üµÄ¶ĢėÄʬ¶Ī£¬ÕāŠ©¶ĢėÄʬ¶Ī×ŌÉķÄܹ»¾Ū¼ÆŠĪ³ÉµäŠĶµÄµķ·ŪŃłĻĖĪ¬£¬²¢ĒŅŌŚĶźÕūµ°°×ÖŹŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ¹ż³ĢÖŠ·¢»Ó¾ö¶ØŠŌ×÷ÓĆ£¬Ņ²±»³ĘĪŖµķ·ŪŃł¶ĢėÄ£¬Ķعż¶ŌÕāŠ©¶ĢėÄ×Ō×é×°ŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ»śÖĘ½ųŠŠŃŠ¾æ£¬Ņ²ŠķÄÜŌŚµ°°×ÖŹÕūĢåŠņĮŠĻąĖĘŠŌ²»“óµÄĒéæöĻĀ£¬ÕŅµ½¾Ö²æŠņĮŠÉĻµÄĻąĖĘŠŌ£¬“Ó¶ų½āŹĶ²»Ķ¬µÄµ°°×ÖŹŠĪ³ÉĻąĖʵĵķ·ŪŃłĻĖĪ¬µÄŌĄķ£¬¾ßÓŠÖŲŅŖµÄĄķĀŪŅāŅ唣ĮķŅ»·½Ćę£¬ŃŠ¾æµķ·ŪŃł¶ĢėĵÄ×Ō×é×°»śÖĘ¼°Ęä¶ŌĶźÕūµ°°×ÖŹ¾Ū¼ÆŠŠĪŖµÄÓ°Ļģ£¬ÓŠĄūÓŚ²ūĆ÷ÕāŠ©µ°°×ÖŹŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ·Ö×Ó»śÖĘ£¬“Ó¶ųĪŖĻą¹Ų¼²²”µÄŌ¤·Ą”¢Õļ¶ĻŗĶÖĪĮĘĢį¹©ÖŲŅŖµÄĄķĀŪ»ł“”(¼ūFig.1)”£
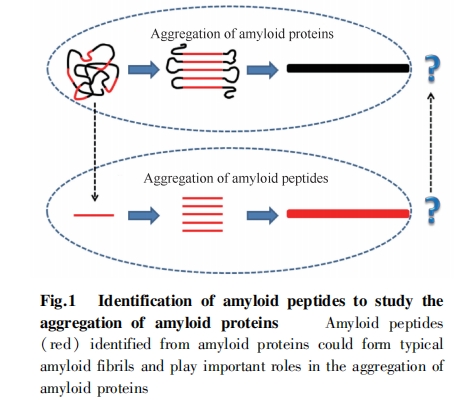
½üÄźĄ“£¬ŌŚŠķ¶ąÓėČĖĄą¼²²”Ļą¹ŲµÄŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄµ°°×ÖŹÖŠ£¬¶¼¼ų¶Ø³öÕāŃłµÄ¶ĢėÄʬ¶Ī”£ÕāŠ©Ą“×Ō²»Ķ¬µ°°×ÖŹµÄ¶ĢėÄʬ¶ĪŌŚŠņĮŠĢŲÕ÷ÉĻĢåĻÖ³öŅ»¶ØµÄĻąĖĘŠŌ£¬Ņ²ĪŖ½ųŅ»²½²ūĆ÷ĶźÕūµ°°×ÖŹŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ»śÖĘĢį¹©ĮĖĖ¼Ā·”£±¾ĪĽ«Ö÷ŅŖÕė¶Ō“Óø÷ÖÖČĖĄą¼²²”Ļą¹Ųµķ·ŪŃłµ°°×ÖŠ·¢ĻֵĶĢėÄÕ¹æŖ×ŪŹö£¬ŅŌĘŚĪŖ×īÖÕ²ūĆ÷µķ·ŪŃłĻĖĪ¬ŠĪ³ÉµÄ·Ö×Ó»śÖĘ£¬²¢Öøµ¼Ļą¹Ų¼²²”µÄŌ¤·Ą”¢Õļ¶ĻŗĶÖĪĮĘĢį¹©ÓŠÓƵÄĻßĖ÷”£
£± µķ·ŪŃł¶Ģėĵļų¶Ø
ŌŚµķ·ŪŃł²”±äÖŠ£¬ŗź¹ŪÉĻ±ķĻÖĪŖµķ·ŪŃł°ßæéµÄ²”ĄķŠŌ³Į»ż£¬ŌŚĪ¢¹ŪÉĻŹĒÓɵ°°×ÖŹŅģ³£¾Ū¼ÆŠĪ³ÉµÄÄÉĆ×¼¶µķ·ŪŃłĻĖĪ¬²ų½į¶ų³É”£ÄæĒ°£¬µķ·ŪŃłĻĖĪ¬µÄ¼ģ²āÖ÷ŅŖÓĆŅŌĻĀµäŠĶĢŲÕ÷ĪŖÖø±ź:ĶāŠĪĪŖÖ±¾¶Ō¼10nm£¬¹ā»¬²»·Ö²ęµÄÄÉĆ×ĻĖĪ¬£¬×é³Éµķ·ŪŃłĻĖĪ¬µÄµ°°×ÖŹ»ņ¶ĢėijŹµäŠĶµÄ¦ĀÕŪµž¶ž¼¶½į¹¹£¬¾øÕ¹ūŗģČ¾É«Ź±£¬µķ·ŪŃłĻĖĪ¬ŌŚĘ«¹āĻŌĪ¢¾µĻĀ³ŹĻÖ³ö»ĘĀĢÉ«µÄĖ«ÕŪÉä¹ā£¬µķ·ŪŃłĻĖĪ¬ÓėĮņ»ĒĖŲT½įŗĻŹ±£¬»įŌŚ495nmø½½ü³öĻÖĢŲÕ÷ŠŌµÄÓ«¹ā·å”£ÄæĒ°£¬ÉĻŹöĢŲÕ÷ŅŃ³ÉĪŖ¼ų¶Øµķ·ŪŃłĻĖĪ¬µÄ½š±ź×¼[9]£¬¶ų“ÓĢģČ»µ°°×ÖŹµÄŠņĮŠÖŠ¼ų¶Øµķ·ŪŃł¶Ģėĵď×ŅŖ±ź×¼£¬¾ĶŹĒÕāŠ©¶ĢėÄŅŖÄܹ»×Ō×é×°ŠĪ³É¾ßÓŠÉĻŹöĢŲÕ÷µÄµķ·ŪŃłĻĖĪ¬”£
