
摘要:
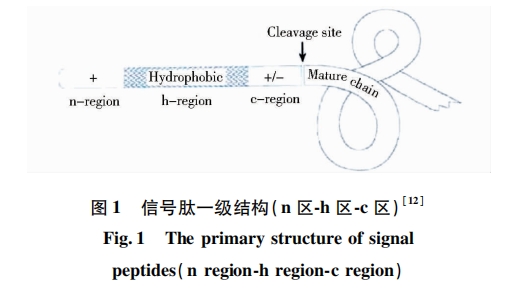
信号肽位于分泌蛋白的N端,一般由10~40个氨基酸残基组成,并大致分为3个区段:①n区(n-region)为带正电荷的氨基酸,如赖氨酸和精氨酸;②疏水区(h-region)由9个或更多的以中性氨基酸为主组成的疏水核心区,能够形成一段L螺旋结构,常见有亮氨酸、异亮氨酸等;③加工区(c-region)是信号肽酶切割信号肽的部位,由极性、小分子氨基酸(如甘氨酸、丙氨酸、丝氨酸等)组成。一般认为疏水区氨基酸序列越长、疏水性越强的蛋白质分泌效率就越高,因此疏水区的长度及疏水性对蛋白质分泌十分重要[10, 11]。信号肽结构如图 1所示。
2.2 信号肽转运机制
信号肽介导的蛋白质转运可分为两大类: 若某个蛋白质的合成与转运是同时发生的,则属于翻译-转运同步机制; 如果蛋白质从核糖体上释放后才发生转运,则属于翻译后转运机制。
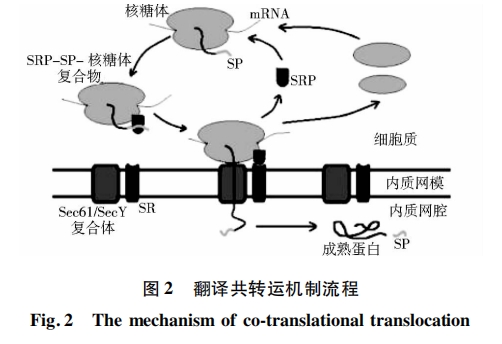
(1)翻译共转运途径[13]。核糖体串联成聚核糖体,准备合成蛋白质→信号识别颗粒(SRP)识别并结合在核糖体上→在聚核糖体上,由mRNA开始合成信号肽→SRP识别信号肽并与之结合,多肽链合成暂时停止→SRP-信号肽-聚核糖体复合物与内质网膜上SRP受体及核糖体受体结合→暂时停止合成的多肽链恢复合成,新生肽链随信号肽延伸,接着信号肽被膜内侧的信号肽酶水解,新生肽链继续延伸,形成有高级结构的成熟蛋白质。翻译共转运途径如图 2所示。
(2)翻译后转运机制。特定条件下蛋白质在其合成结束后才开始转运,采用这种机制定位的蛋白质一般都是可溶性蛋白质,它们有一段适度疏水的信号肽序列而使它们在合成时避开了 SRP 的识别。
3 信号肽对抗体蛋白分泌效率的影响
3.1 一级结构氨基酸组成
大量研究表明,对于动物细胞而言,信号肽的疏水核心长度和疏水性对于抗体蛋白的分泌效率至关重要[14, 15],强疏水性能够高效引导新生肽的转运。有研究表明,在对全球热门抗体药物阿瓦斯汀(Avastin)的重链信号肽疏水核心区进行一级结构改造后,用新的疏水性较低的信号肽取代原有信号肽,由此造成阿瓦斯汀产量降低[16]。此外,研究酵母 PhoA 信号肽疏水核心Leu与Ala 的比值时发现,当 Leu/Ala 为 6∶4 时,前体蛋白分泌加工能力最高,在此比值附近,信号肽分泌加工能力随疏水性的增强而提高[17]。
3.2 二级结构的空间构象
信号肽序列的二级空间构象能够与mRNA翻译起始因子相结合,但过于复杂的二级结构却会引起翻译阻遏效应。研究表明,全人源化抗HBsAg抗体的原信号肽经过优化后二级结构能量从原来的ΔG=-19.7kcal/mol降至ΔG=-15.9kcal/mol,最终优化后的轻链信号肽序列蛋白质表达量比原轻链信号肽序列表达载体高4~8倍[18]。
4 信号肽序列优化策略
研究发现,许多原核和真核细胞甚至不同物种间的信号肽在功能上是通用的,国外进行基因表达所采用的分泌信号大致有以下几种情况:①与表达蛋白质同源的分泌信号肽;②与宿主表达细胞同源的分泌信号肽;③机体内表达的分泌蛋白的分泌信号肽。不同物种间的信号肽都可以用于某一特定宿主细胞表达系统中表达免疫球蛋白[19]。在动物细胞表达系统中表达重组抗体蛋白时常用到的信号肽除了自身所携带的信号肽还包括:①替换高效表达蛋白质的信号肽序列作为重组抗体的信号肽,如乳蛋白中的酪蛋白或血清白蛋白的信号肽序列等,因为这些蛋白质能在细胞中被高效分泌;②对原有抗体信号肽序列进行一级结构改造;③替换某些病毒的蛋白质信号肽序列。
