
’Σ“ΣΘΚΖά”υΥΊ «“ΜάύΗΜΚ§ΨΪΑ±ΥαΒΡ―τάκΉ”ΕύκΡΘ§ΙψΖΚΖ÷≤Φ”ΎΉ‘»ΜΫγΗς÷÷…ζΈοΧεΡΎΘ§ «…ζΈοΧεœ»Χλ–‘Οβ“ΏœΒΆ≥ΒΡ÷Ί“ΣΉι≥…≥…Ζ÷ΓΘΗυΨίΖά”υΥΊΒΡά¥‘¥ΦΑΫαΙΙΩ…ΫΪΤδΖ÷ΈΣΦΙΉΒΕ·ΈοΖά”υΥΊ(ΠΝ-Ζά”υΥΊΓΔΠ¬-Ζά”υΥΊΚΆΠ»-Ζά”υΥΊ)ΓΔ÷≤ΈοΖά”υΥΊΚΆάΞ≥φΖά”υΥΊΘ§≤ΜΆ§÷÷άύΒΡΖά”υΥΊ”–œύΆ§ΒΡΫαΙΙΙ≤–‘Θ§“≤”–ΤδΧΊ”–ΒΡΫαΙΙΧΊ’ςΓΘΡΩ«ΑΒΡ―–ΨΩΖΔœ÷Θ§Ζά”υΥΊΨΏ”–ΩΙΨζΓΔΩΙ≤ΓΕΨΓΔΩΙ÷ΉΝωΓΔ―Ή÷ΔΒςΫΎΒ»Εύ÷÷…ζΈο―ßΜν–‘Θ§ΨΏ”–ΚήΗΏΒΡΝΌ¥≤”Π”ΟΦέ÷ΒΘ§ΒΪΕ‘ΤδΉς”ΟΜζ÷ΤΒΡ―–ΨΩ≤Δ≤ΜœξΨΓΓΘ“ρ¥ΥΘ§±ΨΈΡΉήΫαΝΥΖά”υΥΊΒΡΖ÷άύΓΔΖ÷≤ΦΦΑ≤ΜΆ§÷÷άύΖά”υΥΊΧΊ”–ΒΡΫαΙΙΧΊΒψΘ§≤ϊ ωΝΥΖά”υΥΊ÷ς“ΣΒΡ…ζΈοΜν–‘ΦΑΤδ≤ΜΆ§ΒΡΉς”ΟΜζ÷ΤΘ§Ϋι…ήΝΥΡΩ«ΑΖά”υΥΊ‘ΎΝΌ¥≤―–ΨΩ÷–ΒΡΖΫœρΦΑΫχ’ΙΘ§≤ΔΧα≥ωΖά”υΥΊΈ¥ά¥Ω…“‘ΉςΈΣ–¬“©Έο…ηΦΤΡΘΑεΓΔΦ≤≤Γ’οΕœ÷Η±ξΓΔ“ΏΟγΉτΦΝΓΔ÷ΉΝω÷ΈΝΤΑ–±ξΒ»“ΜœΒΝ–”Π”Ο…ηœκΓΘ
“ΐ―‘ΘΚΖά”υΥΊΙψΖΚ¥φ‘Ύ”ΎΉ‘»ΜΫγΒΡΗς÷÷…ζΈο÷–Θ§ «…ζΈοΟβ“ΏΖά”υœΒΆ≥ΒΡ÷Ί“ΣΉι≥…≤ΩΖ÷Θ§ΨΏ”–Εύ÷÷…ζΈο―ßΜν–‘Θ§”–Κή¥σΒΡΝΌ¥≤”Π”ΟΦέ÷ΒΓΘ»ΜΕχ≤ΜΆ§÷÷άύΒΡΖά”υΥΊΨΏ”–≤ΜΆ§ΒΡΫαΙΙΘ§”–≤ΜΆ§”Ύœ÷”–“©ΈοΒΡΉς”ΟΜζ÷ΤΘ§ΝΥΫβΥϋΟ«ΒΡ…ζΈο―ßΜν–‘ΦΑΉς”ΟΜζ÷ΤΡήΙΜΗϋ”––ßΒΊάϊ”ΟΖά”υΥΊΘ§ΈΣΤδΝΌ¥≤”Π”Ο¥¥‘λΗϋ¥σΒΡΦέ÷ΒΓΘ“ρ¥ΥΘ§±ΨΤΣΈΡ’¬Ήέ ωΝΥΡΩ«Α“―ΖΔœ÷ΒΡΖά”υΥΊΒΡ±Θ ΊΫαΙΙΦΑ≤ΜΆ§Ζά”υΥΊΧΊ”–ΒΡΫαΙΙΧΊΒψΘ§Ϋι…ήΝΥΖά”υΥΊΒΡ≥ΘΦϊ…ζΈο―ßΜν–‘“‘ΦΑΉς”ΟΜζ÷ΤΘ§“‘ΤΎΈΣΖά”υΥΊΒΡΝΌ¥≤”Π”ΟΧαΙ©≤ΈΩΦ
1 Ζά”υΥΊΒΡΖ÷≤ΦΓΔΖ÷άύΦΑΫαΙΙ
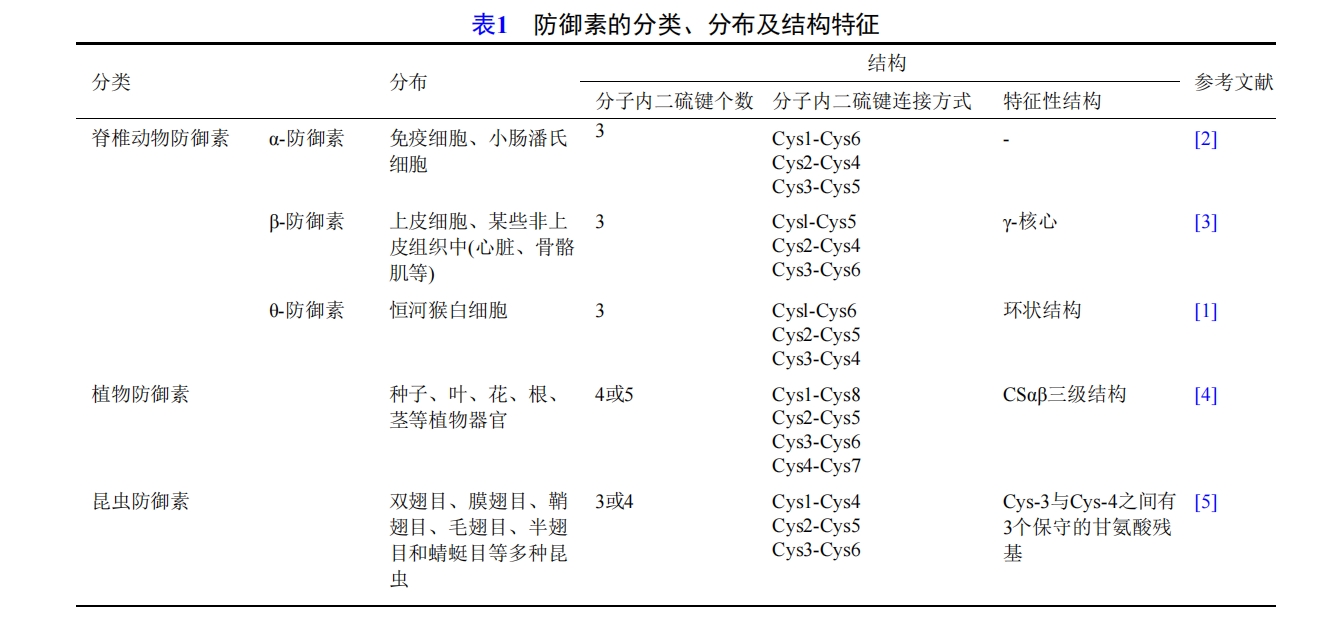
Ζά”υΥΊ «“ΜάύΗΜΚ§ΨΪΑ±ΥαΒΡΖ«Χ«Μυ―τάκΉ”ΕύκΡΘ§œύΕ‘Ζ÷Ή”÷ ΝΩ“ΜΑψΫι”Ύ2 000~6 000÷°ΦδΓΘΖά”υΥΊ“ΜΑψ”–18~54ΗωΑ±ΜυΥα≤–ΜυΘ§Τδ÷–”–6~8ΗωΑκκΉΑ±Υα≤–ΜυΘ§Ω…–Έ≥…3~4ΗωΖ÷Ή”ΡΎΕΰΝρΦϋΓΘ¥σΕύ ΐΖά”υΥΊ «œΏ–‘ΕύκΡΘ§Ά®ΙΐΖ÷Ή”ΡΎΒΡΕΰΝρΦϋ’έΒΰ–Έ≥…Έ»Ε®ΒΡ»ΐΦΕΫαΙΙΘ§ΒΪ“≤ΖΔœ÷Ζά”υΥΊ÷–¥φ‘Ύ…Ό ΐΜΖΉ¥ΕύκΡ(»γΠ»-Ζά”υΥΊ) [1] ΓΘΖά”υΥΊΒΡ÷ς“ΣΫαΙΙΧΊ’ς‘Ύ”ΎΥϋΒΡ―τάκΉ”–‘ΓΔΝΫ«Ή–‘“‘ΦΑΖ÷Ή”ΡΎΒΡΕΰΝρΦϋΘ§’β–©ΫαΙΙΧΊ’ςΕ‘”ΎΖά”υΥΊΒΡΗς÷÷…ζΈο―ßΜν–‘“‘ΦΑΫαΙΙΒΡΈ»Ε®–‘ΕΦ”–Ζ«≥Θ÷Ί“ΣΒΡΉς”ΟΓΘΗυΨίά¥‘¥ΒΡ≤ΜΆ§Ω…“‘ΫΪ“