肽水凝胶的本质在于肽分子通过非共价键自组装形成纳米网络结构,从而在空隙中容纳水分子。在非共价键相互作用中,芳香环-芳香环相互作用、疏水相互作用和氢键对肽的自组装有积极影响,而静电相互作用则起着更为复杂的作用,包括静电排斥和静电吸引。尽管肽序列本身在自组装中起着至关重要的作用,但凝胶化的触发条件也发挥着重要作用。动力学和热力学因素能够调节肽的组装效率和形态。肽分子趋向于结晶态(热力学最低能态)还是凝胶态(动力学亚稳态),也受到热力学和动力学的极大影响。
肽分子与溶剂分子之间的氢键可能会因温度和超声波而减弱,溶液的离子强度和pH值会抑制静电相互作用。另一方面,提高温度可能会增强疏水效应。与溶剂可及疏水性(SAH)较小的肽相比,具有较大SAH值的肽会形成溶剂可及疏水性(SAH)较小的纤维。具有较大SAH值的肽在较低温度下会更快地形成纤维网络。离子浓度通过影响静电相互作用在肽的组装中起作用。例如,在高浓度的氯化钠(≥0.5m)条件下,肽分子之间的静电排斥力可能会减弱,导致它们之间的堆积角减小。因此,初始的纳米纤维结构会转变为纳米管形态。溶剂的切换会影响肽不同部分的溶解性,从而影响其组装。Aβ42蛋白质从有机相迁移到水相会导致其易于聚集的构象发生变化。在此过程中,亲水侧链的脱水、极性骨架的脱水以及疏水侧链的暴露显著增加了Aβ42蛋白的溶剂化自由能。此外,浓度与水凝胶的弹性不一定呈正相关,也不一定与低浓度下肽的弹性相关。浓度可能比高浓度时高出一个数量级。光介导的构象变化可导致利用液-液相分离(LLPS)的肽组装的进展或终止。与具有高成核势垒的传统单步自组装机制相比,LLPS介导的多步自组装机制通过增加LLPS形成的液滴中的溶质浓度显著降低了成核势垒,为精确自组装提供了一种有效策略。相分离后,肽形成高浓度液滴,组装从液滴内部向外自组装并生长。在连续组装过程中,液滴中的肽分子被稀释并溶解在溶液中,然后组装体可以从溶液中分离出来。谢明键团队为肽设计了一个两步成核计算模型,并提出了类似的假设。与实验结果相比,他们的计算模型正确预测了在Aβ(16-22)凝胶过程的初始阶段,水溶液中乙腈含量的增加会导致形成更少、更大的液滴,从而加速组装。不同的溶剂组成可以在成核过程之前调节粒子相的性质、大小和数量。
肽凝胶的形成、结构和性能会受到分子层面多种调控方法组合的影响。某些肽分子在pH值和温度的双重作用下会表现出独特的自组装行为。肽Py-VFFAKK(P1)将富电子发色团芘与肽片段相结合。在碱性条件下,P1形成纳米颗粒,而在中性和酸性条件下则形成纳米纤维。在pH值为7时,温度触发P1形成更稳定的螺旋结构,并诱导形成水凝胶。另一方面,在pH值为3的条件下,温度触发只能使P1在溶液中自组装,其机械强度比pH值为7时低10个数量级。在纯丝素蛋白(HS)水凝胶中,丝素蛋白链形成链内和链间氢键,倾向于形成晶体并形成以无孔结构为主的结构。添加核黄素和增加离子浓度可显著增强链内氢键结合,并提高HS水凝胶的孔隙率。离子的存在还能通过吸引水分子到水凝胶表面来增强保水能力。在食品工业中,pH值和离子浓度的结合会影响离子键、氢键和疏水相互作用,从而实现蛋白质在气液界面的稳定组装和吸附。
不同的凝胶化方法会影响其与其他抗癌疗法联合使用时所采取的策略。例如,某些生物分子无法承受高温或高离子强度,而许多小分子对pH值或光照敏感。水凝胶的硬度和凝胶化时间也是决定其能否在癌症治疗中临床应用的重要参数。在加热冷却过程中,加热后冷却至室温通常需要1至2小时,而有些水凝胶需要老化2天。有一种环肽,HOOC-Cyclo(L-Lys-L-Glu)-Fmoc(6),加热溶液后仅需1分钟即可凝胶化。在上述加热冷却水凝胶的硬度信息中,G'的值约为10³,G"的值约为10²。从二甲基亚砜(DMSO)切换到水的溶剂交换法仅需5分钟,但从水切换到丙酮则需要搅拌一夜。离子强度机制形成水凝胶需要2至3小时,G'的值在500至10000之间,G"的值在0.1至100之间。在没有任何辅助手段的情况下,通过简单涡旋实现凝胶化的时间,大部分在30分钟到12小时之间,但E3C16E6除外,它需要老化24小时。对于这些水凝胶,G'的范围是100到1000,G''为10到100。仅需15到20分钟的超声处理,RADA-KLA肽就能在30分钟内自组装成G'(5000)和G''(500)的水凝胶。基于相同的超声处理方法,SH水凝胶不仅需要3天的老化时间,而且其G'和G''的值比RADA-KLA水凝胶低10倍。光交联法是触发水凝胶形成最快的方法;一条肽链或一个肽环只需几秒钟就能形成水凝胶。通过pH反应实现凝胶化的时间通常不超过24小时,从5分钟到3小时不等。储存模量在9到1000之间,G''在3到100之间。由液-液相分离(LLPS)触发的肽水凝胶至少需要10分钟才能凝胶化。LLPS肽水凝胶中最软的能达到G'=60和G''=30,而最硬的能达到G'(2400)和G''(700)。线性肽水凝胶的G'和G''值可通过额外添加诸如TPPS、阿霉素(Dox)和羟基喜树碱(HCPT)等化疗药物来提高,当这些药物添加到纯水凝胶中时,可使这些值提高10倍,但环肽水凝胶比线性肽水凝胶更硬。环肽水凝胶的G'和G''最小值分别为200和50,最大值分别为104和103。环肽水凝胶的凝胶化时间从即刻到数小时不等。然而,一些肽水凝胶仍需要数天甚至一周的时间来老化(表1)。肽水凝胶的凝胶化时间从即刻到数小时不等,但一些环肽水凝胶仍需要数天甚至数周的时间来老化。