4.1 替换高效分泌蛋白的信号肽
当所构建的细胞株无法利用自身的天然信号肽获得高表达分泌目标抗体时,利用高表达蛋白质的信号肽替代原信号肽可以提高目标抗体的表达量,往往同一物种的信号肽比其他物种的信号肽表达量要更高[7]。研究表明,α因子信号肽对引导小分子质量的蛋白的分泌很有效,如表皮生长因子,单链抗体Fv片段等利用α因子信号肽获得了较理想的表达量。Lars等[20]应用CHO细胞证明,与来自不同物种的或天然免疫球蛋白G信号肽衍生的信号肽相比,白蛋白和天青杀素的天然信号肽介导了抗体的最佳表达。
4.2 改造信号肽一级结构
信号肽一级结构中存在3个功能区,根据研究报道,增加芽孢杆菌信号肽N端的正电荷或增加信号肽疏水核心h区的疏水性或长度有利于提高信号肽的加工效率,信号肽C端氨基酸残基的缺失使H区更靠近切割位点,可能有利于蛋白质的易位;而增加信号肽C端的极性,则可能会使信号肽的空间结构发生变化,降低其加工效率[21]。
4.3 考虑宿主密码子偏爱性
编码同一种氨基酸的密码子称为同义密码子,蛋白质合成过程中,同义密码子的使用概率并不相同,某一物种或某一基因通常倾向于使用一种或几种特定的同义密码子,即为密码子的偏爱性[22, 23]。各种生物的密码子使用并不是随机的,而有一定的偏爱性,在选择信号肽时,应考虑到该表达宿主相应的偏爱密码子,再进行信号肽的优化。Valent等使用偏爱密码子的信号肽所得目标蛋白产量更高,稳定性也较天然信号肽好[23]。Jalah等对1L-15的天然信号肽进行优化时,联合使用偏爱密码子,使得1L-15的表达水平提高了近100倍[24]。
4.4 使用某些病毒表达载体所带的信号肽序列
某些病毒的表达蛋白是高分泌蛋白,如昆虫杆状病毒表达载体和丙型肝炎病毒系统表达外源基因的表达量高,而且能完成真核基因产物必要的转录后加工修饰[25]。但是在表达抗体药物时一般不会使用病毒载体,因为其安全性无法得到保障。
5 小 结
基于信号肽的功能对重组抗体分泌具有重要的作用,对于其的应用也越来越广泛。通过添加一段合适的信号肽序列来引导重组抗体的分泌不仅可以实现提高分泌效率和简化下游的纯化过程,而且对抗体的稳定性和活性都具有积极的作用,当然信号肽序列的选择和优化要根据不同的蛋白质及表达系统来进行。目前,高效的信号肽也正在不断被研究和挖掘出来[26]。另外,可以利用在线工具进行查询和检测信号肽序列,从基因组中快速地预测出信号肽[27],结果的可信度也比较高。例如,使用UTR Tailortech库和SignalP及Tatp软件等来预测筛选适合目的蛋白最佳分泌效率的信号肽[28, 29]。
展望未来,随着抗体药物的发展和进步,关于提高重组抗体的表达量和稳定性的研究会不断的深入和发展,方法也会越来越多,而基于信号肽优化的方法或策略将会很好地促进上述研究。
免责声明:本文为行业交流学习,版权归原作者所有,如有侵权,可联系删除,参考文献可阅读期刊原文

电话:0551-65177703 邮箱:pb@peptidesbank.com 地址:安徽省合肥市四川路868号云谷创新园A6栋3层
合肥肽库生物(Taikubio)只为有资质的科研机构、医药企业基于科学研究或药证申报的用途提供医药研发服务, 不为任何个人或者非科研性质的、非用于药证申报使用等其他用途提供服务。