ХЄТЄЈє±ѕОДЦШµгЅйЙЬБЛДФґ©НёлДєНДФїЙЙшНёµД PDCs ЧчОЄµЭЛНФШМеЈ¬Ѕ«РЎ·ЦЧУТ©ОпёєФШОпґ©№эСЄДФЖБХПЈЁBBBЈ©µЭЛНЦБДФКµЦКЈЁНј 1 єННј 2Ј©µДЧоРВ·ўХ№ЎЈОДЦРЧЬЅбБЛДФДЪЖ¤Пё°ыЙП±нґпµДлДј°ЖдМШТмРФКЬМеЎўТ©ОпёєФШОпµДСЎФсєН PDCs µДБ¬ЅУЧУЎўДФЧЄФЛ»ъЦЖТФј°ПтДФµДµЭЛНР§ВКЎЈ
BBBґ©ЛулДКЗНЁ№эКЙѕъМеХ№КѕјјКх·ўПЦµДЈ¬»тХЯґУМмИ»ЙсѕУЄСшµ°°Ч»тДіР©ІЎ¶ѕЦРСЬЙъ¶шАґЈ¬ІўАыУГКЬМеЅйµјµДПё°ыДЪНМ·ЦЧУ»ъЦЖАґїзФЅСЄДФЖБХПЎЈДФСЄЖБХПЈЁBBBЈ©їЙЙшНёµДлДЕјБЄТ©ОпЈЁPDCsЈ©УЙ BBB ґ©ЛулДЎўБ¬ЅУЧУєНТ©Оп·ЦЧУЧйіЙЈ¬АыУГДЪФґРФЧЄ°ыНМЧчУГ»ъЦЖІўУХК№ґуДФФКРнХвР©ЙъОп»оРФ·ЦЧУґ©№э BBBЈ¬ТСіЙОЄТ»ЦЦєЬУРЗ°ѕ°µДЦРКаЙсѕПµНіТ©ОпµЭЛНПµНіЎЈФЪґЛЈ¬ОТГЗСРѕїБЛДФЙшНёлДґ©ЛуОпєНДФїЙЙшНё PDCs ЧчОЄ·ЦЧУФШМеЅ«РЎ·ЦЧУТ©ОпУРР§ФШєЙµЭЛНґ©№э BBB µЅґпДФКµЦКµДЧоРВ·ўХ№ЎЈЦШµгЅйЙЬБЛДФДЪЖ¤Пё°ыЙП±нґпµДлДј°ЖдМШТмРФКЬМеµД№±ПЧЎўТ©ОпУРР§ФШєЙµДСЎФсЎўPDCs µДЙијЖЎўДФЅшИл»ъЦЖТФј°ПтґуДФµДµЭЛНР§ВКµИРВЦЄК¶ЎЈ
ФЪЦРКаЙсѕПµНіТ©ОпїЄ·ўЦРЈ¬КµПЦЙсѕЦОБЖТ©Опґ©№эСЄДФЖБХПЈЁBBBЈ©ИФИ»КЗТ»ёцѕЮґуµДМфХЅЈ¬УИЖдКЗ¶ФУЪµ°°ЧЦКєНєЛЛбЦОБЖТ©Оп¶шСФЎЈСЄДФЖБХПј«ґуµШПЮЦЖєНїШЦЖОпЦКЅшИлґуДФµДБч¶ЇЈ¬Іўідµ±ЧЕґуДФµДКШГЕФ±ЅЗЙ«Ј¬ґУ¶шФцјУБЛТ©ОпіЙ№¦µЅґпґуДФµДДС¶ИЎЈѕЭ№АјЖЈ¬Ц»УР2%µД·З»оРФНвЕЕЧЄФЛµ°°ЧµЧОпЎўЗЧЦ¬РФРЎ·ЦЧУЙсѕЦОБЖТ©ОпЈЁ·ЦЧУБї< 500 DaЈ¬LogPЅУЅь2Ј©ДЬ№»ґ©№эСЄДФЖБХПІўµЅґпЦРКаЙсѕПµНіЦОБЖ°РµгЈ»і¬№э98%µДРЎ·ЦЧУєНјёєх100%µДґу·ЦЧУТ©ОпЈ¬°ьАЁлДАаЎўЦШЧйµ°°ЧЎўµҐБґї№МеЎў»щУЪRNAёЙИЕЈЁRNAiЈ©µДТ©ОпєН»щТтБЖ·ЁЈ¬¶јІ»·ыєПґ©НёСЄДФЖБХПµДТЄЗуЎЈХвј«ґуµШЧи°БЛХвР©»ЇєПОпіЙОЄУРР§µДЦРКаЙсѕПµНіЦОБЖТ©ОпµДЗ±Б¦ЎЈ
ОЄБЛїЛ·юПтґуДФКдЛНТ©ОпµДХП°Ј¬ТСѕїЄ·ўіцБЛ¶аЦЦІЯВФАґПтЦРКаЙсѕПµНіКдЛНЙсѕЦОБЖТ©ОпЎЈЦРКаЙсѕПµНіТ©ОпКдЛНІЯВФЦчТЄ·ЦОЄБЅАаЈєЗЦИлРФєН·ЗЗЦИлРФТ©ОпКдЛНЎЈЗЦИлРФіМРтЦјФЪОпАнґ©НёСЄДФЖБХПЈ¬АэИзНЁ№эКЦКхЦІИлДФКТµј№ЬЈ¬И»єуЅ«Т©ОпЧўИлДФКТЗ»Ј¬НЁ№эЗКДЪЧўЙдПтСь¶ОЦлНшД¤ПВЗ»ёшТ©Ј¬НЁ№эѕ±¶ЇВц№аЧўёЯЙшИЬТєИзёКВ¶ґј»тѕ±¶ЇВц¶ЇВц№аЧўСЄ№Ь»оРФОпЦКИз»єј¤лД»т»єј¤лДАаЛЖОпАґФЭК±ґтїЄСЄДФЖБХПЈ¬АыУГОўЕЭѕЫЅ№і¬ЙщФЭК±ЖЖ»µСЄДФЖБХПЈ¬»тХЯФЪДФІїЦІИл»ЇС§ЦОБЖѕЫєПОп±ЎЖ¬ЎЈХвР©іМРт¶Ф»јХЯІ»УСєГЈ¬ІўЗТґжФЪЗ±ФЪµДЅЎїµ·зПХЎЈ·ЗЗЦИлРФ·Ѕ·ЁЦјФЪФЪІ»ЖЖ»µСЄДФЖБХПОпАнНкХыРФµДЗйїцПВґ©НёСЄДФЖБХПЎЈХвР©·Ѕ·Ё°ьАЁАыУГСЄДФЖБХПЦРУлЙъОпС§Па№ШµДДЪФґРФЧЄФЛ»ъЦЖЈ¬ИзКЬМеЅйµјЎўОьёЅЅйµјєНФШМеЅйµјµДЧЄФЛПµНіЈ¬ЅьДкАґХвР©ПµНіФЪУЄСшОпЦКЎўј¤ЛШєНЧЄФЛµ°°Ч·ЅГжµГµЅБЛєЬєГµДСРѕїЈ»»тХЯНЁ№э»ЇС§РЮКОТ©ОпЈ¬АыУГЦ¬ЦК»ЇјјКхЅ«Т©ОпУлі¤БґЦ¬·ѕЛб№ІјЫБ¬ЅУЈ¬ТФФцјУТ©ОпНЁ№эСЄДФЖБХПµДНЁµАЈ¬ґУ¶шМбёЯТ©ОпµДЦ¬ЦКИЬЅвРФєНСЄДФЖБХПµДЙшНёРФЎЈ
Т»ЦЦєЬУРЗ°ѕ°µД·ЗЗЦИлРФІЯВФКЗК¶±рСЄДФЖБХПґ©НёлД»тСЄДФЖБХПґ©Луµ°°ЧЧчОЄДФІїµЭЛН·ЦЧУФШМеЈ¬ТФФцјУґ©№эСЄДФЖБХПµДФЛКдЎЈ·ЦЧУІЯВФЦјФЪНЁ№эЅ«ЦОБЖјБ»ЇС§Б¬ЅУµЅСЄДФЖБХП°РПтЕдМе»тСЄДФЖБХПґ©Луµ°°ЧЙПЈ¬ґУ¶шЅЩіЦСЄДФЖБХПµДДЪФґРФБчИлФЛКдЈ¬ёГЕдМе»тґ©Луµ°°ЧУлЖдКЬМеЅбєПЈ¬ґҐ·ўЧЄ°ыНМЧчУГІўЅ«Т©ОпФЛКдµЅСЄДФЖБХПµДЗ»ГжЎЈСЄДФЖБХПФЛКдлДєНСЄДФЖБХПґ©Луµ°°Ч-Т©Оп№№ЅЁМеУРїЙДЬЖЫЖґуДФФКРнХвР©·ЦЧУНЁ№эЈ¬ґУ¶шґЩЅшТ©ОпЅшИлДФКµЦКЎЈґУЦРКаЙсѕПµНіТ©ОпµЭЛНµДЅЗ¶ИАґїґЈ¬ХвЦЦКЬМеЅйµјµДФЛКдКЗТ»ЦЦ·ЗіЈУРОьТэБ¦µДІЯВФЈ¬ТтОЄАнВЫЙПЈ¬ёчЦЦ·ЦЧУЦКБї»тОпАн»ЇС§РФЦКµД·ЦЧУ»хОпЈ¬°ьАЁРЎ·ЦЧУєНЙъОпЦЖјБЈ¬¶јїЙТФНЁ№эХвТ»№эіМЅшРРФЛКдЎЈСЄДФЖБХПґ©Луµ°°Ч-Т©Оп№№ЅЁМе»тДФЙшНёлД-Т©ОпЕјБЄОпЈЁPDCЈ©ТСіЙОЄТ»ЦЦєЬУРЗ°ѕ°µД·ЗЗЦИлРФТ©ОпµЭЛН·Ѕ·ЁЦ®Т»ЎЈ
ФЪСЄДФЖБХПЈЁBBBЈ©µДґ«µЭ·ЅГжЎЈУл·ЦЧУБїі¬№э 150 З§µА¶ы¶ЩЗТСЄДФЖБХПґ©НёДЬБ¦УРПЮµДї№МеТ©ОпЕјБЄОпЈЁADCЈ©Па±ИЈ¬лДТ©ОпЕјБЄОпЈЁPDCsЈ©·ЦЧУБїёьµНЈ¬ТЧУЪґу№жДЈµНіЙ±ѕєПіЙЈ¬ІўЗТФКРн¶Ф°±»щЛбЅшРРРЮКОЈ¬ТФФцЗїФЪС»·ЦРµДОИ¶ЁРФєНСЄДФЖБХПµДЙшНёРФЎЈ
2 | СЄДФЖБХПКЗЦРКаЙсѕПµНіТ©ОпµЭЛНµДЦчТЄХП°
ґуДФКЗТ»ёц·ЗіЈёґФУµДЦШТЄЖч№ЩЈ¬°ьє¬ёчЦЦАаРНµДДФПё°ыЈ¬ГїЦЦПё°ы¶јУРІ»Н¬µД№¦ДЬЎЈЙсѕФЄКЗµзРЛ·ЬПё°ыЈ¬УГУЪґ«µЭРЕПўЈ¬ДЪЖ¤Пё°ыРОіЙДФСЄ№ЬЈ¬РЗРОЅєЦКПё°ыОЄЙсѕФЄ№¦ДЬМṩ֧іЦЈ¬РЎЅєЦКПё°ыКЗґуДФµДіЈЧ¤ѕЮКЙПё°ыЈ¬ёєФрґуДФµДГвТЯјаКУЎЈЖдЛыґуДФѕЮКЙПё°ыЈ¬ИзСЄ№ЬЦЬО§ѕЮКЙПё°ыЈ¬О»УЪСЄ№ЬНв±нГжЈ¬ІўФЪДФКµЦКУлС»·Ц®јдµДЅзГжЦґРР»о¶ЇЈЁНј1єННј2Ј©ЎЈИЛАаґуДФУµУРТ»ёці¬№э400УўАпі¤µДСЄ№ЬНшВзЈ¬°ьАЁґуДФ¶ЇВцЎўРЎ¶ЇВцєНГ«ПёСЄ№ЬЎЈјёєхГїёцЙсѕФЄ¶јУРЧФјєµДґуДФГ«ПёСЄ№Ь№©СЄЎЈ
ЦРКаЙсѕПµНіТ©ОпФЪґуДФЦРµДґ«µЭґжФЪИэёцЦчТЄХП°ЎЈКЧПИЈ¬СЄДФЖБХПЈЁBBBЈ©КЗСЄ№ЬУлДФКµЦКЦ®јдµДТ»µАЙъАнЖБХПЈЁНј1Ј©ЎЈЖдґОЈ¬СЄДФј№ТєЖБХПЈЁBCSFBЈ©КЗСЄТєС»·УлДФј№ТєС»·Ц®јдµДЖБХПЎЈBCSFBУЙТФПВЅб№№РОіЙЈє
ВцВзґФЅфГЬБ¬ЅУµДЙПЖ¤Пё°ыГжПтСЄДФј№ТєЎЈµЪИэЈ¬ОЮСЄ№ЬµДЦлНшД¤ЖБХПО»УЪУІД¤ПВЈ¬ІўНкИ«·в±ХБЛЦРКаЙсѕПµНіЈ¬ФЪЦРКаЙсѕПµНіТ©ОпПтґуДФµДЧЄФЛЦР·ў»УЧЕЦШТЄЧчУГЎЈФЪХвР©ЖБХПЦРЈ¬СЄДФЖБХПКЗЧоЦШТЄµДЖБХПЈ¬¶ФДФПё°ыµДЦ±ЅУОў»·ѕіїШЦЖЧоОЄСПёсЎЈ
