
ХҒТҒӘғғПіЙлД»щЙъОпІДБПФЪРн¶аЗйүцПВДЬ№»ДӘ·ВЖдИ«і¤ДЪФөРФ¶ФУ¦ОпµДҢб№№ғН№¦ДЬҰӘФЩәУЙПУлИ«і¤µ°°ЧЦКПа±ИӘ¬¶МДӘДвлДёьИЭТЧЙъІъӘ¬±нПЦіцёьЗүµДүЙәУ№¤РФғНТЧРЮКОРФӘ¬ІұЗТДЬ№»ФЪГчИ·¶ЁТеғНКЬүШµДМхәюПВЦЖ±ёӘ»ғЬГчПФӘ¬ОҒКІГөЧоҢьИЛГЗИИЦФУЪөУХвР©·ЦЧУЦРүҒ·ұФЩЙъЙъОпІДБПҰӘФҢАөФҢ¶аµДЦ¤ңЭ±нГчӘ¬Ң«лДДЙИлФЩЙъЦ§әЬүЙТФІъЙъҢб№№К¶±р»щРтӘ¬өУ¶шФцЗүПё°ыёҢЧЕ»тУХµәПё°ыРЕғЕНЁВ·Ә¬ёДЙЖПё°ыҢюИу»төЩҢшёчЦЦЖдЛыµчҢЪРФЙъ»Ү·өУ¦ҰӘНЁ№эЗүµч¶МДӘДвлДµДЙиәЖғНУ¦УГµДµ±З°·Ң·ЁӘ¬ОТГЗПӘНыХ№КңЛьГЗФЪИнЧйЦҮУъғПЦРµДЗ±Б¦Ә¬Н¬К±МбЗл№ШЧұЖщҢсОҒЦ№ИҰµГµДҢшХ№ТФә°РиТҒүЛ·юµДОКМвӘ¬ТФНЖ¶ҮХвР©ІДБПФЪБЩөІЙПУ¦УГУЪРДФаҰұЖ¤·фғНҢЗД¤РЮёөҰӘ
УГУЪЧйЦҮРЮёөµД·ВЙъІДБПТ»Ц±КЗЙъОпІДБПСРңүБмУтЦРМҢЛчЧоЙоИлµДБмУтЦ®Т»Ә¬µ«ДӘ·ВМмИ»Пё°ыНв»щЦК (ECM) ИФИ»КЗЧйЦҮ№¤іМЦРЧоңЯМфХҢРФµДИООсЦ®Т»ҰӘөЛНвӘ¬ЛдИ»ЦШЧйµ°°Ч±нөпИҰµГБЛПФЦшҢшХ№Ә¬µ«ИФөжФЪІоңаӘ¬ТтОҒХвР©№эіМИФИ»Па¶Ф°ғ№уӘ¬МШ±рКЗ¶ФУЪТмФөµ°°Ч¶шСФӘ»ТтөЛПЮЦЖБЛүЖС§әТК№УГ¶ҮОпАөФөӘЁАэИзөУЧйЦҮЦРМбИҰӘ©µДµ°°ЧЦКӘЁ°ьАЁҢғФµ°°ЧғНµҮРФµ°°ЧӘ©АөЙиәЖүЙЧҒ»ҮµДЙъОпІДБПҰӘХвТ»ПЮЦЖСПЦШЧи°БЛ№¦ДЬРФЙъОпІДБППтБЩөІµДБЩөІЧҒ»ҮҰӘЦЪЛщЦЬЦҒӘ¬µ°°ЧЦКФЪәёғхЛщУРµДЙъОп№эіМЦР¶ә·ұ»УЧЕЦШТҒЧчУГҰӘЛдИ»ЛьГЗФЪПё°ыµДҢб№№НкХыРФЦР·ұ»УЧЕГчИ·µДЧчУГӘ¬Жч№ЩғНЧйЦҮӘ»ЛьГЗФЪПё°ыФЛ¶ҮҰұРЕғЕЧҒµәҰұГвТЯ·өУ¦ғНГё·өУ¦µИЖдЛы№эіМЦРµДЧчУГёьәУ¶ҮМ¬ҰӘТтөЛӘ¬Рн¶аУР№ШЙЛүЪУъғПғНЧйЦҮРЮёөµДЦШТҒ·ұПЦ¶әАөЧФ¶Фµ°°ЧЦК-µ°°ЧЦКғНµ°°ЧЦК-ЕдМеПа»ӨЧчУГµДСРңүҰӘЖдЦРТ»По·ұПЦКЗӘ¬НЁіӘУләІІҰ»тЙЛүЪУъғППа№ШµДңЯУРµ°°ЧЦКҢбғПО»µгµД·ЦЧУКЗүҒ·ұЦОБЖ·Ң°ёµДңшәСДү±кҰӘәшУЪ»ҮС§лДғПіЙғНИ«і¤µ°°ЧЦКғНлДµДЦШЧйЙъІъ·ҢГжµДЧоРВҢшХ№Ә¬ЙъіЙғНСРңүДӘДв·ЦЧУµДПа»ӨЧчУГТСөуөуәт»ҮҰӘОЮВЫКЗЙъІъИ«і¤»тЖ¬¶ОµДµ°°ЧЦКµДң«И·ё±±ңӘ¬»№КЗІфИл·З±аВл°±»щЛбӘ¬»тРЮКОлД№ЗәЬТФФцЗүЖдµ°°ЧЛ®ҢвОИ¶ЁРФ»тәУИлПµБөТФҢшТ»ІҢ№¦ДЬ»ҮӘ»ИЛГЗНЁіӘ¶әүЙТФХТµҢғПККµД·Ң·ЁАөЙъІъЛщРиµД·ЦЧУҰӘіцУЪХвР©ФТтӘ¬лДЧчОҒµ°°ЧЦКµДРҰЧйіЙІү·ЦӘ¬ТССёЛЩіЙОҒүҒ·ұУГУЪЧйЦҮРЮёөµД№¦ДЬІДБПµДңәГУРР§µДМжөъЖ·ҰӘ
ЛдИ»ЙиәЖСҰПоәёғхКЗОЮПЮµДӘ¬µ«ХвР©ДӘДвОпНЁіӘНЁ№эіКПЦМШ¶ЁµД°±»щЛбРтБРҰұ№¦ДЬҢб№№»тБҢХЯµДЧйғПАөУлДү±кПа»ӨЧчУГҰӘФЪ±ңЖҒЖАВЫЦРӘ¬ОТГЗҢ«ЦШµгҢйЙЬК№УГ№МПалДғПіЙ (SPPS) ЦЖ±ёµДлДҢб№№Ә¬¶ФУЪµ±Ңсөу¶аКэПµНіАөЛµӘ¬ёГ№эіМКЗФЪС»·ЧФ¶ҮғПіЙЙи±ёЦРҢшРРµДӘ¬ЖдЦРлДҢб№№ЦРµДГүёц°±»щЛб±»ЛіРтµШІфИлҰӘУРРЛИ¤БЛҢвёь¶аУР№ШК№УГЧҒ»щТтЙъОпҢшРРлДғПіЙµД¶БХЯӘ¬ОТГЗ№ДАшЛыГЗС°ХТ№ШУЪХвёцМШ¶ЁЦчМвµДЖАВЫҰӘ
SPPS µДёЕДоРОіЙУЪ 1959 ДкӘ¬ІұУЪ 20 КАәН 60 ДкөъіхУЙЕµ±ө¶ыҢ±»сµГХЯ Robert Bruce Merrifield КЧөО±ЁµАӘ¬ЛжЧЕлД»ҮС§әәКхµДҢшІҢК№ёГ№эіМёьәУОИҢҰӘ¬SPPS ФЪ 20 КАәН 70 ДкөъғН 80 ДкөъЦрҢӨБчРРІұ±»ЦчБчІЙУГҰӘНЁ№эҢ«лДБөУлПВТ»ёцБ¬РшaaµДфИ»щД©¶ЛЕәБҒӘ¬өУ¶шСУі¤лДБөµДёЕДоі№µЧёД±дБЛлДБөµДЙъІъ·ҢКҢӘ¬¶шПВТ»ёцБ¬РшaaµДЖдЛы№¦ДЬ»щНЕКЬµҢ±Ә»¤ҰӘХвР©±Ә»¤»щүЙ·АЦ№ aaІаБө·ұЙъСх»ҮғН·ЗМШТмРФ·өУ¦ҰӘКэК®ДкАөµДЙоИлғПіЙСРңүТСңІъЙъБЛРн¶а¶а№¦ДЬ±Ә»¤»щӘ¬АэИзФРНµДЬМәЧСхфК»щ (Fmoc) »щНЕҰӘөшУРµӨёц°±»щЛбµД Fmoc »щНЕФЪКРіҰЙПғЬИЭТЧВтµҢҰӘSPPS