“ĖĶā£¬ŃŠ¾æµķ·ŪŃł¶ĢėĵľŪ¼ÆŠŠĪŖ£¬×īÖÕµÄÄæµÄŹĒŃŠ¾æĘäĶźÕūµ°°×ÖŹŅģ³£¾Ū¼Æµ¼ÖĀ¼²²”µÄ»śÖĘ£¬ Ņņ“Ė£¬Ņ»øöÓŠŃŠ¾æ¼ŪÖµµÄµķ·ŪŃł¶ĢėÄ£¬²»½ö×ŌÉķÄܹ»¾Ū¼ÆŠĪ³Éµķ·ŪŃłĻĖĪ¬£¬»¹ÄܶŌĘäĖłŌŚµ°°×ÖŹµÄ¾Ū¼ÆŠŠĪŖ·¢»ÓÖŲŅŖµÄ×÷ÓĆ£¬ ĄżČē£¬µķ·ŪŃł¶ĢėÄʬ¶ĪµÄȱŹ§»ņĶ»±ä£¬Äܹ»ŌŚŅ»¶Ø³Ģ¶ČÉĻĘĘ»µøƵ°°×ÖŹµÄ¾Ū¼ÆŠŠĪŖ”£
ĪÄĻ×ÖŠĶس£°Ń¾ßÓŠÉĻŹöĮ½øöĢŲµć£¬Ą“×ŌĢģČ»µ°°×ÖŹµÄ”¢³¤¶ČĪŖ¼øøöµ½Ź®¼øøö°±»łĖį²Š»łµÄ¶ĢėÄʬ¶Ī³ĘĪŖµķ·ŪŃł¶ĢėÄ£¬ÕāŠ©¶ĢėÄÄæĒ°ŅŃ³ÉĪŖµķ·ŪŃł²”±äĻą¹Ų¼²²”ĮģÓņµÄŃŠ¾æČČµć”£“ĖĶā£¬Ņ²ÓŠŅ»Š©ČĖĪŖÉč¼ĘµÄ¶ĢėÄʬ¶Ī£¬ĘäÄܹ»×Ō×é×°ŠĪ³É¾ßÓŠµäŠĶµķ·ŪŃłĻĖĪ¬ĢŲÕ÷µÄÄÉĆ×ĻĖĪ¬£¬ÓŠŹ±Ņ²±»³ĘĪŖµķ·ŪŃł¶ĢėÄ”£
£² ŃŠ¾æµķ·ŪŃł¶Ģėĵķ½·Ø
Õė¶ŌÉĻŹö¼ų¶Øµķ·ŪŃł¶Ģėĵıź×¼£¬ÓŠŠķ¶ą¾µäµÄ¼ģ²ā¼¼ŹõŗĶŃŠ¾æ·½·Ø£¬µķ·ŪŃł¶ĢėÄĻą¹ŲŃŠ¾æµÄ¹ć·ŗæŖÕ¹£¬µĆŅęÓŚÕāŠ©¼¼Źõ·½·ØµÄĶźÉĘŗĶĘÕ¼°”£ĘäÖŠ£¬±ķÕ÷ÄÉĆ×ĻĖĪ¬½į¹¹µÄĶøÉäµē¾µŗĶŌ×ÓĮ¦ĻŌĪ¢¾µ”¢±ķÕ÷¶ž¼¶½į¹¹µÄŌ²¶žÉ«¹āĘ×”¢£ŲÉäĻßŃÜÉäŗĶŗģĶā¹āĘ×£¬ŅŌ¼°øÕ¹ūŗģČ¾É«”¢Įņ»ĒĖŲ£ŌČ¾É«µČČ¾É«¼¼Źõ£¬ŅŃ³ÉĪŖ³£ÓĆµÄ¼¼ŹõŹÖ¶Ī£¬ĢŲ±šŹĒĮņ»ĒĖŲ£ŌČ¾É«£¬ĖüÄܹ»ŌŚČÜŅŗדĢ¬ĻĀŹµŹ±¼ģ²āµķ·ŪŃłĻĖĪ¬µÄŠĪ³É£¬ŅŃ³ÉĪŖ³õ²½ÓėæģĖŁ¼ų¶Øµķ·ŪŃł¶ĢėÄ£¬²¢ŃŠ¾æĘä¾Ū¼Æ¶ÆĮ¦Ń§µÄÖŲŅŖ·½·Ø”£
³żĮĖÉĻŹö³ÉŹģµÄ¼¼ŹõŹÖ¶Ī£¬µķ·ŪŃł¶ĢėĵÄŃŠ¾æĶ¬Ź±Ņ²µĆŅęÓŚČÕ½„³ÉŹģµÄ¶ąėĹĢĻąŗĻ³É¼¼Źõ£¬ŃŠ¾æĶźÕūµ°°×ÖŹŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ»śÖĘ£¬Ķس£ŠčŅŖ¹¹½ØĻąÓ¦µ°°×ÖŹµÄ±ķ“ļŌŲĢ壬ŌŚŌŗĖ»ņÕęŗĖĻµĶ³ÖŠ½ųŠŠ±ķ“ļ“æ»Æ£¬²»½ö·ŃŹ±·ŃĮ¦£¬¶ųĒŅŗÜÄŃ»ńµĆøß“æ¶ČµÄµ°°×ÖŹ½ųŠŠŃŠ¾æ£¬¶ų³£¼ūµÄµķ·ŪŃł¶ĢėÄŠņĮŠ£¬³¤¶Č¶ąŌŚ¼øøöÖĮŹ®¼øøö°±»łĖį²Š»łÖ®¼ä£¬Ņņ“Ė·Ē³£ČŻŅץūÓƶąėĹĢĻąŗĻ³É¼¼Źõ£¬»ńµĆøß“æ¶ČµÄ¶ĢėĽųŠŠŃŠ¾æ”£“ĖĶā£¬¼ĘĖ滜øØÖśµÄ·ÖĪö¼¼Źõ£¬ĄżČē·Ö×Ó¶ÆĮ¦Ń§Ä£Äā£¬Ņ²ŌŚ½üÄźĄ“±»¹ć·ŗÓĆÓŚ¶Ōµķ·ŪŃł¶ĢėĵľŪ¼ÆŠŠĪŖ½ųŠŠÄ£Äā·ÖĪö[10].ŌŚÕāŠ©³ÉŹģ¶ųĻČ½ųµÄ¼¼ŹõµÄÖ§³ÖĻĀ£¬µķ·ŪŃł¶ĢėĵÄŃŠ¾æŅ»Ö±ŹĒ½üÄźĄ“¹ś¼ŹÉĻµÄŃŠ¾æČČµć”£
£³ ø÷ÖÖ¼²²”ÖŠµÄµķ·ŪŃł¶ĢėÄ
3.1 Éń¾ĶĖŠŠŠŌ¼²²”ÖŠµÄµķ·ŪŃł¶ĢėÄ
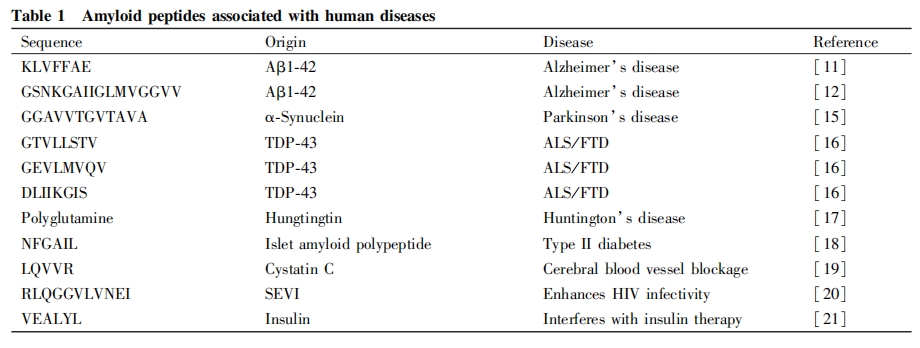