2.1 | СЄДФЖБХПµДМШРФ
СЄДФЖБХПКЬµЅСПёсµчЅЪЈ¬Ц»ФКРн±ШРиµДАлЧУЎўУЄСшОпЦКєНј¤ЛШЅшИлґуДФЈ¬ІўЧиЦ№їЙДЬУРє¦µДСЄФґРФНвФґРФОпЦКЅшИлґуДФЎЈСЄДФЖБХПКЗУЙДЪЖ¤Пё°ыЧйіЙµДБ¬РшІгЈ¬ЕЕБРФЪґуДФГ«ПёСЄ№ЬДЪЈ¬ЦЬО§°ь№ьЧЕ»щµЧД¤ЎўЦЬПё°ыєНРЗРОЅєЦКПё°ыµДЦХЧгЎЈДФОўСЄ№ЬДЪЖ¤Пё°ыЦ®јдѕЯУРМШКвµДЅфГЬБ¬ЅУЈ¬СЄ№ЬЗ»НкИ«·в±ХЈЁНј 1Ј©ЎЈРЗРОЅєЦКПё°ыЎўЦЬПё°ыЎўДЪЖ¤Пё°ыЎў»щµЧД¤ЎўРЎЅєЦКПё°ыєНЙсѕФЄ¶јФЪСЄДФЖБХПµДНкХыРФЦР·ў»УЧЕ№ШјьЧчУГЈЁНј 1Ј©ЎЈ
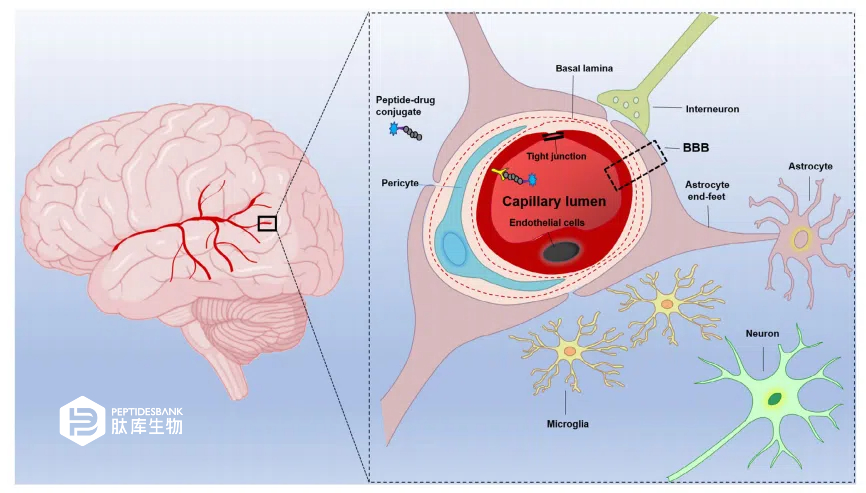 Нј1 СЄДФЖБХПЈЁBBBЈ©єНЖдЛыЙсѕСЄ№ЬµҐФЄЧйјюµДКѕТвНјЎЈСЄДФЖБХПКЗУЙБ¬РшµДДФДЪЖ¤Д¤ЧйіЙµДПё°ы¶аЖБХПЈ¬НЁ№эЅфГЬБ¬ЅУЎўЦЬПё°ыєНРЗРОЅєЦКПё°ыЧгД©ЅфГЬ·в±ХЎЈСЄДФЖБХПЅ«ДФКµЦКУлНвЦЬСЄС»·ёфАлїЄАґЈ¬ІўЧчОЄЦРКаЙсѕПµНіУлНвЦЬЦ®јдµДСЄ№ЬЅзГжЈ¬µчЅЪБЅХЯЦ®јдµДНЁРЕЎЈДФґ©НёСЄДФЖБХПґ©ЛулДЕјБЄТ©ОпїЙТФНЁ№эАыУГДЪФґРФФЛКд·ЦЧУ»ъЦЖЈ¬ґ©№эСЄДФЖБХПЈ¬µЅґпДФКµЦКЎЈ
Нј1 СЄДФЖБХПЈЁBBBЈ©єНЖдЛыЙсѕСЄ№ЬµҐФЄЧйјюµДКѕТвНјЎЈСЄДФЖБХПКЗУЙБ¬РшµДДФДЪЖ¤Д¤ЧйіЙµДПё°ы¶аЖБХПЈ¬НЁ№эЅфГЬБ¬ЅУЎўЦЬПё°ыєНРЗРОЅєЦКПё°ыЧгД©ЅфГЬ·в±ХЎЈСЄДФЖБХПЅ«ДФКµЦКУлНвЦЬСЄС»·ёфАлїЄАґЈ¬ІўЧчОЄЦРКаЙсѕПµНіУлНвЦЬЦ®јдµДСЄ№ЬЅзГжЈ¬µчЅЪБЅХЯЦ®јдµДНЁРЕЎЈДФґ©НёСЄДФЖБХПґ©ЛулДЕјБЄТ©ОпїЙТФНЁ№эАыУГДЪФґРФФЛКд·ЦЧУ»ъЦЖЈ¬ґ©№эСЄДФЖБХПЈ¬µЅґпДФКµЦКЎЈ
УлНвЦЬµДДЪЖ¤Пё°ыІ»Н¬Ј¬ДФДЪЖ¤Пё°ыИ±·¦ґ°їЧЈЁФКРнСЄТєєНЧйЦЇЦ®јдїмЛЩЅ»»»·ЦЧУµДїЧП¶Ј©Ј¬ґУ¶шПЮЦЖБЛ·ЦЧУµДБч¶ЇЎЈЅфГЬБ¬ЅУµДµДґжФЪПЮЦЖБЛПаБЪДЪЖ¤Пё°ыЦ®јд·ЦЧУµДБчИлЈЁПё°ыЕФНѕѕ¶Ј»Нј2Ј©ЎЈЅб№№ЙПЈ¬ПаБЪДФДЪЖ¤Пё°ыЦ®јдµДЅфГЬБ¬ЅУРОіЙБЛТ»ёцБ¬РшµДНшВзЈ¬УЙЖЅРРµДЎўП໥Б¬ЅУµДїзД¤єНПё°ыЦКµ°°ЧБґЧйіЙЈ¬ТФ·в±ХПё°ыЕФјдП¶Ј¬ґУ¶шЧчОЄПё°ыЕФГЕПЮЦЖЗЧЛ®РФ·ЦЧУµДПё°ыЕФБч¶ЇЎЈХвР©МШРФК№µГДФОўСЄ№ЬДЬ№»РОіЙТ»ІгЅфГЬµДПё°ыІгЈ¬ЖдїзДЪЖ¤µзЧиЈЁTEERЈ©і¬№э1000 ¦ёcm 2ЎЈСЄДФЖБХПЈЁBBBЈ©ПФКѕДЪЖ¤Пё°ы¶ФТєµОµДНМКЙЧчУГЈЁјґ»оПё°ыЙгИЎТєµОµД№эіМЈ©ґуФјјхЙЩБЛ 100 ±¶ЎЈ
Нј 2 СЄДФЖБХПЈЁBBBЈ©·ЦЧУФЛКдµДЦчТЄВ·ПЯµДКѕТвНјЎЈ·ЦЧУґ©№эСЄДФЖБХПµДЦчТЄНѕѕ¶°ьАЁ±»¶ЇА©ЙўЈЁПё°ыЕФєНїзПё°ыНѕѕ¶Ј©ЎўФШМеЅйµјµДПё°ыДЪНМЈЁCMTЈ©ЎўКЬМеЅйµјµДПё°ыДЪНМЈЁRMTЈ©ЎўОьёЅЅйµјµДПё°ыДЪНМЈЁAMTЈ©єНПё°ыЅйµјµДПё°ыДЪНМЎЈСЄДФЖБХПЙП±нґпБЛТ©ОпНвЕЕЧЄФЛµ°°ЧЈ¬ТФПЮЦЖРн¶а·ЦЧУЅшИлґуДФЎЈДФґ©НёСЄДФЖБХПґ©ЛулДєНлД-Т©ОпЕјБЄОпЈЁPDCsЈ©їЙТФАыУГСЄДФЖБХПЙП№э¶И±нґпµДКЬМе»тЧЄФЛµ°°ЧЈ¬НЁ№эКЬМеЅйµјµДПё°ыДЪНМЅйµјТ©Опґ©№эСЄДФЖБХПЎЈ
СЄДФЖБХПІў·ЗКЗѕІМ¬µДЈ¬¶шКЗёЯ¶И¶ЇМ¬µДЎЈДЪЖ¤Пё°ыЎўРЗРОЅєЦКПё°ыЎўЦЬПё°ыєНУлСЄДФЖБХПГЬЗРЅУґҐµДЙсѕФЄЈЁЙсѕСЄ№ЬµҐО»»тNVUЈ©Ц®јдµДёчЦЦПё°ыДЪєНПё°ыјдРЕєЕКВјюїШЦЖЧЕ·ЦЧУµДНЁНёРФЈЁНј1Ј©ЎЈДЪЖ¤Пё°ыЗ»ІаЈЁСЄТєІаЈ©єНД¤НвЈЁґуДФІаЈ©Д¤ЙПµДМШКвµ°°ЧЦКµчЅЪЖПМСМЗµИґъР»ІъОпµДНЁ№эЈ¬ІўїШЦЖ·ЦЧУµДЅшіцЈЁНј2Ј©ЎЈСЄДФЖБХПСПёсїШЦЖНвЦЬГвТЯПё°ыЅшИлґуДФЎЈОўЅєЦКПё°ыЈЁЦРКаЙсѕПµНіЧ¤БфµДѕЮКЙПё°ыСЬЙъПё°ыЈ©µДґжФЪУРЦъУЪµчЅЪЦРКаЙсѕПµНіКµЦКЦРµДГвТЯ·ґУ¦ЎЈГвТЯПё°ыИзTБЬ°НПё°ыФЪЦРКаЙсѕПµНіЛрЙЛєНјІІЎЖЪјдїЙТФЅшИлґуДФЎЈТ»ПоРВµД·ўПЦ±нГчЈ¬ДіР©СЄЅ¬µ°°ЧПтґуДФµДФЛКдЛжДкБд¶ш±д»ЇЈ¬ґУЅЎїµДкЗбРЎКуµДЕдМеМШТмРФЎўКЬМеЅйµјµДЧЄ°ыНМЧчУГЧЄ±дОЄАПДкРЎКуµД·ЗМШТмРФРЎОСЙгИЎЎЈЛщУРХвР©МШХчК№µГСЄДФЖБХПідµ±БЛОпАнЖБХПЈЁЅфГЬБ¬ЅУЈ©ЎўЎ°ФЛКдЖБХПЎ±ЈЁМШ¶ЁµДФЛКдПµНіЈ©ЎўЎ°ґъР»ЖБХПЎ±ЈЁЧЁГЕµДГёПµНіЈ©ТФј°Ў°ГвТЯЖБХПЎ±ЎЈ
2.2 | ·ЦЧУґ©№эСЄДФЖБХПФЛКдµДНѕѕ¶
УЙУЪСЄДФЖБХПЈЁBBBЈ©µДЎ°ЅфГЬРФЎ±Ј¬·ЦЧУµДЕФВ·Пё°ыФЛКдЧоРЎ»ЇЈ»Т©ОпєН±ШРи·ЦЧУЦчТЄНЁ№э±»¶ЇА©ЙўЎўФШМеЅйµјµДФЛКдЎўКЬМеЅйµјµДЧЄ°ыНМСКЎўОьёЅЅйµјµДЧЄ°ыНМСК»тНЁ№эПё°ыЅйµјµДЧЄ°ыНМСКµДЧчУГЅшИлґуДФЈЁНј2Ј©ЎЈЧЄ°ыНМСККЗТ»ёц№эіМЈ¬НЁ№эД¤°ьДТЅ«·ЦЧУґУПё°ыµДТ»ІаЧЄТЖµЅПё°ыµДДЪІїЈ¬И»єуЧЄТЖµЅПё°ыµДБнТ»ІаЎЈПа¶ФУЪНвЦЬДЪЖ¤Пё°ыЈ¬ДФДЪЖ¤Пё°ы±нПЦіцПа¶ФЅПµНµДДТЕЭФЛКдЛЩВКЎЈ
2.2.1 | ±»¶ЇА©Йў
±»¶ЇА©ЙўКЗЦё»ЇєПОпФЪЕЁ¶ИМЭ¶ИµД·ЅПтЙПТЖ¶ЇЈ¬ОЮРиДЬБїЈ¬ЗТІ»їЙ±ҐєНЎЈјтµҐА©ЙўКЗґу¶аКэЗЧЦ¬РФРЎ·ЦЧУЦРКаЙсѕПµНіТ©ОпµДЦчТЄЅшИл»ъЦЖЎЈЧоґуПё°ы-Г«ПёСЄ№ЬѕаАлОЄ20ОўГЧЈ¬РЎ·ЦЧУїЙТФФЪ°лГлДЪЙшНёЎЈОпЦКЅшИлґуДФµДА©ЙўїЙТФ·ЦОЄПё°ыЕФА©ЙўєНПё°ыДЪА©ЙўЈЁНј2Ј©ЎЈПё°ыЕФА©ЙўКЬµЅДФДЪЖ¤Пё°ыЦ®јдЅфГЬБ¬ЅУµДПЮЦЖЎЈПё°ыДЪА©ЙўЦчТЄИЎѕцУЪ·ЦЧУµДЙшНёРФЎЈРЎЗЧЦ¬РФОпЦКИзѕЖѕ«єНАа№Мґјј¤ЛШїЙТФНЁ№э·З±ҐєНА©ЙўЅшРРПё°ыДЪЙшНёЎЈёЯ¶ИЦ¬ИЬРФТ©ОпµШИыГЧЛЙїЙТФЗбТЧµШґ©№эСЄДФЖБХПЈ¬ІъЙъХтѕІЧчУГЎЈТ»°гАґЛµЈ¬РЎ·ЦЧУБїЈЁ<500 µА¶ы¶ЩЈ©ЎўИхЗвјьЈЁЙЩУЪ 6 ёцЗвјьЈ©ЎўЗЧЦ¬РФЈЁLogP > 2Ј©ТФј°І»ґжФЪЧФУЙРэЧЄјьєНј«РФ±нГж»эЈЁPSAЈ©< 60 ЦБ 70 ЖЅ·Ѕ°ЈЈ¬¶јУРАыУЪНЁ№эА©Йўґ©№эСЄДФЖБХПЎЈФЪІвБїНЁ№э±»¶ЇА©ЙўµДТ©ОпЙшНёРФК±Ј¬ЛщУРХвР©ІОКэ¶јКЗЧЫєПїјВЗµДЈ¬¶шІ»КЗµҐ¶АїјВЗЎЈ¶ФУЪМмИ»лДЈ¬іэ·ЗлДѕЯУРБЅЗЧРФЅб№№»тНЁ№эєПіЙ·Ѕ·ЁК№ЖдѕЯУРЗЧЦ¬РФЈ¬·сФт±»¶ЇА©Йў·ЗіЈУРПЮЎЈ