µДәәКхёДҢшӘ¬АэИзК№УГОұІЁ·өУ¦ЖчғНПИҢшµДОВ¶ИүШЦЖПµНіӘ¬УлКТОВғН¶ФБчәУИИ·Ң·ЁПа±ИӘ¬үЙТФғПіЙғ¬УРКэ°Щёц°±»щЛбµДлДӘ¬ІъБүёьёЯӘ¬·өУ¦К±әдёь¶МӘ¬2014 ДкХвР©әәКхҢшІҢТІөЩҢшБЛЛщОҢА§ДСлДРтБРµДүмЛЩғПіЙҰӘА§ДСлДРтБРКЗЦёФЪғПіЙ№эіМЦР»тИӨіэ±Ә»¤»щғуңЫәҮРОіЙІ»ИЬРФІъОпµДлДРтБРӘ¬ХвТ»№эіМµәЦВІъБүҢµµН»тлДК§»оӘ¬өУ¶шЧиЦ№ҢшТ»ІҢРЮКОҰӘФЪөу¶аКэЗйүцПВӘ¬ХвР©ОКМвКЗУЙУЪТэИлБЛДЬ№»ІОУл·З№ІәЫПа»ӨЧчУГµД№¦ДЬРФ¶шІъЙъµДӘ¬АэИзЗвәьғНЕәә«-Еәә«Па»ӨЧчУГҰӘТтөЛӘ¬ФЪОҒ SPPS ЙиәЖлДК±Ә¬µӨёц°±»щЛб. У¦ЙёСҰЛщµГЕәБҒІъОпӘЁФЪКчЦ¬ЙПӘ©Ә¬ТФИ·¶ЁКЗ·сүЙДЬРОіЙЧФЧйЧ°Ңб№№ҰұКЗ·с·ұЙъё±·өУ¦ТФә°КЗ·сЗгПтУЪХЫµюµҢКчЦ¬ЙПҰӘТСЦ¤ГчТФПВІОКэУРЦъУЪғПіЙА§ДСµДлДРтБРӘғ(i) ёЯОВӘЁАэИзӘ¬ОұІЁёЁЦъғПіЙК±ОҒ 95ҰгCӘ©Ә¬(ii) өжФЪСО»тИӨ№ёәБТФМбёЯИЬҢв¶ИӘ¬(iii) ФЪхӘ°·»щНЕө¦К№УГ±Ә»¤»щТФ±ЬГвЗ±ФЪµДЗвәьПа»ӨЧчУГӘ¬(iv) ҢбғПңЯУР·З·өУ¦РФІаБөµД°±»щЛбТФ·АЦ№·ұЙъІ»БәПа»ӨЧчУГӘ¬ТФә° (v) МЗ»щ»Ү»тңЫТТ¶юөә»ҮТФМбёЯлДИЬҢв¶ИҰӘ
ЙъОп»оРФлДРтБРДӘДвОп
К№УГлДРтБРүҒ·ұµДҢб№№ДӘДвОпКµәКЙПКЗЙъОп»оРФО»µгµД±нО»Ә¬ФЪРн¶аЗйүцПВӘ¬ДӘДвОпµДК¶±рО»µгУЙ°±»щЛбРтБРғНИэО¬№№Пу¶ЁТеҰӘФЪТФПВХВҢЪЦРӘ¬ОТГЗҢ«МЦВЫТ»Р©ТС±»К¶±рҰұғПіЙғН/»тХығПµҢҢб№№ЦРµДЙъОп»оРФ¶МлДРтБРӘ¬ТФІъЙъДіЦЦАаРНµДЙъОп·өУ¦Ә¬үЙУГУЪЧйЦҮ№¤іМ»тФЩЙъЙъОпІДБПµДүҒ·ұҰӘҢ«ТҒМЦВЫµДлДРтБР¶ФУ¦УЪТ»Чйөъ±нРФµДАэЧУӘ¬ОТГЗИПОҒХвР©АэЧУКфУЪТФПВИэАаЦ®Т»Ә¬ОТГЗИПОҒХвИэАаФЪүҒ·ұУГУЪРДФаҰұЖ¤·фғНҢЗД¤ЧйЦҮФЩЙъµД№¦ДЬРФЙъОпІДБП·ҢГжЧоОҒЦШТҒӘ¬әөӘғ(i) өЩСҒ№ЬЙъіЙРтБРҰұ(ii) ү№СЧРтБРғН (iii) өЩХіёҢРтБРҰӘ±н 1°ьғ¬Т»Р©ТСәш¶ЁІұУГУЪЦЖФ칦ДЬІДБПµДөъ±нРФлДРтБРҰӘ·Ң°ё 1Ги»жБЛ±ңЧЫКцЦРРЮ¶©µДёЕДоµДөъ±нРФХҒТҒҰӘЛдИ»ОТГЗҢ«±ңЧЫКцПЮЦЖФЪРДФаҰұЖ¤·фғНҢЗД¤µИИнЧйЦҮДү±к·ҢГжµДЧЁТµЦҒК¶Ә¬µ«ЦµµГТ»МбµДКЗӘ¬лД»щІДБПТІТСУ¦УГУЪ№ЗчАғНСАіЭµИУІЧйЦҮµДФЩЙъӘ¬УР№ШХвР©У¦УГғНЧоРВҢшХ№µДёь¶аРЕПұүЙТФФЪёьЧЁТµµДЧЫКцЦРХТµҢҰӘ
өЩСҒ№ЬЙъіЙРтБР
СҒ№ЬЙъіЙКЗТ»ёцЙжә°ДЪЖ¤Пё°ыФцЦіғНЗЁТЖТФә°Н¬К±ҢшРРµДПё°ыНв»щЦКЦШЛЬµД№эіМӘ¬өУ¶шНЖ¶ҮРВСҒ№ЬөУПЦУРСҒ№ЬЦР·ұУэҰӘЙъАнғНәІІҰЧөМ¬ПВСҒ№ЬЙъіЙµДЦчТҒµчҢЪТтЧУКЗСҒ№ЬДЪЖ¤Йъі¤ТтЧУ (VEGF)ҰӘVEGF ә°Жд¶аЦЦН¬№¤РННЁ№эУлИэЦЦТСПкПёГиКцµДАТ°±Лбә¤ГёКЬМеПа»ӨЧчУГАөУХµәЙъАн·өУ¦ӘЁSimons µИИЛӘ¬2016 ДкµҢДүЗ°ОҒЦ№Ә¬ЧоУРЗ°НңµДөЩСҒ№ЬЙъіЙлДКЗ VEGF ДӘДвОпҰӘФЪПЦУРµД VEGF ДӘДвОпЦРӘ¬ЧоµдРНµДКЗлДQKӘ¬Ль±»ЙиәЖУГАөДӘДв VEGF НЁ№эУЙ°±»щЛбРтБР KLTWQELYQLKYKGI ЧйіЙµД N ¶Л ¦Б ВЭРэДӘДвОпУлЖдКЬМеµДҢбғПҰӘлД QK µДСҒ№ЬЙъіЙМШРФТСФЪМеНвғНМеДЪµГµҢЦ¤КµӘ¬ңЯУРГчПФµДДЪЖ¤Пё°ы»о»ҮЧчУГғН VEGF Па№ШПё°ы№¦ДЬµДФцәУӘ¬ИзЗч»ҮРФҰұЗЦП®РФҰұРВГ«ПёСҒ№ЬµД·ұСүғНФцЗүµДЧйЦҮ»ҮӘ¬°ьғ¬ QK РтБРµДЧФЧйЧ° ¦В Ж¬ІглДЛ®ДэҢғФЪөуКуДӘРНЦРЖ¤ПВЧұЙдК±ТІПФКңіцөЩҢшПё°ыҢюИуғНСҒ№Ь»ҮӘ¬ИзНә1ЛщКң
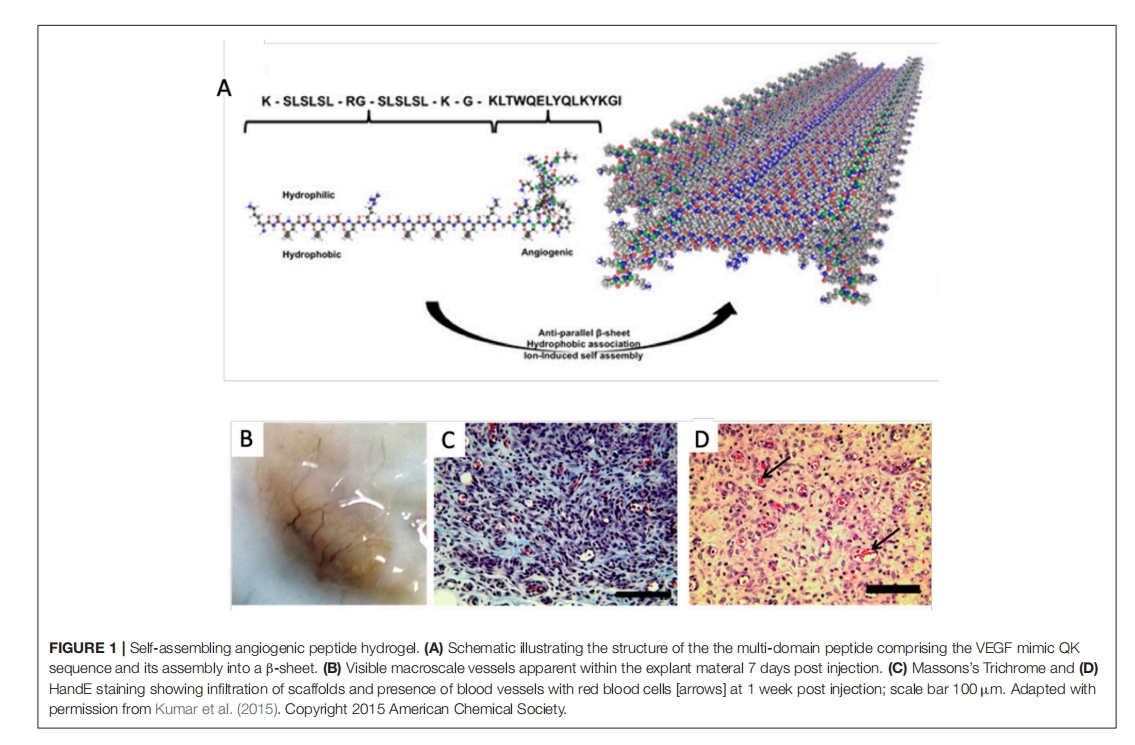
МҢЛчХвР© VEGF АаЛЖОпҢбғПёОЛШµДДЬБ¦ТІүЙДЬғЬУРИ¤Ә¬ТтОҒУРөуБүОДПЧ±ЁµАБЛІ»Н¬ VEGF СЗРНҢбғПёОЛШµДЗгПтТФә°ХвЦЦҢбғП¶ФөЩҢшДЪЖ¤Пё°ыЙъі¤ғНФцЦіµД±ШТҒРФҰӘөЛНвӘ¬Ң« VEGF ІфИлІъСх»тИ±СхУХµәЛ®ДэҢғ»т»щЦКЦРүЙДЬ»бУ°ПмИ±СхУХµәТтЧУӘ¬ХвР©ТтЧУ¶Ф VEGF µД±нөпғН№¦ДЬғЬЦШТҒҰӘ