ŌŚø÷ÖÖ¼²²”Ļą¹Ųµķ·ŪŃłĻĖĪ¬µÄŃŠ¾æÖŠ£¬ÓėÉń¾ĶĖŠŠŠŌ¼²²”Ļą¹ŲµÄµķ·ŪŃłĻĖĪ¬ŹĒµ±Ē°ŃŠ¾æµÄÖŲµć”£¶ųŌŚÄæĒ°ŅŃ±ØµĄµÄµķ·ŪŃł¶ĢėÄÖŠ£¬ŃŠ¾æµĆ×ī¶ąµÄŹĒÓė°¢×Čŗ£Ä¬Ö¢Ļą¹ŲµÄµķ·ŪŃł¶ĢėÄA¦Ā16-22”£°¢×Čŗ£Ä¬Ö¢ŹĒÄæĒ°×īĮ÷ŠŠµÄÉń¾ĶĖŠŠŠŌ¼²²”£¬Ęä·¢²”»śÖĘÓė»¼ÕßÄŌ²æµķ·ŪŃł¶ąėÄA¦Ā1-42Ņģ³£¾Ū¼Æ£¬²¢ŠĪ³Éµķ·ŪŃł°ßæéĆÜĒŠĻą¹Ų£¬ŌŚĪ§Čʵķ·ŪŃł¶ąėÄA¦Ā1-42¾Ū¼ÆŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ·Ö×Ó»śÖĘ½ųŠŠµÄ“óĮæŃŠ¾æÖŠ£¬·¢ĻÖøƶąėÄÖŠÓɵŚ16Ī»ÖĮ22Ī»°±»łĖį²Š»ł×é³ÉµÄŅ»øö7ėÄʬ¶ĪKLVFFAE (¼“A¦Ā16-22)Äܹ»×Ō×é×°ŠĪ³ÉŅŌ¦ĀÕŪµžĪŖ»ł“”µÄµķ·ŪŃłĻĖĪ¬£¬ŌŚA¦Ā1-42µÄŅģ³£¾Ū¼Æ¹ż³ĢÖŠ·¢»Ó׏Ų¼ü×÷ÓĆ[11]”£×ī½üµÄŃŠ¾æ±ØµĄ±ķĆ÷£¬A¦Ā1-42ÖŠÓɵŚ25Ī»ÖĮ40Ī»°±»łĖį²Š»ł×é³ÉµÄʬ¶ĪA¦Ā25-40£¬Ņ²¾ßÓŠŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄÄÜĮ¦£¬æÉÄܶŌA¦Ā1-42µÄ¾Ū¼ÆŅ²·¢»ÓĮĖÖŲŅŖ×÷ÓĆ[12]£¬×ī½üÓŠŃŠ¾æ·¢ĻÖ£¬Ļą±ČÓŚŠĪ³Éµķ·ŪŃłĻĖĪ¬£¬£Į¦Ā ŠĪ³ÉµÄ¹Ń¾ŪĢå·“¶ųŹĒĻø°ū¶¾ŠŌøüĒæµÄ¾Ū¼ÆדĢ¬£¬¶ųŠĪ³Éµķ·ŪŃłĻĖĪ¬ÓŠæÉÄÜŹĒ½µµĶµķ·ŪŃł¾Ū¼ÆĪļĻø°ū¶¾ŠŌµÄŅ»ÖÖ×ŌĪŅ±£»¤»śÖĘ[13]£¬Ņņ“Ė£¬½«Ą“¶ŌÓŚ £Į¦Ā ÖŠÕāĮ½øö¹Ų¼üʬ¶ĪµÄŃŠ¾æ£¬Ņ²Ó¦øĆøü¶ą¹Ų×¢ĖüĆĒŌŚ¹Ń¾ŪĢåŠĪ³É¹ż³ĢÖŠµÄ×÷ÓĆ”£
ÅĮ½šÉÖ¢ŹĒ¼Ģ°¢×Čŗ£Ä¬Ö¢Ö®ŗóµÄµŚ¶ž“óÉń¾ĶĖŠŠŠŌ¼²²”£¬Ęä²”ĄķĢŲÕ÷±ķĻÖĪŖ»¼ÕßÄŌ²æ“óĮæLewyŠ”ĢåµÄŠĪ³ÉŗĶ²ś¶ą°Ķ°·ÄÜÉń¾ŌŖµÄĖĄĶö”£ĻÖŅŃÖ¤Źµ£¬LewyŠ”ĢåµÄÖ÷ŅŖ³É·ŻŹĒÓÉ140øö°±»łĖį²Š»ł×é³ÉµÄ¦Į-Ķ»“„ŗĖµ°°×ĶعżŅģ³£¾Ū¼ÆŠĪ³ÉµÄµķ·ŪŃłĻĖĪ¬£¬øƵ°°×ÖŹÖŠÓɵŚ61-95Ī»°±»łĖį×é³ÉµÄ·Ē¦Āµķ·ŪŃłėÄ(NAC) ĒųÓņŗ¬ÓŠ“óĮæŹčĖ®ŠŌ°±»łĖį²Š»ł£¬±»ČĻĪŖŹĒŠĪ³É¦ĀÕŪµžŗĶµķ·ŪŃłĻĖĪ¬µÄÖ÷ŅŖĒųÓņ[14]£¬ĮķÓŠŃŠ¾æ±ķĆ÷£¬¦Į-Ķ»“„ŗĖµ°°×ÖŠµŚ67ÖĮ78Ī»°±»łĖį²Š»ł×é³ÉµÄʬ¶Ī£¬¶Ōµķ·ŪŃłĻĖĪ¬µÄŠĪ³É¼°Ęä¶ŌÉń¾Ļø°ūµÄ¶¾ŠŌ·¢»Ó¹Ų¼ü×÷ÓĆ[15]”£
¼”Ī®Ėõ²ąĖ÷Ó²»Æ (ALS)ŹĒŅ»ÖÖĘĘ»µ»¼ÕßÉń¾Ļø°ū£¬²¢µ¼ÖĀ¼”ČāÖš½„ĪŽĮ¦ŗĶĪ®ĖõµÄÉń¾ĻµĶ³¼²²”£¬Ķس£Óė¶īņØŅ¶ŠĶŹ§ÖĒÖ¢(FTD)ŗĻ²¢·¢×÷£¬Ņņ“ĖĮ½ÕßŗÜæÉÄܾßÓŠĶ³Ņ»µÄ·¢²”»śÖĘ”£ŃŠ¾æ·¢ĻÖ£¬·“Ź½¼¤»ī·“Ó¦DNA½įŗĻµ°°× (TDP-43)æÉŠĪ³É·ŗĖŲ»ÆµÄ°üŗĢåŌŚĻø°ūÄŚ³Į»ż£¬ŅżĘš¼”Ī®Ėõ²ąĖ÷Ó²»ÆŗĶ¶īņØŅ¶ŠĶŹ§ÖĒÖ¢µÄ·¢Éś£¬Ņņ“ĖTDP-43·¢ÉśŅģ³£¾Ū¼ÆµÄ·Ö×Ó»śÖĘŅ²ŹĒŃŠ¾æÕßĆĒ¹Ų×¢µÄÖŲµć£¬ ×ī½ü£¬KumarµČ[16] “ÓTDP-43ŠņĮŠĄļ¼ų¶Ø³ö£³øöÄܹ»ŌŚĢåĶā×Ō·¢¾Ū¼ÆŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄʬ¶Ī:GTVLLSTV”¢GEVLMVQVŗĶDLIIKGIS£¬ÕāŠ©Ę¬¶ĪŗÜæÉÄÜÖ÷µ¼ĮĖČ«³¤TDP-43µ°°×ŌŚĢåÄŚµÄ¾Ū¼ÆŠŠĪŖ”£