2.2.2 | ФШМеЅйµјµДПё°ыДЪНМЧчУГ
±ШРиУЄСшЛШИзЖПМСМЗЎў°±»щЛбєНєЛЬХЛбНЁ№эёЯ¶ИСЎФсРФєНБўМеМШТмРФЧЄФЛµ°°ЧЈ¬НЁ№эФШМеЅйµјµДїзПё°ыФЛКдЈЁCMTЈ©»ъЦЖґ©№эСЄДФЖБХПЎЈЧЄФЛµ°°ЧФЪЗ»ІаК¶±рµЧОпЈ¬И»єуµЧОп-ЧЄФЛµ°°ЧЅбєППµНіґҐ·ўФШМеµ°°ЧґУНвПтµЅДЪПтЧґМ¬µД№№Пу±д»ЇЈ¬µјЦВµЧОпНЁ№эЕЁ¶ИМЭ¶ИПтД¤µДЗ»НвІаФЛКдЈЁНј2Ј©ЎЈИз№ы»ЇєПОпРиТЄДжЕЁ¶ИМЭ¶ИТЖ¶ЇЈ¬ATPїЙДЬМṩДЬБїАґґЩЅшХвТ»№эіМЎЈґуДФДЪЖ¤Пё°ыД¤ЦРУРјёёцФЛКдПµНіУГУЪУЄСшОпЦКєНДЪФґРФ»ЇєПОпµДФЛКдЈ¬АэИзGLUT1ЖПМСМЗЧЄФЛµ°°ЧУГУЪЖПМСМЗєНДіР©јєМЗµДЧЄФЛЈ¬ЛбРФ°±»щЛбЧЄФЛПµНіУГУЪ№И°±ЛбєНМ춬°±ЛбµДЧЄФЛЈ¬ТФј°LAT1ґуЦРРФ°±»щЛбЧЄФЛµ°°ЧУГУЪ±Ѕ±ы°±ЛбєНЖдЛыЦРРФ°±»щЛбµДЧЄФЛЎЈLAT1»№ЧЄФЛТ»Р©Т©ОпЈ¬ИзL-¶а°Н°·ЎўјУ°НЕз¶Ў»тГА·ЁВШЈ¬ХвР©Т©ОпУлДЪФґРФµЧОпѕЯУРПаЛЖµД·ЦЧУЅб№№єНґуРЎЎЈ
2.2.3 | КЬМеЅйµјµДЧЄ°ыНМЧчУГ
ДФГ«ПёСЄ№ЬДЪЖ¤Пё°ыѕЯУРКЬМеЅйµјµДЧЄ°ыНМЧчУГЈЁRMTЈ©»ъЦЖЈ¬ЧчОЄМШ¶ЁєНСЎФсРФЙгИЎДіР©ґу·ЦЧУµДКЦ¶ОЎЈТ»°гАґЛµЈ¬С»·ЦРО»УЪДФДЪЖ¤Пё°ыЗ»ІаµДЕдМеУлЖдМШТмРФКЬМеЅбєПЈ¬КЬМе-ЕдМеёґєПОпНЁ№эДТЕЭФЛКд»ъЦЖґ©№эПё°ыДЪКТЈ¬НЁіЈНЁ№эДЪМе/ИЬГёМеПµНіЎЈИ»єуЈ¬КЬМе-ЕдМеёґєПОпЅвАлЈ¬ЕдМеґУёґєПОпЦРКН·ЕµЅДЪЖ¤Пё°ы·ЗЗ»ІаµДПё°ыНвїХјдЈ¬КЬМе±»»ШКХЦБДЪЖ¤Пё°ыµДЗ»ІаЈЁНј2Ј©ЎЈСЄТєЦРµДДіР©ґу·ЦЧУНЁ№эТСід·Ц±нХчµДДЪФґРФКЬМеЈЁИзТИµєЛШЎўЧЄМъµ°°ЧєНКЭЛШЈ©ґ©№эСЄДФЖБХПЎЈ№ШУЪІ»Н¬µДЦРКаЙсѕПµНіјІІЎИзєОУ°ПмКЬМеµД±нґпј°ЖдµчЅЪЈ¬ДїЗ°БЛЅвУРПЮЎЈКЬМеЅйµјµДЧЄ°ыНМЧчУГУРїЙДЬ±»АыУГЧчОЄПтґуДФµЭЛНЦОБЖРФ·ЦЧУµДІЯВФЎЈ
2.2.4 | ОьёЅЅйµјµДПё°ыДЪНМЧчУГ
ОьёЅЅйµјµДЧЄ°ыНМСКЈЁAMTЈ©КЗТ»ЦЦ·ЗМШТмРФµДЧЄ°ыНМСКЈ¬ЛьКЗУЙДіР©ґу·ЦЧУµДґшХэµзєЙІї·ЦУлє¬УРТхАлЧУТТхЈёОЛШµ°°ЧѕЫМЗµДДФДЪЖ¤Пё°ыД¤Ц®јдµДѕІµзП໥ЧчУГЛщґҐ·ўµДЎЈAMTµД№эіМ°ьАЁѕІµзёґєПОпµДДЪПЭЈ¬ЛжєуРОіЙДЪєМеЈ¬ґУЗ»ІаФЛКдµЅ·ЗЗ»ІаЈ¬ІўКН·ЕAMTµЧОпЎЈФЪAMTґЩЅшµДЦРКаЙсѕПµНіТ©ОпµЭЛНІЯВФЦРЈ¬НЁіЈК№УГ¶аСфАлЧУµ°°ЧЈ¬ИзУгѕ«µ°°Ч»тє¬УРСфАлЧУРтБРµДПё°ыґ©НёлДЈ¬ИзSynBлДЈЁНј2Ј©ЎЈУРИ¤µДКЗЈ¬µјЦВCOVID-19±¬·ўµДSARS-CoV-2ІЎ¶ѕµД1РНµ°°ЧїЙТФНЁ№эТ»ЦЦАаЛЖУЪAMTµД»ъЦЖґ©№эРЎКуСЄДФЖБХПЎЈУлСфАлЧУµ°°Ч»тлДПа№ШµДЗ±ФЪИ±µгКЗЛьГЗµДЛж»ъ·ЦІјЈ¬ТтґЛИ±·¦¶ФґуДФµДСЎФсРФЈ¬ТФј°УлДЪЖ¤ЛрЙЛПа№ШµДЗ±ФЪ¶ѕРФЎЈ
2.2.5 | Пё°ыЅйµјµДЧЄ°ыНМЧчУГ
Пё°ыЅйµјµДЧЄ°ыНМСККЗЧоЅь·ўПЦµДТ»ЦЦТ©Опґ©№эСЄДФЖБХПЈЁBBBЈ©µДФЛКдНѕѕ¶ЎЈµҐєЛПё°ы±»ИПОЄКЗТ»ЦЦЎ°МШВеТБДѕВнЎ±Ј¬±»ІЎФМеУГЧчЅшИлЦРКаЙсѕПµНіµДЅ»НЁ№¤ѕЯЎЈТ»Р©ІЎФМеЈ¬ИзИЛАаГвТЯИ±ПЭІЎ¶ѕЈЁHIVЈ©Ј¬НЁ№эХвЦЦФЛКдНѕѕ¶ЅшИлґуДФЈЁНј2Ј©Ј¬ЖдЦРёРИѕHIVµДµҐєЛПё°ыєН/»тѕЮКЙПё°ыФЪХэіЈЗЁТЖ№эіМЦРґ©№эСЄДФЖБХПЅшРРГвТЯјаКУЈ¬»тХЯУЙУЪІъЙъґЩСЧЅйЦКёД±дСЄ№ЬНЁНёРФЎЈРн¶аЙсѕПµНіјІІЎЈ¬Из°ў¶ыґДєЈД¬ІЎЎўЕБЅрЙІЎЎўДФБцєН°¬ЧМІЎПа№ШіХґфЈ¬¶јУРСЧЦўіЙ·ЦЎЈФЪСЧЦў№эіМЦРЈ¬№г·єХРДј°ЧПё°ыЈЁµҐєЛПё°ыєНЦРРФБЈПё°ыЈ©ЎЈХвР©Пё°ыНЁ№э±»іЖОЄПё°ыНвЙшєНЗч»ЇЧчУГµД№эіМПтСЧЦўІїО»ЗЁТЖЈ¬ХвКЗ¶АМШµДЎЈПё°ыЅйµјµДЧЄ°ыНМСКТС±»їЄ·ўіЙТ»ЦЦБоИЛРЛ·ЬµДІЯВФЈ¬УГУЪКµПЦСЄДФЖБХПµДЦОБЖРФТ©ОпµЭЛНЎЈ»щУЪГвТЯПё°ыЎўЙсѕёЙПё°ыєНјдідЦКёЙПё°ыµДДФБцЦОБЖТ©ОпФШМеТС±»МЅЛчЎЈ
2.2.6 | »оРФНвЕЕЧЄФЛ
СЄДФЖБХПµДГ«ПёСЄ№ЬДЪЖ¤Пё°ыЙП±нґпЧЕТ©ОпНвЕЕЧЄФЛµ°°ЧЈ¬ХвПЮЦЖБЛРн¶аТ©ОпЅшИлґуДФµДДЬБ¦ЎЈХвР©НвЕЕЧЄФЛµ°°ЧДЬ№»Ѕ«Т©Оп±ГіцПё°ыЈ¬ґУ¶шНЁ№эЅ«ЛьГЗЕЕіэФЪґуДФДЪЖ¤Пё°ыЦ®НвЈ¬¶ФЅшИлСЄДФЖБХПРОіЙБЛ¶оНвµДПЮЦЖЈЁНј2Ј©ЎЈґу¶аКэХвР©НвЕЕЧЄФЛµ°°ЧКфУЪATPЅбєПєРі¬јТЧеµ°°ЧЈ¬°ьАЁP-МЗµ°°ЧЈЁP-gpЈ©Ўў¶аТ©ДНТ©Па№Шµ°°ЧЈЁMRPЈ©єНИйПЩ°©ДНТ©µ°°ЧЎЈЧЬЦ®Ј¬СЄДФЖБХПСПёсµчЅЪЧЕ±ШРиОпЦКєНРЕПў·ЦЧУЈЁИзлДЈ©µДБчИлєНБчіцЈ¬ІўЧчОЄЦРКаЙсѕПµНіУлСЄТєЦ®јдµДНЁРЕЅУїЪЎЈ
3 | СЄДФЖБХПґ©ЛулДµД·ўПЦ
3.1 | УГУЪДФІїµЭЛНµД BBB ґ©ЛулД
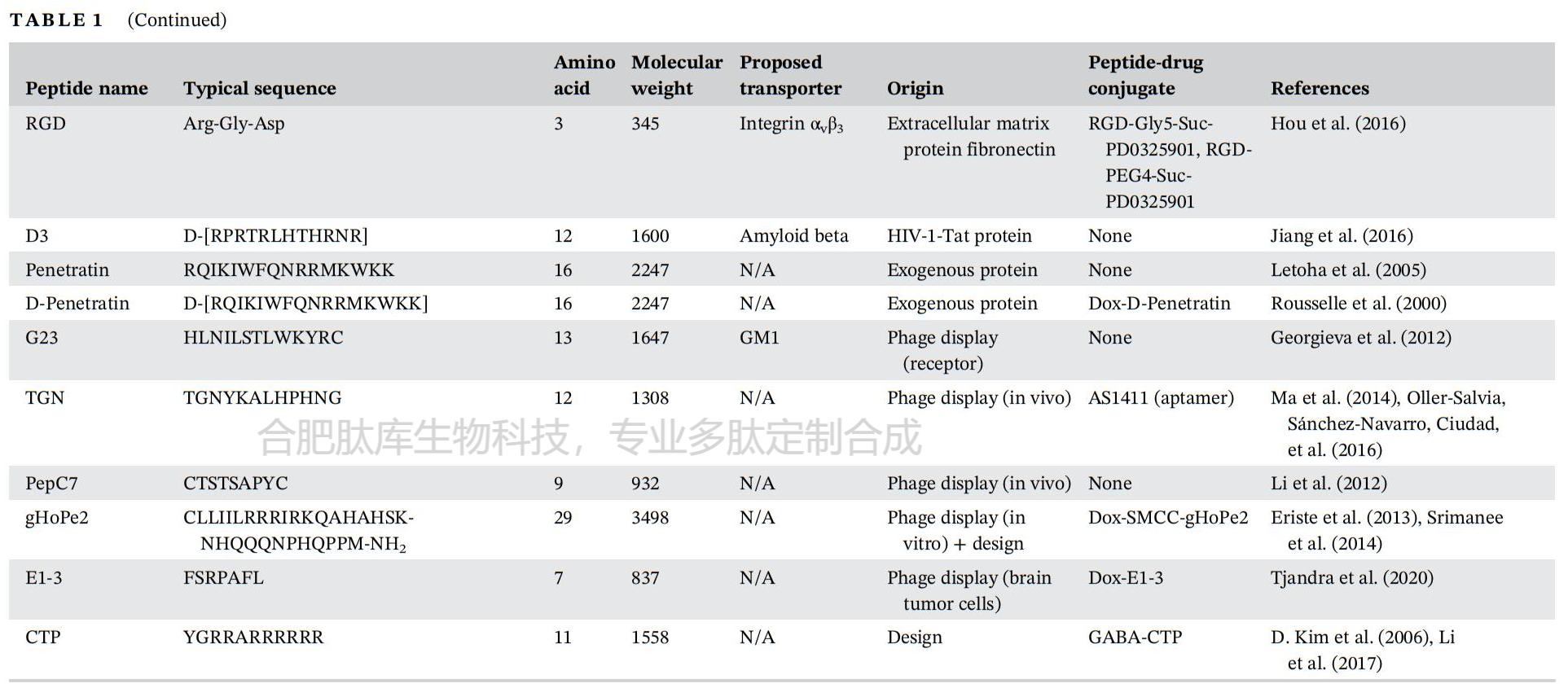
І»¶ПФці¤µДСРѕїРЛИ¤ТЄЗу·ўПЦДЬ№»Ѕ«ЙсѕЦОБЖ»хОпЈЁИзµ°°ЧЦКЎў»щТтєНТ©ОпЈ©ФЛКдµЅДФКµЦКµДДФ№йіІєНДФґ©Нё·ЦЧУФШМеЎЈЖдЦРЈ¬СЄДФЖБХПЈЁBBBЈ©ґ©НёлД»тСЄДФЖБХПґ©ЛулДТтЖдТЧУЪєПіЙєНРЮКОЎўГвТЯФРФЅПµНТФј°УлѕµдµДСЄДФЖБХПґ©Луµ°°ЧЈЁИзМШВеТБВнї№МеЈ©Па±ИіЙ±ѕЅПµН¶шКЬµЅ№ШЧўЎЈЅьДкАґЈ¬є¬УР¶аґп50ёц°±»щЛбІР»щµДПЯРФ»т»·лДТС±»·ўПЦЧчОЄСЄДФЖБХПґ©ЛулДЈ¬Іў±»УГЧчФШМе»т°РПтФЄЛШЅ«Т©ОпµЭЛНЦБґуДФЎЈХвР©лДѕЯУРТ»Р©№ІН¬МШХчЈ¬°ьАЁЈЁaЈ©¶ФМШ¶ЁКЬМеµДЗїЗЧєНБ¦Ј¬ХвР©КЬМеНЁіЈФЪґуДФСЄ№ЬµДЗ»Іа±нґпЈ¬ТФґҐ·ўДЪ»ЇЈ»ЈЁbЈ©ЅйµјПё°ыНМСКµДДЬБ¦Ј»ТФј°ЈЁcЈ©ДЬ№»ТФ·ЗЗЦИлРФµД·ЅКЅґЩЅш»хОпФЛКдµЅДФКµЦКЈ¬¶шІ»У°ПмСЄДФЖБХПµДНкХыРФЈЁНј2Ј©ЎЈТ»Р©СЄДФЖБХПґ©ЛулДґ©НёСЄДФЖБХПЈ¬¶шІ»РиТЄКЬМеЎЈХвЦЦлДЦчТЄКфУЪПё°ыґ©НёлДЈЁCPPЈ©јТЧеЈ¬УЙ¶МµДБЅЗЧРФєН/»тСфАлЧУРтБРЧйіЙЈ¬НЁ№э·ЗАлЧУМе»ъЦЖЈЁAMT »ъЦЖЈ©ѕЯУРёЯ¶Иґ©НёПё°ыД¤µДДЬБ¦ЎЈСЄДФЖБХПґ©ЛулДЦчТЄФґЧФКИЙсѕµДДЪФґРФµ°°ЧЦКЎўЙсѕ¶ѕЛШЎўДіР©ІЎ¶ѕТФј°ДЪФґРФлДЈ¬»тХЯНЁ№эКЙѕъМеХ№КѕЙъОпЙёСЎєНРЮКО¶ш·ўПЦЎЈ