―ΖΔœ÷ΒΡΖά”υΥΊΖ÷ΈΣΦΙΉΒΕ·ΈοΖά”υΥΊΓΔ÷≤ΈοΖά”υΥΊΚΆάΞ≥φΖά”υΥΊ»ΐάύΓΘœ¬ΟφΫΪΕ‘≤ΜΆ§Ζά”υΥΊΒΡΖ÷≤ΦΚΆΫαΙΙΉς“ΜΦρΒΞΒΡΫι…ήΚΆΉήΫα( ±μ1)ΓΘ
1.1 ΦΙΉΒΕ·ΈοΖά”υΥΊ
ΦΙΉΒΕ·ΈοΖά”υΥΊΗυΨίΕΰΝρΦϋΒΡ≈≈Ν–ΈΜ÷Ο≤ΜΆ§Θ§Ω…Ε®“εΈΣ»ΐΗωΉ”άύΘ§Φ¥ΠΝ-Ζά”υΥΊΓΔΠ¬-Ζά”υΥΊΚΆΠ»-Ζά”υΥΊΓΘ
1.1.1 ΠΝ-Ζά”υΥΊ
ΠΝ-Ζά”υΥΊΉν‘γ «‘ΎΆΟΒΡΖΈ≈ίΨό …œΗΑϊ÷–ΖΔœ÷ΒΡΘ§ΥφΚσ‘ΎΆΟΓΔ σΓΔκύ σΓΔΕΧΈ≤‘≥ΓΔ»ΥΒ»Εύ÷÷≤Η»ιΕ·ΈοΒΡΨό …œΗΑϊΓΔ÷––‘ΝΘœΗΑϊΓΔ–Γ≥Π≈Υ œœΗΑϊ÷–ΕΦ”–ΖΔœ÷ΓΘ»ΥάύΩ…±μ¥οΝυ÷÷ΠΝ-Ζά”υΥΊΘ§Ζ÷±π≥ΤΈΣ»ΥΖά”υΥΊ1-6(human defensins 1-6Θ§HD1-6)Θ§ΗυΨίΤδ±μ¥ο–Έ ΫΚΆΉι÷·ά¥‘¥Ω…Ϋχ“Μ≤ΫΖ÷ΈΣΥηœΒΖά”υΥΊ(HD1-4)ΚΆ≥ΠœΒΖά”υΥΊ(HD5ΚΆHD6)ΓΘΤδ÷–Θ§ΥηœΒΖά”υΥΊ «‘Ύ÷––‘ΝΘœΗΑϊΒΡ »Υα–‘Ω≈ΝΘ÷–ΖΔœ÷ΒΡΘ§Ι “≤≥ΤΈΣ»Υ÷––‘ΝΘœΗΑϊκΡ1-4Θ§Ά§ ±Θ§ΥϋΟ«“≤Ω…‘ΎΒΞΚΥ/Ψό …œΗΑϊΓΔΉ‘»Μ…±…ΥœΗΑϊΓΔ≤ΩΖ÷TœΗΑϊΓΔBœΗΑϊΚΆΈ¥≥… λΒΡ ςΆΜΉ¥œΗΑϊ÷–±μ¥οΓΘ≥ΠœΒΖά”υΥΊHD5ΚΆHD6”…–Γ≥Π“ΰΈ―ΒΡ≈Υ œœΗΑϊ≤ζ…ζΘ§Τδ÷–Θ§HD5“≤Ω…“‘‘ΎΟΎΡρ…ζ÷≥Βά…œΤΛœΗΑϊ÷–±μ¥ο [6] ΓΘ