ŗąĶ¢¶ŁĪčµø²”ŹĒŅ»ÖÖŗ±¼ūµÄÉń¾ĶĖŠŠŠŌ¼²²”£¬ŅŌÓ°ĻģŌĖ¶Æ¹¦ÄÜĪŖÖ÷£¬Ķ¬Ź±Ņ²»į½„½ųŠŌµŲµ¼ÖĀČĻÖŖŗĶ¾«Éń·½ĆęµÄÕĻ°£¬ŗąĶ¢¶ŁĪčµø²”µÄ·¢Éś±»ČĻĪŖŹĒÓÉ3144øö°±»łĖį²Š»ł×é³ÉµÄµ°°×ÖŹHungtingtinµÄĶ»±äŗĶ¾Ū¼Æµ¼ÖĀµÄ£¬Ī»ÓŚøƵ°°×ÖŹNÄ©¶ĖµÄÖŲø“¹Č°±õ£°·ŠņĮŠ(polyglutamine)ŹĒµ°°×ÖŹ·¢ÉśŅģ³£¾Ū¼ÆµÄ¹Ų¼üŠņĮŠ£¬ŃŠ¾æ±ķĆ÷£¬øĆŠņĮŠÖŠ¹Č°±õ£°·µÄŹżÄæŌ½¶ą£¬HungtingtinŌ½ČŻŅ×·¢Éś¾Ū¼Æ[17]”£
ŌŚÉĻŹöÕāŠ©Éń¾ĶĖŠŠŠŌ¼²²”ÖŠ£¬µķ·ŪŃł³Įµķ“ÓŠī»żµ½ÖĀ²”ŹĒŅ»øö³¤ĘŚŗĶ»ŗĀżµÄ¹ż³Ģ£¬ŌēĘŚ¼«ÄŃ·¢ĻÖ£¬¶ųŅ»µ©·¢²”Ö®ŗóÓÖÄŃŅŌÄę×Ŗ£¬Ņņ“Ė£¬ŃŠ¾æÕāŠ©¼²²”µÄ·¢Éś»śÖĘŹĒµ±Ē°ÉśĪļŅ½Ń§·Ē³£ČČĆŵÄŃŠ¾æĮģÓņ£¬¶ųÕāŠ©¼²²”ÖŠĻąÓ¦µķ·ŪŃł¶ĢėĵÄŃŠ¾æ£¬Ņ²ŅņĘäŌŚ½ŅŹ¾µķ·ŪŃł³ĮµķŠĪ³ÉµÄ·Ö×Ó»śÖĘ·½ĆęµÄÖŲŅŖŅāŅ壬ŹÜµ½¹ć·ŗµÄÖŲŹÓ”£
3.2 ĘäĖū¼²²”Ļą¹Ųµķ·ŪŃł¶ĢėÄ
Ņȵŗµķ·ŪŃł¶ąėÄŹĒŗ¬ÓŠ37øö°±»łĖį²Š»łµÄ¶ąėļ¤ĖŲ£¬¾ßÓŠµ÷½ŚĢĒ“śŠ»µČ¶ąÖÖÉśĪļѧ×÷ÓĆ£¬ŅŃÓŠŃŠ¾æ±ķĆ÷£¬Ņȵŗµķ·ŪŃł¶ąėÄŌŚŅȵŗÖŠµÄŅģ³£³Į»żÓėIIŠĶĢĒÄņ²”µÄ·¢ÉśĆÜĒŠĻą¹Ų£¬¶ŌĘäŠņĮŠµÄÉīČėŃŠ¾æ·¢ĻÖ£¬Ņȵŗµķ·ŪŃł¶ąėÄÖŠµÄŅ»øöNFGAILʬ¶Ī£¬Äܹ»ŌŚĢåĶā×Ō×é×°ŠĪ³ÉµäŠĶµÄµķ·ŪŃłĻĖĪ¬£¬ŌŚøƶąėÄ·¢Éśµķ·ŪŃł¾Ū¼ÆµÄ¹ż³ĢÖŠ·¢»Ó¾ö¶ØŠŌ×÷ÓĆ[18]
ČĖė×ŅÖĖŲ( cystatin)CŹĒÓÉ120øö°±»łĖį²Š»ł×é³ÉµÄŠ”·Ö×Óµ°°×ÖŹ£¬ŹōÓŚė×ŅÖĖŲ¼Ņ×壬¹ć·ŗ“ęŌŚÓŚČĖĢåø÷øö²æĪ»µÄĻø°ūŗĶĢåŅŗÖŠ£¬ė×ŅÖĖŲCĶعżŅÖÖĘ°ėė×°±Ėįµ°°×Ćø»īŠŌ£¬ŌŚŠķ¶ąÕż³£ÉśĄķ»ī¶ÆÖŠ·¢»Óµ÷æŲ×÷ÓĆ£¬ŌŚ²”ĄķĢõ¼žĻĀ£¬ė×ŅÖĖŲCŌŚÄŌ²æ¶ÆĀöŃŖ¹ÜÖŠŠĪ³Éµķ·ŪŃł³Į»ż£¬¶ĀČūŃŖ¹Ü£¬ŃĻÖŲŹ±ÉõÖĮÓÕ·¢Ī£¼°ÉśĆüµÄÄŌŅēŃŖ£¬ŃŠ¾æ±ķĆ÷£¬ė×ŅÖĖŲCÖŠŠņĮŠĪŖLQVVRµÄĪåėÄʬ¶ĪÄܹ»ŠĪ³Éµķ·ŪŃłĻĖĪ¬£¬ŌŚøƵ°°×ÖŹµÄŅģ³£¾Ū¼Æ¹ż³ĢÖŠ·¢»ÓÖŲŅŖ×÷ÓĆ[19]