КЙѕъМеХ№КѕјјКхЈЁPDTЈ©УЙG.P.К·ГЬЛ№ЈЁ1985ДкЈ©ТэИлЈ¬ОЄ·ўПЦЙъОп»оРФлДєНї№МеґтїЄБЛТ»ЙИГЕЎЈЛьКЗЙъІъґуБїІ»Н¬лДєНµ°°ЧЦКЈЁ¶аґп10-10)Ј©ТФј°МеНвєНМеДЪјш¶ЁУлЙъОп°Р±кЅбєПµДЕдМеµДУРјЫЦµ№¤ѕЯЎЈКЙѕъМеХ№КѕјјКхµД»щ±ѕФАнКЗµ°°ЧЦК±нРНЈЁХ№КѕµДлДЈ¬НЁіЈМШТмРФµШУлёРРЛИ¤µД°Р±кЅбєПЈ©УлЖдПаУ¦µД»щТтРНЈЁ±аВлёГµ°°ЧЦКµДDNAЈ»ЕБК©їЛЈ¬2006ДкЈ©Ц®јдµДОпАнБЄПµЎЈКЙѕъМеКЗТ»ЦЦПЛО¬ЧґµДІЎ¶ѕЈ¬ёРИѕПёѕъЛЮЦчЎЈКЙѕъМеХ№КѕКЗЦёНЁ№эЅ«±аВлНвФґРФµ°°ЧЦКµД»щТтІеИлКЙѕъМе»щТтЧйµДТВїЗµ°°Ч»щТтЦРЈ¬К№µ°°ЧЦК»тлДАаЅбєПОпФЪКЙѕъМе±нГж±нґпµД№эіМЎЈНЁ№эЗЧєНСЎФсКЙѕъМеХ№КѕµДЛж»ъївФЪ№М¶Ё»Ї°Р±к»тМеДЪЧйЦЇЙПЈ¬ТІіЖОЄЙъОпЙёСЎЈ¬Аґјш¶Ёµ°°ЧЦК»тлДАаЅбєПОпЈЁНј3(a)Ј©ЎЈ·єЛШ»Ї№эіМµДЦчТЄІЅЦи°ьАЁЈє(a) КЙѕъМеївУл№М¶Ё»Ї°Р±кЈЁМеНвЙъОп·єЛШ»ЇЈ©»тКЙѕъМеЧўИл¶ЇОпЈЁМеДЪЙъОп·єЛШ»ЇЈ©µД·хУэЈ»(b) ЗеПґТФИҐіэ·ЗЅбєПКЙѕъМеЈ»(c) К№УГЛб»тёЯСОПґНСЅбєПОпЈ»(d) НЁ№эёРИѕЛЮЦчґуі¦ёЛѕъПё°ыАґА©ФцПґНСµДКЙѕъМеїЕБЈЈЁНј3(a)Ј©ЎЈЙъОп·єЛШ»ЇїЙТФФЪСЄЗеСщ±ѕЎў·ЦАлµДКЬМеЎўПё°ы±нГжКЬМе»тМШ¶ЁЖч№ЩЈЁМеДЪЈ©ЙПЅшРРЈ¬НЁіЈЅшРРИэµЅОеВЦТФ»сµГ¶Ф°Р±кѕЯУРЧоёЯЗЧєНБ¦µДлД»тµ°°ЧЦКЎЈЙъОп·єЛШ»ЇєуЈ¬НЁ№эКЙѕъМеDNAµДІвРтАґјш¶ЁлДРтБРЈЁНј3Ј©ЎЈЅУПВАґЈ¬НЁ№эУлВТРтлДЅшРР№¦ДЬ·ЦОцАґИ·ИП·ЦАллДµДМШТмРФЎЈЖдЛы·Ѕ·ЁЈ¬Из¶Ф№йіІКЙѕъМеµДГвТЯИѕЙ«»тХЯ¶ФКЙѕъМе»тєПіЙлДЅшРРУ«№в»т·ЕЙдРФ±кјЗЈ¬ТФИ·¶ЁЖдФЪЖч№ЩЧйЦЇЦРµДСЎФсРФ·ЦІјЎЈ
Нј 3 ФґЧФКЙѕъМеХ№КѕЙъОпЙёСЎјјКхєНЙсѕКИРФµ°°ЧµДСЄДФЖБХПґ©ЛулДµДКѕТвНјЎЈЈЁaЈ©Хл¶ФМеНв°Рµг»тМеДЪЧйЦЇµДКЙѕъМеХ№КѕЙъОпЙёСЎ№эіМЎЈКЙѕъМеОДївУлМеНв№М¶ЁФЪ±нГж»тМеДЪЧўЙдµЅ¶ЇОпМеДЪµД°РµгЅбєПЎЈѕ№э·хУэЈ¬ОґЅбєПµДКЙѕъМе±»ПґµфЈ¬ЅбєПµДКЙѕъМеНЁ№эПґНСКХјЇІўЅшТ»ІЅА©ФцЎЈЅшРР¶аВЦСЎФсТФё»јЇЗЧєНБ¦ЗїµДКЙѕъМеЎЈёЯЗЧєНБ¦µДКЙѕъМеЅбєПОпНЁ№э DNA ІвРтЅшРРґї»ЇєНІвРтЈ¬ТФИ·¶Ё°РµгЅбєПлДµДРтБРЎЈХвР©°РПтлДЅшТ»ІЅєПіЙЈ¬ІўФЪМеНвСЄДФЖБХП Transwell јмІвЦРЅшРРЖА№АЈ¬ТФјш¶ЁСЄДФЖБХПґ©ЛулДЎЈЈЁbЈ©јш¶ЁёєФрЙсѕКИРФµ°°Чґ©№эСЄДФЖБХПµДЖ¬¶ОЎЈЈЁcЈ©КЙѕъМеХ№КѕјјКхјш¶ЁіцХл¶ФРЎКуґуДФЦРµНГЬ¶ИЦ¬µ°°ЧКЬМеЈЁLDLRЈ©µДСЄДФЖБХПґ©ЛулД LRPep2ЎЈХ№КѕБЛ LRPep2 µДЅб№№ТФј°лД - µ°°ЧЦКЅбєПµДјЖЛг¶ФЅУЈЁ¶ҐНјЈ©Ј¬ТФј°ВЮµ¤Гч±кјЗµД LRPep2-rhoЈЁємЙ«Ј©Ул LDLRЈЁВМЙ«Ј©ФЪРЎКуДФЗРЖ¬ЙПµД№І¶ЁО»ЈЁµЧНјЈ©ЎЈ
АыУГХвЦЦКЙѕъМеХ№КѕјјКхЈ¬НтµИИЛЅ« C7C КЙѕъМеХ№КѕївНЁ№э±ЗЗ»ёшґуКуК№УГЈ¬ґУДФЧйЦЇЦР»ШКХКЙѕъМеЈ¬Іўјш¶ЁіцТ»ЦЦїЛВЎ7µДлДРтБРЈЁACTTPHAWLCGЈ©Ј¬ЛьДЬ№»НЁ№эРбѕхЗшУтґУ±ЗЗ»µЅґуДФµДНЁВ·ИЖ№эСЄДФЖБХПЎЈК№УГКЙѕъМеХ№КѕјјКх·ўПЦµД7ёцІР»щµДлД pepC7ЈЁCTSTSAPYCЈ»±н 1Ј©ПФКѕіцПтДФЧйЦЇµДМШРФЈ¬ЖдЧЄО»Р§ВК±ИЛж»ъївКЙѕъМеёЯ 41 ±¶ЎЈІј¶ыМ©µИИЛАыУГКЙѕъМеХ№КѕјјКхјш¶ЁіцТ»ЦЦєЬУРЗ°ѕ°µДСЄДФЖБХПґ©№эµДлДРтБР LRPep2ЈЁHPWCCGLRLDLRЈ»±н 1Ј©Ј¬ЛьХл¶ФµНГЬ¶ИЦ¬µ°°ЧКЬМеЈЁLDLRЈ©µДПё°ыНвЅб№№УтЈ¬ХвФЪРЎКуДФЗРЖ¬ЙПСЄ№ЬЦР LRPep2 Ул LDLR №І¶ЁО»µДГвТЯУ«№вИѕЙ«ЦеõЅБЛЦ¤ГчЈЁНј 3(c)Ј©ЎЈНЁ№эКЙѕъМеХ№КѕјјКх·ўПЦµДСЄДФЖБХПґ©ЛулДµДЖдЛыАэЧУЈ¬Из L57ЎўVH4127ЎўлД 22ЎўTHRЎўCRTЎўHAIЎўSLSЎў12-mer-1.2Ўў12-mer-1.3ЎўG23ЎўTACL08ЎўTGNЎўgHoPe2 єН E1-3Ј¬ЧЬЅбУЪ±н 1 ЦРЎЈ
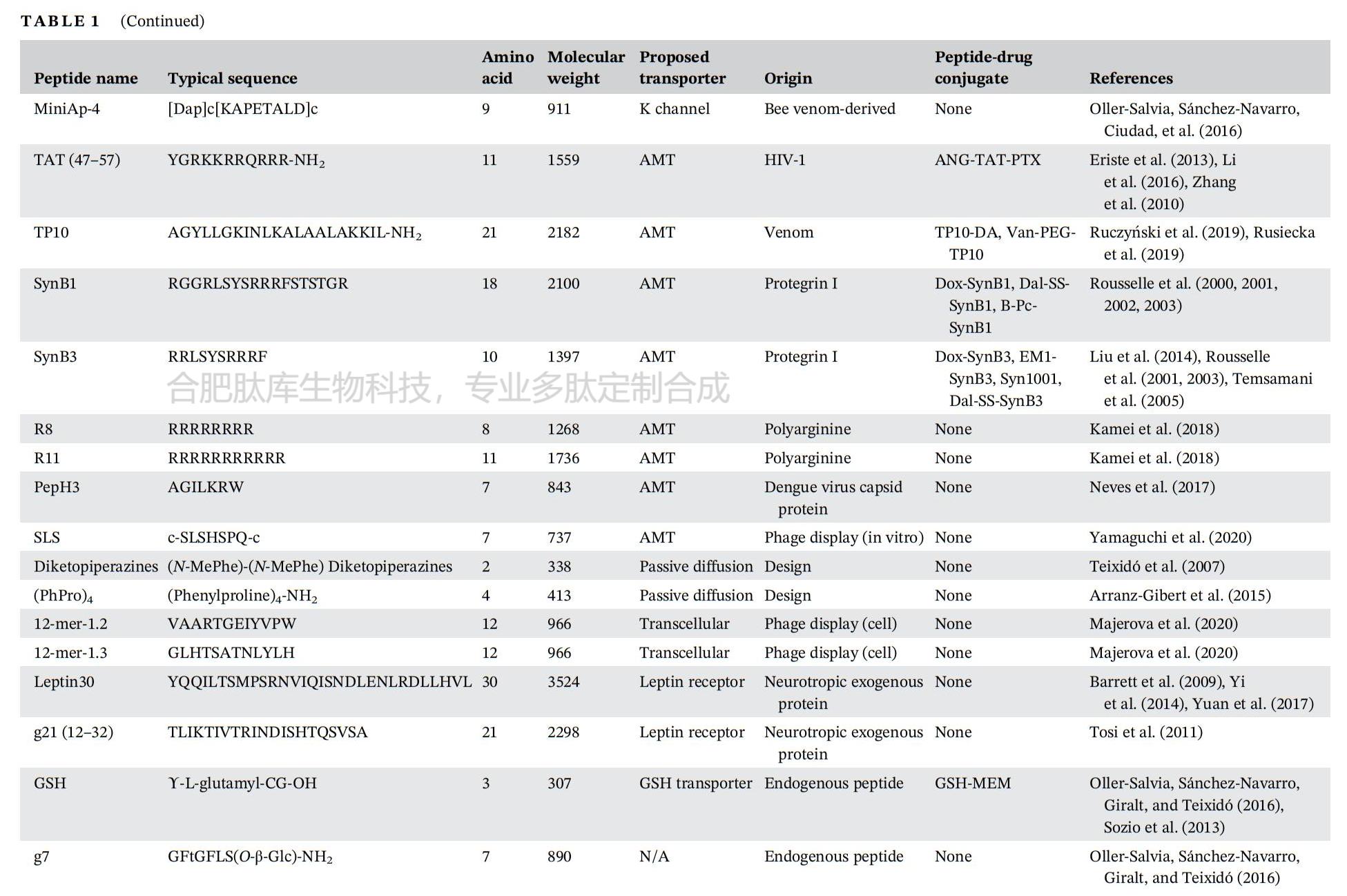
ЛхРґЈєAMTЈ¬ОьёЅЅйµјµДПё°ыЧЄИѕЈ»B-Pc-SynB1Ј¬ЬР»щЗаГ№ЛШ - SynB1Ј»D-[°±»щЛб]Ј¬D- РэПт°±»щЛбЈ»Dal-SS-SynB1Ј¬D- ѕ«°±Лб - SS - SynB1Ј»GABA-CTPЈ¬¦Г-°±»щ¶ЎЛб -Пё°ыЦКЧЄµјлДЕјБЄОпЈ»GM1Ј¬µҐНЩТєЛбЛДјєМЗЙсѕЅЪЬХЦ¬Ј»GSH-MEMЈ¬№ИлЧёКлД-ГАО¬МШЈ»K НЁµАЈ¬јШНЁµАЈ»LDLRЈ¬µНГЬ¶ИЦ¬µ°°ЧКЬМеЈ»LRP-1Ј¬µНГЬ¶ИЦ¬µ°°ЧПа№Шµ°°Ч 1Ј»LRP-2Ј¬µНГЬ¶ИЦ¬µ°°ЧПа№Шµ°°Ч 2Ј»N/AЈ¬І»їЙУГЈ»nAchRЈ¬СМјоРНТТхЈµЁјоКЬМеЈ»TfRЈ¬ЧЄМъµ°°ЧКЬМеЈ»TP10-DAЈ¬TP10-¶а°Н°·Ј»Van-PEG-TP10,Нт№ЕГ№ЛШ - ѕЫТТ¶юґј
3.3 | ФґЧФЙсѕУЄСшµ°°ЧєНІЎ¶ѕµД BBB ґ©ЛулД
һЩѪДФЖБХПЈЁBBBЈ©ґ©ЛулДФґЧФДЪФґРФЙсѕУЄСшµ°°ЧЎЈНЁ№эРтБР±И¶ФЎўєПіЙЖ¬¶ОµДЙёСЎ»тЙжј°XЙдПЯєНєЛґЕ№ІХсЈЁNMRЈ©µДЅб№№СРѕїЈ¬їЙТФјш¶ЁіцёєФрСЄДФЖБХПФЛКдµДлД¶ОЈЁНј3(b)Ј©ЎЈАэИзЈ¬К№УГєЛґЕ№ІХсјјКхЈ¬ТСЦ¤ГчФШЦ¬µ°°ЧEЈЁApoEЈ©µ°°ЧµДРтБР»щРтУлСЄДФЖБХПЙПµДКЬМеLRP-1ЧЄФЛµ°°ЧП໥ЧчУГЈ¬ІўёєФрДФЧЄ°ыНМЧчУГЎЈДіР©ЙсѕУЄСшІЎ¶ѕТСЦЄѕЯУРґ©№эСЄДФЖБХПІўµЅґпДФКµЦКµДДЬБ¦ЎЈФґЧФHIV-1µДTATлДЈЁ±н1Ј©µИлДФШМеїЙТФґ©№эДФДЪЖ¤Пё°ыФЛКдІ»Н¬µД»хОпЎЈК±јдФґЧФ 2 РНµЗёпИИІЎ¶ѕТВїЗµ°°ЧµДлД¦Б-ВЭРэЅб№№Ут PepH3ЈЁ±н 1Ј©НЁ№э»щУЪєЛґЕ№ІХсЈЁNMRЈ©µДјјКхµГТФјш¶ЁЈ¬ІўЗТНЁ№э AMT »ъЦЖПФКѕіцБјєГµДДФІїґ©НёДЬБ¦ЎЈ