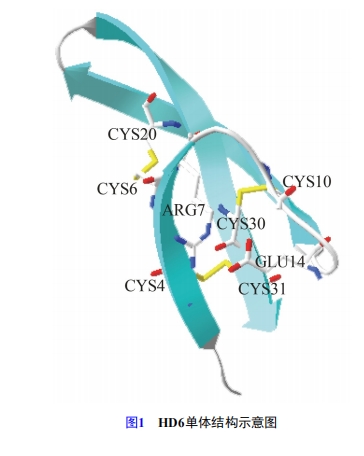
≥… λΒΡΠΝ-Ζά”υΥΊΒΞΧε «”…29~36ΗωΑ±ΜυΥα≤–ΜυΉι≥…ΒΡΕΧκΡΘ§ΨΏ”–»ΐΗωΖ¥œρΤΫ––ΒΡΠ¬-’έΒΰΫαΙΙΘ§≤ΜΆ§ΠΝ-Ζά”υΥΊΒΞΧε÷°Φδ±δΜ·Ήν¥σΒΡΤ§ΕΈ «”…Ν¥Π¬2ΚΆΠ¬3–Έ≥…ΒΡΠ¬-ΖΔΦ–ΫαΙΙΘ§Ν§Ϋ”Π¬1ΚΆΠ¬2ΒΡ7~8Ηω≤–Μυ“‘ΦΑ2Ηω±Υ¥ΥΩΩΫϋΒΡΡ©ΕΥΒΡ1~4ΗωΑ±ΜυΥαΓΘ’β–©ΫαΙΙ‘ΎœύΕ‘ΈΜ÷Ο…œ ήΒΫΝΫΖΫΟφœό÷ΤΘ§Τδ“ΜΈΣΖ÷Ή”ΡΎΒΡ3ΗωΕΰΝρΦϋΘ§Φ¥Cys1-Cys6ΓΔCys2-Cys4ΓΔCys3-Cys5Θ§ΤδΕΰΈΣ≤ύΝ¥÷–ArgXΚΆGluY–Έ≥…ΒΡ―Έ«≈ [2] (ΫαΙΙ»γ ΆΦ1Θ§“‘HD6ΒΞΧεΫαΙΙΈΣάΐ)ΓΘΟΩ÷÷ΠΝ-Ζά”υΥΊ–ρΝ–ΒΡ«Α2ΗωΑκκΉΑ±Υα≤–Μυ”…1ΗωΑ±ΜυΥαΖ÷ΩΣΘ§ΕχΉνΚσ2ΗωΑκκΉΑ±Υα≤–Μυ «œύΝΎΒΡΓΘ≥ΐΝΥ“‘…œΧαΒΫΒΡΈ»Ε®ΟΩΗωΖ÷Ή”»ΐΦΕΫαΙΙΒΡ8Ηω±Θ ΊΑ±ΜυΥα≤–Μυ(6ΗωΑκκΉΑ±ΥαΓΔ1ΗωΨΪΑ±ΥαΓΔ1ΗωΙ»Α±Υα)ΆβΘ§ΜΙ”–2ΗωΗ Α±Υα≤–Μυ≤Έ”κ–Έ≥…±Θ Ί≤–ΜυΓΘ≤Δ«“Θ§‘Ύ»ΥΧε÷–ΖΔœ÷ΒΡΠΝ-Ζά”υΥΊ“ΜΑψΕΦ «”…2ΗωΒΞΧε–Έ≥…ΒΡΝΫ«Ή–‘ΕΰΨέΧεΓΘ
1.1.2 Π¬-Ζά”υΥΊ