1 加热 - 冷却过程
较高的温度能够通过让更多的粒子自由移动和相互作用来加速自组装,从而形成新的连接和结构。随着溶液冷却,新形成的结构会巩固并固化,形成类似凝胶的网络。冷却步骤使肽网络能够固定住溶液,从而形成稳定的凝胶。然而,如果温度超过特定阈值,所形成的结构可能会崩塌,导致凝胶重新变成溶液。
通常情况下,许多肽在中性环境(pH值7.4)中加热冷却后能够自组装。李等人将具有广泛免疫系统活性的促吞噬素(TKPR)与疫苗佐剂Nap-GDFDFDY相结合,创造了一种新型自组装分子Nap-GDFDFDYTKPR(化合物3),该分子在pH值为7.4的磷酸盐缓冲盐水(PBS)溶液中加热冷却后能够形成水凝胶。化合物3能够与巨噬细胞表面的NRP-1受体更牢固地结合,促进细胞摄取。尽管促吞噬素和Nap-GDFDFDY都能诱导抗原呈递细胞的吞噬作用,但化合物3的效果更为显著。与TKPR相比,化合物3能产生两倍的TNF-α、IFN-γ、IL-6和IL-12,并且能更有效地激活CD8+T细胞3.18倍。同时,它能通过多种给药方式(如尾静脉注射、皮下注射和腹腔注射)以相当的方式抑制肿瘤,如图1a所示。在小鼠乳腺癌肿瘤模型(4T1)中,与PBS组相比,用化合物3凝胶处理的小鼠最终肿瘤体积约为38.1%。FKFEYYSV(1-YSV)溶液在70°C下加热15分钟,然后冷却至25°C,于pH值为7.4时形成可注射水凝胶,其抗肿瘤活性比游离YSV提高了两倍,解决了YSV需要毫摩尔浓度才能发挥细胞活性的问题。静电相互作用对于1-YSV的自组装是必需的。正负氨基酸相互作用(K和E)促进了FKFE部分的自组装。1-YSV与10-羟基喜树碱(HCPT)的共组装改善了水凝胶的粘弹性,使储能模量值提高了10倍,并通过HCPT从扩散内吞作用转变为微胞饮作用穿透细胞(图1b)。1-YSV/HCPT水凝胶的肿瘤抑制效果是6.0毫克/克HCPT水溶液的两倍。
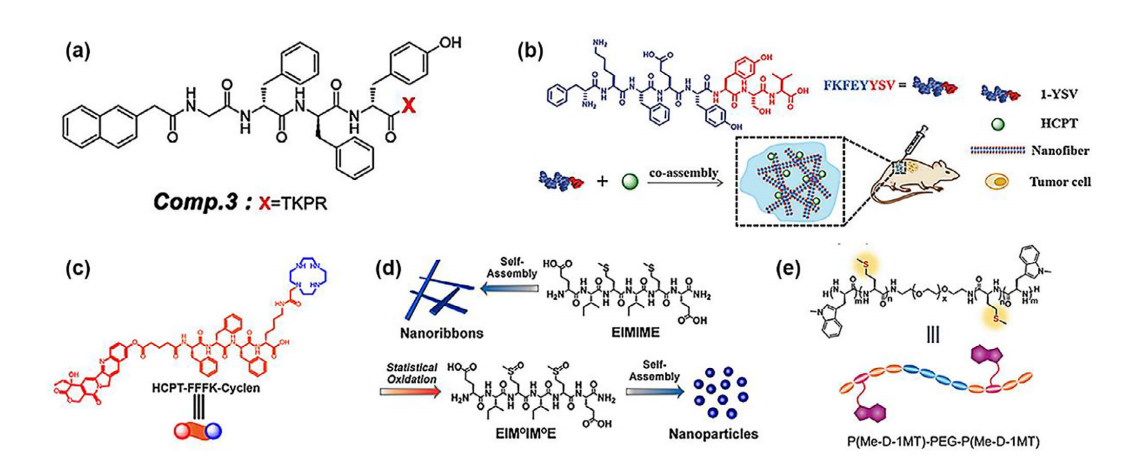
a)自组装肽Nap-GDFDFDYTKPR(化合物3)的结构。b)用于癌症治疗的1-YSV和10-羟基喜树碱(HCPT)共组装示意图。c)HCPT-FFFK-cyclen的化学结构。d)肽EIMIME及其氧化对应物的化学结构以及它们分别自组装成纳米带或纳米粒子的过程。e)P(Me-D-1MT)-PEG-P(Me-D-1MT)的结构及由活性氧触发的聚合物疏水性转变。
喜树碱(CPT)或羟基喜树碱(HCPT)在细胞核内的活性会抑制DNA拓扑异构酶I,从而阻止DNA链重新连接,导致DNA链断裂,最终导致细胞死亡。然而,由于水溶性低和全身毒性,它们的临床应用受到了限制。HCPT-FFFK-环己二胺在pH值为7.4时加热和冷却可形成水凝胶。此外,三苯丙氨酸(FFF)为水凝胶单元提供自组装能力,而HCPT作为封端基团。大环胺环己二胺具有两个优点:它可逆转细胞内ATP耗竭,从而减少ATP依赖性药物外排,并且由于其正电荷仅存在于癌细胞的酸性环境中,所以能特异性靶向癌细胞的细胞核。尽管与单独使用环己二胺相比,使用环己二胺与HCPT结合能改善ATP耗竭,但在体内抗癌试验中,环己二胺或FFFK-环己二胺与游离HCPT几乎没有抗肿瘤作用。只有这种自组装纤维结构才有利于HCPT-FFFK-cyclen进入细胞,并且在癌细胞酸性环境的激活下,其末端的cyclen具有靶向细胞核的能力,从而增加了HCPT在细胞核内的聚集和药效(图1c)。与PBS组相比,在人非小细胞肺癌(A549)的肿瘤模型中,HCPT-FFFK-cyclen组的肿瘤抑制率高达80%。
肽的序列对于维持自组装结构尤为重要。在bola型两亲性六肽的疏水部分添加一个或两个甲硫氨酸残基,会形成三种不同的结构:EIMIIE、EIIIME和EIMIME。这些肽分别形成超螺旋、扭曲的纳米带和纳米带。活性氧(ROS)会将甲硫氨酸肽中的硫醚转化为亚砜基团,但只有氧化后的EIMIME能够生成均匀的纳米颗粒。将光敏剂血卟啉e6(Ce6)和喜树碱(CPT)与EIMIME连接,然后共组装成水凝胶DPEIM。要获得水凝胶,需将新鲜溶液在pH值为7.4和80°C的条件下加热30分钟,然后立即冷却至室温并老化两天。从溶酶体逃逸后,DPEIM可通过激光照射用于同时进行光动力和化疗研究。喜树碱会通过活性氧与谷胱甘肽(GSH)响应性二硫键的相互作用而释放,该二硫键存在于肽与喜树碱之间。活性氧的释放还会将细胞外的纳米纤维DPEIM转化为能够深入肿瘤内部引发化疗的纳米颗粒。仅含Ce6肽的PEIM在激光照射后抗肿瘤能力相比未照射时显著提高,从50%提升至80%,加入CPT肽后,抗肿瘤能力进一步提升至约100%(图1d)。