ОЄБЛУЕ»ЇСЄДФЖБХПЈЁBBBЈ©ґ©ЛулДЈ¬РиТЄЅшРР»ЇС§РЮКОєНУЕ»ЇЈ¬ТФёДЙЖЛьГЗµДОпАн»ЇС§РФЦКЎўФЛКдЙшНёРФ»тЕдМе-КЬМеЅбєПЗЧєНБ¦ЎЈУЕ»Ї№эіМ°ьАЁјхЙЩлДЙПµДЗвјьµҐО»ЎўµчХыLogPЦµЈ¬»тґУМмИ»°±»щЛбЧЄ±дОЄ·ЗМмИ»°±»щЛб»тлДАаЈ¬ТФК№ЛьГЗ¶Фµ°°ЧГёёьОИ¶ЁЎЈЅ«D-¶ФУіМе°±»щЛбµДХыєПКЗФцЗї¶Фµ°°ЧЛ®ЅвГёЅµЅвµЦї№РФµДУРР§·Ѕ·ЁЈ¬ТтОЄГё¶ФL-лДјьѕЯУРµЧОпМШТмРФЎЈАэИзЈ¬ЈЁPhProЈ©4 [(Phenylproline)4-NH2]єН¶юНЄЯЯаєЈЁ±н1Ј©±»їЄ·ўіцАґТФФцјУЛ®ИЬРФєНСЄДФЖБХПЙшНёРФЎЈХвР©лДµД·ЦЧУБїРЎУЪ500µА¶ы¶ЩЈ¬НЁ№э±»¶ЇА©Йўґ©№эСЄДФЖБХПµДЗ±Б¦ёьґуЎЈУлЈЁN-MePheЈ©4лДПа±ИЈ¬ЈЁPhProЈ©4µДИЬЅвРФФцјУБЛ1000±¶Ј¬ЙшНёРФТІПФЦшФцјУЎЈТ»ёц12ёцІР»щµДСЄДФЖБХПґ©ЛуTHRЈЁTHRPPMWSPVWPЈ»±н1Ј©ФЪСЄЗеЦРПФКѕіцµНОИ¶ЁРФЎЈ·ґКЅ-ПыРэ°ж±ѕµД THRЈЁTHRreЈ©±»ЦЖФміцАґТФФцЗїОИ¶ЁРФЈ»¶Фє¬УРБЅёц№ІјЫБ¬ЅУлДµДЦ§»Ї THRre лДµДЅшТ»ІЅРЮКОК№СЄДФЖБХПЈЁBBBЈ©µДФЛКдДЬБ¦МбёЯБЛ 2.6 ±¶ЎЈEriste µИИЛНЁ№эЅ« CPP pVEC (LLIILRRRIRKQAHAHSK-NH2) УлНЁ№э№в¶ЇБ¦БЖ·ЁЈЁPDTЈ©·ўПЦµДХл¶ФЙсѕЅєЦКБцµДлД gHo (H2NQQQNPHQPPM-NH2) ЕјБЄЈ¬єПАнµШЙијЖБЛТ»ЦЦСЄДФЖБХПґ©ЛуИЪєПлДФШМе gHoPe2ЈЁ±н 1Ј©ЎЈЅ« PDT УГУЪ·ўПЦРВлДєН CPP ТФФцЗїПё°ыЙгИЎµДБЄєП·Ѕ·ЁЈ¬ІъЙъБЛТ»ЦЦФЪВДЪЦЧБцЦР»эАЫµДУЕ»ЇµД gHoPe2 лДЈ¬¶ш gHo лДФтГ»УРЎЈ
4 | ДФґ©НёлДµДіЈјыЧчУðег
4.1 | ЧЄМъµ°°ЧКЬМе
ЧЄМъµ°°ЧКЬМеЈЁTfRЈ©КЗСЄДФЖБХПЈЁBBBЈ©ЦРRMTНЁВ·µДТ»ёцМШХчГчИ·µДКЬМеЈ¬КЗТ»ЦЦД¤МЗµ°°ЧКЬМеЈ¬ЅйµјМъЅбєПµ°°ЧЧЄМъµ°°ЧµДФЛКдЈ¬ІўЧчОЄµчЅЪМъЙгИЎєНОИМ¬µДКШГЕФ±ЎЈѕЎ№ЬTfRґжФЪУЪДФГ«ПёСЄ№ЬДЪЖ¤Пё°ыЎўВцВзґФЙПЖ¤Пё°ыЎўЙсѕФЄЈ¬їЙДЬ»№УРЙсѕЅєЦКПё°ыЙПЈ¬µ«ЛьЦчТЄФЪёОПё°ыєНємПё°ыЦР±нґпЈ»ТтґЛЈ¬TfRІў·ЗСЄДФЖБХПЛщ¶АУРЎЈґу¶аКэАаРНПё°ыЙПTfRµД±нґпЛ®ЖЅИЎѕцУЪМъµД№©У¦Л®ЖЅј°ЖдФцЦіВКЎЈTfRЅйµјµДЧЄ°ыНМЧчУГ№эіМФКРнС»·МъЅбєПЧЄМъµ°°ЧНЁ№эСЄДФЖБХПІўЅшИлДФјдЦКїХјдЈ¬ХвУлТИµєЛШєНТИµєЛШСщЙъі¤ТтЧУНЁ№эСЄДФЖБХПµДТИµєЛШКЬМеФЛКдµЅґуДФµД·ЅКЅАаЛЖЎЈ¶сРФПё°ыУЙУЪЖдЙъі¤РиТЄёЯЛ®ЖЅµДМъЈ¬ТтґЛTfRµД±нґпЛ®ЖЅЙэёЯЎЈТтґЛЈ¬ЧЄМъµ°°ЧТС±»УГЧчФШМе»т°РПтЕдМеЈ¬Ѕ«ї№°©Т©ОпЎўДЙГЧїЕБЈЎўµ°°ЧЦКєН»щТтµЭЛНЦБ№э¶И±нґпЧЄМъµ°°ЧКЬМеµДФцЦі°©Пё°ыЎЈУлЧЄМъµ°°ЧКЬМеЅбєПµДї№МеТС±»Ц¤ГчДЬ№»СЎФсРФµШ°РПтСЄДФЖБХПДЪЖ¤Пё°ыЎЈOX-26КЗТ»ЦЦґуКуЧЄМъµ°°ЧКЬМ嵥Бґї№МеЈ¬ТС±»іЙ№¦УГУЪїзФЅСЄДФЖБХПµЭЛНѕЫєПОпМеєНДЙГЧїЕБЈЎЈУ뵥¶АѕІВцёшТ©Па±ИЈ¬НЁ№эЅ«OX-26УлЦОБЖјБЅбєПЈ¬ДФІїТ©ОпЕЁ¶ИПФЦшМбёЯЎЈѕЯУРУЕ»ЇУлЧЄМъµ°°ЧКЬМеЅбєПµДЛ«МШТмРФї№МеЈ¬їЙ°РПт¦В-·ЦГЪГёЈ¬ДЬ№»ґ©№эСЄДФЖБХПІўјхЙЩРЎКуДФІїµн·ЫСщµ°°Ч-¦ВЈЁA¦ВЈ©ЎЈТ»ЦЦУЙ№¤іМ»ЇОЄУлЧЄМъµ°°ЧКЬМеЅбєПµДµ°°ЧЦКЖ¬¶ОУлї№¦В-·ЦГЪГёї№ФЅбєПЖ¬¶ОЈЁFabЈ©ИЪєП¶шіЙµДФЛКдФШМеЈ¬їЙПФЦшФцЗїРЎКуєНґуКуµДДФІїФЛКдєНЦРКаЙсѕПµНі·ґУ¦ЎЈИЛГЗ·ўПЦЦоИз THRЎўCRT єН HAI µИ BBB ґ©ЛулДЈЁ±н 1Ј©ТФЧЄМъµ°°ЧКЬМеЈЁTfRЈ©ОЄ°РµгЎЈ
4.2 | µНГЬ¶ИЦ¬µ°°ЧКЬМе
µНГЬ¶ИЦ¬µ°°ЧКЬМеЈЁLDLRЈ©Ѕйµјё»є¬µЁ№МґјµДµНГЬ¶ИЦ¬µ°°ЧЈЁLDLЈ©µДДЪНМЧчУГЈ¬ІўТС±»№г·єСРѕїУГУЪЅ«µ°°ЧЦКєНЦОБЖТ©Опґ©№эСЄДФЖБХПЈЁBBBЈ©ФЛКдЦБЦРКаЙсѕПµНіЈЁCNSЈ©ЎЈLDLR ФЪЦ§Жш№ЬЙПЖ¤Пё°ыТФј°ЙцЙППЩєНЖ¤ЦКЧйЦЇЦР±нґпЧоОЄПФЦшЎЈЛьФЪСЄДФЖБХПТФј°ёчЦЦЦЧБцПё°ыЈЁИзЙсѕЅєЦКБцПё°ыЈ©ЦРТІёЯ¶И±нґпЎЈФґЧФ LDLR ЅбєПО»µгµДФШЦ¬µ°°Ч BЈЁApoBЈ©єНФШЦ¬µ°°Ч EЈЁApoEЈ©лДЈЁ±н 1Ј©ТС±»Ц¤ГчФЪЦРКаЙсѕПµНіјІІЎЦР¶Ф LDLR °РПтЦОБЖУРР§Ј¬ѕЎ№ЬУлДЪФґРФ LDL ѕєХщТФј°ИЕВТґуДФЦРµДµЁ№МґјОИМ¬Т»Ц±КЗТ»ёцБоИЛµЈУЗµДОКМвЎЈНЁ№эКЙѕъМеХ№КѕЙъОпЙёСЎ·ЁїЄ·ўµД Peptide-22ЈЁ±н 1Ј©¶Ф LDLR УРЗЧєНБ¦Ј¬ЗТУлДЪФґРФ LDL ОЮѕєХщЈ¬ДЬ№»ёЯР§ЗТїмЛЩµШЧЄТЖµЅЦРКаЙсѕПµНіЎЈ
4.3 | µНГЬ¶ИЦ¬µ°°ЧКЬМеПа№Шµ°°Ч 1 єН 2ЈЁLRP-1 єН LRP-2Ј©
LRP-1 єН LRP-2 КЗ LDLR і¬јТЧеЦРґуРНµДЎў¶а№¦ДЬµДЎў¶аЕдМеµДДЪНМКЬМеЎЈЛьГЗЅйµј¶аЦЦЕдМеµДДЪ»ЇєНЅµЅвЈ¬ХвР©ЕдМеЙжј°ёчЦЦґъР»Нѕѕ¶ЎЈLRP КЬМеУРБЅёцЦчТЄ№¦ДЬЈєЈЁaЈ©ЧчОЄЗеµА·тКЬМеІОУлЖдЦЪ¶аЕдМеµДДЪНМЧчУГЈ»ЈЁbЈ©ЧчОЄРЕєЕКЬМеµчЅЪёчЦЦПё°ы№эіМЎЈLRP КЬМеУл№г·єµДЈЁ>40 ЦЦЈ©·ЦГЪµ°°ЧєНПё°ы±нГжЧ¤Бф·ЦЧУП໥ЧчУГЈ¬°ьАЁФШЦ¬µ°°Ч EЎў¦Б2-ѕЮЗтµ°°ЧЈЁ¦Б2MЈ©ЎўКЬМеПа№Шµ°°ЧЎўЧйЦЇРНПЛИЬГёФј¤»оОпЎўПЛИЬГёФј¤»оОпТЦЦЖјБ 1Ўўµн·ЫСщЗ°Меµ°°ЧЎўИйМъµ°°ЧєНєЪЛШЧЄФЛµ°°ЧЎЈ¶аДкАґЈ¬LRP-1 єН LRP-2 КЬМеТС±»їЄ·ўУГУЪНЁ№э RMT Нѕѕ¶Ѕ«Т©Оп°РПтµЅґуДФЎЈ
LRP-1 КЬМеЈЁТІіЖОЄ CD91 »т¦Б2MКЬМеЈ¬600 З§µА¶ы¶ЩЈ©ФЪ°ьАЁёОФаєН·ОФЪДЪµД¶аЦЦЧйЦЇЦРЖХ±й±нґпЈ¬ІўІОУлµЁ№МґјґъР»ЎўДЪНМЧчУГєНПё°ыРЕєЕґ«µјЎЈLRP-1 ФЪХыёцЦРКаЙсѕПµНі№г·є±нґпЈ¬ЦчТЄФЪґуДФЖ¤ЦКЎўВцВзґФЎўєЈВнМеєНРЎДФЎўДФДЪЖ¤Пё°ыЎўСЄ№ЬЖЅ»¬јЎПё°ыЎўЦЬПё°ыЎў»о»ЇµДРЗРОЅєЦКПё°ыєНРЎЅєЦКПё°ыЦРЎЈLRP-1 ФЪ¶сРФРЗРОПё°ыБцЦР№э¶И±нґпЈ¬¶шФЪ°ў¶ыґДєЈД¬ІЎЦР LRP-1 µД±нґпЅµµНЎЈ№ШУЪ LRP-1 ФЪґуДФЦРµД№¦ДЬµДґу¶аКэСРѕї¶јјЇЦРФЪЛьµчЅЪµн·ЫСщЗ°Меµ°°ЧФЛКдЎўµн·ЫСщ¦ВґУДФКµЦКЗеіэТФј°СЄДФЖБХПНЁНёРФµДЧчУГЙПЎЈ
LRP-2 КЬМеЈЁТІіЖОЄ megalinЈ©КЗТ»ЦЦ 600 З§µА¶ы¶ЩµДїзД¤µ°°ЧЈ¬ТІКЗј№Чµ¶ЇОпЦРґжФЪµДЧоґуµДПё°ы±нГжМЗµ°°ЧЦ®Т»ЎЈLRP-2 µДЅб№№Ул LRP-1 ·ЗіЈПаЛЖЎЈLRP-2 ЦчТЄФЪЙцФаЎў°тлЧєНґуДФЦР±нґпЎЈФЪЅЎїµµДґуДФЦРЈ¬LRP-2 ФЪґуДФЖ¤ЦКЎўєЈВнМеЎўОЖЧґМеЎўЗрДФЎўРбЗтєНРЎДФµДІ»Н¬ЙсѕФЄЦР±нґпЎЈLRP-2 ТІФЪЕЕБРФЪДФКТ±ЪµДКТ№ЬД¤Пё°ыЎўГ«ПёСЄ№ЬЎўВцВзґФЎўЙсѕФЄєНРЗРОЅєЦКПё°ыЦР±нґпЎЈФЪ»јУР°ў¶ыґДєЈД¬ІЎµД»јХЯµДДФЧйЦЇЦРЈ¬·ўПЦ LRP-2 ФЪКЬЛрЙсѕФЄЦРЙПµчЎЈ
