1 pH��Ӧ�ĵ����
����ҩ�ﻯѧ�����﹤������õ��˹㷺���о�����21�ֳ��õİ�������Ϊ�ĵ���Ҫ�ɷ֣�����������Ľ���֮��ͨƪ�õ���ĸ������ָ�����ǡ�21�ֵ��Ͱ�����ͨ�����ǵIJ��������֣����Է�Ϊ���ࣺ
-
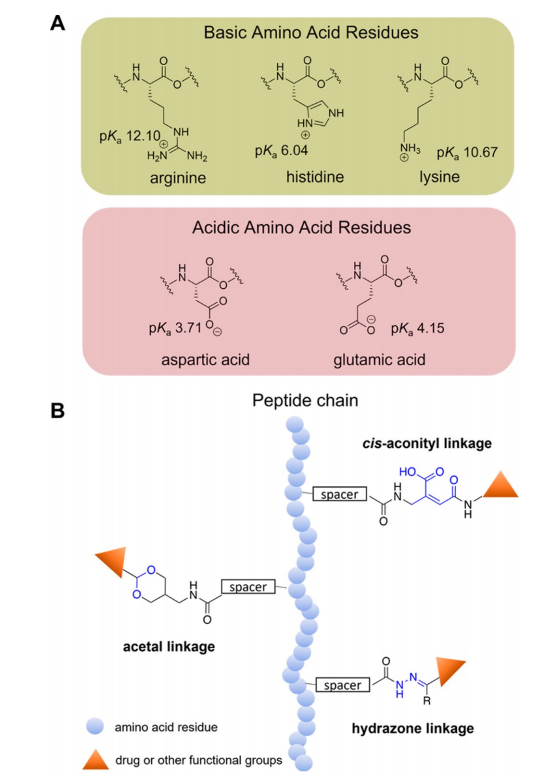
���д���ɲ����İ�����[������(R)���鰱��(H)��������(K)���춬����(D)�Ȱ���(E)]��
-
���м��Բ�����ɲ����İ�����[˿����(S)���հ���(T)���춬����(N)�Ȱ�����(Q)]��
-
������ˮ�����İ�����[������(A)���Ӱ���(V)����������(I)��������(L)��������(M)����������(F)���Ұ���(Y)��ɫ����(W)]��
-
�Լ������������[���װ���(C)�������װ���(U)���ʰ���(G)������(P)]��
�����а������У����д�������İ����������pH��Ӧ������������Ҫ����Ϊ�����ӻ���ȥ���ӻ�ʱ��ˮ��/��ˮ�Իᷢ���仯��Ȼ����������ĵ�һpKaֵͨ��������Ԥ����̬�仯���ܷ�����pHֵ�����磬�ڽ������ڴ����벻ͬ�����Ĵ���ͬ��ɵİ������Ӱ�����ǵ�pKaֵ��Williams������һϵ�ж�����֤��������Ӱ�죬��Щ�����д�����ɵ�D�л��������������ӣ���������pKa����D�л����������Ӷ����͡��ڸ����ӵ���ϵ�У������������������ͬ���͵ĵ�ɣ���pKaֵ��ͬ������ݺ��ɭ-�������Ͷ��շ��̹��㣬����pKaֵҲ��������ͬ�����⣬����ͬʱ���м��Ժ����Բл����ģ���pH��Ӧ��Ҳ���ܵ�Ӱ�졣
����ϸ������֯��쳣�ı仯��������Χ����pHֵ�ı仯��ͼ1)��pH��Ӧ�Ŀ�������������ض���֢�Ͱ������ơ���ˣ��ڱ����У����ǽ�����pH��Ӧ�ĵ���ƣ��ص��ע��(1)������ѡ��(2)ͨ��pH���м����ӿ��ѽ�Ĺ��ܻ��š�
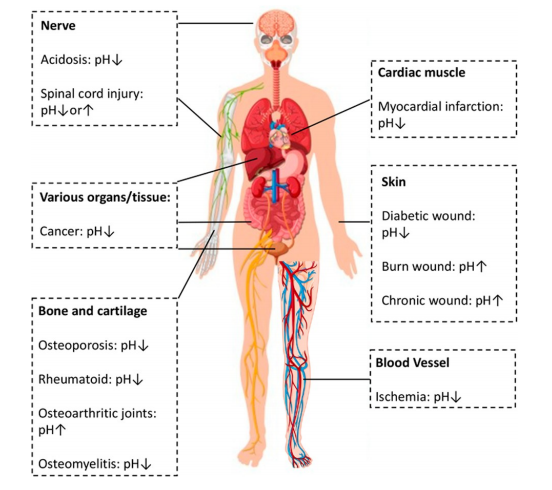
ͼ 1. ����������������ʱ���� pH ֵ�ı仯
1.1. �������pH����Ӧ