此外,加热冷却法可用于构建基于肽的溶胶-凝胶转变材料。例如,于等人设计了一种含聚(L-甲硫氨酸)(PMe)的肽,作为连接大分子引发剂(胺封端的聚乙二醇)和右旋-1-甲基色氨酸(D-1MT)的多肽块,以及一种吲哚胺2,3-双加氧酶1(IDO-1)通路抑制剂。由于PMe的亚磺酸醚基团可被H2O2氧化成亚砜/砜,该水凝胶具有药物释放和H2O2清除性能。该材料表现出常规的热响应性溶胶-凝胶相变,且随着浓度的增加相变温度降低。他们选择了8.0wt.%的共聚物溶液,其具有合适的溶胶-凝胶相变温度(约22°C),并在PBS(pH7.4)中自组装成纤维。有趣的是,在低浓度(0.2毫克/毫升)时,共聚物呈球形。当将αPD-L1装载到凝胶中时,它能引发有效的T细胞介导的免疫反应,并对肿瘤细胞产生显著的杀伤效果。值得注意的是,共聚物的纯纤维比游离D-1MT治疗具有更强的CD8T细胞浸润(图1e)。αPD-L1负载的P(Me-D-1MT)-PEG-P(Me-D-1MT)组小鼠的存活率与PBS组相比显著提高,超过10天。这种负载有αPD-L1的肽水凝胶对黑色素瘤的抑制率为60%。当使用1,4-丁二醇二缩水甘油醚(BDDE)将肽与透明质酸(HA)交联时,溶胶-凝胶转变温度会发生变化。将BDDE加入肽溶液(pH7.4)并在50°C下加热3小时后,这些原材料会转化为水凝胶。所选的抗癌生物活性肽(ACBP;专利号ZL961222236.0)在体外和体内均能降低人胃癌干细胞(GCSCs)的增殖,诱导其凋亡并降低其致瘤性。具有独特结构的肽基水凝胶对胶原酶-I具有高度抗降解性,在24天内重量损失为40±5%。他们将水凝胶递送至肿瘤组织附近。他们发现,这种肽基水凝胶能够选择性地抑制胃癌细胞的生长,通过降低癌干细胞(CSC)的增殖,同时提高成纤维细胞的黏附和生长比例。这种肽基水凝胶对MKN-45肿瘤的抑制率为40%。
大多数自组装过程是在固定的pH值下进行的,但脂质在较宽的pH范围内发生修饰,并通过与肽类似物的不同末端相互作用构建出多种结构。在这项工作中,讨论了将肽整合到具有延长疏水域的脂质中,消除了静电相互作用的影响,并且不受肽结合过程中pH值和序列剧烈变化的影响。将溶液在80°C下加热30分钟并冷却至室温后,纳米纤维可在较宽的pH值范围内(5.0、7.4和9.0)自组装,如表1所示。注意到N端连接的E3C16-GG-C12-AAGG肽形成了具有反平行β-折叠氢键的扭曲纳米带。相反,C端接枝的纳米带表现出平行β-折叠氢键,并且被发现是扁平结构。
2 溶剂切换法
溶剂切换法是一种促进聚合物及其他分子自组装的常用技术。第一种溶液确保肽完全溶解,而第二种溶液(一种选择性溶剂)则使分子中的某一成分变得不溶。所选溶剂促使分子中不溶的部分聚集,从而形成各种自组装结构。采用“溶剂切换法”制备了Fmoc-FF水凝胶,即Fmoc-FF肽从Fmoc-FF-DMSO溶液中稀释至双蒸水,在室温下形成凝胶。使用TWEEN60/SPAN60(肽浓度为5×10−3wt.%,直径约130纳米,zeta电位约为-20.0/-25.0mV)制备的纳米凝胶(NGs)能够特异性靶向Caveolin1高表达的MDA-MB-231细胞。Caveolin1作为细胞质支架,促进膜内陷,有助于形成小窝,使纳米凝胶能够通过小窝介导的内吞作用进入细胞(图2a)。纳米凝胶(NGs)在24小时后可将细胞活力降低至40%以内。
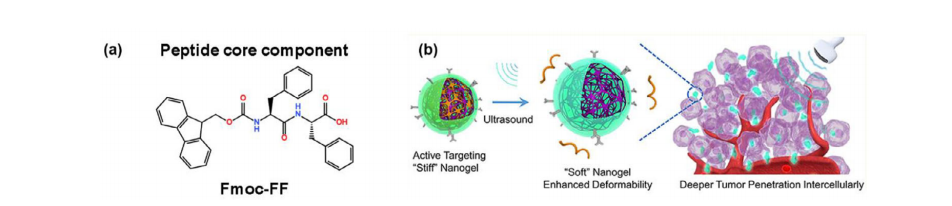
a)核心肽的化学结构。b)在外部超声作用下,GenPLPFT可通过脉冲触发的普朗尼克F127脱落转变为更软更大且具有更强变形能力的纳米凝胶,在肿瘤部位进行超声处理后,GenPLPFT/EPI会转变为更软更大且具有更强变形能力的纳米凝胶,从而实现对肿瘤的深层渗透。
“溶剂切换法”不仅可以通过用水稀释有机溶液来形成水凝胶,还可以通过用有机溶液稀释水来实现。为了触发由不良溶剂体系引发的自组装过程,向双蒸水溶液中加入约七倍体积比的丙酮,以帮助聚赖氨酸、经细胞间黏附分子-1(ICAM-1)抗体修饰的聚赖氨酸以及泊洛沙姆F127形成水凝胶。使用京尼平来稳定纳米结构,京尼平能够交联聚赖氨酸侧链上的氨基,然后将化疗药物(表柔比星)负载到纳米凝胶中(设计为GenPLPFT/EPI)。传统的三阴性乳腺癌(TNBC)治疗手段难以穿透深部肿瘤,难以治愈患者。在GenPLPFT/EPI中,ICAM-1是对抗TNBC的有力靶点,同时,京尼平交联会在肿瘤组织的细胞外酸性环境(即pH值6.0)下引起膨胀,京尼平交联的电荷反转使水凝胶能更快地从溶酶体中逃逸。GenPLPFT/EPI能够将表柔比星直接且特异性地递送至三阴性乳腺癌(TNBC)的深部肿瘤,并降低表柔比星引发急性心肌梗死的风险。同时,超声处理会降低肿瘤组织的间质压力,使水凝胶变软,通过从纳米凝胶中释放无害的F127来增强肿瘤渗透性,如图2b所示。在超声辅助下使用GenPLPFT/EPI进行的体内小鼠实验几乎完全治愈了肿瘤。其治疗效果是表柔比星的30倍。