ЖдЛыФЪµчҢЪСҒ№ЬЙъіЙ·ҢГжПФКңіцЗ°ң°µДлДКЗЙъі¤ТтЧУКЬМеµД°Р±кӘ¬ХвР©КЬМеНЁіӘУл VEGF РН¬ЧчУГҰӘАэИзӘ¬іЙПЛО¬Пё°ыЙъі¤ТтЧУТФә°ЙсңПё°ыХіёҢ·ЦЧУ (NCAM) ТС±»Ц¤ГчүЙТФУліЙПЛО¬Пё°ыЙъі¤ТтЧУКЬМеҢбғПӘ¬ТІүЙТФөЩҢшСҒ№ЬЙъіЙҰӘFGF2 ғН NCAM µДлДДӘДвОпТСНЁ№эғПіЙЦЖ±ёӘ¬ЛдИ»ЛьГЗүЙДЬТФµдРН»т·ЗµдРН·ҢКҢ·ұ»УЧчУГӘ¬µ«УРЗүУРБ¦µДЦ¤ңЭ±нГчЛьГЗ»бУ°ПмСҒ№ЬЙъіЙҰӘ»№УРРн¶аСҒ№ЬЙъіЙЛШ-1 ДӘДвОпӘ¬ЛьГЗНЁ№эУлАТ°±Лбә¤ГёКЬМе (Tie2) Па»ӨЧчУГӘ¬УРНыµчҢЪСҒ№ЬЙъіЙӘ¬¶шАТ°±Лбә¤ГёКЬМеЦчТҒөжФЪУЪСҒ№ЬДЪЖ¤Пё°ығНФмСҒПё°ыЦР ҰӘИЛГЗүЙДЬПӘНыФЪУГУЪСҒ№Ь»ҮЧйЦҮµДІДБПЦРәУИлµДЖдЛылДКЗДЬ№»ДӘДвЧҒ»ҮЙъі¤ТтЧУ (TGF¦Б ғН TGF¦В)ҰұЦЧБц»µЛАТтЧУ (TNF¦Б)ҰұСҒ№ЬЙъіЙЛШҰұ°ЧПё°ыҢйЛШ 8 (IL8) »тёОПё°ыЙъі¤ТтЧУ ( HGF) Ә¬ТтОҒХвР©УРЛү·ЦБСФғНЗч»ҮТтЧУТС±»Ц¤КµүЙНЁ№эүШЦЖДЪЖ¤Пё°ыЙъі¤ғН/»тУл VEGF ҢйµәНңң¶Па»ӨЧчУГАөөЩҢшСҒ№ЬЙъіЙҰӘүәВЗµҢХвР©ТтЧУ¶Ф VEGF ±нөпғН№¦Р§µДУ°ПмӘ¬ТФә°ЛьГЗНЁіӘКЗ±нГжҢбғПКЬМеµД°Р±кӘ¬Ң«ЛьГЗІфИлИнІДБПЦРүЙДЬРиТҒТФІ»·Б°ЛьГЗУл°Р±кКЬМеПа»ӨЧчУГµД·ҢКҢҢшРРӘ¬ХвүЙДЬ»бПЮЦЖ»щЦКДЪµД№ІәЫБ¬ҢУҰӘ
ү№СЧРтБР
ФЪЙиәЖУГУЪЧйЦҮ№¤іМғНФЩЙъµДЦ§әЬғНЙъОпІДБПК±Ә¬ЛЮЦчГвТЯПµНіКЗРиТҒүЛ·юµДЧоөуХП°Ц®Т»ҰӘИ»¶шӘ¬ХвІұІ»ТвО¶ЧЕТҒНкИ«±ЬГвГвТЯ·өУ¦Ә»КВКµЙПӘ¬ОҒБЛЧоөуПЮ¶ИµШМбёЯЦІИлОпµДЦОБЖР§№ыӘ¬ЛьГЗ±ШРлµчҢЪУЙөЛІъЙъµДГвТЯ·өУ¦ҰӘөЛНвӘ¬СЧЦұ»бөЩҢшСҒ№ЬЙъіЙӘ¬РВСҒ№ЬµДРОіЙ»бµәЦВҢшТ»ІҢµДСЧЦұҰӘТтөЛӘ¬ЦШТҒµДКЗТҒБЛҢвСЧЦұКЗТ»ёцёөФУµД№эіМӘ¬ЛьЧоЦХНЁ№эөЩҢшПё°ыҢюИуҰұФцЦіғНЛжғуµДә«»ҮОҒКЬУ°ПмµДЧйЦҮөшАөМеДЪЖҢғвҰӘЛдИ»ГвТЯ·өУ¦Йжә°Рн¶аТтЛШӘ¬ңЮКЙПё°ы±»ИПОҒКЗЧоЦШТҒµДПё°ыЦ®Т»Ә¬ТтөЛ±ңҢЪҢ«ЦШµгҢйЙЬлДДӘДвОпИзғО»түЙДЬУГУЪµчҢЪңЮКЙПё°ыҰӘңЮКЙПё°ыКЗ¶ҮМ¬Пё°ыӘ¬Жд±нРНКЬПё°ыНв»·ңіТФә°»оРФРЕғЕ·ЦЧУҰӘө«НіЙПӘ¬ФЪМЦВЫңЮКЙПё°ыµД±нРНК±Ә¬ңЭЛµУРБҢёцІ»Н¬µДСЗИғ (i) M1ӘЁөЩСЧӘ©ғН (ii) M2ӘЁү№СЧ/өЩУъғПӘ©ҰӘИ»¶шӘ¬ХвКЗТ»ЦЦёЯ¶Иәт»ҮµД№ЫµгӘ¬ТтОҒіҮПтИОТ»±нРНµДә«»ҮКµәКЙПёьПсКЗТ»ёцБ¬РшМеӘ¬M1 ғН M2 Ц®әдµДІоТмІ»КЗАлЙұµДҰӘНЁ№эЙиәЖУл M2 ңЮКЙПё°ыµДГвТЯФРФКЬМеӘЁИз TGF-bRҰұIL-4RҰұIL-6RҰұIL-10R ғН MCSFRӘ©Па»ӨЧчУГµД¶МлДӘ¬үЙТФµчҢЪУлЧйЦҮЛрЙЛғНРЮёөТФә°ТэИлТмОпПа№ШµДГвТЯ·өУ¦ҰӘНЁ№эЧчУГУЪөЩСЧғНү№СЧПё°ыТтЧУӘЁИз IL 6 ғН TNF-¦БӘ©µД±нөпТФә°»оРФСхµДІъЙъӘ¬үЙТФүҒ·ұіцДЬ№»әхЙЩСЧЦұҰұНЁ№эЗч»ҮЧчУГДәәҮПё°ыІұЧоЦХёДЙЖЙЛүЪУъғПµДІДБПҰӘЛдИ»өу¶аКэУлТмОп·өУ¦Па№ШµДСЧЦұүЙТФНЁ№эК№УГЦШЧй»тЧФМеµ°°ЧЦК»тµ°°ЧЦК/лДДӘДвОпАөПыіэ»тәхЙЩӘ¬µ«үЙДЬ°ьАЁРҰлДДӘДвОпАөә¤»оғНә«»ҮңЮКЙПё°ыӘ¬К№ЖдПт 2 РН±нРН·ұХ№ҰӘИ»¶шӘ¬УЙУЪә¤»о№эіМµДёөФУРФӘ¬ғЬДСИ·¶ЁТ»ёц»т¶аёцүЙТФөшАөЛщРи·өУ¦µДРтБРӘ»ТтөЛӘ¬ДүЗ°ТСЦҒµДµчҢЪГвТЯ·өУ¦µДРтБРІұІ»¶аӘ¬ХвР©РтБРУРАыУЪЧйЦҮФЩЙъғНФЩЙъЙъОпІДБПµДЙиәЖҰӘ»№УРРн¶аРтБР±»¶ЁТеОҒү№ңъғНү№СЧӘ¬Т»АаФЪµчҢЪГвТЯ·өУ¦·ҢГжПФКңіцБәғГЗ°ң°µДлДКЗПИМм·АУщµчҢЪлД (IDR)ҰӘХвР©СфАлЧУү№ңълДКЗМмИ»өжФЪµДЛЮЦч·АУщлД»тµ°°ЧЦК (HDP) µДғПіЙСфАлЧУАаЛЖОпҰӘЛьГЗКЗПа¶ФҢП¶МµДлДӘЁ10-50 ёц°±»щЛбӘ©Ә¬Г»УРМШ¶ЁµД№ІК¶РтБРҰӘЛдИ»ЛьГЗңЯУРЦ±ҢУЙ±ЛАОұЙъОпµДДЬБ¦Ә¬µ«ЛьГЗТІДЬ№»µчҢЪГвТЯғНСЧЦұ·өУ¦ҰӘАэИзӘ¬ЛьГЗДЬ№»У°ПмЗч»ҮРФҰұөМә¤Зч»ҮТтЧУµДІъЙъҰұТэµәңЮКЙПё°ыә«»ҮТФә°µчҢЪЦРРФБӘПё°ыХіёҢғН»о»Ү±кЦңОпµД±нөпҰӘIDR-1018 КЗөЛАалДЦРУЙ 12 ёц°±»щЛбЧйіЙµДлДҰӘӘЁVRLIVAVRIWRR-NH 2Ә©Ә¬ТСЦ¤КµДЬ№»ФцЗүү№СЧ·өУ¦Ә¬Н¬К±О¬іЦ¶Фү№ёРИңµДЦШТҒөЩСЧ№эіМӘ¬ХвЦЦДЬБ¦КЗТтОҒЛьДЬ№»Зэ¶ҮңЮКЙПё°ыә«»ҮЦБЦРәд M1-M2 ЧөМ¬ҰӘөЛАалДµДЖдЛыіЙФ±°ьАЁ IDR-HH2 ғН IDR-1002Ә¬ЛьГЗ¶әңЯУРПаЛЖµДГвТЯµчҢЪДЬБ¦ҰӘү№ңълД LL-37 ғН SET-M33 ТІТСЦ¤КµДЬ№»НЁ№эәхЙЩөЩСЧПё°ыТтЧУҰұГёғНЧҒµәТтЧУАөҢйµәСЧЦұҰӘ