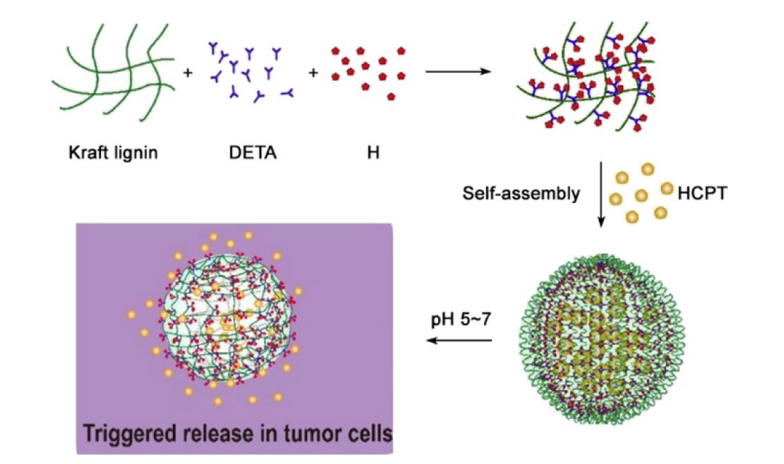
���簱����ɷ�Ϊ�������ᣨ������ɣ����������ᣨ������ɣ����������ᣨR��H��K����������pH��Ӧ������װ�Ļ���ϵ�У�ͼ2A���������������²������ӻ�����һ�������ṹ����̬��ҩ���ͷ����ʵı仯�����磬�콨�����������һ��pH��Ӧ��ϵ������KKGRGDS��Ϊ��ˮ������Ӧ��ͷ����VVVVVV��Ϊ��ˮβ��������������������pH��pH7.0��������װ�γɽ���������pH5.0�£����ڴ����ӻ���K�л�֮����ų�ľ�������ã��������塣��������ҩ�ﰢù�أ�Dox��������pH7.0���������Ľ����к���pH5.0�¹۲쵽ҩ����������ͷţ��Ӷ�����������������һ�ֻ���������H�㷺�������pH��Ӧ��ϸ������(CPP)��CPP�����ڸ����������������pH�������ġ�H�л��ϵ����������pKa�ӽ�6��ʹ����״̬���������pHֵ�·����仯�����磬Lei������������һ��pH��Ӧ������װľ����-H�����������и�����ҩ��10-�ǻ�ϲ����(HCPT)��30-40�������������Ǵ���ϸ��Ĥ������ߴ磬�������������������������£�������ŷ������ӻ����������ӷֽ⣬�ͷų�ҩ�ͼ3�������������������ͨ������ʹ�ü������ὫpH��Ӧ����������ĵ�ҩ������ϵͳ�У����ڰ�֢���ơ�
ͼ2.(A)�������������л��Ļ�ѧ�ṹ��pKaֵ��25��C�£���(B)����pH���м��Ľṹ��
���˼��������⣬��������Ҳ��pH��Ӧ������װ��������еij��ò��ϡ����⣬����������������᳣������ڵ��ת����ϵ����ơ����磬Kim����ͬ�����ýṹΪ�ۣ���ϩ�ǰ���(PEI)�CKn�CEm(PKE�ۺ���)��ABC��Ƕ�ι�������л��ݡ���Ϊһ��pH��Ӧ�Ե��ת���ۺ��EmǶ�������ڻ�������������pHֵ����ˣ�������Ʋ��ϳ��˾��в�ͬ����E�л���7��9��13����Ԫ����PKE�����о����ǵ�����װ�ͻ�������������������뵥����PEI��ȣ�PK5E9����װ������������������������е�תȾЧ�ʡ������תȾʵ�����������pH��Ӧ�Ե��ת������ЩPK5E9��۸�������Ч�����������壬������pH6.0��ʹϸ����ȡ�������1.5-2.5����תȾЧ�ʱ�pH7.4�������50����ͻ�����������pH��Ӧ������ƻ������ϵͳ��
ͼ3.����װ����AL-His/HCPT��������ʾ��ͼ��DETA������������H���鰱�HCPT��10-�ǻ�ϲ���
1.2. �����Ŷ�pH����Ӧ
����ͨ������ѡ����������pH��Ӧ��֮�⣬�������pH���ѽ���Ĺ��ܻ���Ҳ���ṩ�������ƿռ䡣��Щ������������pH���ȶ����������Ի���������»ᷢ���ѽ��ṹ�仯�����ַ�������ѹ㷺Ӧ����pH��Ӧ������װ�Ļ��������������Ϊ��pHֵ�����仯ʱ���ǹ����Է��ӻ��þ��й����ԣ��Ӷ��ṩһ�������Եġ����ܡ��������Ʒ���������pH���ȶ����������ꡢ˳ʽ-��ͷ����������������������ȩ(ͼ2B)������pH��Ӧ��ϵ�����磬Balci��Top�����һ��pH��Ӧ��ҩ�����ϵͳ����mPEG-����Doxͨ������ѽ�����ż�������������ڳ��¼����������µ�ҩ���ͷš�ż����AT1����pH��Ӧ�Ե�H6�ṹ�����DZȽ���ͨ����ֱ����Doxż����mPEG��mPEG-HYD-Dox����ͨ�������ӵ�mPEG-��ż���mPEG-AT1-Dox����ҩ���ͷź�ϸ�����������������mPEG-AT1-Dox��pH5.0��72Сʱ�ͷŵ�Dox��ԼΪmPEG-HYD-Dox��3������ϸ�����Ը���mPEG-HYD-Dox������������pH��Ӧ�Ե������Ȼ�����ڴ���ҩ���ͷţ�������ͨ���ڵ���ϵͳ����м�������pH��Ӧ�Ե�������ǿ��Ч����
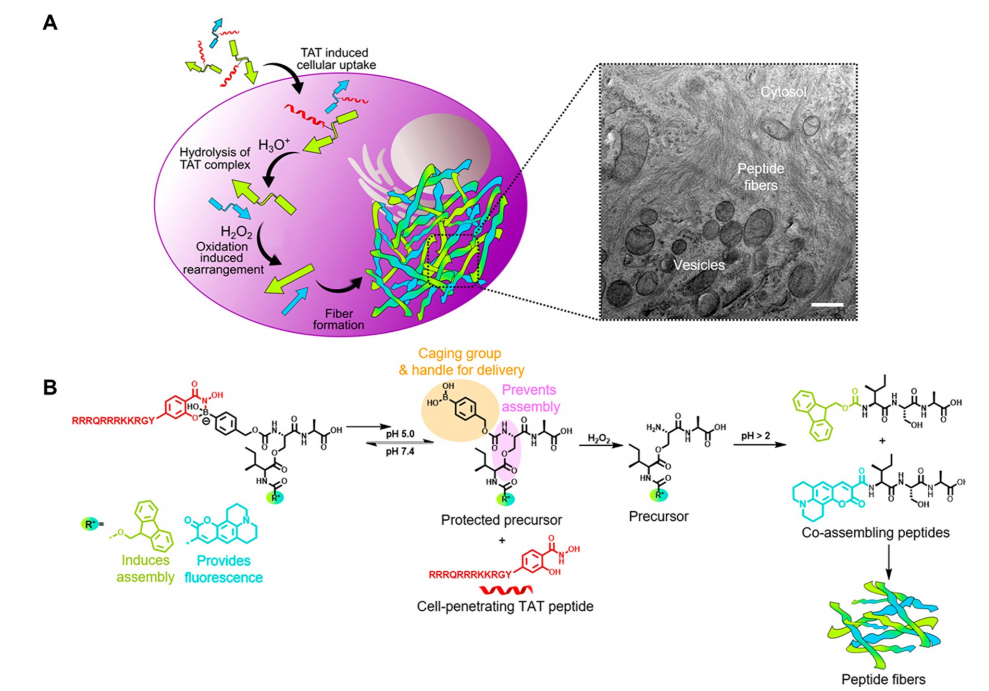