3 电解质强度
电解质强度的增加通常会降低疏水部分在水中的溶解度,并抑制带电基团之间的静电排斥作用,使肽链彼此靠得更近,促进疏水段的聚集。根据其浓度(电解质强度),金属盐可以稳定或破坏肽结构。特定离子,如钙离子,能够特别与肽链的主链或侧链结合,稳定所构建的结构。二苯丙氨酸(FF)序列与叶酸(FA)的结合使得目标分子能够自组装。EE序列中的众多羧酸根以及作为碱性磷酸酶(ALP)底物的酪氨酸磷酸酶(Yp),分别使肽能够对钙离子和碱性磷酸酶作出响应。因此,在pH值为7.4的条件下,肽混合物与ALP和钙离子(1:1比例)反应会生成肽水凝胶。
肽FA-(Nal)FFKYpGEE被设计用于在正常细胞表面生成纳米纤维,以保护它们免受诸如顺铂之类的化疗药物的侵害,同时还能形成用于细胞培养的三维水凝胶(图3a)。单独使用顺铂(15微米)可抑制50%的细胞,但当培养细胞与肽水凝胶和顺铂一起使用时,细胞存活率可提高到70%。另一方面,Xu及其同事首次利用碱性磷酸酶(ALP),这种酶在癌细胞表面会过度释放和过度表达。因此,他们设计了一种肽D-Phe-D-Phe-D-Tyr(ffy),其中的Tyr被磷酸化。在37°C和pH7.4的条件下与癌细胞孵育2小时后,癌细胞表面和分泌的磷酸酶会使肽中的Tyr去磷酸化,导致癌细胞表面形成纳米纤维。水凝胶/纳米网显著将细胞与环境隔离开来,导致癌细胞发生依赖于半胱天冬酶的死亡(图3b)。肽水凝胶可将HeLa细胞、多药耐药性子宫肉瘤细胞系和Ect1/E6E7细胞的存活率分别降低至10%、30%和60%。
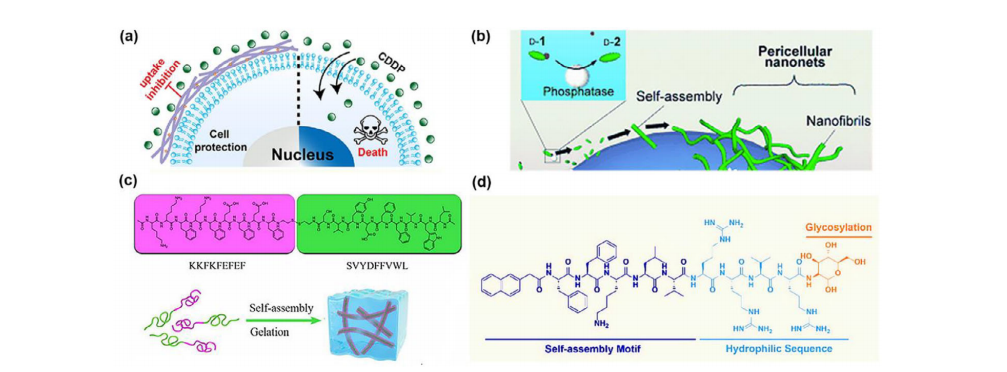
a)肽水凝胶能够保护正常细胞免受顺铂的损害。b)酶催化形成细胞周水凝胶/纳米网以诱导细胞死亡。c)KKEF-TRP2肽的氨基酸序列及自组装示意图。d)设计的糖肽(PG)的分子结构。
大多数肽可以通过使用氯化钠或磷酸盐缓冲液调节离子强度来触发形成水凝胶。此外,还可以通过使用氯化钠或磷酸盐缓冲盐水(PBS)来调整水凝胶的离子强度。KKEF-TRP2在暴露于氯化钠或PBS后,在浓度为20毫克/毫升时自组装成水凝胶。水不溶性表位SVYDFFVWL(TRP2)已通过二硫键连接到亲水部分和自组装肽KKFKFEFEF上,形成KKEF-TRP2。TRP2是B16黑色素瘤细胞上由人类HLA-A2和小鼠主要组织相容性复合体(MHC)I类分子H-2Kb限制的表位。与商业AddaVax佐剂肽疫苗相比,皮下接种KKEF-TRP2纳米纤维使肿瘤内活化的CD8+T细胞浸润增加了1.37倍。KKEF-TRP2治疗后,肿瘤重量仅为PBS组的三分之一,治疗效果比AddaVax佐剂肽疫苗好10%(图3c)。
为了促进肿瘤的快速生长,大量癌细胞会过度表达葡萄糖转运蛋白1(GLUT1),这是基础葡萄糖摄取所必需的。吴等人通过依次连接自组装基序(Nap-FFKLV)与亲水序列(RRVR)和糖基化(甘氨胺)来合成一种糖肽,并将等体积的肽溶液与2×PBS缓冲液(pH=7.4,37°C)混合,使糖肽自组装成纳米纤维,然后形成水凝胶。甘氨胺与GLUT1的相互作用使糖肽能够通过内吞作用进入癌细胞,随后糖肽在溶酶体中自组装,促进GLUT1的破坏。GLUT1回收的中断对需要高能量的癌细胞来说是毁灭性的。在溶酶体中自组装的糖肽(PG)会拉伸溶酶体,导致溶酶体功能障碍和癌细胞死亡。PG可使GLUT1表达降低60%,并产生比对照组多20.2倍的细胞内活性氧。治疗后,肿瘤体积仅为原来的二分之一;治疗效果接近PBS组,表明其治疗效果与PBS组相当,提示其治疗效果与紫杉醇相似(图3d)。
离子强度应根据肽的特性进行调整。例如,某些肽只需少量氯化钠溶液就能形成水凝胶。将Toll样受体激动剂(TLR7/8a,咪喹莫特)与SmacN7肽(AVPIAQK)连接,然后连接自组装肽KEF9(KKFKFEFEF)。由于KEF9之间的氢键、疏水和静电相互作用,滴加少量氯化钠溶液即可形成Smac-TLR7/8肽纳米纤维。Smac肽可直接与凋亡抑制蛋白(IAPs)结合,降低其抑制细胞凋亡的作用,从而选择性地提高癌细胞对辐射的敏感性。此外,TLR7/8激动剂用于诱导巨噬细胞极化为M1表型,从而重建免疫抑制性肿瘤微环境(ITM)。经Smac-TLR7/8和辐射处理后,不仅巨噬细胞的吞噬能力增强,而且极化的M1型巨噬细胞还可能释放肿瘤坏死因子、促炎细胞因子,并产生氧化分子,从而提高癌细胞对辐射的敏感性。Smac-TLR7/8联合放疗的肿瘤抑制率为67.7%,几乎是αPD-1联合放疗(36.2%)的两倍。