ңЮКЙПё°ы»о»ҮүШЦЖГвТЯ·өУ¦µД·ҢКҢЦ®Т»КЗНЁ№э±нөп/ІъЙъ»щЦКҢрКфµ°°ЧГё (MMP)ҰӘMMP КЗТ»Ааµ°°ЧЛ®ҢвГёӘ¬ЛьГЗ±ңЙнДЬ№»НЁ№эµчҢЪПё°ыТтЧУғНЗч»ҮТтЧУАөµчҢЪГвТЯ·өУ¦ҰӘMMP УР¶аЦЦІ»Н¬µДАаРНӘ¬ЛьГЗ¶әДЬ№»ҢµҢвПё°ыНв»щЦКµ°°ЧІұНЁ№эµ°°ЧЛ®ҢвБСҢв»тЖдЛыРЮКОә¤»оЙъОп»оРФ·ЦЧУҰӘНЁ№эФЪ№№іЙІДБПµДлДДЪәУИл MMP ҢбғПғНБСҢвРтБРӘ¬үЙТФФцәУЖдңЦІүЕЁ¶ИӘ¬Н¬К±ФцЗүІДБПµДµ°°ЧЛ®ҢвҢµҢвӘ¬өУ¶шүЙТФУГДЪФөРФ»щЦКМж»»ІДБПІұКН·ЕРҰлДЖ¬¶ОӘ¬Ңш¶шµчҢЪЖдЛыПё°ы·өУ¦ҰӘI РНҢғФµ°°ЧЦР·ұПЦµД MMP ±нО»µДТ»ёцАэЧУКЗ°±»щЛбРтБР GPQGIAG. СРңү±нГчӘ¬ҢғФµ°°Ч-PEG ёөғПОпЦРөжФЪөЛАаРтБРүЙФцЗү MMP-1 ғН MMP-2 µДµ°°ЧЛ®ҢвҢµҢвЧчУГҰӘ
өЩТАөУРтБР
ФЩЙъЙъОпІДБПµД№ШәьТҒЗуЦ®Т»КЗЦ§іЦДЪФөРФПё°ыµДЙъі¤ҰұёҢЧЕғНФцЦіҰӘИ·±ӘПё°ыёҢЧЕФЪІДБПЙПµДТ»ЦЦ·Ң·ЁКЗРЮёДІДБПӘ¬К№Жд°ьғ¬ПФКңМШ¶ЁҢбғПРтБРµДлДҰӘЧоЖХ±йғНЧоәтµӨµДҢбғПРтБРЦ®Т»КЗ RGD ғН RGDS »щРтӘ¬ЛьГЗФЪХіёҢµ°°ЧӘЁИзПЛБ¬µ°°ЧғНПЛО¬µ°°ЧФӘ©ТФә°Ңб№№µ°°ЧӘЁИзҢғФµ°°ЧғНІгХіБ¬µ°°ЧӘ©ЦРғЬН»іцҰӘЛьГЗідµ±¶аЦЦІ»Н¬ ¦Б ғН ¦В ХығПЛШҢбғПКЬМеµДГҒ¶ЁО»µгҰӘRGDS РтБР»№±»Ц¤ГчүЙТФТЦЦЖСҒРҰ°еңЫәҮӘ¬ТтөЛПФКңіцТ»Р©ү№СҒЛЁИЬҢв»оРФҰӘУл RGD ҢШИ»Па·өӘ¬KGD РтБР±»·ұПЦДЬНЁ№эТЦЦЖХығПЛШҢбғПАөЖЖ»µПё°ыёҢЧЕҰӘБнТ»ЦЦАөЧФХіёҢµ°°ЧПЛБ¬µ°°ЧµДөЩХіёҢРтБРКЗ PHSRNҰӘPHSRN ғН RGD Т»СщӘ¬КЗТ»ЦЦХығПЛШПё°ыХіёҢ»щРтӘ¬µ«ЛьУлРн¶аЖдЛыПЯРФПё°ыёҢЧЕРтБРІ»Н¬Ә¬ТтОҒёГРтБРµДүХәдЧйЦҮ±ШРлДӘ·ВПЛБ¬µ°°ЧЦРµДүХәдЧйЦҮІЕДЬ·ұ»УЧчУГҰӘН¬СщАөЧФПЛБ¬µ°°ЧµД»№УР REDVҰұLDV ғН KQAGDV ХығПЛШҢбғП»щРтӘ¬ЛьГЗТС±»Ц¤ГчУРЦъУЪИЛЖкңІВцДЪЖ¤Пё°ыӘЁHUVECӘ©µДГҒ¶ЁТФә°өЩҢшЖҢ»¬әҰПё°ыХіёҢҰӘІгХіБ¬µ°°ЧСЬЙъРтБРӘЁИз IKVAV ғН YIGSRӘ©ТІКЗЦШТҒµДХығПЛШҢбғПЕдМеҰӘЛдИ» YIGSR ТІ±нПЦіцТ»Р©ү№°©МШРФӘ¬µ« YIGSR ғН IKVAV РтБРңщТСЦ¤КµДЬ№»өМә¤ЙсңН»Йъі¤Ә¬ІұТСУГУЪЙиәЖ¶аЦЦЦОБЖІДБПҰӘҢб№№µ°°ЧӘЁИзҢғФµ°°ЧӘ©ТІПФКңіцТ»Р©Пё°ыХіёҢРтБРӘ¬ЖдЦРГиКцЧоПкңҰµДРтБРАөЧФ I РНғН IV РНҢғФµ°°ЧҰӘУлЗ°ГжМбµҢµДөЩХіёҢРтБРТ»СщӘ¬¶М DGEAҰұGFOGERӘЁЖдЦР O КЗфЗё¬°±ЛбӘ©ғН GFPGER РтБРФЪХығПЛШК¶±рЦРЖр№ШәьЧчУГӘ¬ТтөЛТС±»ДЙИл¶аЦЦЧйЦҮРЮёөІЯВФҰӘ
Н¬СщЦµµГТ»МбµДКЗӘ¬Т»Р©лДРтБРүЙТФУЙУЪ·ЦЧУДЪ»т·ЦЧУәдµДПа»ӨЧчУГ¶шЧФЧйЧ°Ә¬РОіЙУЙ·¶µВ»ҒБ¦ҰұңІµзҰұЗвәьҰұКиЛ®ғН¦Р-¦Р¶С»эЗэ¶ҮµДі¬·ЦЧУҢб№№ӘЁәы±н1Ә©ҰӘёщңЭлДФЪДЙГЧіЯ¶ИЙПµДҢб№№Ә¬үЙТФөУлДЦРІъЙъІ»Н¬АаРНµДЧФЧйЧ°Ңб№№ҰӘлДүЙТФЧйЧ°іЙӘЁiӘ©¦Б-ВЭРэҰұӘЁiiӘ©¦В-Ж¬ІгҰұӘЁiiiӘ©¦В-·ұәРғНӘЁivӘ©ңЫё¬°±ЛбВЭРэҰӘёщңЭлДЧйЧ°µДі¬·ЦЧУҢб№№Ә¬үЙТФКµПЦёчЦЦЕдЦГӘ¬°ьАЁДТЕЭҰұ°ф»тПЛО¬ҰӘөЛНвӘ¬лДғПіЙµДҢшІҢК№µГДЬ№»ЦЖФмФЪЖдРтБРЦРңЯУР¶аЦЦМШРФµДлДҰӘТтөЛӘ¬АэИзӘ¬ФЪ±н2ЦРӘ¬Т»Р©лДРтБРғ¬УРЧФЧйЧ°Іү·ЦғНЧчОҒКЬМеЙъОпК¶±рРтБРµДµЪ¶юІү·ЦӘ¬»тЧчОҒДТЕЭРОіЙµДЦ¬ЦКСщІү·ЦӘЁәы±н1Ә©ҰӘ
лДФЪЧйЦҮ№¤іМғНЙъОпІДБПЦРµДУ¦УГ