LRP-1 єН LRP-2 ФЪЅ«·ЦЧУФЛКдµЅПё°ыДЪ·ЅГжХ№ПЦіцБЛБоИЛЦхДїµДДЬБ¦ЎЈСРѕїТ»Ц±јЇЦРУЪАыУГ LRP КЬМеµД»хО﹦ДЬЈ¬НЁ№эЧЄ°ыНМЧчУГЅ«Т©Опґ©№эСЄДФЖБХПЈЁBBBЈ©ЎЈТСИ·¶ЁБЛХл¶Ф LRP КЬМеµДУГУЪДФІїТ©ОпФЛКдµДлДЈЁ±н 1Ј©ЎЈѕЭ±ЁµАЈ¬ѕЫЙЅАжхҐ 80ЈЁTween-80Ј©°ьІгµДѕЫ¶Ў»щЗи»щ±ыП©ЛбхҐДЙГЧїЕБЈФЪѕІВцЧўЙдєуДЬґУСЄТєЦРОьёЅФШЦ¬µ°°Ч EЈЁApoEЈ©єНФШЦ¬µ°°Ч BЈЁApoBЈ©Ј¬ІўНЁ№э LRP-1 ЅйµјµДЧЄ°ыНМЧчУГНѕѕ¶ґЩЅшСЄДФЖБХПµДґ©ФЅЎЈ
4.4 | КЭЛШКЬМе
КЭЛШКЗТ»ЦЦ16 kDaµДј¤ЛШЈ¬ЦчТЄУЙ°ЧЙ«НвЦЬЦ¬·ѕПё°ыІъЙъЈ¬ФЪµчЅЪКіОпЙгИлєНО¬іЦДЬБїЖЅєв·ЅГж·ў»УЦШТЄЧчУГЎЈКЭЛШ»№ІОУлµчЅЪС§П°єНјЗТд№эіМЎЈКЭЛШКЬМеТС±»·ўПЦґжФЪУЪЙсѕЅєЦКПё°ыЎўРЗРОЅєЦКПё°ыЎўВцВзґФєНДФГ«ПёСЄ№ЬДЪЖ¤Пё°ыЦРЎЈКЭЛШґУЦ¬·ѕПё°ыКН·ЕµЅСЄТєЦРЈ¬ІўНЁ№эУл±нґпФЪДФКµЦКЦРµДКЭЛШКЬМеП໥ЧчУГ¶ш±»ЙгИЎЎЈДФДЪЖ¤Пё°ы»бУ°ПмЅшКіЎўІъИИЎў»о¶ЇµИЙъАнєНРРОЄ·ЅГжЎЈФЪ·КЕЦ»јХЯЦРЈ¬УЙУЪКЭЛШЛ®ЖЅЙэёЯЈ¬КЭЛШКЬМеїЙДܻᱥєНЎЈТ»ЦЦФґЧФКЭЛШµДлДРтБРg21ЈЁ12-32ёц°±»щЛбЈ»±н1Ј©Ж¬¶ОТС±»УГУЪУлѕЫИйЛб-фЗ»щТТЛб№ІѕЫОпЈЁPLGAЈ©ДЙГЧїЕБЈ±нГжЅбєПЈ¬ХвР©ДЙГЧїЕБЈФЪѕІВцЧўЙдєуґ©№эСЄДФЖБХПЈ¬ІўФЪ2РЎК±єуµЅґпґуДФЈ¬ХјЧўЙдјББїµД0.16%ЎЈБнТ»ЦЦФґЧФКЭЛШµД30ёц°±»щЛбµДлДЈЁLeptin30Ј»±н1Ј©±»їЄ·ўОЄДЙГЧїЕБЈµДДФ°РПтЕдМеЎЈ
4.5 | СМјоРНТТхЈµЁјоКЬМе
СМјоРНТТхЈµЁјоКЬМеЈЁnAchRЈ©КфУЪЕдМеГЕїШАлЧУНЁµАі¬јТЧеЈ¬°ьАЁ¦Г-°±»щ¶ЎЛбAРНєНCРНКЬМеЎўёК°±ЛбКЬМеєН5-фЗЙ«°·КЬМеЎЈnAchRЅйµјЙсѕєНјЎИвПё°ыЦ®јдµДН»ґҐґ«µЭЎЈnAchR¶ФЙсѕµЭЦКТТхЈµЁјоєНЙПс«РФ¶ѕЖ·Дб№Е¶Ў¶јУР·ґУ¦ЎЈnAchRЦчТЄФЪЦРКаЙсѕПµНіЎўНвЦЬЙсѕПµНієНјЎИвЦР±нґпЎЈЙсѕФЄnAchRУЙОеёцїзД¤СЗ»щЧйіЙЈ¬ХвР©СЗ»щО§ИЖЧЕТ»ёцЦРСлідВъЛ®µДїЧП¶ЕЕБРЎЈТС±ЁµАµД505ёц°±»щЛбµДЙсѕ¶ѕРФїсИ®ІЎ¶ѕМЗµ°°ЧЈЁRVGЈ©УлnAchRП໥ЧчУГЈ¬Іў±»УГУЪґ«µЭёчЦЦДЙГЧїЕБЈЎўRNAєНDNAЖ¬¶ОЎЈRVGСЬЙъµД29ёц°±»щЛбµДлДRVG29ЈЁ±н1Ј©УлPAMAMЈЁѕЫхЈ°·°·Ј©ДЙГЧїЕБЈЅбєПЈ¬ФЪМеНвСЄДФЖБХПДЈРНЦРПФКѕіц±Иґ«НіДЙГЧїЕБЈёьёЯµДСЄДФЖБХПґ©ФЅР§ВКЎЈБнТ»ЖЄ±ЁµАіЖЈ¬Т»ЦЦ 16 ёц°±»щЛбµД D-CDX лДЈЁ±н 1Ј©їЙНЁ№э nAchR ЅйµјµДПё°ыДЪНМЧчУГґЩЅшДФ°РПтТ©ОпµЭЛНЎЈ
5 | УГУЪ PDCS µД BBB ґ©ЛулД
ѕЎ№ЬХл¶ФСЄДФЖБХПЈЁBBBЈ©ЙП±нґпµДЧЄФЛµ°°ЧµДї№Ме»тДЪФґРФµ°°ЧЦКЕдМеУРЛщЅшІЅЈ¬µ«Т»Р©ёЯЗЧєНБ¦ї№МеУЙУЪ¶ФКЬМеµДёЯ¶ИЗЧєНБ¦ТФј°ФЪґуДФКµЦКЦРОЮР§µД»хОпКН·Е¶шЧи°БЛ»хОпµДґуДФµЭЛНЎЈґЛНвЈ¬µ°°ЧЦКµДёЯіЙ±ѕЙъІъєНёґФУЅб№№ПЮЦЖБЛХвЩѪДФЖБХПґ©Луµ°°ЧµД№г·єУ¦УГЎЈРЎіЯґзлДТтЖдДЬ№»±нПЦіціцЙ«µД°РПтМШТмРФЎўµН¶ѕРФєНГвТЯФРФЈ¬ТФј°ЧоРЎµДєПіЙєНЦЖФмМфХЅ¶шФЅАґФЅКЬµЅ№ШЧўЎЈёчЦЦСЄДФЖБХПґ©ЛулДТС±»СРѕїТФЖА№АЖдґ©№эСЄДФЖБХПµДДЬБ¦Ј¬ХвР©лДФЪ±н1ЦРЅшРРБЛЧܽᡣһЩѪДФЖБХПґ©ЛулДНЁ№эАыУГСЎФсРФRMTНЁВ·µДКЬМеТААµ»ъЦЖґ©№эСЄДФЖБХПЈЁНј2Ј©Ј¬¶шТ»Р©єПіЙ»тµ°°ЧЦКСЬЙъµДПё°ыґ©НёлДЈЁCPPЈ©ФтНЁ№эКЬМе·ЗТААµ»ъЦЖЈ¬ЦчТЄКЗНЁ№эAMTНЁВ·Ј¬ЧчОЄґуДФµЭЛНФШМеПФКѕіцПЈНыЎЈCPPs НЁіЈУЙ 5 ЦБ 30 ёц°±»щЛбЧйіЙЈ¬ґуЦВ·ЦОЄБЅАаЈєёЯСфАлЧУРНєНБЅЗЧРФРНЎЈCPPs µДХэµзєЙєНБЅЗЧРФКЗФКРн CPPs ґ©№эЙъОпД¤ІўЅ«ЙъОп»оРФ»хОпЗэ¶ЇЅшИлПё°ыµД№ШјьМШХчЎЈТ»Р© CPPs ±нПЦіцСЄДФЖБХПґ©НёДЬБ¦ЎЈПВГжЦШµгЅйЙЬБЛТ»Р©іЈУГµДФґЧФ PDTЎўCPP »тМмИ»µ°°ЧЦКµДСЄДФЖБХПґ©ЛулДЈ¬УГУЪ№№ЅЁ PDCЎЈ
5.1 | Angiopep-2
Angiopep-2КЗТ»ЦЦ19ёц°±»щЛбµДлДЈЁTFFYGGSRGKRNNFKTEEYЈ¬·ЦЧУБї2.3 kDaЈ»±н1Ј©Ј¬КЗНЁ№эЅ«6.5 kDaµД¶алДaprotininЈЁLRP-1єНLRP-2µДЕдМеЈ©УлЖдЛыИЛАаµ°°ЧЦКЈЁ°ьАЁbikuninЎў·ЦГЪµн·ЫСщЗ°Меµ°°ЧєНKunitzТЦЦЖјБ-1З°Меµ°°ЧЈ©ЅшРРРтБР±И¶Ф¶шїЄ·ўµДЈ¬ХвИэЦЦµ°°ЧЦК¶јѕЯУРТСЦЄ¶ФLRPКЬМеУРЗЧєНБ¦µДKunitzЅб№№УтЎЈК№УГМеНвСЄДФЖБХПЕЈДФГ«ПёСЄ№ЬДЪЖ¤Пё°ыTranswellКµСйЈ¬Angiopep-2µДЧЄ°ыНМЧчУГДЬБ¦±ИaprotininёЯИэ±¶Ј¬±ИЧЄМъµ°°ЧєНИйМъµ°°ЧёЯ50±¶ТФЙПЎЈґЛНвЈ¬ЅьємНвУ«№вИѕБПCy5.5±кјЗµДAngiopep-2НЁ№эМеДЪіЙПсєНДФЗРЖ¬µДУ«№в·ЦОцПФКѕЈ¬ДЬ№»СёЛЩЅшИлДФКµЦКЈЁНј4(a)Ј©ЎЈCy5.5-Angiopep-2ЈЁємЙ«Ј©О»УЪДФКµЦКЦРЎЈ
ФЪѕІВцЧўЙдєу 1 РЎК±Ј¬№ЫІмµЅУ«№в±кјЗµД Angiopep-2ЈЁВМЙ«Ј©їїЅьДФПё°ыµДПё°ыєЛЈЁА¶Й«Ј©Ј¬¶ш Cy5.5-Angiopep-7ЈЁТхРФ¶ФХХЈ©Оґґ©№эДФГ«ПёСЄ№ЬЈЁВМЙ«Ј»Нј 4(a)Ј©ЎЈНЁ№э»оМеіЙПсЈ¬ФЪѕІВцёшТ© 5 Ммєу№ЫІмµЅУ«№в±кјЗµД Angiopep-2ЈЁВМЙ«Ј©ФЪ»оМеРЎКуґуДФµДДФД¤єНКµЦКЦРµД»эАЫЈЁНј 4(b)Ј©ЎЈФЪѕІВцёшТ©єу 30 ·ЦЦУЦБ 24 РЎК±µДК±јд·¶О§ДЪЈ¬МеДЪРЎКуґуДФіЙПсПФКѕ Cy5.5-Angiopep-2 ФЪґуДФЦРµД»эАЫПФЦшёЯУЪ Cy5.5-Angiopep-7ЎЈCy5.5-Angiopep-2 У«№вµД AUC Цµ±И Cy5.5-Angiopep-7 ёЯ 9.5 ±¶Ј¬±нГч Angiopep-2 ¶ФґуДФµДСЎФсРФЙгИЎЈЁНј 4(c)Ј©ЎЈAngiopep-2 µДЧоіхУ¦УГІаЦШУЪ»ЇС§єПіЙ Angiopep-2 РЎ·ЦЧУТ©ОпЕјБЄОпЈ¬°ьАЁЧПЙјґјЎўТАНРІґЬХЎў¶аИб±ИРЗЎўВр·ИЎўВр·И-6-¦В-ЖПМСМЗИ©ЛбхҐЎўї§·ИЛбєНЙсѕµЭЦКµДЕјБЄОпЈЁНј 6Ј©ЎЈЛжєу¶Ф Angiopep-2 µДАыУГКЗЅ«ФШМеЈЁИзДЙГЧБЈЧУЎўЅєКшЎўМјДЙГЧ№ЬєНї№МеЈ©ЅшРРёДФмЈ¬ТФ±гПтґуДФКдЛНТ©ОпЈ¬ХвЦ§іЦБЛСЄДФЖБХПґ©Луµ°°Ч Angiopep-2 ФЪОЄёчЦЦЦРКаЙсѕПµНіјІІЎКдЛНРЎ·ЦЧУєНґу·ЦЧУ·ЅГжµДНЁУГРФЎЈ