有趣的是,降低乙酸盐缓冲液的含量也能促进肽自组装成纳米纤维。例如,Coil29纳米纤维在没有佐剂的情况下也能引发体液和细胞免疫反应,而且在Coi29的N端连接的B细胞和T细胞表位不会破坏其正常的纳米纤维结构。为了打破a/d和c/f位置残基之间的疏水相互作用,从而调节纤维状肽生物材料的长度,同时稳定纳米纤维的上下表面,创建了Coil29Capa/d(将A、L改为S、T)和Coil29Capc/f(将I、A改为T、S)。将肽溶液在乙酸盐缓冲液(pH4.0)中稀释四倍后,在室温下肽会自组装成纤维。可能较短的纳米纤维更易于从内体中释放出来,进入细胞质,从而到达交叉呈递途径。较短的纳米纤维能使SIINFEKL的呈递增加1.5倍,并使针对SIINFEKL表位的CD8+T细胞数量翻倍。
4 肽浓度
各种肽类似物的结构特征具有天然形成自组装的能力,从而无需混合任何支撑材料即可形成水凝胶。氢键、范德华力、静电相互作用和疏水作用等非共价相互作用均有助于肽的自组装。
这些肽水凝胶可用作疫苗来刺激免疫反应,并具有其他生物医学功能。该水凝胶基于自组装肽KWKAKAKAKWK(K肽)或EWEAEAEAEWE(E肽),其表位通过化学方式与三个甘氨酸(GGG)残基相连。研究人员发现,与表位相连的K或E肽在水溶液中可自组装成纳米颗粒。此外,如果两种成分中有一种不溶于水,它们就会自组装成纳米纤维,否则就会形成纳米颗粒。共组装肽水凝胶疫苗能够特异性地靶向肿瘤细胞,将表位携带入内体或溶酶体,并通过非炎症途径增强T细胞反应,这与传统佐剂不同。共组装肽疫苗在树突状细胞中触发了MyD88依赖的NF-κB信号通路,提高了肽抗原的成熟和交叉呈递效率。治疗后,凝胶疫苗的最终肿瘤体积小于表位疫苗组的50%,仅为铝佐剂表位疫苗组的80%。
此外,利用肽水凝胶进行免疫治疗可以通过直接呈递表位或改变肿瘤微环境来实现。DBT-2FFGYSAYPDSVPMMS(DBT-2FFGYSA)是由两个FFGYSA肽连接在环境敏感荧光团4,7-二(噻吩-2-基)-2,1,3-苯并噻二唑(DBT)的两侧构成。YSAYPDSVPMMS(YSA)能特异性结合在许多癌症中过度表达且在癌症进展中起重要作用的跨膜受体酪氨酸激酶Eph受体A2(EphA2)。令人惊讶的是,由于存在两个芳香族苯丙氨酸(FF),FFG对于杀死EphA2受体过度表达的癌细胞至关重要,同时也有助于改善肽的自组装。当DBT-2FFGYSA与EphA2受体相互作用时,它为DBT创造了一个相对疏水的环境,从而显著增强了荧光并减少了分子内电子转移(ICT)。它可以在实时和特异性地在EphA2受体区域进行成像。DBT-2FFGYSA诱导了癌细胞中EphA2受体的聚集。它不仅通过半胱天冬酶依赖性途径(MAPK)和半胱天冬酶非依赖性途径(AIF和EndoG)激活癌细胞凋亡,还能诱导内质网应激和钙网蛋白表面暴露。此外,如图4a所示,DBT-2FFGYSA疗法成功地将冷肿瘤微环境转化为热肿瘤。治疗3周后,DBT-2FFGYSA自组装组的肿瘤体积降至生理盐水组的1/4。

a)DBT-2FFGYSA将免疫冷肿瘤转化为免疫热肿瘤的拟议机制示意图。b)由谷胱甘肽还原诱导的内化肽E3C16E6和ER-EVMSeO的原位自分类,以形成同时靶向内质网和高尔基体的肽组装体,促进细胞器功能障碍和随后的细胞死亡。c)用于光动力治疗的LND-K和TPPS4自组装成可注射制剂。d)纳米凝胶被用作可激活和程序性可调节的中性粒细胞模拟物,用于癌症的磁热酶联疗法(METT)。该纳米凝胶表现出优异的物理化学稳定性和生物安全性。可以在癌细胞中实现磁激活和程序性上调活性氧。这种方法成功地展示了单线态氧介导的细胞/肿瘤增殖抑制。
在另一个例子中,水凝胶(HGs)是通过将拉帕尼特纳米粘土(NCs)、肽聚树状分子(PPDs)和表面活性剂在去离子水中涡旋混合而制备的。肽聚树状分子是通过合成具有多价精氨酸残基的肽树状分子,并利用硫辛酸段的开环聚合而制得的。PPDs可通过氢键和静电相互作用与NCs相互作用,使NC的─OH离子暴露在自组装纳米纤维的外部。动态的─OH离子使HGs能够中和肿瘤产生的酸性代谢物,从而逆转肿瘤微环境,并产生特异性的抗肿瘤效果,以及良好的生物相容性。在体内和体外实验中,水凝胶能够诱导细胞凋亡。纯HGs的肿瘤抑制效果与低剂量药物治疗(1.00/0.10毫克/千克)相当,这种药物治疗涉及联合使用TRAIL(肿瘤坏死因子相关凋亡诱导配体)和阿霉素(一种小分子化疗药物)。与单纯低浓度和高浓度(2.50/0.25毫克/千克)的双药疗法相比,结合水凝胶的双药疗法对肿瘤的抑制率分别提高了3倍和2倍。