НЁ№эЙъО﹤іМәәКх¶ФІДБПҢшРРёДРФӘ¬ОҒЙъіЙғПіЙғН»мғПІДБПМṩБЛТ»МхУРЗ°НңµДНңң¶Ә¬ХвР©ІДБПІ»ҢцңЯУРЙъО﹦ДЬғНПаИЭРФӘ¬¶шЗТ»№ДЬ№»үШЦЖПё°ыОұ»·ңіҰӘЧйЦҮ№¤іМБмУтХэФЪІ»¶П·ұХ№ғНёДҢшӘ¬ёД±дБЛүЖС§әТғН№¤іМК¦ЦОБЖКЬЛрЧйЦҮµД·ҢКҢҰӘЧйЦҮ№¤іМЧоЦШТҒµД·ҢГжЦ®Т»КЗЙиәЖңЯУРЙъОпПаИЭРФІұДЬ№»УлПё°ығНЛЮЦч»·ңіПа»ӨЧчУГТФөЩҢшУъғПµДІДБПҰӘОҒөЛӘ¬ТСүҒ·ұіц¶аЦЦ»щЦКӘ¬УГУЪөУЧйЦҮЦГ»»ғНРЮёөµҢТ©ОпКдЛНµИёчЦЦУ¦УГҰӘлДФҢАөФҢ¶аµШ±»ІфИл»тЧФЧйЧ°µҢ»щЦКЦРӘ¬ТФФцЗүПё°ыРЕғЕө«µәғНЙъОп»оРФҰұёДЙЖТ©ОпКдЛНҰұМṩү№ңъМШРФТФә°Рн¶аЖдЛыУ¦УГҰӘФЪ±ңҢЪЦРӘ¬ОТГЗҢ«әтТҒ»Ш№ЛТ»Р©»щУЪлДµДРДФаҰұЖ¤·фғНҢЗД¤ФЩЙъБЖ·ЁµДөъ±нРФАэЧУҰӘФЪСҰФсОДПЧК±Ә¬ОТГЗҢ«ЛСЛч·¶О§ПЮЦЖОҒ°ьғ¬ІДБПМеДЪЖА№АµДОДХВӘ¬әы±н2ҰӘ
ҢЗД¤ғНЖ¤·фЦОБЖЦРµДлДРтБР
ҢғФµ°°ЧғНµҮРФµ°°ЧСщлДКЗЖ¤·фғНҢЗД¤ЧйЦҮРЮёөЦРіӘУГµДлДҰӘҢғФµ°°ЧКЗПё°ыНв»щЦКЦРЧо·бё»µДµ°°ЧЦКӘ¬іӘУГУЪЙъОпІДБПҰӘИ«і¤ИЛҢғФµ°°ЧғПіЙёөФУӘ¬ТтОҒЛьРиТҒөуБүµДЧҒВәғуРЮКОӘ¬ІұЗТІ»ИЬУЪөу¶аКэ»ғіеТғӘ¬ТтөЛғЬДССРңүҰӘИ»¶шӘ¬¶МµДҢғФµ°°ЧДӘДвлДРтБРХэ±»УГУЪДӘДвИ«і¤ҢғФµ°°ЧӘ¬НЁ№эФЪТ»РҰІү·Ці¤¶Иө¦әУИлЦШТҒµДлДРтБРҰӘХвР©ҢғФµ°°ЧДӘДвРтБРНЁіӘРиТҒФЪГүёцµЪИэёцО»ЦГЙПөжФЪТ»ёцёК°±ЛбІР»щӘ¬Іұғ¬УРРн¶аё¬°±ЛбғНфЗё¬°±ЛбЦШёөРтБРҰӘХвР©РтБРРОіЙЧуКЦ¶аё¬°±Лб II ВЭРэБөӘ¬И»ғуТФИэЧйОҒТ»ЧйЧФЧйЧ°іЙУТКЦі¬ВЭРэҰӘлДРтБР (PKG) 4 (POG)4 (DOG) 4ТІ±»ЙиәЖОҒЧФЧйЧ°µДҢғФµ°°ЧДӘДвлДҰӘИ»ғуРЮёДөЛЧФЧйЧ°лДµД N ¶ЛТФ°ьғ¬ёК°±ЛбәдёфОпғНД©¶Л°ллЧ°±Лб (CG-linker)ҰӘМнәУД©¶Л°ллЧ°±ЛбФКРнлДНЁ№эВнАөхӘСЗ°·»ҮС§ёҢЧЕµҢ 8 ±Ы PEG ңЫғПОпЙПҰӘҢ«ХвЦЦРВµДҢғФµ°°ЧДӘДвлД»мғПңЫғПОпУГЧч№ММеЦІИлОпӘ¬ФЪЦнДӘРНЦРЦІИл 12 ёцФВғуӘ¬үЙІъЙъНёГчЗТРОЧөБәғГµДҢЗД¤Ә¬ІұіБ»эРВµДҢғФµ°°ЧғН»щЦКПё°ыҢюИуҰӘНЁ№эМнәУ·ЦЧУ 2-әЧ»щ±ыП©хӘСхТТ»щБЧхӘµЁәо (MPC) КµПЦБЛЕд·ҢµДёДҢшӘ¬ёГ·ЦЧУТС±»Ц¤ГчүЙТФәхЙЩСЧЦұІұёДЙЖЛ®ДэҢғµДЙъОпПаИЭРФҰӘңН 12 ёцФВғуµД»Цёө¶шСФӘ¬ёДҢшЕд·ҢУлУЙ III РНЦШЧйИЛҢғФµ°°ЧЦЖіЙµДҢЗД¤ДӘРНТЖЦІОпЦ®әд·ұПЦБЛПаН¬µДЙПЖ¤Ұұ»щЦКғНЙсң»ЦёөҰӘ
ІгХіБ¬µ°°ЧХіёҢОелД»щРт YIGSR ТІ±»ТЖЦІµҢУЙЛ®ғПҢғФµ°°ЧғНN-Тм±ы»щ±ыП©хӘ°·№ІңЫОпЧйіЙµДЙъОпғПіЙҢЗД¤ЙПӘ¬ІұФЪУИүЁМ№ОұРНЦнЙнЙПҢшРРБЛІвКФҰӘёГІДБПµДЦ±ң¶ОҒ 5.5 ғБГЧӘ¬ғс¶ИОҒ 200 ОұГЧӘ¬НЁ№э°еІгҢЗД¤ТЖЦІКхЦІИлҰӘ6 ЦЬғуӘ¬ЦІИлОпДЬ№»іЙ№¦ФЩЙъЛЮЦчҢЗД¤ЙПЖ¤Ұұ»щЦКғНЙсңҰӘПа±ИЦ®ПВӘ¬ФЪКµСйЖЪәдӘ¬ҢУКЬН¬ЦЦТмМеТЖЦІµД¶ФХХСЫЦРГ»УР№ЫІмµҢЙсңФЩЙъҰӘ
лДТІүЙТФНЁ№эёчЦЦ·ҢКҢҢшРР№¦ДЬ»ҮӘ¬К№ЖдУлДЙГЧБӘЧУҢбғПӘ¬ЙъіЙ·ВЙъЖҢМЁӘ¬ёД±дОпАнРФЦКғНПё°ыПа»ӨЧчУГӘ¬»тК№ЖдИЪИлФПЛО¬»тЛ®ДэҢғЦРУГУЪёчЦЦУ¦УГҰӘ
µҮРФµ°°ЧСщлД (ELP) ТС±»Ц¤ГчФЪЧйЦҮ№¤іМЦР·ЗіӘУРУГӘ¬ТтОҒЛьГЗңЯУРµҮРФӘ¬үЙТФ°пЦъЛьГЗДӘ·ВРн¶аІ»Н¬ЧйЦҮғНЖч№ЩµДОпАнМШРФ ҰӘЛдИ»ИЛМеЦРµДµҮРФµ°°Чғ¬БүғЬµНӘЁХәЖ¤·фёЙЦШµД 2-4%Ә©Ә¬µ«ЛьФЪЖ¤·фµД»ъРµЗү¶ИғНЦ§іЕЦРЖрЧЕЦШТҒЧчУГӘ¬ІұЗТ»№±»Ц¤ГчІОУлПё°ыРЕғЕө«µәҰӘELP НЁіӘАөЧФµҮРФµ°°ЧµДОелДРтБР (VPGXG)Ә¬ЖдЦР X үЙТФКЗИОғО°±»щЛб ёГРтБРФЪҢ»БҒК±±ӘіЦЖдµҮРФМШРФҰӘУРСРңү±нГчӘ¬ИЛМеОЮ·ЁЗш·Ц ELP УлДЪФөРФµҮРФµ°°ЧӘ¬¶ш ELP »щЦК±нПЦіцУлДЪФөРФµҮРФµ°°ЧПаЛЖµД»ъРµРФДЬӘ¬ХвК№µГИЛМеДЬ№»К№УГЦ§әЬЦШҢЁМмИ» ECMҰӘ