�����Weil����ͬ�½�pH���е�����-ˮ���������(SHA)��������һ�������ĸ������У�ּ����ϸ���ڽ��ж��ת��������ư���һ���ɴ�ϸ����TAT�ģ����������ε��������Ľ�ϣ�������������е�SHA���ţ��������������Ի�������ˮ�⡣�����������ͷ������ϸ���ڵĹ��������ѽ⣬��������O��N����ת�ƣ���һ���γ��ģ���Щ����A549ϸ���ڹ���װ����ά���磨ͼ4������ϸ�������������յ������Ͷ��ת��Ϊ�����ϸ�������ṩ��ƽ̨�����⣬�����о�������˸÷�����δ����չ�����ص��ǽ���������pH��Ӧ�ԺͶԴ�л����������Ӧ�Ĺ��ܻ������ϵ�����װ���ṹ�У��Ӷ�ʵ�ֶ��ƻ�ҽ�ơ�
ͼ4. (A) ����ϸ���ڹ���װ������ʾ��ͼ�������һ�����γ��˿ɼ�����ά���磬��ͨ�������������TEM��������500���ף��۲쵽��(B) ϸ���ڷ����Ļ�ѧ��Ӧʾ��ͼ��
2. ��Ӧ����
��pH��Ӧ�����У��IJ����ĵ���״̬ͨ��������Ƭ�ε���ˮ�Ի���ˮ�ԣ��Ӷ���������ϵ�Ļ�ѧ��/����̬��Ӧ������Ҫ����Ҫ���ǵ��IJ�������һ����ȫ�����롣�����IJ��������ŵ�����Ա˴˶���������Ը��ݺ��ɭ-�������Ͷ��շ��̹�������pKa������������£����㹻��IJ��������ŵ����ȥ����ʱ�����ܻᷢ������װ��̬�仯����ȡ���ڲ���pH��ӦƬ�ε������ԣ����������ĵ������ԡ������пɵ�������İ���������������pKaʱ��Ԥ����̬�仯���ܷ�����ʲôpHֵ�±�ø�������ֵ������������һ�������б˴˽ӽ�ʱ��Ȼ������ҩ����ͺ���֯�����еĴ����pH��Ӧ��Ӧ���У���pH�����仯ʱ�ĵĵ�ɻ���٣��Ӷ���������װ����pH�����仯ʱ���ĵĵ��Ҳ�����ӣ���������װ��̬����չ����ȫ���塣���pH�仯��������������װ���ṹ֮�����̬ת��������������Ӧ���ƣ�����װ����չ/�������̬ת�����У�pH����������װ������Ϊֱ�ө�������Ƿ����ܽ�ģ�Ȼ��pH�仯���µ���״̬����ˮ/��ˮƽ�ⷢ���仯���յ�����װ�����ṹ����չ/�������̬ת�����ƿ��ܸ����ӡ�ͨ���������ĵ�pH��Ӧϵͳ����ʹ���״̬������װ�����ṹ�����Ż���pHֵ�ı仯�������ĵ�pH��Ӧϵͳ�е�ijЩƬ�ο