此外,纳米纤维能够靶向特定的细胞器以发挥抗肿瘤功能。肽溶液在室温、大气条件下配制,pH值为7.4。谷胱甘肽(GSH)会将纳米颗粒中的反自组装结构释放为两条肽链,例如二硫键,该二硫键连接着E3C16E6中的E6结构域和位于ER-EVMSeO(EVMSeO:Ts-EVMSeO=9:1)疏水区内的亲水硒氧化物基团。随后,E3C16SH自组装成扭曲的纳米纤维,而肽EVMSe则与Ts-EVMSe形成扁平的纳米带。由于E3C16SH中的巯基被GSH还原,能够与高尔基体上的富含半胱氨酸的蛋白质结合,而Ts-EVMSeO则利用Ts(N端Ts单元)作为靶向基团靶向内质网(ER),这种自分类组装赋予它们靶向内质网和高尔基体的能力,从而导致细胞器功能障碍并引发癌细胞死亡(图4b)。混合物E3C16E6+ER-EVMSeO能够将人宫颈癌细胞(HeLa)和人非小细胞肺癌细胞系(A549)的存活率降低至约30%。[46]此外,吴等人合成了LND肽(FFRFK-RGD-K)共轭物(LND-K),其中洛尼达明部分(LND)作为封端基团促进自组装,FFRFK结构域靶向线粒体。RGD三肽通过靶向过表达的αvβ3整合素受体进入癌细胞,并平衡肽的两亲性。在C末端,一个阳离子赖氨酸残基不仅在LND-K自身降低ATP水平、诱导细胞内活性氧(ROS)生成以及消融线粒体功能方面发挥关键作用,而且还能通过静电相互作用与四苯基卟啉-4,4,4-四磺酸(TPPS4)形成稳定的共组装体。TPPS4是一种阴离子光敏剂,可将凝胶化时间从30分钟缩短至1分钟。LND-K能够在pH7.4的水溶液中自组装成超分子水凝胶。在自组装过程中,疏水环境(例如π-π堆积)使得LND-K/TPPS4产生ROS的效率高于游离的TPPS4。LND-K肽在酸性环境中的电荷反转过程使LND-K/TPPS-4水凝胶能够靶向癌细胞并缓慢释放TPPS-4。LND-K能够通过有效抑制线粒体功能增强对癌细胞的细胞毒性,并破坏线粒体活性,最终使癌细胞对光动力TPPS4治疗更敏感(图4c)。在体外和体内实验中,LND-K/TPPS4的肿瘤治疗效果均超过单独使用TPPS4的两倍以上。
在另一项研究中,张等人制备了一种可注射且磁性引导的纳米凝胶MNP-CPO@Nanogels,这种纳米凝胶能够更有效地在肿瘤中积聚,同时对健康组织安全无害。难以溶解的氯过氧化物酶(CPO)和掺杂锌的Fe3O4磁性纳米颗粒(MNPs)通过Fmoc-Tyr-OH共同包封。Fmoc-Tyr(H2PO3)-OH兼具亲水和疏水区域,但H2PO3的存在破坏了它们之间的相互作用。他们在MNPs表面放置了表面酸性磷酸酶(AP),然后Fmoc-Tyr(H2PO3)-OH被AP转化为Fmoc-Tyr-OH,随后在PBS缓冲液(pH=6)中围绕MNP核心自组装形成水凝胶。静电吸附作用使肽水凝胶能够从溶液中收集CPO。MNP-CPO@Nanogels能够成功进入癌细胞的溶酶体。通过受控的交变磁场激活MNPs,增加了细胞内的活性氧(ROS)和过氧化氢(H2O2)。结果,CPO的催化反应将H2O2转化为高毒性的1O2,从而导致细胞凋亡(图4d)。在体内肿瘤治疗中,所有组在第一周均呈上升趋势,但只有磁性纳米颗粒-环丙烯基过氧化物酶@纳米凝胶组在第三周消除了肿瘤。
5 超声处理
超声处理最常见的是通过完全混合肽聚集体、将其分解成更小的片段,以及增强氢键和疏水相互作用来促使肽水凝胶的形成。此外,超声处理会产生热量,有助于更有效地溶解肽。超声处理还能改变肽的二级结构,使其从无规卷曲转变为β-折叠。本节将阐明超声处理对肽自组装成水凝胶的影响。
带电氨基酸残基形成的肽结构也可能在超声作用下形成水凝胶。其中最典型的例子是RADA-16肽,它由带正电和带负电的氨基酸组成,以及促凋亡肽KLA(序列KLAKLAKKLAKLAK),它们能够在溶液中有序自组装形成纳米结构。两个RADA-16序列通过Gly-Gly二肽作为连接基团与KLA肽相连,经过20分钟的超声处理后,它们会转变成一种水凝胶,这种水凝胶不仅能诱导癌细胞死亡,还能阻止肝癌细胞相互黏附和迁移(图5a)。这种RADA-KLA肽水凝胶在80微米浓度下几乎完全抑制肝癌细胞(HepG2)黏附。同时,在该浓度下,HepG2细胞迁移受到75%的抑制。PuraMatrix已经将RADA-16商业化。
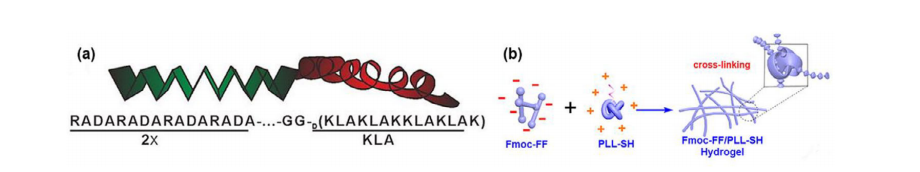
a)RADA-KLA肽分子结构的图示。b)基于静电相互作用触发共组装的Fmoc-FF/PLL-SH水凝胶形成示意图。
基于静电相互作用,带正电荷的聚-L-赖氨酸(PLL)和带负电荷的二肽N-芴甲氧羰基二苯丙氨酸(Fmoc-FF)可用于增强自组装纤维之间的附着,从而形成具有可调流变特性的可注射水凝胶。在pH值为7.5的条件下,经15分钟超声处理后,混合溶液在室温(25°C)下转变为水凝胶。在向PLL中添加硫醇基团后,Fmoc-FF/PLL-SH纤维形成类似于菌毛抗原的α-螺旋形状,从而能够被免疫系统识别。通过形成二硫键连接,添加硫醇基团还能进一步稳定人工螺旋纤维结构(Fmoc-FF/PLL-SH)。水凝胶疗法使荷瘤小鼠体内CD4+、CD8+和CD3+T细胞数量显著增加。然而,水凝胶能够抑制肿瘤生长,但无法引发免疫反应以完全清除肿瘤组织(图5b)。经过12天的治疗,对照组的肿瘤体积增长至约3000立方毫米,而治疗组的肿瘤体积仅为对照组的五分之一。