“ĖĶā£¬¾«ŅŗĄ“Ō“µÄ²”¶¾øŠČ¾ŌöĒæ×Ó (SEVI)Äܹ»Ķعżµķ·ŪŃłĻĖĪ¬µÄŠĪ³ÉŌö¼ÓHIV²”¶¾µÄøŠČ¾Į¦[20]£¬ŅȵŗĖŲŌŚ“¢“ę¹ż³ĢÖŠ·¢Éśµķ·ŪŃł¾Ū¼Æ»įÓ°ĻģĘäŅ©Š§[21]”£ŌŚÕāŠ©µ°°×ÖŹÖŠ£¬Ķ¬Ńł·¢ĻÖÄܹ»ŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ¶ĢėÄʬ¶Ī£¬ÕāŠ©¶ĢėÄĪŖ½ųŅ»²½Ąķ½āµ°°×ÖŹ·¢ÉśŅģ³£¾Ū¼ÆµÄ·Ö×Ó»śÖĘĢį¹©ÖŲŅŖµÄĻßĖ÷£¬Table1øÅĄØĮĖÉĻŹöµķ·ŪŃł¶Ģėĵĥ“Ō“¼°Ęä°±»łĖįŠņĮŠ”£
£“ µķ·ŪŃł¶ĢėÄÖŠµÄ³£¼ū°±»łĖį
ÉĻŹö“Ó²»Ķ¬µ°°×ÖŹÖŠ¼ų¶Ø³öĄ“µÄµķ·ŪŃł¶ĢėÄ£¬ĖäČ»ŌŚ¾ßĢåµÄŠņĮŠÉĻČŌČ»ø÷²»ĻąĶ¬£¬µ«ŹĒĶعż·ÖĪöĖüĆĒŗ¬ÓŠµÄ°±»łĖį²Š»ł¼°ĘäŠŌÖŹ£¬²»ÄŃ·¢ĻÖĘäÖŠÓŠŅ»Š©³öĻÖʵĀŹ½Ļøߵݱ»łĖį²Š»ł£¬ÕāŠ©°±»łĖį²Š»łĖł¾ßÓŠµÄĢŲŹāŠŌÖŹ£¬ÓŠĶū³ÉĪŖ½āŹĶ²»Ķ¬µÄ¶ĢėÄČēŗĪŠĪ³É¾ßÓŠĻąĖĘĢŲÕ÷µÄµķ·ŪŃłĻĖĪ¬µÄ¹Ų¼üĻßĖ÷”£
4.1 ŹčĖ®ŠŌ°±»łĖį
ŹčĖ®×÷ÓĆ×÷ĪŖµ°°×ÖŹŗĶ¶ąėÄ×Ō·¢¾Ū¼ÆµÄŌŹ¼Ēż¶ÆĮ¦Ö®Ņ»£¬ŌŚµķ·ŪŃłĻĖĪ¬µÄŠĪ³É¹ż³ĢÖŠ·¢»ÓÖŲŅŖ×÷ÓĆ”£µ°°×ÖŹŠņĮŠÖŠµÄŹčĖ®ĒųŹĒĘä·¢Éśµķ·ŪŃł¾Ū¼ÆµÄÖ÷ŅŖĒųÓņ£¬ÕāŅŃŹĒ¹²Ź¶[22]£¬TableµÄÄŚČŻĻŌŹ¾£¬ÄæĒ°ŅŃ±ØµĄµÄ“Óø÷ÖÖĢģČ»µķ·ŪŃłµ°°×ÖŹÖŠ¼ų¶Ø³öĄ“µÄ¹Ų¼ü¶ĢėÄʬ¶Ī£¬¼øŗõ¶¼ŹĒŅŌ±ū°±Ėį (£Į)”¢ ēÓ°±Ėį(£Ö)”¢ĮĮ°±Ėį(£Ģ)”¢ŅģĮĮ°±Ėį(£É)”¢±½±ū°±Ėį(£Ę)”¢¼×Įņ°±Ėį(£Ķ)µČŹčĖ®ŠŌ°±»łĖįĪŖÖ÷£¬ĮķŅ»·½Ćę£¬ŌŚĪŅĆĒŅŌĶłŃŠ¾æÖŠ£¬ČĖ¹¤Éč¼ĘµÄŅ»Š©Äܹ»×Ō×é×°ŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄÄÉĆ׶ĢėIJÄĮĻ£¬Ņ²ŹĒÓÉ“óĮæĮ¬ŠųµÄŹčĖ®ŠŌ°±»łĖį²Š»łĖł×é³É[23,24]£¬ÓÉ“ĖČĻĪŖ£¬ŹčĖ®ŠŌ°±»łĖį£¬ÓČĘäŹĒĮ¬ŠųµÄŹčĖ®ŠŌ°±»łĖįʬ¶Ī£¬ŹĒµķ·ŪŃł¶ĢėĵÄŅ»øöÖŲŅŖĢŲÕ÷£¬ŹĒ·ńŗ¬ÓŠ×ć¹»µÄŹčĖ®ŠŌ°±»łĖį£¬ŹĒŌ¤²āŅ»øö¶ĢėÄŠņĮŠÄÜ·ńŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄÖŲŅŖÖø±ź£¬Ķ¬Ź±Ņ²ŹĒ²ūĆ÷ĢģČ»µ°°×ÖŹŅģ³£¾Ū¼Æ·Ö×Ó»śÖʵÄÖŲŅŖĻßĖ÷”£
4.2 øŹ°±Ėį(£Ē)
øŹ°±ĖįĖäČ»Ķس£±»¹éĄąĪŖĒ×Ė®ŠŌµÄ°±»łĖį£¬µ«ŹĒÓÉÓŚĘä²ąĮ“ĪŖ²»æɽāĄėµÄ-£Č£¬ĘäŠŌÖŹĘ䏵ŹĒ½éÓŚĒ×Ė®ŗĶŹčĖ®Ö®¼äµÄ£¬“ĖĶā£¬øŹ°±ĖįŅ²ŹĒ²ąĮ“×īŠ”ŗĶ×ī¾ßČįČĶŠŌµÄ°±»łĖį£¬ŌŚ¶ĢėÄŗĶµ°°×ÖŹ·¢Éś¹¹Ļó±ä»Æµ¼ÖĀ¾Ū¼ÆµÄ¹ż³ĢÖŠ£¬Äܹ»Ģį¹©ČįČĶŠŌĮ¬½Ó¶ų“Ł½ų¾Ū¼ÆµÄ·¢Éś£¬ŌŚ×ŌČ»½ēÖŠ£¬ŠĪ³ÉøßĒæ¶ČĻĖĪ¬µÄÖėĖæµ°°×£¬ŌŚĘäŗĖŠÄĒųÓņ¾Ķŗ¬ÓŠ“óĮæÖŲø“µÄøŹ°±ĖįŠņĮŠ[25]£¬ŌŚÉč¼Ę¹¦ÄܻƵÄ×Ō×é×°¶ĢėÄÄÉĆ×ĻĖĪ¬²ÄĮĻŹ±£¬øŹ°±ĖįŅ²³£±»ÓĆ×÷×Ō×éװʬ¶ĪŗĶ¹¦ÄÜʬ¶ĪÖ®¼äµÄĮ¬½ÓŌŖ¼ž[26]£¬¶ų·ÖĪöÄæĒ°ŅŃŃŠ¾æµÄµķ·ŪŃł¶ĢėÄ£¬²»ÄŃ·¢ĻÖøŹ°±ĖįŅ²“óĮæ³öĻÖŌŚÕāŠ©¶ĢėÄŠņĮŠÖŠ(Table1)£¬±ķĆ÷øŹ°±ĖįŌŚ¶ĢėÄ×Ō×é×°ŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ¹ż³ĢÖŠ·¢»ÓÖŲŅŖ×÷ÓĆ”£