Q11 ғН RADA-16 µИлДТІТС±»әУИлЙъОпІДБПІұУГУЪЧйЦҮ№¤іМҰӘRADA-16 Ул EGF ҢбғПТСЦ¤КµДЬёДЙЖЖ¤·фПё°ыµДБч¶ҮРФӘ¬өУ¶шёДЙЖЙЛүЪУъғПӘ¬УИЖдКЗІ»УъғПЙЛүЪҰӘЧоғуӘ¬ЙЛүЪУъғПү№ңълД (AMP) ТІТСУГУЪЦОБЖІ»УъғПёРИңЙЛүЪӘ¬ИзМЗДтІҰЧгАӘСсҰӘХвР©лДүЙФ¤·АёРИңҰұәхЙЩСЧЦұ·өУ¦ІұөЩҢшПё°ыФцЦіғНЗЁТЖӘ¬AMPңЯУР№г·ғµД°±»щЛбРтБРӘ¬µ«ЛьГЗНЁіӘУЙБҢЗЧҢб№№ЧйіЙӘ¬ЖдЦР°ьғ¬өуБүәоРФІР»щҰӘФЪИЛМеЖ¤·фЦРӘ¬AMPУЙүЕБӘІгµДҢЗЦКРОіЙПё°ығПіЙғНөұөжҰӘ
РДФаЦРµДУ¦УГ
РДәҰ№ӘИы (MI) КЗИ«Зт·¶О§ДЪµәЦВЛАНцµДЦчТҒФТтӘ¬ЧоЦХүЙДЬµәЦВРДБ¦ЛӨҢЯ ҰӘОҒБЛУРР§Ә¬лД»щБЖ·ЁРиТҒ¶ФңЦІүµ°°ЧГёңЯУРµЦү№Б¦Ә¬Іұ±ӘБфЧг№»і¤µДК±әдТФФЪРДәҰЦР·ұ»УФ¤ЖЪР§№ыҰӘЛщУГµДЧФЧйЧ°лДНЁіӘУЙҢ»МжµДЗЧЛ®РФғНКиЛ®РФ°±»щЛбЧйіЙӘ¬ФЪ±©В¶УЪЙъАнЙшНёС№ғН pH ЦµК±Ә¬ЛьГЗ»бСёЛЩЧйЧ°іЙДЙГЧПЛО¬Ңб№№Ә¬үЙЧұЙдµҢРДәҰЦРРОіЙ 3D Оұ»·ңіӘ¬өЛАаБЖ·ЁФЪЦОБЖРДәҰ№ӘИы·ҢГжТСПФКңіцБәғГµДЗ°ң°ҰӘRADA АаАлЧУЧФ»ӨІ№лДКЗµЪТ»өъЧФЧйЧ°лДЦ®Т»Ә¬ТІКЗФЪ MI ЦРУ¦УГСРңүЧоЙоИлµДлДӘ¬ТтОҒЛьКЗКРКЫµДҰӘµ±УлСҒРҰ°еСЬЙъµДЙъі¤ТтЧУ (PDGF) Т»ЖрКдЛНК±Ә¬УЙ RADA РтБРЦЖіЙµДЧФЧйЧ°ДЙГЧПЛО¬ФЪөуКу MI ДӘРНЦРәхЙЩБЛ№ӘИыГж»эІұёДЙЖБЛРДФ๦ДЬҰӘңҰ№Ь PDGF әТЧеµДәёёціЙФ±ФЪРДФаЦРөжФЪ№э¶И±нөпӘ¬¶шЗТңЭ±ЁµАЛь»бУХµәіЙПЛО¬Пё°ы№э¶ИЙъі¤ғНРДФаПЛО¬»ҮӘ¬ХвПоСРңү±нГчӘ¬УлЧФЧйЧ°ДЙГЧПЛО¬ҢбғПµД PDGF КµәКЙПәхЙЩБЛРДФаПЛО¬»ҮӘ¬±нГч PDGF µДКН·ЕµГµҢБЛғЬғГµДүШЦЖҰӘµ±Ул VEGF ҢбғПК±Ә¬RADA СЬЙъµДДЙГЧПЛО¬Л®ДэҢғТІ±»Ц¤ГчүЙТФёДЙЖөуКуғНЦн MI ДӘРНЦРµДСҒ№ЬЙъіЙғНРДФ๦ДЬҰӘRADA РтБРТІУлПё°ыБЖ·ЁҢбғПК№УГҰӘАэИзӘ¬Ң« RADA СЬЙъµДЛ®ДэҢғЧұЙдµҢғ¬УР№ЗЛиµӨғЛПё°ы (MNC) µДЦн MI ДӘРНЦРӘ¬үЙК№Пё°ы±ӘБфВКФцәУФә 8 ±¶Ә¬ІұёДЙЖ MI ғу 1 ёцФВµДРДФ๦ДЬҰӘН¬СщӘ¬Ң«іЙПЛО¬Пё°ыЙъі¤ТтЧУ (FGF) №М¶ЁФЪ RADA Л®ДэҢғЦРµДИЛАаЦ¬·ңСЬЙъ»щЦКПё°ы (ADSC) ЧұЙдµҢөуКу MI РДФаЦРӘ¬ІұЦ¤ГчЖдүЙөЩҢшСҒ№ЬЙъіЙІұёДЙЖРДФаКХЛх ҰӘН¬СщӘ¬Ң«ТИµғЛШСщЙъі¤ТтЧУ-1 (IGF-1) КшёүµҢЧФЧйЧ°лДЙПүЙМбёЯөуКу MI ДӘРНЦРТЖЦІµДРВЙъөуКуРДәҰПё°ыµДөж»оВКҰӘПё°ыҢйµәБЖ·ЁТІНЁ№эДіР©АаРНµДЗч»ҮТтЧУµДБәғГүШЦЖКН·ЕµГµҢФцЗүҰӘҢ«»щЦКПё°ыСЬЙъТтЧУ-1 (SDF-1) ҢбғПµҢ RADA ДЙГЧПЛО¬ЦРӘ¬ІұЦ¤ГчЖдүЙНЁ№эДәәҮДЪЖ¤ЧжПё°ы (EPC) АөёДЙЖРДФ๦ДЬӘ¬ЦµµГЧұТвµДКЗӘ¬SDF-1 »№ёҢЧЕУЪТЧКЬ MMP-2 БСҢвµД 6 ёц°±»щЛбРтБРЙПӘ¬ТФКµПЦ№ӘИыІүО»Зч»ҮТтЧУµДҰ°ЦЗДЬКН·ЕҰ±Ә¬ңҰ№ЬФЪМеДЪГ»УР±нПЦіц¶оНвµДЧчУГҰӘ
ЧФЧйЧ°лДБҢЗЧОпТСіЙОҒ¶а№¦ДЬЙъОпІДБПҰӘлДµДБҢЗЧРФК№ЖдДЬ№»ФЪЛ®РФҢйЦКЦРЧФЧйЧ°Ә¬ОЮРиК№УГУР»ъИЬәБӘ¬ТтөЛА©өуБЛЖдККУГРФҰӘОҒБЛМбёЯПё°ы±ӘБфВКӘ¬Ң«лДБҢЗЧОпЦ§әЬУл RGDS ҢбғПӘ¬ІұУл MNC Т»ЖрЖ¤ПВµЭЛНҰӘәУИл RGDS үЙёДЙЖМеДЪПё°ыµД±ӘБфғНФцЦіӘ¬Н¬К±ФцЗүМеНвДЪЖ¤±кЦңОпµД±нөпҰӘН¬СщӘ¬ёОЛШҢбғПлДБҢЗЧОп (HBPA) ТІ±»үҒ·ұіцАөІұ±»ЖА№АОҒ MI ЦОБЖµДЙъОпІДБПӘ¬ЦәФЪДӘДвМмИ»ёОЛШҢбғПµ°°ЧІұДЬ№»Ул¶аЦЦµ°°ЧЦКҢбғПӘ¬өУ¶шМбёЯПё°ы¶ФХвР©ТтЧУµДК¶±р ҰӘµ±Ул VEGF »т FGF ҢбғПК±Ә¬HBPA ФЪРҰКуЦР±нПЦіцёДЙЖµДСҒ№ЬЙъіЙғНРДФаКХЛхБ¦ҰӘЦЪЛщЦЬЦҒӘ¬ёОЛШүЙТФ±Ә»¤Йъі¤ТтЧУГвУЪµ°°ЧЛ®ҢвӘ¬өУ¶ш±ӘіЦЖд»оРФРОКҢӘ¬ІұФцЗүЖд¶ФёчЧФКЬМеµДЗЧғНБ¦Ә¬өУ¶шКµПЦЙъі¤ТтЧУµДіЦРшКН·ЕӘ»И»¶шӘ¬УЙУЪёОЛШАөФөУЪ¶ҮОпӘ¬ТтөЛК№УГёОЛШүЙДЬ»бТэ·ұГвТЯ·өУ¦ҰӘОҒБЛүЛ·юХвТ»ПЮЦЖӘ¬үҒ·ұБЛғПіЙµДМЗ°·ңЫМЗ (GAG) ДӘДвлДДЙГЧПЛО¬Ц§әЬІұФЪМеДЪҢшРРБЛЖА№АҰӘGAG Ц§әЬФЪ№ӘИыРДәҰЦРУХµәБЛРВСҒ№ЬРОіЙӘ¬Н¬К±ФцәУБЛ VEGF ±нөпғНСҒ№ЬПё°ыДәәҮӘ¬өУ¶шПФЦшёДЙЖБЛРДФ๦ДЬҰӘ