6 氢离子浓度(pH 值)
氢离子的浓度在肽水凝胶的成功构建中也起着关键作用。在不同的 pH 值条件下,酸性氨基酸(如天冬氨酸和谷氨酸)以及碱性氨基酸(如赖氨酸和精氨酸)能够接受或失去质子,从而导致氨基酸侧链的离子化状态促进或阻碍水凝胶网络的形成。
凝胶形成条件可以从碱性到酸性。例如,C16-VVAEEE的三肽(VVA中性氨基酸)核心促进氢键和β折叠的形成。通过平衡VVA两侧的烷基链和谷氨酸序列,可以调节自组装过程的pH敏感性。C16-VVAEEE在略酸性条件下(约pH6.8)自组装,然后在室温下搅拌24小时形成凝胶。C16-VVAEEE纳米纤维靶向内质网,在低酸性条件(pH6.8和7.1)的细胞中引起显著的细胞毒性,但在pH7.5的细胞中则没有。有趣的是,纳米纤维可以通过经皮给药来治疗肿瘤(图6a)。通过经皮给药实现最佳的肿瘤治疗效果,治愈率为60%。此外,腺嘌呤这种亲水性核碱基与疏水性驱动结构(三个苯丙氨酸)结合形成纳米纤维单元(Ade-FFF)。这些互补的核碱基可以通过独特的氢键(沃森-克里克相互作用)相互作用,同时肽基团通过π-π堆积相互作用,从而实现纳米纤维单元的自组装。当浓度设定在0.45%以上且pH值降至7.5时,采用pH开关法实现了Ade-FFF凝胶化。此外,当向Ade-FFF水凝胶中加入阿霉素(Dox)时,它能够嵌入腺嘌呤堆积结构之间,使水凝胶的储能模量比单独的Ade-FFF提高约十倍。因此,Ade-FFF水凝胶能够容纳大量阿霉素,并在Ade-FFF水凝胶降解过程中持续释放阿霉素(图6b)。负载阿霉素的Ade-FFF水凝胶不仅将生存时间延长了至少20%,而且最终肿瘤体积仅为阿霉素溶液组的一半。
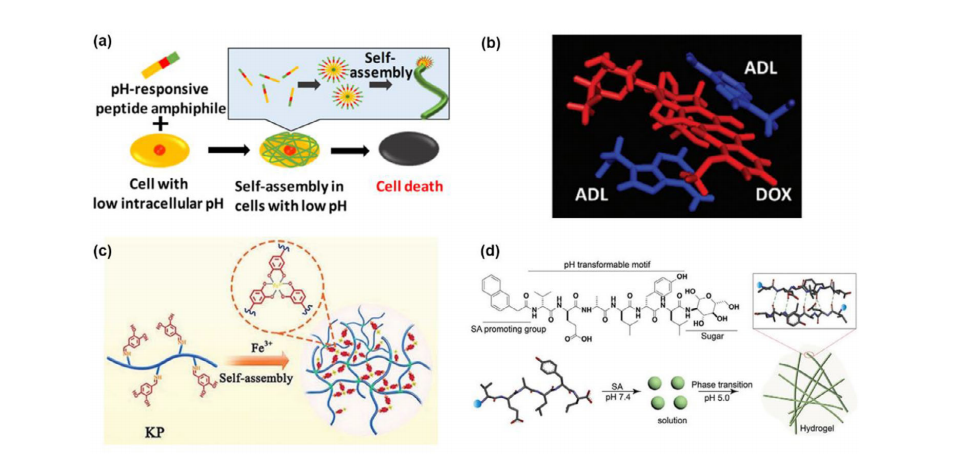
a)一种细胞内pH响应性肽两亲分子的分子自组装诱导选择性细胞死亡的示意图。b)Ade-FFF自组装与阿霉素(Dox)的分子动力学模拟显示药物嵌入腺嘌呤堆积结构之间。c)KL、PCA和Fe3+离子的抗癌肽自组装制备可注射水凝胶的示意图。d)典型的LTP分子结构,由SA促进基团、pH可转换基序以及C端有或无糖修饰组成。LTP在pH7.4的水溶液中可形成纳米颗粒,并在酸性条件下(pH5.0)通过非共价相互作用转变为纳米纤维,进一步缠绕形成自组装水凝胶。
有趣的是,凝胶的形成条件也可以从酸性变为碱性。在室温下,将含有赖氨酸(碱性氨基酸)和亮氨酸(中性氨基酸)组合而成的抗肿瘤肽KLLKLLKKLLKLLK-NH₂(KL)、4,4-二羟基苯甲醛(PCA)和氯化铁的溶液的pH值从6.5升至7.4时,会发生凝胶化形成KPFe水凝胶。KL与PCA之间亚胺键和Fe(III)邻苯二酚配位连接的溶解与重新建立是KPFe水凝胶pH敏感性的原因。KPFe水凝胶还表现出温度敏感性,在60°C时可转变为溶液。此外,加入Fe³⁺离子会增加分子交联密度,增强机械强度(G₀=1272帕),并且在凝胶基质中通过芬顿反应将H₂O₂分解为・OH自由基。KPFe水凝胶在808纳米处的光热转换效率为24.7%,这归因于Fe(III)-邻苯二酚配合物。如图6c所示的示意图,KPFe+近红外(NIR)组的抗肿瘤能力优于KPFD(KPFe包裹盐酸多柔比星)组,但KPFD+近红外组与KPFe+近红外组相比,可增加细胞坏死率(从38.53%提高到47.36%)和细胞凋亡率(从16.84%提高到23.94%)。
肿瘤微环境中的pH值变化也会改变肽自组装的纳米结构。从胰岛素聚集的关键部分选取了酸性响应区域VEALY作为核心,N端添加2-萘乙酰基,C端进行糖基化,分别提高了自组装能力和蛋白水解稳定性。肽(LTP)在癌细胞溶酶体中生成水凝胶,使癌细胞对化疗药物更敏感,并通过扩大溶酶体和改变膜通透性导致细胞坏死或凋亡。在pH值为7.4时,肽自组装成纳米颗粒,通过小窝介导的内吞作用进入细胞并在溶酶体中富集,因此在pH值为5.0(与溶酶体pH值相当)时,羧基去离子化,加速分子自组装形成纳米纤维(图6d)。治疗后,与PBS组相比,纯LTP可使肿瘤重量减轻40%,与阿霉素联合使用时,肿瘤重量可减轻80%。
7 光交联