4.3 ±½»·½į¹¹
ŌŚ³£¼ūµÄŹčĖ®ŠŌ°±»łĖįÖŠ£¬±½±ū°±ĖįŌŚ¶Ģėĵĵķ·ŪŃł¾Ū¼ÆÖŠ·¢»Ó×ÅÓČĘäÖŲŅŖµÄ×÷ÓĆ£¬ÕāŹĒŅņĪŖ³żĮĖ±¾ÉķŹĒŹčĖ®ŠŌ¼«ĒæµÄ°±»łĖįÖ®Ķā£¬±½±ū°±Ėį²ąĮ“ÖŠµÄ±½»·½į¹¹»¹ÄÜĢį¹© ¦Š-¦Š ¶Ń»ż×÷ÓĆ£¬ĪŖ¶ĢėĵľŪ¼ÆĢį¹©¶īĶāµÄĒż¶ÆĮ¦£¬ÓŠŃŠ¾æ±ķĆ÷£¬½«KLVFFAEÖŠµÄ±½±ū°±ĖįĢę»»ĪŖĶ¬ŃłŹĒŹčĖ®ŠŌ°±»łĖįµÄ±ū°±Ėįŗó£¬Ęä¾Ū¼ÆŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄÄÜĮ¦¼õČõ[27]£¬“ĖĶā£¬ĄŅ°±ĖįĖäČ»²»ŹĒµäŠĶµÄŹčĖ®ŠŌ°±»łĖį£¬ŌŚµķ·ŪŃłĻĖĪ¬µÄŠĪ³ÉÖŠŅ²·¢»ÓÖŲŅŖµÄ×÷ÓĆ£¬ĄżČēÓŠŃŠ¾æ±ķĆ÷£¬¦Į-Ķ»“„ŗĖµ°°×µŚ39Ī»µÄĄŅ°±Ėį£¬æÉŅņĪŖĘä²ąĮ“ÖŠ±½»·½į¹¹µÄ“ęŌŚ£¬ĪŖ¶ĢėĵľŪ¼ÆĢį¹©¦Š-¦Š¶Ń»ż×÷ÓĆ[28]”£
4.4 ¹Č°±õ£°·(£Ń)
ÓÕµ¼¶ĢėÄ·¢Éśµķ·ŪŃł¾Ū¼ÆµÄĮķŅ»øöÖŲŅŖµÄ°±»łĖįŹĒ¹Č°±õ£°·£¬ĖäČ»¹Č°±õ£°·Ķس£±»ČĻĪŖŹĒĒ×Ė®ŠŌ°±»łĖį£¬µ«ŹĒÄæĒ°·¢ĻֵĊķ¶ąµ°°×ÖŹ»ņ¶Ģėĵĵķ·ŪŃł¾Ū¼ÆŠŠĪŖ£¬¶¼Óė¹Č°±õ£°·ĆÜĒŠĻą¹Ų£¬ŌŚŅżĘšŗąĶ¢¶ŁĪčµø²”µÄHuntingtinµ°°×ÖŠ£¬ĘäN¶Ė¶ą¾Ū¹Č°±õ£°·ÖŲø“ŠņĮŠ±»ČĻĪŖŹĒ¾Ū¼Æ·¢ÉśµÄÖŲŅŖŌŖ¼ž[17]£¬ŌŚĄ“×Ō½ĶÄøSup35µ°°×µÄGNNQQNYµķ·ŪŃł¶ĢėÄÖŠ£¬Ņ²ÓŠ2øö¹Č°±õ£°·²ĪÓė¶ĢėÄ×Ō×é×°µÄ·¢Éś[29]”£ ŌŚ ¦Į-Ķ»“„ŗĖµ°°×ÖŠ£¬µ±µŚ50Ī»µÄ×é°±ĖįĶ»±äĪŖ¹Č°±õ£°·Ź±£¬øƵ°°×ÖŹ·¢Éś¾Ū¼ÆµÄÄÜĮ¦Ć÷ĻŌĢįøß[30-31]”£ÕāŠ©ŃŠ¾æ½į¹ūĢįŹ¾£¬¹Č°±õ£°·ŌŚµķ·ŪŃł¾Ū¼ÆµÄ¹ż³ĢÖŠ·¢»Ó×ÅÖŲŅŖ×÷ÓĆ£¬ÕāÓė¹Č°±õ£°·²ąĮ“Ä©¶ĖµÄõ£°·»łÖ®¼äæÉŠĪ³ÉĒā¼ü£¬ÓÕµ¼×Ō×é×°µÄ·¢ÉśÓŠ¹Ų[32].ĪŅĆĒ×ī½üµÄŃŠ¾æ·¢ĻÖ£¬ĖäČ»¹Č°±õ£°·ŹĒĒ×Ė®ŠŌ°±»łĖį£¬Ęä²ąĮ“ÉĻµÄ¶žŃĒ¼×»łŹĒ¾ßÓŠŅ»¶ØŹčĖ®ŠŌµÄ»łĶÅ£¬ ÕāÖÖ¾Ö²æµÄŹčĖ®ŠŌ£¬Ķ¬ŃłŌŚÓÕµ¼¶Ģėĵĵķ·ŪŃł¾Ū¼ÆÖŠ·¢»Ó×Ų»ČŻŗöŹÓµÄ×÷ÓĆ[33]”£
£µ ŃŠ¾æµķ·ŪŃł¶ĢėĵÄŅāŅåŗĶÓ¦ÓĆ
5.1 Ļą¹Ų¼²²”µÄŌ¤·ĄŗĶÖĪĮĘ