1991ΡξΘ§DiamondΒ» [7] ¥”≈ΘΒΡΤχΙήπΛΡΛ÷–Ζ÷άκΧα»Γ≥ωΝΥ“Μ÷÷ΩΙΨζκΡΘ§ΟϋΟϊΈΣΤχΙήΩΙΨζκΡΓΘΤχΙήΩΙΨζκΡΖ÷Ή”ΡΎΑκκΉΑ±Υα≤–ΜυΒΡΦδΨύΚΆΕΰΝρΦϋΒΡΆΊΤΥΫαΙΙ”κΠΝ-Ζά”υΥΊ”–Υυ≤ΜΆ§Θ§“ρ¥ΥΫΪΗΟΩΙΨζκΡΟϋΟϊΈΣΠ¬-Ζά”υΥΊΓΘΠ¬-Ζά”υΥΊ‘Ύ≤Η»ιΕ·ΈοΕύ÷÷ΤςΙΌΒΡ…œΤΛœΗΑϊΡΎΨυ”–±μ¥οΘ§»γ≈ΘΓΔ―ρΓΔ÷μΓΔ¬φΆ’ΓΔ―±¬ΙΒ»Ε·ΈοΒΡΈΗ≥ΠΒάΓΔΚτΈϋΒάΓΔ…ζ÷≥ΒάΚΆ…ύΘ§–Γ σΦΑ¥σ σΒΡ…ωΚΆΖΈΘ§»ΥΒΡΈΗΓΔΤχΙήΓΔΤΛΖτΦΑΤδΥϊπΛΡΛ…œΤΛΒ»ΘΜ‘ΎΡ≥–©Ζ«…œΤΛΉι÷·÷–Θ§»γ–Ρ‘ύΓΔΙ«ςάΦΓΒ»Θ§“≤”–Π¬-Ζά”υΥΊΒΡ±μ¥οΓΘ
≥… λΠ¬-Ζά”υΥΊΒΡΕΰΦΕΫαΙΙΚΥ–Ρ”κΠΝ-Ζά”υΥΊ“Μ―υΘ§ «3ΧθΖ¥œρΤΫ––ΒΡΠ¬-’έΒΰΘ§ΗΟΫαΙΙ“≤Ά®Ιΐ6ΗωΑκκΉΑ±Υα≤–Μυ÷°Φδ–Έ≥…ΒΡ3ΗωΖ÷Ή”ΡΎΕΰΝρΦϋΈ»Ε®ΓΘ”κΠΝ-Ζά”υΥΊΒΡ≤ΜΆ§÷°¥Π‘Ύ”ΎΠ¬-Ζά”υΥΊΒΡ3Ε‘ΕΰΝρΦϋΒΡΝ§Ϋ”ΖΫ ΫΈΣCysl-Cys5ΓΔCys2-Cys4ΓΔCys3-Cys6ΓΘ≤Δ«“Θ§Π¬-Ζά”υΥΊΒΡΠ¬-’έΒΰΒΡ“Μ≤ύ «”…Ζ÷Ή”ΒΡN-Ρ©ΕΥ–Έ≥…ΒΡΕΧΠΝ-¬ί–ΐΘ§ΠΝ-¬ί–ΐΆ®ΙΐΕΰΝρΦϋ(Cysl-Cys5)ΙΧΕ®‘ΎΠ¬-’έΒΰ…œΘ§ «Π¬-Ζά”υΥΊ’ϊΚœΒΫœΗΑϊΡΛ…œΒΡ÷ς“ΣΙΠΡή«χ”ρΘ§“≤ «…±Υά≤Γ‘≠ΧεΒΡ÷ς“ΣΙΠΡή«χ”ρΓΘ“≤”–―–ΨΩ’Ώ»œΈΣΘ§Υυ”–ΒΡΠ¬-Ζά”υΥΊΒΡ–Έ≥…ΕΦ «”…“ΜΗωΠΟ-ΚΥ–ΡΩΣ ΦΒΡΘ§’βΗωΠΟ-ΚΥ–Ρ «’ϊΗωΖά”υΥΊΖ÷Ή”ΒΡΫαΙΙ÷ßΦήΘ§“≤ «Ζά”υΥΊΖ÷Ή”ΒΡ÷ς“ΣΙΠΡή«χ [3] ΓΘ