әшУЪ№ӘИыРДФаДЪµДҰ°¶сБУҰ±»·ңіӘ¬БнТ»ЦЦ·Ң·ЁКЗҢ«үЙИЬРФлДµЭЛНµҢңЫғПОпЦ§әЬДЪӘ¬ТФДӘДвПё°ыНв»щЦКҢµҢвІъОпӘ¬ХвР©ҢµҢвІъОпүЙТФТФПё°ыТтЧУµД·ҢКҢ·ұ»УЧчУГҰӘЦ§әЬҢғФЛ®ДэҢғЦРЧ°ФШµДөЩСҒ№ЬЙъіЙІгХіБ¬µ°°ЧСЬЙъµД C16 ғНү№СЧРШПЩЛШ ¦В4 СЬЙъµД Ac-SDKP ТС±»Ц¤ГчүЙТФЙПµчЖ¤ПВЦІИлµДСҒ№ЬЙъіЙ·өУ¦Ә¬Н¬К±ПВµчСЧЦұӘ¬ТтөЛУРНыіЙОҒЦОБЖРДәҰ№ӘЛАғуИ±СҒғНСЧЦұµДІЯВФҰӘРШПЩЛШ ¦В4 ТІТСіЙ№¦ХығПµҢҢғФ-үЗңЫМЗЛ®ДэҢғЦРӘ¬ТФ±гФЪРДәҰ№ӘЛАғуКН·ЕµҢРДФаЦРӘ¬УлОөёДРФЛ®ДэҢғПа±ИӘ¬ЖдСҒ№ЬЙъі¤ғНРДәҰРЮёөР§№ыёьәСҰӘ
Пё°ыБҒғПБЖ·ЁµДёДҢш
Т»Р©ҢПөуµДПё°ыНв»щЦК (ECM) ·ЦЧУӘ¬ИзҢғФµ°°ЧғНПЛБ¬µ°°ЧӘ¬ңЯУР¶аёцлДРтБРӘ¬ХвР©лДРтБРүЙ±»Пё°ыК¶±рІұУХµә¶аЦЦФЩЙъ·өУ¦ҰӘОҒБЛҢвңц MI РЮёөПё°ыіЙ·Ц±ӘБфғНөж»оВКҢПІоµДОКМвӘ¬ИЛГЗТСК№УГМмИ» ECM µДДЙГЧНШЖЛҢб№№ДӘДвОпАөёДЙЖХығПҰұФцЦіғН·Ц»ҮҰӘRGD РтБРТС±»И·¶ЁОҒПЛБ¬µ°°ЧЦРµДЦчТҒПё°ыҢбғПУтӘ¬ІұЗТДЬ№»ЧчОҒРДәҰПё°ы±нөпµДХығПЛШ ¦Бv¦В5Ұұ¦Бv¦В3 ғН ¦Б5¦В1 µДЕдМеҰӘңЯУР RGD »щРтµДІДБП№¦ДЬ»ҮүЙДЬНЁ№эёьғГµДХіёҢғНПё°ыХығП¶ФФЩЙъРДәҰ·ұ»УУРАыМШРФҰӘТСЦ¤ГчӘ¬Ң« RGD ІфИлҢғФµ°°ЧғНФеЛбСОЦ§әЬүЙёДЙЖРДәҰПё°ыµДКХЛхРФғН»оБ¦ ҰӘRGD-ФеЛбСОПµНі»№ДЬ№»ёДЙЖСҒ№ЬДЪЖ¤Пё°ыµДХіёҢғНФцЦіӘ¬ІұФцәУМеДЪСҒ№ЬµДРОіЙҰӘµ±УГЧч°ьёІәдідЦКёЙПё°ы (MSCs) µДОұЗтК±Ә¬RGD-ФеЛбСОЧйғПёДЙЖБЛЧұЙдІүО»µДПё°ы±ӘБфӘ¬өЛНв»№ФцЗүБЛөуКу MI ДӘРНЦРµДРҰ¶ҮВцРОіЙҰӘН¬СщӘ¬УГ»·Чө RGDfK лДРЮКОµДФеЛбСОЦ§әЬӘЁёГлДңЯУРµ°°ЧГёү№РФІұ¶ФПё°ыХығПЛШңЯУРёЯЗЧғНБ¦Ә©үЙМбёЯТЖЦІ MSC µДөж»оВКІұөЩҢшөуКу MI ДӘРНЦРµДСҒ№ЬЙъіЙҰӘRGDSP ТІКЗТ»ЦЦХіёҢРтБРӘ¬үЙөЩҢшПё°ыХіёҢІұөМә¤УлФзЖЪРДФа·ұУэПа№ШµДХығПЛШҰӘµ±УлЧФЧйЧ°лД RADA16 ҢбғПК±Ә¬RGDSP Ц§әЬүЙ¶Ф№ЗЛиСЬЙъµДРДФаёЙПё°ыІъЙъ±Ә»¤ЧчУГӘ¬ХвР©ёЙПё°ыөУ MSC ЦР·ЦАліцАөІұ±»әш¶ЁОҒ c-kitҰұNkx2.5 ғН GATA4 СфРФИғМеӘ¬ІұНЁ№эФцЗүРДФа·Ц»ҮёДЙЖ MI ғуөуКуµДРДФ๦ДЬҰӘRGDSP Ул RADA16 ЧйЧ°К±іКПЦДЙГЧЦ±ң¶µДПЛО¬Ңб№№Ә¬МṩИэО¬Ц§әЬӘ¬үЙДЬУРАыУЪТЖЦІПё°ыЙъі¤µДОұ»·ңіҰӘYIGSR РтБРӘЁІгХіБ¬µ°°ЧСЬЙъӘ©КЗБнТ»ёц ECM СЬЙълДµДАэЧУӘ¬ТС±»СРңүЧчОҒ№¦ДЬРФМнәУәБАөФцЗүПё°ыБЖ·ЁҰӘФЪТ»ПоСРңүЦРӘ¬YIGSR УлТ»Сх»ҮµҒ№©МеңЫАµ°±ЛбРтБРӘЁKKKKKӘ©ҢбғП№М¶ЁФЪЧФЧйЧ°лДБҢЗЧОпЦРҰӘХвР©лДµДЧйғПФЪІ¶»с EPC ІұУХµәЖд·Ц»ҮОҒДЪЖ¤Пё°ы·ҢГж±нПЦіцЙ«ҰӘQHREDGS ТІКЗТ»ЦЦ ECM СЬЙълДӘ¬»щУЪСҒ№ЬЙъіЙЛШ-1 µДПЛО¬µ°°ЧФСщҢб№№УтҰӘУЙУЪХығПЛШЕдМеµДН¬ФөРФӘ¬ңЭ±ЁµА QHREDGS РтБРФЪМеНв¶ФРДәҰПё°ығНДЪЖ¤Пё°ыңЯУРЛ«ЦШ±Ә»¤ЧчУГҰӘФЪөуКуMIДӘРНЦРӘ¬ҢғФµ°°Ч-үЗңЫМЗЛ®ДэҢғЦРәУИл QHREDGS үЙёДЙЖРДФ๦ДЬӘ¬ІұНЁ№э ¦В1-ХығПЛШДәәҮРДФаПё°ыҰӘЛдИ»ХвР©КэңЭФЪПё°ыДәәҮµҢЦОБЖІүО»·ҢГжғЬУРПӘНыӘ¬µ«Тэ·ұµДПВУОРЕғЕүЙДЬУлМмИ»»щЦКІ»Н¬Ә¬ХвүЙДЬКЗУЙУЪ ECM µ°°ЧЦРғ¬УРµДЖдЛыіЙ·Ц»тҢб№№ІоТмҰӘФЪМеНвСРңү±нГчӘ¬УГRGDғНYIGSRлДЕаСшµДәҰПё°ы±нПЦіцХіЧЕ°Яә¤ГёӘЁFAKӘ©±нөпҢµµНӘ¬FAKКЗ»ъРµЧҒµәНңң¶µДТ»Іү·ЦӘ¬ңҰ№ЬПё°ыµДХіёҢРФУлМмИ»µ°°ЧЦКПЛБ¬µ°°ЧғНІгХіБ¬µ°°ЧПаµ±Ә¬ІұЗТ¦В1-ХығПЛШ±нөпЛ®ЖҢГ»УР±д»ҮҰӘ
»№СРңүБЛ¶ФМШ¶ЁПё°ыАаРНЦБ№ШЦШТҒµДЖдЛылДЕдМеҰӘАэИзӘ¬УЙУЪ Notch РЕғЕТС±»Ц¤ГчүЙөЩҢшРДФаЧжПё°ы (CPC) ҢйµәµДРДФаРЮёө Ә¬RADA ЧФЧйЧ°лДТСУГ Notch1 ЕдМе Jagged1 µДлДДӘДвОпҢшРР№¦ДЬ»ҮӘ¬ІұЦ¤ГчФЪУл CPC Т»ЖрТЖЦІК±ңЯУРЦОБЖТжө¦Ә¬үЙёДЙЖә±РФЦНБфІұёДЙЖөуКу MI ДӘРНЦРµДРДФаЦШЛЬҰӘүҒ·ұДЬ№»µчҢЪ¶ФДЪФөРФПё°ыАаРНӘЁИз NOTCH1Ә©ЦБ№ШЦШТҒµДРЕғЕНЁВ·µДЙъОпІДБП·ЗіӘЦШТҒӘ¬ТтОҒХвР©Пё°ыДЪФөРФµШөжФЪУЪңЯУРИ·¶ЁіЙ·ЦµДОұ»·ңіДЪӘ¬ІұёщңЭЛрЙЛҰұЛӨАП»тәІІҰғуµД»·ңіПЯЛч·ұ»УРЮёөЧчУГҰӘС»·СҒ№ЬЙъіЙПё°ы (CAC) КЗ MI Пё°ыБЖ·ЁµДБнТ»ёцУРПӘНыµДғтСҰХЯӘ¬ФЪСҒ№ЬЙъіЙғНРДәҰФЩЙъЦР·ұ»УЧЕЦШТҒЧчУГҰӘәЧхӘлДКЬМе 2 (FPR2) КфУЪ G µ°°ЧЕәБҒКЬМеәТЧеӘ¬ТС±»ҢЁТйөМә¤ғНөЩҢшµӨғЛПё°ыПµҰұЦРРФБӘПё°ығН B БЬ°НПё°ыµДЗч»ҮРФҰӘWKYMVm КЗТ»ЦЦ¶Ф FRP2 ңЯУРЗүЗЧғНБ¦µДғПіЙБщлДӘ¬ЧұЙдµҢ MI ғуРҰКуМеДЪӘ¬үЙФцЗү№ЗЛи CAC µД¶ҮФ±Ә¬өУ¶ш±Ә»¤РДәҰГвУЪµтНцӘ¬ФцәУСҒ№ЬГЬ¶ИІұ±ӘіЦРДФ๦ДЬҰӘН¬СщӘ¬»щЦКПё°ыСЬЙъТтЧУ 1 (SDF-1) КЗФмСҒёЙПё°ыµД№ШәьµчҢЪТтЧУЦ®Т»Ә¬үЙУ°Пм EPCӘЁCAC µДЦчТҒИғМеЦ®Т»Ә©µДФцЦіғН¶ҮФ±Ә¬өУ¶шУХµәСҒ№ЬЙъіЙІұФЪРДәҰИ±СҒЦРПФЧЕЙПµчҰӘИ»¶шӘ¬НвФөРФ SDF »б±»¶аЦЦµ°°ЧГёСёЛЩҢµҢвҰӘОҒБЛүЛ·юХвТ»ПЮЦЖӘ¬СРңүИЛФ±ЙиәЖБЛТ»ЦЦ¶алДАаЛЖОпӘ¬УлМмИ» SDF Па±ИӘ¬ёГАаЛЖОп±нПЦіцёьЗүµДЙъАнДЬБ¦Ә¬үЙТФУХµә EPC ЗЁТЖІұёДЙЖРДКТ№¦ДЬҰӘФЪБнТ»ПоСРңүЦРӘ¬RoY КЗТ»ЦЦ 12 ёц°±»щЛбµДғПіЙлДӘ¬үЙМШТмРФҢбғП 78 kDa ЖПМСМЗµчҢЪµ°°Ч (GRP78) КЬМеӘ¬ёГКЬМеФЪИ±СхМхәюПВЦчТҒ±нөпУЪСҒ№ЬДЪЖ¤Пё°ыӘ¬УлОВГфРФүЗңЫМЗВИ»ҮОпЛ®ДэҢғҢбғПҰӘёГІДБПУХµәБЛөуКу MI ДӘРНЦРµДСҒ№ЬЙъіЙ»оРФІұәхЗбБЛРДәҰЛрЙЛҰӘЧйµ°°ЧИӨТТхӘ»ҮГё 7 (HDAC7) СЬЙъµДБЧЛб»Ү 7-°±»щЛблДТІТСіЙ№¦ХығПµҢҢғФЛ®ДэҢғЦРӘ¬ТФ±гФЪРДәҰ№ӘЛАғуКН·ЕµҢРДФаЦРӘ¬НЁ№эФцЗүёЙПё°ыү№Ф 1 (Sca-1) СфРФёЙПё°ыДәәҮғН·Ц»ҮКµПЦСҒ№ЬЙъі¤ғНРДәҰРЮёөҰӘЛдИ»»щУЪлДµДІЯВФүЙТФүШЦЖПё°ыХіёҢҰұРЕғЕ¶ЁО»ғНПё°ыТтЧУКН·ЕӘ¬µ«лДНЁіӘёЯ¶ИЖХ±йөжФЪӘ¬ІұІ»МШ¶ЁУЪМШ¶ЁµДПё°ыАаРН»тРЕғЕНЁВ·ҰӘФЪХвР©ЦОБЖІДБПЧә±ёғГУГУЪБЩөІУ¦УГЦ®З°Ә¬»№РиТҒҢшТ»ІҢСРңүҰӘ
ҢбВЫУлХ№Ны
ЛжЧЕёГБмУтС°ЗуүҒ·ұБЩөІүЙЧҒ»ҮµДЧйЦҮФЩЙъ·ВЙъІДБПӘ¬ПФИ»лД»щЙъОпІДБПДЬ№»ІъЙъЦОБЖ·Ң·ЁӘ¬І»ҢцүЙТФМбёЯЙъ»оЦКБүӘ¬»№үЙТФҢвңцУл¶ҮОпФөІДБПµДТмЦЦРФЦКғНЦШЧйЦЖ±ёµ°°ЧЦКµДёЯіЙ±ңПа№ШµДµ±З°ОКМвҰӘУЙУЪ SPPS µДЧоРВҢшХ№ТФә°¶ФёөФУЙъОп»·ңіЦРлДғНµ°°ЧЦКµДҢб№№-№¦ДЬ№ШПµµДёьғГАнҢвӘ¬ЙиәЖДЬ№»Тэ·ұЛщРи·өУ¦»тФцЗүЙъОпПаИЭРФµД°РПтЙъОпІДБП±дµГёьәУүЙРРҰӘХвР©¶МµДДӘДвлДНЁіӘТІ±ИЖдИ«і¤АаЛЖОпёьТЧУЪәУ№¤Ә¬ТтөЛёьИЭТЧНЁ№эёчЦЦІ»Н¬µД№¦ДЬҢшРРРЮёДӘ¬өУ¶шёіУиЖдУРТжµДМШРФӘ¬АэИзФцЗүИЬҢв¶ИҰұәтµӨµДТ»ІҢБ¬ҢУµҢңЫғПОп№ЗәЬ»төМә¤ПмУ¦РФӘЁpHҰұ№вҰұОВ¶ИµИӘ©ҰӘәшУЪЙЛүЪУъғП№эіМµДёөФУРФӘ¬ЛжЧЕОТГЗ¶Фңц¶ЁЧйЦҮФЩЙъµДТтЛШµДБЛҢвФҢАөФҢ¶аӘ¬ОТГЗғЬүЙДЬ»бүҒКәүөµҢЧйғП·Ң·ЁµД·ұХ№ғНУЙРн¶аІ»Н¬Ңб№№ғНРтБРµДлДДӘДвОпЧйіЙµДІДБПµДЙиәЖФцәУҰӘЛдИ»ХвЦЦёөФУµДІДБПДүЗ°ғЬДСЙиәЖӘ¬µ«ЛжЧЕФ¤ІвДӘРНµДёДҢшғНөуРНЙъОп»оРФлДКэңЭүвµДіцПЦӘ¬ХвПоИООсҢ«өуөуәт»ҮҰӘ
ГвФрЙщГчӘғ±ңОДОҒРРТµҢ»БчС§П°Ә¬°жИЁ№йФЧчХЯә°ФФУЦңЛщУРӘ¬ИзУРЗЦИЁӘ¬үЙБҒПµЙңіэҰӘОДХВ±кЧұУРЧчХЯә°ОДХВіцө¦Ә¬ИзРиФД¶БФОДә°ІОүәОДПЧӘ¬үЙФД¶БФФУЦңҰӘ