Õė¶ŌĄ“×ŌĢģČ»µ°°×ÖŹµÄµķ·ŪŃł¶ĢėÄÕ¹æŖŃŠ¾æ£¬²ūĆ÷ĖüĆĒŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ·Ö×Ó»śÖĘ£¬ÓŠÖśÓŚ·ÖĪöĘäĖłŌŚµÄĶźÕūµ°°×ÖŹŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄŌĄķ£¬“Ó¶ų×īÖÕ²ūĆ÷Ļą¹Ų¼²²”µÄ·¢²”»śÖĘ£¬ĪŖÕāŠ©¼²²”µÄŌ¤·Ą”¢Õļ¶ĻŗĶÖĪĮĘĢį¹©ĄķĀŪ»ł“”£¬ĮķŅ»·½Ćę£¬ŌŚÖ¤Źµµķ·ŪŃł¶ĢėĶŌÓŚĶźÕūµ°°×ÖŹŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄÖŲŅŖŠŌÖ®ŗó£¬Ņ²æÉÕė¶ŌÕāŠ©¶ĢėÄæŖÕ¹½ųŅ»²½µÄŃŠ¾æ£¬ŌŚĻą¹Ų¼²²”µÄŌ¤²āŗĶÖĪĮĘ·½Ćę·¢»Ó×÷ÓĆ”£
¼ų¶Ø³öĢģČ»µ°°×ÖŹŠĪ³Éµķ·ŪŃłĻĖĪ¬¹ż³ĢÖŠµÄ¹Ų¼ü¶ĢėÄŠņĮŠŗó£¬æÉŅŌ½ųŅ»²½¼ų¶Ø³ö×ī¹Ų¼üµÄ°±»łĖį²Š»ł£¬²¢Ķعż¶ŌøĆ²Š»łµÄŠŽŹĪĄ“µ÷æŲµķ·ŪŃłĻĖĪ¬µÄŠĪ³É”£ĄżČēGodinµČ[34] ±ØµĄ£¬IAPPµŚ21Ī»µÄĢģ¶¬°±Ėį¶Ōµķ·ŪŃłĻĖĪ¬µÄŠĪ³É¼°ĘäĻø°ū¶¾ŠŌ·¢»ÓÖŲŅŖµÄµ÷æŲ×÷ÓĆ,½«ĘäĢę»»ĪŖ±½±ū°±Ėįŗó,øƶąėÄÄܹ»ĘĘ»µĻø°ūĤ½į¹¹µÄĶźÕūŠŌ,ŅżĘš¦ĀĻø°ūµņĶö£¬ÕāŅ»·¢ĻÖĪŖŌ¤²āIIŠĶĢĒÄņ²”øß·ēĻÕ»łŅņĶ»±äĢį¹©ĄķĀŪÖøµ¼”£XiongµČ[12]·¢ĻÖ£¬½«µķ·ŪŃł¶ąėÄA¦Ā1-42µŚ32Ī»µÄŅģĮĮ°±ĖįĢę»»ĪŖ±ū°±Ėį,ĘäŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄÄÜĮ¦ĻŌÖų¼õČõ£¬ÕāŅ²ĪŖ½ńŗóÓĆ»łŅņ±ą¼µÄ·½·ØŌ¤·ĄŗĶÖĪĮĘ°¢×Čŗ£Ä¬Ö¢µÄ·¢ÉśĢį¹©ĮĖæÉÄÜ”£
ŌŚ·¢ĻÖµ°°×ÖŹÖŠµÄ¹Ų¼üµķ·ŪŃł¶ĢėÄŠņĮŠŗó£¬Ņ²ÄÜÕė¶ŌøĆŠņĮŠĄ“Éč¼ĘŗĶÉøŃ”ĻąÓ¦µÄŅÖÖĘ¼Į£¬“Ó¶ųŅÖÖĘĘäµķ·ŪŃłĻĖĪ¬µÄŠĪ³É£¬²¢ÓŠĶūæŖ·¢³öŅÖÖĘĻąÓ¦ĶźÕūµ°°×ÖŹŅģ³£¾Ū¼ÆµÄŅ©Īļ£¬ĄżČē£¬ŌŚMaoµČ[35],ŃŠ¾æÖŠ£¬Ņ»ĻµĮŠČĖ¹¤Éč¼ĘŗĻ³ÉµÄ¶Ģėı»ÓĆÓŚŅÖÖĘIAP11-20ʬ¶Īµķ·ŪŃł¾Ū¼ÆµÄ·¢Éś£¬ŌŚKatyalµČ[29]ŃŠ¾æÖŠ£¬ŗ£ŌåĢĒ±»ÓĆÓŚŅÖÖĘGNNQQNY·ŪŃł¾Ū¼ÆµÄ·¢Éś,ChenµČ[36] ×ī½üµÄŃŠ¾æ·¢ĻÖ£¬Ńõ»ÆŹÆÄ«Ļ©Äܹ»ŅÖÖʵķ·ŪŃł¶ĢėÄA¦Ā33-42µÄ¾Ū¼Æ£¬ÕāŠ©ŃŠ¾æ£¬ÓŠĶūĪŖ½«Ą“æŖ·¢Õė¶Ōµķ·ŪŃł²”±äµÄŠ”·Ö×ÓŅ©ĪļĢį¹©Ė¼Ā·”£
5.2 ÄÉĆ×ÉśĪļ²ÄĮĻµÄæŖ·¢
“ĖĶā£¬Ä£ÄāĢģČ»ŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ¶ĢėÄŠņĮŠ£¬Ä³Š©Äܹ»ŠĪ³ÉÄÉĆ×ĻĖĪ¬½į¹¹µÄ¶ĢėÄ£¬Ņ²±»Éč¼Ę×÷ĪŖŠĀŠĶÄÉĆ×ÉśĪļ²ÄĮĻ£¬ŌŚÄÉĆ×Ņ½Ń§”¢ÄÉĆ×¼¼ŹõµČĮģÓņ·¢»ÓĘä×÷ÓĆ”£ĄżČēTangµČ[37] Éč¼ĘĮĖŅ»ĻµĮŠŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ5ėÄ£¬Äܹ»æģĖŁŠĪ³ÉæÉ×¢ÉäµÄĖ®Äż½ŗ£¬ŌŚ×°ŌŲĻø°ūæŖÕ¹×éÖƹ¤³ĢÓ¦ÓĆ·½ĆęĻŌŹ¾³ö¾Ž“óµÄĒ±Į¦£¬ČĖ¹¤Éč¼ĘµÄKFFEAAAKKFFEæÉŠĪ³ÉµäŠĶµÄµķ·ŪŃłĻĖĪ¬£¬²¢×÷ĪŖÄ£°åĄ“ÖʱøŅ»Ī¬¹čÄÉĆ×Ļß[38]£¬ĪŅĆĒ×ī½üµÄŃŠ¾æ·¢ĻÖ£¬ŅŌĮ¬ŠųµÄŹčĖ®ŠŌ°±»łĖį²Š»łĪŖÖ÷ĢåÉč¼ĘµÄ¶ĢėÄA6KŅ²ÄÜŠĪ³ÉµäŠĶµÄµķ·ŪŃłĻĖĪ¬£¬²¢æÉ×÷ĪŖŹčĖ®Ņ©ĪļµÄŌŲĢ壬ĢįøßĘäŌŚĖ®ČÜŅŗÖŠµÄĪČ¶ØŠŌ[39]£¬ÕāŠ©ŃŠ¾æ±ķĆ÷£¬³żĮĖŌŚĻą¹Ų¼²²”ÖŠµÄĄķĀŪŅāŅ壬µķ·ŪŃł¶ĢėÄŅ²æÉÄܳÉĪŖŅ»ĄąŠĀŠĶµÄÉśĪļ²ÄĮĻ£¬æŖ·¢ÕāŠ©µķ·ŪŃł¶ĢėÄŌŚÉśĪļŅ½Ń§¹¤³ĢĮģÓņµÄĒ±ŌŚŹµÓĆ¼ŪÖµ£¬Ņ²ŹĒŅ»øöÖµµĆ¹Ų×¢µÄ·¢Õ¹·½Ļņ”£
£¶ ĪŹĢāÓėÕ¹Ķū
×ŪÉĻĖłŹö£¬ÄæĒ°ŅŃ¾“ÓŠķ¶ąĢģČ»µ°°×ÖŹĄļ·¢ĻÖĮĖÖŲŅŖµÄµķ·ŪŃł¶ĢėÄŠņĮŠ£¬²¢¶ŌĘä½ųŠŠĮĖÉīČėŃŠ¾æ£¬ÕāŠ©µķ·ŪŃł¶ĢėÄĶس£ŗ¬ÓŠ½Ļ¶ąµÄŹčĖ®ŠŌ°±»łĖį£¬ŌŚŹčĖ®×÷ÓĆ”¢¦Š-¦Š ¶Ń»ż×÷ÓĆŗĶĒā¼üµČ·Ö×Ó¼ä·Ē¹²¼Ū×÷ÓĆĮ¦µÄĒż¶ÆĻĀ£¬Ķعż¶ĢėĦĀÕŪµž¶ž¼¶½į¹¹Ļą»„¾Ū¼Æ£¬“Ó¶ųŠĪ³ÉµäŠĶµÄµķ·ŪŃłĻĖĪ¬£¬ŃŠ¾æÕāŠ©µķ·ŪŃł¶ĢėĵÄ×Ō×é×°»śÖĘ£¬ÓŠÖśÓŚ½ųŅ»²½²ūĆ÷ĘäĖłŌŚĢģČ»µ°°×ÖŹŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ·Ö×Ó»śÖĘ”£Č»¶ų£¬µķ·ŪŃł¶ĢėĵÄŃŠ¾æŅ²ÓŠĘä×ŌÉķµÄĪŹĢāŗĶ¾ÖĻŽŠŌ£¬ĄżČē£¬ĶźÕūµ°°×ÖŹČēŗĪ»łÓŚÕāŠ©¹Ų¼üµÄ¶ĢėÄŠņĮŠŠĪ³Éµķ·ŪŃłĻĖĪ¬£¬ÄæĒ°ČŌȻȱ·¦Ņ»øö³ä·ÖŗĻĄķµÄ½āŹĶ£¬“ĖĶā£¬µ°°×ÖŹµÄŅģ³£¾Ū¼ÆŅ²²»ĶźČ«ÓÉŠņĮŠ±¾Éķ¾ö¶Ø£¬ĢŲ¶ØĪ»µćµÄŠŽŹĪŅ²»įÓ°ĻģŅģ³£¾Ū¼ÆµÄ·¢Éś£¬ĄżČē£¬Ņ»Š©°±»łĖįĪ»µćµÄŅģ³£ŠŽŹĪ»įµ¼ÖĀ¾Ū¼ÆµÄ·¢Éś[40]£¬¶ųĮķŅ»Š©ŠŽŹĪ£¬ČēĢĒ»ł»ÆŠŽŹĪ£¬»į½µµĶ£Į¦ĀµÄ¾Ū¼Æ³Ģ¶Č£¬µ¼ÖĀ¶¾ŠŌøüĒæµÄ¹Ń¾ŪĢåµÄÉś³É[41]£¬Ņņ“Ė£¬Ī§ČĘŅŃÖŖµÄµķ·ŪŃł¶ĢėÄ£¬ČŌȻӊ“óĮæµÄÉŠĪ“Ē峞µÄ»śÖĘÖµµĆ½ųŅ»²½Ģ½Ė÷£¬ŌŚŅżĘšµķ·ŪŃł²”±äµÄµ°°×ÖŹÖŠ£¬ČŌȻӊŠķ¶ąĪ“ÖŖµÄµķ·ŪŃł¶ĢėÄŠčŅŖČ„·¢¾ņ£¬³żĮĖµķ·ŪŃł¶Ģėı¾ÉķµÄ°±»łĖįŠņĮŠ£¬µ°°×ÖŹµÄŠŽŹĪŅ²ŹĒÖµµĆ¹Ų×¢µÄÖŲµć£¬ ÕāŠ©¹¤×÷½«ĪŖ×īÖÕ²ūĆ÷ĢģČ»µ°°×ÖŹŠĪ³Éµķ·ŪŃłĻĖĪ¬µÄ·Ö×Ó»śÖʵģ¶Ø»ł“”£¬ĮķŅ»·½Ćę£¬µķ·ŪŃł¶ĢėÄŠĪ³ÉµÄÄÉĆ×ĻĖĪ¬£¬Ņ²æɱ»×÷ĪŖŅ»ĄąĢģČ»µÄÄÉĆ×ÉśĪļ²ÄĮĻ£¬ŌŚÖŚ¶ąĮģÓņ·¢»ÓĘäÓ¦ÓĆ¼ŪÖµ”£
ĆāŌšÉłĆ÷£ŗ±¾ĪÄĪŖŠŠŅµ½»Į÷ѧĻ°£¬°ęČعéŌ×÷Õß¼°ŌŌÓÖ¾ĖłÓŠ£¬ČēÓŠĒÖČØ£¬æÉĮŖĻµÉ¾³ż”£ĪÄÕĀ±ź×¢ÓŠ×÷Õß¼°ĪÄÕĀ³ö“¦£¬ČēŠčŌĶĮŌĪļ°²Īæ¼ĪÄĻ×£¬æÉŌĶĮŌŌÓÖ¾