5.2 | Хл¶Ф LRP-1ЎўLRP-2 єН LDLR µДлД
Т»ЦЦ 18 ёцІР»щµДФШЦ¬µ°°Ч E лД¶ОєНТ»ЦЦ 39 ёцІР»щµДФШЦ¬µ°°Ч B лД¶ОЈЁ±н 1Ј©ФґЧФДЪФґРФЙсѕЗчПтµДФШЦ¬µ°°Ч E єНФШЦ¬µ°°Ч B µ°°ЧЈ¬НЁ№эУлґуДФЦРµДµНГЬ¶ИЦ¬µ°°ЧКЬМеП໥ЧчУГПФКѕіцСЄДФЖБХПґ©НёР§№ыЎЈЧоРВµДКЙѕъМеХ№КѕЙъОпЙёСЎ»№·ўПЦБЛјёЦЦСЄДФЖБХПґ©ЛулДЈ¬°ьАЁХл¶ФµНГЬ¶ИЦ¬µ°°ЧКЬМеµД Peptide-22ЎўVH4127ЎўLRPep2 єН AEP лДЈ¬ТФј°Хл¶Ф LRP-1 КЬМеµД L57 лДЈЁ±н 1Ј©ЎЈ
5.3 | Хл¶ФЧЄМъµ°°ЧКЬМеЈЁTfRЈ©µДлД
НЁ№эКЙѕъМеХ№КѕЙъОпЙёСЎ№эіМЈ¬·ўПЦБЛТ»ёц 12 ёцІР»щµД THR лДЈЁTHRPPMWSPVWPЈ©єНТ»ёц 7 ёцІР»щµД HAI лДЈЁHAIYPRHЈ»±н 1Ј©КЗИЛАаЧЄМъµ°°ЧКЬМеµДЅбєПлДЎЈИЪєПВМЙ«У«№вµ°°ЧЈЁGFPЈ©µДЧЄМъµ°°ЧКЬМеЈЁTfRЈ©лДїЙТФ±»ЧЄМъµ°°ЧКЬМе±нґпПё°ыДЪ»ЇЎЈСЄДФЖБХПґ©Лу THR лДТСУлЅрДЙГЧБЈЧУЈЁAuNPЈ©ЕјБЄЈ¬ТФФцјУДЙГЧБЈЧУФЪґуКуґуДФЦРµДЙшНёЎЈTHR лДДЬ№»УлПЩПа№ШІЎ¶ѕ AAV8 ІЎ¶ѕМеЅбєПЈ¬ІўПФЦшФцЗїЛьГЗґ©№эСЄДФЖБХПєНЧЄµјЙсѕФЄПё°ыµДДЬБ¦ЎЈСЄДФЖБХПґ©Лу HAI лДЕјБЄµДЦ¬ЦКМеФЪДФЅєЦКБцРЎКуДЈРНЦРёДЙЖБЛµЭЛНєНЦОБЖР§№ыЎЈHAI лДДЬ№»Ѕ« AuNP µЭЛНµЅґуКуґуДФЈ¬ІўЗТ±И THR лДПФКѕіцёьєГµДФЛКдДЬБ¦ЎЈБнТ»ЦЦСЄДФЖБХПґ©ЛулД CRTЈЁ»· CRTIGPSVC лДЈ¬±н 1Ј©СЎФсРФµШУлЧЄМъµ°°ЧКЬМеП໥ЧчУГЈ¬УХµј±р№№№№Пу±д»ЇЈ¬ґУ¶шФЪ№¦ДЬЙПЎ°ДЈДвЎ±МъЎЈCRT µјПтµДІЎ¶ѕїЕБЈФЪХэіЈРЎКуґуДФЦРПФКѕіцёДЙЖµДСЄДФЖБХПЙшНёРФЈ¬CRT ЕјБЄµДДЙГЧїЕБЈФЪРЎКуДЈРНµДЙсѕЅєЦКБцКµЦКЦРПФКѕіцФцЗїµДґуДФ·ЦІјЎЈ
5.4 | TAT СЬЙълД
TATЈЁИЛАаГвТЯИ±ПЭІЎ¶ѕ 1 РН[HIV-1]µДЧЄµјЅб№№УтЈ©µ°°ЧКЗТ»ЦЦ 86 ёц°±»щЛбµДЧЄВјТтЧУЈ¬ІОУл HIV-1 ІЎ¶ѕµДёґЦЖЦЬЖЪЈ¬ДЬ№»НЁ№эОьёЅДЪНМЧчУГЅшИлПё°ыІўЧЄТЖµЅПё°ыєЛЦРЎЈґУ TAT µ°°ЧµД¦Б-ВЭРэЅб№№УтЦРСЬЙъіцµД 11 ёцІР»щµД TAT лДЈЁ±н 1Ј©КЗµЪТ»ЦЦ±»јш¶ЁОЄСфАлЧУ CPP µДлДЎЈTAT лДТС±»Ц¤ГчДЬФцјУ TAT-¦В-°лИйМЗЬХГёЧєєПОпФЪ°ьАЁґуДФФЪДЪµДГ«ПёСЄ№ЬТФНвµДЖХ±йЧйЦЇґ©НёРФЎЈёчЦЦУл TAT ЧєєПµДДЙГЧБЈЧУПФКѕіцёДЙЖµДґуДФµЭЛНЎЈНЁ№эЅ«Л« TAT єН Angiopep-2 лДУлЧПЙјґјЧєєПєПіЙµД PDC ФЪВДЪЅєЦКБцРЎКуДЈРНЦРПФКѕіцТ©ОпПтґуДФµДЧЄО»µГµЅёДЙЖЎЈ
5.5 | Transportan 10
Transportan 10ЈЁTP10Ј©КЗТ»ЦЦ21ёцІР»щµДБЅЗЧРФCPPЈЁAGYLLGKINLKALAALAKKILЈ»±н1Ј©Ј¬ѕЯУРёЯД¤ЧЄО»ДЬБ¦ЎЈTP10ТС±»УГЧчлДєЛЛбЎўлДєНµ°°ЧЦКµДµЭЛНФШМеЎЈTP10-¶а°Н°·ЧєєПОпЈЁНј6Ј©НЁ№эРЎіЯґзPEGЧчОЄБ¬ЅУЧУЈ¬ДЬ№»ЅшИлДФЧйЦЇЈ¬¶Ф¶а°Н°·D1єНD2КЬМеѕЯУРПа¶ФЅПёЯµДЗЧєНБ¦Ј¬ІўФЪ¶ЇОпДЈРНЦРПФКѕіцї№ЕБЅрЙІЎµД»оРФЎЈБнТ»ЦЦTP10-Нт№ЕГ№ЛШЧєєПОпЈЁНј6Ј©ДЬ№»ґ©№эСЄДФЖБХПЈ¬ДФЧйЦЇЦРЧєєПОпє¬Бї±ИУОАлНт№ЕГ№ЛШёЯіц200±¶ЎЈ
5.6 | SynB
SynB лДФґЧФМмИ»µДї№ѕълД protegrinЈ¬КфУЪСфАлЧУ CPP јТЧеЎЈ18 ёцІР»щµД SynB1ЈЁRGGRLSYSRRRFSTSTGRЈ©єН 10 ёцІР»щµД SynB3ЈЁRRLSYSRRRFЈ©лДЈЁ±н 1Ј©ТС±»Ц¤ГчїЙНЁ№эОьёЅЅйµјµДДЪНМЧчУГФцЗїТ©Опґ©№эСЄДФЖБХПµДФЛКдЎЈL-SynB1ЎўL-SynB3 ј°Жд¶ФУіТм№№Ме D-SynB3 Ул°ўГ№ЛШЕјБЄЈЁНј 6Ј©Ј¬ЕјБЄОпК№°ўГ№ЛШµДґуДФЙгИЎБїПФЦшФцјУФј 30 ±¶ЎЈЎЈSynB1 »№ДЬФцЗїЬРОчБЦ»тВр·И-6-ЖПМСМЗИ©ЛбхҐФЪґуДФЦРµДЧчУГЈ¬ЗТІ»»бЛрє¦СЄДФЖБХПµДНкХыРФЎЈSynB1 лДНЁ№э¶юБтјьБ¬ЅУЧУУл dalargin Б¬ЅУЈЁНј 6Ј©Ј¬ёГЧєєПОпПФЦшёДЙЖБЛ dalargin ФЪґуДФЦРµДµЭЛНєНХтНґ»оРФЎЈ
5.7 | Penetratin
Penetratin КЗТ»ЦЦ 16 ёцІР»щµДСфАлЧУ CPPЈЁRQIKIWFQNRR-MKWKKЈ¬ТІіЖОЄ pAntp-(43-58)Ј©Ј¬ФґЧФ№ыУ¬ґҐЅЗЧгµ°°ЧЈЁ°±»щЛб 43-58Ј»±н 1Ј»Ј©ЎЈPenetratin НЁ№эОьёЅЅйµјµДДЪНМЧчУГПФКѕіцПё°ыґ©НёМШРФЎЈPenetratin ТС±»УГЧчёчЦЦ»хОпЈЁИз GFPЎў№СєЛЬХЛбєНЦ¬ЦКМеЈ©µДПё°ыДЪµЭЛНФШМеЎЈУлѕЫАµ°±ЛбЈЁёЯѕ«°±Лбє¬БїµД CPPЈ©№¦ДЬ»ЇµДДЙГЧБЈЧУПа±ИЈ¬Penetratin №¦ДЬ»ЇµДѕЫєПОпДЙГЧБЈЧУПФКѕіцПФЦшФцЗїµДґуДФЙгИЎЈ¬ІўјхЙЩФЪ·З°РЧйЦЇЦРµД»эАЫЎЈЧЄМъµ°°Ч - Penetratin РЮКОµДЦ¬ЦКМеёДЙЖБЛЦКБЈ ApoE2єНЦКБЈ¦В-°лИйМЗЬХГёµДґуДФµЭЛНЎЈ°ўГ№ЛШ - D-Penetratin ЧєєПОпЈЁНј 6Ј©ПФКѕ°ўГ№ЛШµДґуДФЙгИЎБїёЯіц 6 ±¶ЎЈ
5.8 | Пё°ыЦКЧЄµјлД
Пё°ыЦКЧЄµјлДЈЁCTPЈ©КЗТ»ЦЦ 11 ёцІР»щµДСфАлЧУ CPPЈЁYGRRARRRRRЈ»±н 1Ј©Ј¬ѕ№эєПАнЙијЖЈ¬ДЬ№»ґ©№эПё°ыД¤Ј¬ІўУЕПИѕЫјЇФЪПё°ыЦКЦРЎЈCTP ЕјБЄµД Smac/DIABLO лДєН CTP ИЪєПµД¦В-°лИйМЗЬХГёДЬ№»УРР§µШЅшИлПё°ыµДПё°ыЦКЗ»КТЎЈCTP ЕјБЄµДЙсѕµЭЦК¦Г-°±»щ¶ЎЛбЈЁGABAЈ©ДЬ№»ґ©НёСЄДФЖБХПЈ¬ІўМбёЯґуКуєНРЎКуСЄЅ¬ЦРµД GABA Л®ЖЅЎЈ
5.9 | ѕ«°±Лб-ёК°±Лб-М춬°±ЛблД
RGDЈЁѕ«°±Лб-ёК°±Лб-М춬°±ЛбЈ©лД°РПт¦Бv¦В3ХыєПЛШЈ¬УЙУЪ¦Бv¦В3ХыєПЛШФЪРВЙъСЄ№ЬЦР№э¶И±нґпЈ¬УИЖдКЗФЪДФБцЦРЈ¬ТтґЛТС±»№г·єУГЧчДФІїТ©ОпµЭЛНФШМеЎЈТС±ЁµА RGD лДРЮКОµДѕЫєПОпЎў·ЕЙдРФ±кјЗлДЎўЅєКшєНЦ¬ЦКМеФЪЅєЦКДёПё°ыБцµДХп¶ПєНЦОБЖЦРєЬУРУГЎЈХвР© RGD ДЙГЧБЈЧУ±»ИПОЄНЁ№эДЪ»ЇЅшИл°ЧПё°ыАґЅйµјјдЅУПё°ыДЪНМЧчУГЈ¬°ЧПё°ы»бПмУ¦СЧЦў±»ХРДјµЅґуДФЦРЎЈRGD-PEG4-Suc-PD0325901ЈЁТ»ЦЦ MEK1/2 ТЦЦЖјБЈ©ЕјБЄОпЈЁНј 6Ј©ФЪЦОБЖ U87MG ЅєЦКДёПё°ыБцРЎКуТмЦЦТЖЦІОп·ЅГжПФКѕіцУЕФЅµДЦОБЖР§№ыЈ¬±нГчёГТ©ОпУРНыКµПЦДФІїµЭЛНЎЈ