JM2模拟肽(rrrrrrrr-VFFKGVKDRVKGRSDC)包含CX43微管结合序列(KGVKDRVKGK)以及具有重要细胞穿透肽功能的聚D-精氨酸(rrrrrrrr)。JM2接枝到甲基丙烯酸化透明质酸(HA)上,在光引发剂LAP溶液中,经365纳米紫外线照射10秒即可形成水凝胶。在原位乳腺肿瘤切除的小鼠模型中,JM2-HA水凝胶能够保证抗肿瘤效果,并在体内抑制复发。JM2-HA水凝胶组的复发肿瘤体积仅为对照组的三分之一,且显著小于两组分组(JM2组和HA组)的体积。JM2通过直接降低细胞周期相关蛋白(如细胞周期蛋白A和细胞周期蛋白E)以及参与细胞外基质(ECM)成分降解的MMP2和MMP9等蛋白质的表达,有效抑制癌细胞增殖、迁移和侵袭。当JM2与线粒体竞争微管结合时,释放的线粒体聚集在微管附近。随后,大量聚集的线粒体引发了活性氧诱导的活性氧释放(RIRR)反应以及线粒体介导的癌细胞死亡,其机制如图7所示。
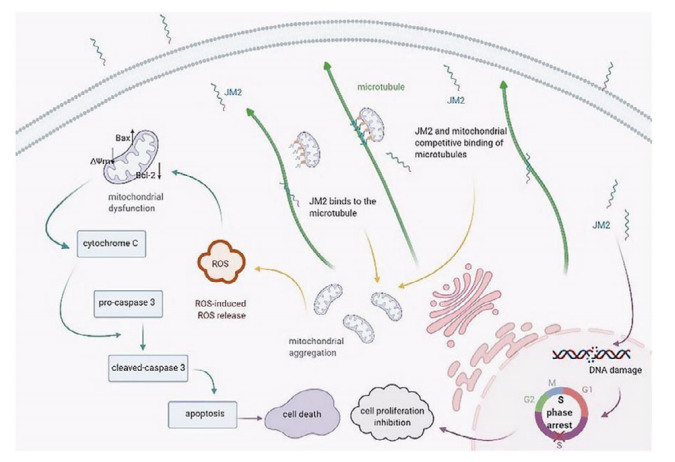
图7 JM2的抗肿瘤机制。JM2应用于肿瘤治疗的可能分子机制图示
8 液-液相分离
近年来,一些学者提出了通过液-液相分离(LLPS)制备水凝胶的方法。严学海团队从冷冻透射电子显微镜(Cryo-TEM)、全原子分子动力学(AAMD)和粗粒分子动力学(CGMD)等多角度阐明了肽在液-液相分离介导下的多步自组装过程。肽分子在液-液相分离过程中从初始液滴到纳米团簇再到纳米纤维,最终形成凝胶的结构演变。银离子的存在延长了液滴的存在时间,这有利于捕捉Fmoc-Ala液-液相分离过程的证据。在早期阶段,液滴中间体主要通过疏水相互作用稳定。在形成具有长程有序结构的纳米纤维时,各种分子间氢键起主导作用。同时,有序的π-π堆积效应不断增强。银离子的配位作用有助于所形成纳米纤维的稳定性。[81]通过溶剂交换法将羧苄基保护的二苯丙氨酸(z-FF)的DMSO溶液用双蒸水稀释四倍,得到了z-FF水凝胶。然而,不同的初始孵育温度(IIT)会导致凝胶化过程早期出现不同的液-液相分离(LLPS)现象,并形成相应的亚稳态液滴。z-FF水凝胶的IIT与最佳机械强度相对应,且随着浓度的增加,该温度也会升高(图8a)。当浓度为0.5毫克/毫升且IIT为40°C时,z-FF水凝胶的机械强度最强(G'约2400帕,G"约700帕),是IIT为80°C时凝胶强度的五倍,是IIT为20°C时凝胶强度的两倍。[52]此外,他们还进行了更详细的相分离实验,发现多步脱溶是引发LLPS介导的肽自组装的关键因素。通过影响早期组装条件,如加热、有机溶剂和水的比例,并更换不同的溶剂。这些过程会影响非共价键力,例如疏水键、π-π堆积和疏水相互作用,从而形成不同的组装结构,可能产生纳米带、纳米管和螺旋纳米纤维。

图8 a)不同初始孵育温度下凝胶化过程中不同液-液相分离(LLPS)的示意图,包括相应亚稳态液滴的形成以及最终水凝胶网络的结构b)通过液-液相分离制备微毛细管网络(µCN)水凝胶的示意图。c)RLP-PEG肽水凝胶制备的进展。
另一位学者田口哲史(Tetsushi Taguchi)也对相分离水凝胶进行了广泛的研究,包括其促进体外血管生成和促进干细胞移植的潜力。将用乙烯砜(sGVS,pH=7.8)改性的猪皮源明胶(sG)、用硫醇基团(sGTH,pH=6.4)改性的sG以及用脲嘧啶酮单元(tGUPy,pH=7.4)改性的猪腱源明胶以1:1:2的比例剧烈混合,并在37°C下放置十分钟,形成LLPS微孔凝胶(µCN水凝胶)。在生理条件下,交联的µCN水凝胶通过内部tGUPy的溶解形成多孔结构。脲嘧啶酮单元之间的四重氢键对于该LLPS系统的稳定形成具有重要意义。µCN水凝胶具有相似的血管网络特征,能够模拟体内营养物质和信号分子的运输途径(图8b)。与非多孔凝胶相比,与小鼠成肌细胞(C2C12细胞)共培养三天后,细胞数量增加了2.3倍。使用微纤维素水凝胶移植间充质干细胞(MSCs)可在14天内治愈患有后肢缺血的小鼠。[83]在另一项研究中,他们进一步对tGUPy的巯基和乙烯基砜基团进行修饰(分别形成tGUPyTH和tGUPyVS),并分别制备了pH值为7.8和6.4的PBS溶液。将pH值为7.4的透明质酸溶液与微纤维素溶液以1:1:2的比例混合,并在37°C下孵育30分钟,形成液液相分离微纤维素水凝胶。与微纤维素水凝胶相比,内皮细胞在微纤维素水凝胶中的铺展和管形成情况得到改善。上述工作对肿瘤血管类器官和癌症治疗中的干细胞移植具有积极影响。
相分离肽水凝胶也可以在纯水环境中制备。黄冠刘通过化学交联固定类弹性蛋白聚合物(RLPs)和聚乙二醇(PEG-4NH2-20k)液液相分离溶液制备了水凝胶。RLP-PEG肽水凝胶包含弹性微区,孔径在20至90微米之间,机械强度各异(图8c)。该水凝胶的G'值约为600帕,杨氏模量约为13千帕。
参考文献:doi.org/10.1002/smll.202503697

电话:0551-65177703 邮箱:pb@peptidesbank.com 地址:安徽省合肥市四川路868号云谷创新园A6栋3层
合肥肽库生物(Taikubio)只为有资质的科研机构、医药企业基于科学研究或药证申报的用途提供医药研发服务, 不为任何个人或者非科研性质的、非用于药证申报使用等其他用途提供服务。