��ܻ�������ɻ�ɣ��Ӷ���ø�����ˮ������ת��������ų���������ͣ������յ��·ֽ�ɷ����ܽ����֡����ߣ��Ŀ������Ϊ����װ������Ƭ�λ���pHֵ�仯ʱʧȥ��ɣ��Ӷ���ø�����ˮ����Щϵͳͨ���ᾭ����̬ת����Ӧ���ơ�����pH��Ӧ�ľ������Ƿֽ����̬ת�����������״̬��Ψһ�ı仯����ô��ЩpH��Ӧϵͳ���Ա��ֳ������ת����Ϊ�����ܿ��ܴ��ڽϴ���ͺ������ǣ������̬ת�����ɻ�ѧ�仯����������ѣ������ģ����ת��ͨ���Dz�����ġ����ǵ���ҩ�����ͺ���֯�����еĴ����Ӧ���в���Ҫ�����ԣ������ڴ��ص��ע���������͵��л����ƣ������������ǵģ�DZ�ڣ������ԡ�
2.1 pH ����������װ
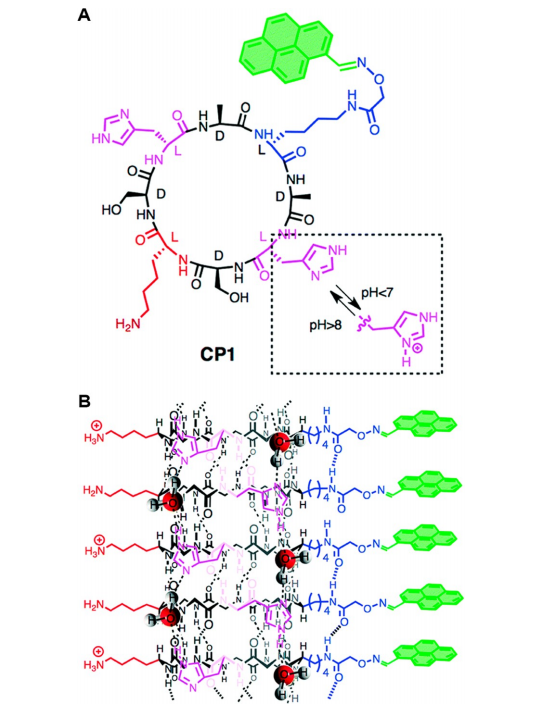
��pH����������װ�����У��ķ���ͨ�����ڲ����ϴ��в��ֵ�ɵ�����·����ܽ⡣pHֵ�ı仯�ᴥ�����ӻ���ȥ���ӻ����Ӷ�����������������ɵ���������������������ľ����ų��������գ���������㻷�ѻ����������Ӽ�����ôٳ�����װ�����磬�Ž��ڼ���ͬ�����������һ��������KRRFFRRK����������pH����������װ���ơ���pH 7.4�£����ı��ֵ���״̬���γ�С�ġ���״������ľۼ��壬����K��R�л������ӻ�����pH������9.4ʱ��K��R�л�����ȥ���ӻ����ھ����ų������ͺ���ˮ�������ǿ�������£��յ�����װ��������ά����һ����������Montenegro��Granja����ͬ�±����ģ����������һ�ֻ�״�ģ�CP1�����ڼ���ͼ5A�����û��ĵĹؼ��ɷ�������H�л���һ��K�л�������������ˮ������ʱ��CP1�ϴ�����ɵ�H��K�������������Ӿ����ų����ö���ֹ�˾ۼ�����ˮ�����к���pHֵ> 8��H��������ȥ���ӻ����Ӷ�����CP1����װ�������ܣ�ͼ5B). ��һ�����Ӽ�¸���H��K����ȥ���ӻ����յ�����װ�ɷּ��������磬�����γ���H�л�����ȶ��Ĵ���ά��
ͼ5. (A) CP1���ĵĽṹ��B) CP1����װ�ɵ��������ܡ�
2.2 pH ���������ͺͷֽ�
���� pH ֵ�ı仯������װ����̬�������ͣ����ͣ���ֽ⣨ͼ6A)������Щ����£�����pHֵ�ı仯���������ӻ���ȥ���ӻ���������������ӣ��Ӷ���ǿ���Ӽ���ų�����ã��������ͺͽ��塣Kim����ͬ�±�����һϵ�п��γɽ�����pH��Ӧ�Ժϳɹ�������ɾۺ϶ȣ�DP��Ϊ55�ľۣ�N-�������ϩ������Ƕ�κͲ�ͬ�ۺ϶ȣ�n =50��75��100��125����H nǶ����ɡ�������pH�£� HnǶ�η������ӻ������������ǿ�����½����ṹ���͡���ͬ������Ķ�̬��ɢ��������ʾ����pHֵ6.5��10�ķ�Χ�ڣ���Һ�еĽ����ߴ籣�ֲ��䡣Ȼ��������ҺpHֵ����5.6ʱ�������ߴ缱������������ҺpHֵ����������pH 2���۲쵽����Ľ���������H nǶ�γߴ���Σ������ߴ���pHֵ�Ľ��������Ƶ����������������������pHֵ�Ľ��ͣ����ӻ���H�л��İٷֱ�ˮƽ���ơ�
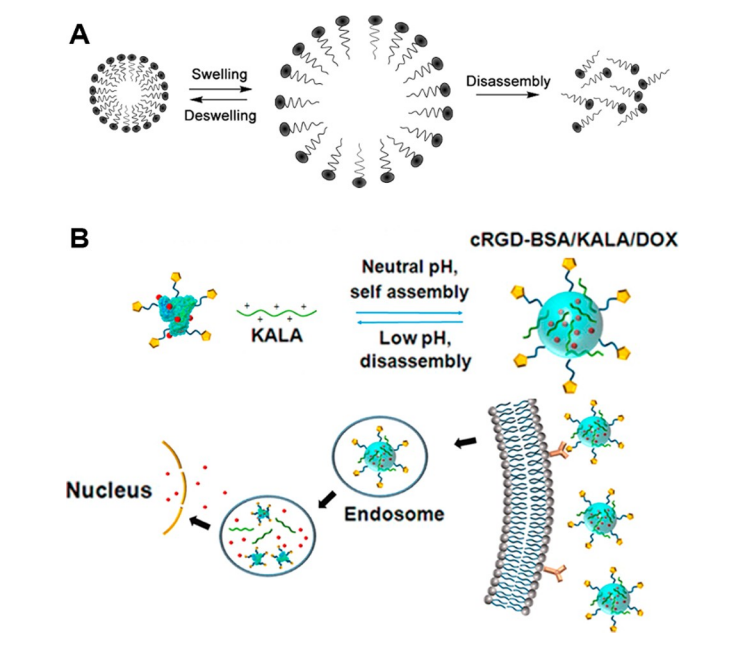
ͼ6.(A)pH�仯�����������������źͽ�����Ӧ����ʾ��ͼ��(B)cRGD-BSA/KALA/Dox�������ӵ�����װ/������Ϊ�Լ�Dox��ϸ���˵�ҩ�����;����
����װ���ѱ���Ƴ��ܹ��ڲ�ͬ��pHֵ�£�ͨ�������pH���м����ѣ�һ����ಽ�ֽ⡣Cheng���������һ�������յ��ֽ�����ӡ����DZ�����һ��˫��������ҩ�����ϵͳ�����д�����ɵ�KALA������װ�γ���㣬��״Arg-Gly-Asp��cRGD�����������ɵ�ţѪ����ף�BSA����ϲ�����Dox�γ�С����������Ϊ���ģ�ͼ6B). ����pHֵ�Ľ��ͣ�BSA��KALA֮��ľ�������ü���������������������ͺͽ��塣���⣬����cRGD-BSA�����и�������ڵ�pHֵ�����ӻ��������������ɵİ�ù�ط��ӵ�����ü��������º��Ľ��岢�ͷŰ�ù�ء�Kataoka������Ƶľ��Ҷ���-Ƕ��-���춬����-��-��ù��Ƕ�ι�����(PEG-PAHD)����һ����pHֵ�������������յ���������ӡ���Ƕ�ι������γ����κ˿ǽṹ��ֱ��ԼΪ65���ס�PEG-PAHD������������pHֵ���ȶ�������pH 4-6ʱ����������Ѻ���ˮ��Dox��Ԫ�ͷŶ�Ѹ�ٷֽ⡣