5.10 | №ИлЧёКлД
№ИлЧёКлДЈЁL-¦Г-№И°±хЈ-L-°ллЧ°±Лб-ёК°±ЛбЈ¬GSHЈ¬±н1Ј©КЗ»оПё°ыЦРЧо·бё»µДЈЁПё°ыДЪёЯґп10 mMЈ¬С»·ЦР5 ¦МMЈ©РЎ·ЦЧУБїБтґјЈ¬ЛьѕЯУРґУЅв¶ѕµЅ±Ј»¤ГвКЬСх»ЇЛрЙЛµИТ»ПµБР№¦ДЬЎЈДЪФґРФGSH±»И·¶ЁОЄСЄДФЖБХПґ©ЛулДЈ¬ХвїЙТФНЁ№эФЪѕ±¶ЇВцЧўЙдєуЈ¬ґуКуґуДФ¶Ф35S±кјЗµДGSHµД±ҐєНЙгИЎ№эіМАґЦ¤ГчЎЈЅ«GSHЧчОЄСЄДФЖБХПґ©ЛулДУГУЪПтґуДФµЭЛНТ©ОпµДЧоіЙ№¦АэЧУКЗG-Technology®ЎЈДЖТААµРФ№ИлЧёКлД-PEGЦ¬ЦКМе±»Ц¤ГчДЬ№»ПтґуДФµЭЛНї№ІЎ¶ѕТ©ОпЎЈGSH»№УлN-јЧ»щ-D-М춬°±ЛбКЬМеЮЧї№јБmemantineЈЁMEMЈ»Нј6Ј©№ІјЫБ¬ЅУЈ¬УГУЪПтґуДФµЭЛНТ©ОпЎЈ
5.11 | ѕЫѕ«°±ЛблД
ѕЫѕ«°±ЛблДЈ¬ИзR8ЎўR11єНR18Ј¬КЗё»є¬ѕ«°±ЛбµДCPPsЈ¬ФЪЙъАнМхјюПВ»бЦКЧУ»ЇЈ¬ІўУлБЧЦ¬Д¤µДґшёєµзєЙµДфИ»щЎўБтЛбСОєНБЧЛб»щНЕП໥ЧчУГЈ¬µјЦВД¤Ѕб№№Е¤ЗъЈ¬ІўТэ·ўЗЧЛ®РФЛ®їЧФЪД¤ЦРЗЁТЖЈ¬ґУ¶шТэ·ў·ЗКЬМеТААµРФПё°ыДЪНМЎЈѕЫѕ«°±ЛбCPPsїЙТФґЩЅш°ьАЁєЛЛбЎў»оРФµ°°ЧЎўБїЧУµгєНДЙГЧїЕБЈµИ·ЦЧУ»хОпґ©№эД¤µДФЛКдЎЈ°Лѕ«°±ЛбЈЁR8Ј»±н1Ј©ЎўD-R8єНR11ТС±»УГЧчСЄДФЖБХПґ©ЛулДЈ¬УГУЪПтґуДФµЭЛНТ©ОпЎЈR8єНD-R8ФЪѕІВцБЄєПёшТ©D-R8єНТИµєЛШ»мєПОпєуЈ¬ПФКѕФцЗїБЛТИµєЛШФЪґуКуґуДФЙµДЙгИЎєН»эАЫЎЈR11лДЈЁ±н1Ј©їЙТФґ©№эХэіЈµДНкХыСЄДФЖБХПЈ¬µЅґпЖ¤ЦКЎўОЖЧґМеєНЗрДФЎЈR11 ЅйµјµДДФІїµЭЛНФЪПµНіёшТ©єуі¤ґп 8 РЎК±µДИ±СЄРЎКуґуДФЦеõЅБЛј«ґуФцЗїЎЈR18 єН D РНї№Ф D-R18 ФЪІ»Н¬ДціЭ¶ЇОпЦР·зДЈРНЦРПФКѕіцЙсѕ±Ј»¤МШРФЈ¬НЁ№эјхЗбИ±СЄРФДФЛрЙЛµДСПЦШіМ¶ИєНёДЙЖ№¦ДЬЅб№ыЎЈХвР©·ўПЦ±нГчЈ¬ѕЫѕ«°±ЛблДКЗєЬУРЗ°ѕ°µДФШМеЈ¬УГУЪїзФЅСЄДФЖБХПµЭЛНЦОБЖТ©ОпЈ¬ѕЯУРЗ±ФЪµДЙсѕ±Ј»¤ЧчУГЎЈ
6 | ґ©НёґуДФµДPDC
ї№МеТ©ОпЕјБЄОпЈЁADCЈ©µД°РПтЦОБЖТСЅшИлБЩґІУ¦УГЈ¬Хвґъ±нБЛНЁ№э°РПтІї·Цї№МеСЎФсРФµШПтЦЧБцКдЛНЎ°ЙсЖжЧУµЇЎ±ХвТ»ёЕДоµДіЙ№¦ЎЈїЙТФЙијЖіцёьРЎµД·ЦЧУґуРЎµДлДЈ¬ТФСЎФсРФµШУлПё°ы±нГж°РµгЅбєПЈ¬ІўК¶±р»јІЎЧйЦЇµД¶АМШ·ЦЧУ°РµгЎЈPDC ЦјФЪНЁ№э»ЇС§Б¬ЅУЧУЅ«°РПтлДУлЦОБЖУРР§ФШєЙБ¬ЅУЖрАґЈЁНј 5Ј©Ј¬КЗ ADC µДМжґъЖ·Ј¬УРїЙДЬА©ґуёчЦЦТ©ОпµДЦОБЖЗ±Б¦ЎЈФЪСЄДФЖБХПЈЁBBBЈ©ґ©ФЅєН°РПтЦРКаЙсѕПµНіјІІЎµДЗйїцПВЈ¬PDC ЦјФЪЅЩіЦДЪФґРФ BBB БчИлФЛКд»ъЦЖЈ¬ІўЅ«Т©ОпНµФЛµЅДФКµЦКЦРЎЈЦОБЖјБїЙТФНЁ№э»ЇС§·Ѕ·ЁУл BBB ґ©ЛулДЕјБЄЎЈ
Нј 5 лДТ©ОпЕјБЄОпЈЁPDCЈ©µДЅб№№ТФј°іЈУГµДБ¬ЅУЧУєНґъ±нРФµДТ©ОпУРР§ФШєЙЎЈPABC ±нКѕ¶Ф°±»щЬРСхфК»щЎЈТ©ОпЕјБЄО»µгТФємЙ«ФІИ¦ПФКѕЎЈ
Улї№МеТ©ОпЕјБЄОпЈЁADCsЈ©Па±ИЈ¬µН·ЦЧУБїµДлДЕјБЄТ©ОпЈЁPDCsЈ©їЙДЬµјЦВёьѕщФИµДО»µгМШТмРФ»ЇС§ЕјБЄЎўФцЗї¶Ф№ММеЧйЦЇµДЙшНёЎўУРР§µДПё°ыФЛКдЈ¬ТФј°ёьјт±гµДєПіЙєНґї»ЇЎЈPDC µДТ»ёцЗ±ФЪИ±µгКЗЈ¬¶ФлДєНЦОБЖјБµД»ЇС§РЮКОїЙДЬµјЦВЖЪНыµДЦРКаЙсѕПµНіЈЁCNSЈ©»оРФµДЙҐК§ЎЈґЛНвЈ¬НЁ№эУлЗЧЦ¬РФлДЕјБЄАґФцјУ·ЦЧУµДЗЧЦ¬РФТІїЙДЬµјЦВіЙОЄНвЕЕ±ГЈЁИз P-gpЈ©µДµЧОпЈ¬АэИз GabathulerЈЁ2010 ДкЈ©ЎЈУ°Пм PDC ЧчОЄСЄДФЖБХПЈЁBBBЈ©їЙЙшНё·ЦЧУФШМеєНУРР§ЦОБЖТ©ОпіЙ№¦µДЦчТЄТтЛШ°ьАЁЈєЈЁaЈ©PDC ПтґуДФФЛКдµДЛЩ¶ИЈ¬ЈЁbЈ©УРР§ФШєЙПтґуДФКН·ЕєНµЭЛНµДіМ¶ИЈ¬ЈЁcЈ©ДФДЪТ©Оп·ЦІјµДТ©ґъ¶ЇБ¦С§Ј¬ЈЁdЈ©»оРФТ©Оп¶Ф CNS °РµгµчЅЪµДҩЧѧЈ¬ЈЁeЈ©УлДЪФґРФµ°°ЧЦК»тлД¶Ф BBB ЧЄФЛµ°°ЧµДѕєХщЈ»(f) УлУРР§ФШєЙКН·ЕПа№ШµДБ¬ЅУМе»ЇС§МШРФТФј°Пё°ыНвєНПё°ыДЪОИ¶ЁРФЈ¬ТФј° (g) ЦОБЖјБµДСЎФсЈ¬°ьАЁР§Б¦ЎўЧчУГ»ъЦЖєНїЙРЮКОµД№¦ДЬ»щНЕЎЈЧРПёОўµч PDC µДМШРФЈ¬ТФЖЅєвСЄДФЖБХПФЛКдєНУРР§ФШєЙµДЦРКаЙсѕПµНі»оРФЈ¬¶ФУЪКµПЦ PDC ЧчОЄ·ЗЗЦИлРФСЄДФЖБХПµЭЛНІЯВФµДЗ±Б¦ЦБ№ШЦШТЄЎЈ
 Нј 6 ДФґ©НёРФ PDCsЈЁлДЈєєЪЙ«Ј¬Б¬ЅУЧУЈєємЙ«Ј¬ЦОБЖУРР§ФШєЙЈєА¶Й«Ј©µДЅб№№ЎЈСЄДФЖБХПґ©ЛулД°ьАЁ Angiopep-2ЎўSynB1ЎўSynB3ЎўPenetratinЎўTP10ЎўRGDЎўgHoPe2ЎўGSH єН E1-3ЎЈЦОБЖУРР§ФШєЙ°ьАЁЧПЙјґјЎўТАНРІґЬХЎў°ўГ№ЛШЎўВр·ИЎўВр·И-6-¦В-ЖПМСМЗИ©ЛбхҐЎўї§·ИЛбЎўPD0325901ЎўЬР»щЗаГ№ЛШЎўНт№ЕГ№ЛШЎў¶а°Н°·ЎўГАЅрёХЈЁMEMЈ©ЎўґпАѕ«єНЙсѕµЭЦКЎЈ
Нј 6 ДФґ©НёРФ PDCsЈЁлДЈєєЪЙ«Ј¬Б¬ЅУЧУЈєємЙ«Ј¬ЦОБЖУРР§ФШєЙЈєА¶Й«Ј©µДЅб№№ЎЈСЄДФЖБХПґ©ЛулД°ьАЁ Angiopep-2ЎўSynB1ЎўSynB3ЎўPenetratinЎўTP10ЎўRGDЎўgHoPe2ЎўGSH єН E1-3ЎЈЦОБЖУРР§ФШєЙ°ьАЁЧПЙјґјЎўТАНРІґЬХЎў°ўГ№ЛШЎўВр·ИЎўВр·И-6-¦В-ЖПМСМЗИ©ЛбхҐЎўї§·ИЛбЎўPD0325901ЎўЬР»щЗаГ№ЛШЎўНт№ЕГ№ЛШЎў¶а°Н°·ЎўГАЅрёХЈЁMEMЈ©ЎўґпАѕ«єНЙсѕµЭЦКЎЈ
ФЪХвЖЄЧЫКцЦРЈ¬ОТГЗСРѕїБЛСЄДФЖБХПґ©ЛулДєНДФґ©НёРФ PDCs ЧчОЄ·ЗЗЦИлРФТ©ОпµЭЛНПµНіЈ¬УГУЪЅ«»ЇБЖТ©ОпєНЖдЛы·ЦЧУ»хОпґ©№эПЮЦЖРФµДСЄДФЖБХПЎЈФзЖЪµДСЄДФЖБХПґ©ЛулДЈ¬ИзПё°ыґ©НёлДЈ¬ЦчТЄАыУГСЎФсРФЅПІоµДОьёЅЅйµјµДЧЄ°ыНМЧчУГАґёДЙЖґ©№эСЄДФЖБХПµДФЛКдЈ¬¶шНЁ№эКЙѕъМеХ№КѕјјКх»тґУМмИ»ЙсѕУЄСшµ°°ЧЦР·ўПЦµДРВРНлДФтАыУГКЬМеЅйµјµДЧЄ°ыНМЧчУГ·ЦЧУ»ъЦЖАґґ©№эСЄДФЖБХПЎЈ