2.3 pH��������̬ת��
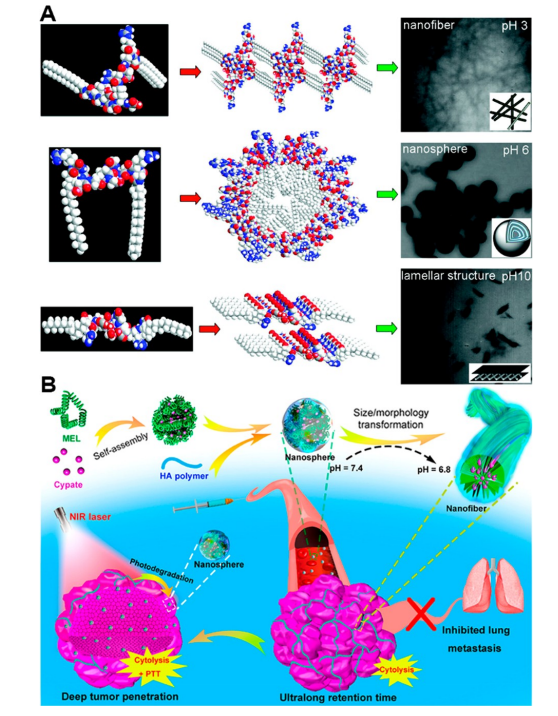
���ֻ���ֲ�ͬ������װ��ò֮���ת���������һ����Ӧ���ơ�����������ѧ���ںϳ����ײ��Ϸ���ȡ���������չ����Щ���ײ��ϵ���ò����pHֵ�ı仯���仯���Ӷ����²������ܵı仯�����磬Zhang���˺ϳ�������������[P1:(Ӳ֬��-GRGDG)2KG��P2:(Fmoc-GRGDG)2KG����P3:(������-GRGDG)2KG]�������ڲ�ͬ��pHֵ������װ�ɸ�����ò������������ά��������Ͳ�״�ṹ��RGD������Ϊ��ˮ�Ǽ����룬Ҳ������Ķ�֮һ�����м��Բл���Ҳ�����Բл����ܹ���ӦpH�ı仯��P1��pH3.0ʱ�ۼ��γ�������ά����pH6.0ʱ����װ�γ�������pHֵ��һ��������10.0ʱ����һ��ת��Ϊ��״�ṹ��ͼ7A).P2��pH3.0ʱ����װ�ɲ�������ṹ������pHֵ�����ߣ���ת��Ϊ������״�ṹ�����ݡ���P1��P2��ȣ�P3��ȱ���ϴ����ˮ���ţ����¸��ض������γ����������ṹ��
ͼ7.(A)P1���ڲ�ͬpHֵ�µ�����װʾ��ͼ��TEMͼ��B��MEL/Cypate@HA��������Ʊ�ʾ��ͼ���Լ��������������������ͽ����⼤�������³����ijߴ�/��̬ת�䡣
Wu���˱�����һ�����ڵİ������ص��ע����̬ѧ��ת�䡣�������о��У�����������ϸ���ķ䶾��(MEL)�����������շ��ӻ�������(cypate)��������(HA)����ϣ��Ʊ���һ����������Ӧ������MEL/Cypate@HA�ṹ������������װ����������pH���͵�����½�һ��ת��Ϊ������ά��Ȼ����NIR�����±��������ͼ7B).��Щ��������pH��Ӧ����γ�Խ��װ/��ж��ʵ������װ���ṹ�ĸ��ӱ仯����һ������δ���ɹ㷺Ӧ��������ҽѧӦ�á�
�ο����ף�doi.org/10.1021/acsabm.2c00188