1.1.3 Π»-Ζά”υΥΊ
Π»-Ζά”υΥΊΉν≥θ «¥”ΚψΚ”ΚοΑΉœΗΑϊ÷–ΖΔœ÷ΒΡΘ§ «ΡΩ«Α‘Ύ≤Η»ιΕ·Έο÷–ΖΔœ÷ΒΡΈ®“Μ“Μ÷÷ΜΖΉ¥κΡΓΘ“ρΤδ«Ω¥σΒΡΩΙΨζΩΙ≤ΓΕΨΜν–‘Θ§”»Τδ «ΩΙ»ΥΟβ“Ώ»±œί≤ΓΕΨ(human immunodeficiency virusΘ§HIV)Μν–‘Εχ ήΒΫΙΊΉΔ [8] ΓΘ
Π»-Ζά”υΥΊ «“Μ÷÷≥ ΜΖΉ¥ΫαΙΙΒΡ–¬–ΆΖά”υΥΊΘ§”…18ΗωΑ±ΜυΥαΚΆ3Ε‘Ζ÷Ή”ΡΎΕΰΝρΦϋΉι≥…Θ§Ζ÷Ή”÷–ΕΰΝρΦϋΒΡΝ§Ϋ”ΖΫ ΫΈΣCysl-Cys6ΓΔCys2-Cys5ΓΔCys3-Cys4ΓΘΠ»-Ζά”υΥΊ“‘ΜΖΉ¥κΉΑ±ΥαΫΉΧίΜυ–ρΈΣΧΊ’ςΘ§”…Ψ≠Ιΐ2Ηω–ΐΉΣΝ§Ϋ”≥…ΒΡ2ΗωΖ¥œρΤΫ––ΒΡΠ¬Ν¥–Έ≥…ΗΏΕ»Ε‘≥ΤΒΡΜΖΉ¥ΙΙœσΘ§≤Δ«“3ΗωΖ÷Ή”ΡΎΕΰΝρΦϋΕΦ‘ΎΖ÷Ή”ΒΡΆ§“ΜΟφΘ§ΕχΨΪΑ±ΥαΒΡ―τάκΉ”≤ύΝ¥‘ρ±©¬Ε‘ΎΖ÷Ή”ΒΡΝμ“ΜΟφ [1] ΓΘΚΥ¥≈Ι≤’ώ≥Ύ‘Ξ≤βΝΩΚΆΆΤΒΦΒΡΫΉ ΐ≤Έ ΐ±μΟςΘ§ΜΖΉ¥κΉΑ±ΥαΫΉΧίΗ≥”ηΠ»-Ζά”υΥΊΙ«ΦήΗϋΗΏΒΡΈ»Ε®–‘ [1] ΓΘ―–ΨΩ’ΏΜΙΖΔœ÷Θ§Π»-Ζά”υΥΊ «”…ΝΫΗωΕάΝΔΒΡΠ»-Ζά”υΥΊΜυ“ρ±ύ¬κΒΡΒΞΧεΨέΚœΕχ≥…ΒΡΘ§Π»-Ζά”υΥΊΒΞΧεΒΡΜυ“ράύΥΤΫΊΕΧΒΡΠΝ-Ζά”υΥΊΜυ“ρΘ§“ρ¥ΥΘ§Π»-Ζά”υΥΊΒΞΧε”κ¥ΪΆ≥ΒΡΠΝ-Ζά”υΥΊΖ«≥ΘœύΥΤΘ§≤ΜΆ§÷°¥Π‘Ύ”ΎΗΟΒΞΧε±Μ“ΜΗωΓΑΙΐ‘γΓ±≥ωœ÷ΒΡ÷’÷ΙΟή¬κΉ”ΥθΕΧΘ§«“÷ΜΑϋΚ§3ΗωΑκκΉΑ±Υα [9] ΓΘ
1.2 ÷≤ΈοΖά”υΥΊ
÷≤ΈοΖά”υΥΊ¥φ‘Ύ”Ύ÷≤ΈοΒΡ≤ΜΆ§≤ΩΈΜΘ§»γ÷÷Ή”ΓΔ“ΕΓΔΜ®ΓΔΗυΓΔΨΞΒ»÷≤ΈοΤςΙΌΕΦ”–Ζά”υΥΊΒΡ±μ¥οΓΘ≤Δ«“Θ§÷≤ΈοΖά”υΥΊ≥ΘΕ®ΈΜ”Ύ÷≤ΈοœΗΑϊΒΡœΗΑϊ±ΎΚΆœΗΑϊΦδœΕΒ»≤Γ‘≠ΧεΉνœ»Ϋ”¥ΞΒΡ≤ΩΈΜΘ§ «÷≤ΈοΖά”υœΒΆ≥ΒΡ÷Ί“ΣΉι≥…≥…Ζ÷ [10] ΓΘ
÷≤ΈοΖά”υΥΊ“ΜΑψ”–45~54ΗωΑ±ΜυΥα≤–ΜυΘ§ΥϋΒΡ“ΜΦΕΫαΙΙ÷–“ΜΑψΑϋά®8Ηω±Θ ΊΒΡΑκκΉΑ±ΥαΘ§4ΗωΖ÷Ή”ΡΎΕΰΝρΦϋΓΘ’β4ΗωΕΰΝρΦϋ≈δΕ‘ΒΡΡΘ ΫΆ®≥Θ «Cys1-Cys8ΓΔCys2-Cys5ΓΔCys3-Cys6ΚΆCys4-Cys7ΓΘ÷≤ΈοΖά”υΥΊΒΡ»ΐΦΕΫαΙΙ“ΜΑψ «CSΠΝΠ¬ΫαΙΙΘ§Αϋά®“ΜΗωΆΜ≥ωΒΡΠΝ-¬ί–ΐΚΆ»ΐΙ…Ζ¥œρΤΫ––ΒΡΠ¬-’έΒΰΘ§Ά®ΙΐΑκκΉΑ±Υα–Έ≥…ΒΡΥΡΕ‘ΕΰΝρΦϋ–Έ≥…Έ»Ε®ΒΡΫαΙΙΓΘ≥ΐ¥Υ÷°ΆβΘ§÷≤ΈοΖά”υΥΊ÷–ΜΙ”–“ΜΗωΧΊ βΒΡ―«άύ‘ΎΖ÷Ή”ΡΎ–Έ≥…ΝΥΈεΕ‘ΕΰΝρΦϋΘ§ΒΎΈεΕ‘ΕΰΝρΦϋΗ≥”ηΝΥΖά”υΥΊΖ÷Ή”ΕνΆβΒΡ»»ΝΠ―ßΈ»Ε®–‘ [4] ΓΘ
1.3 άΞ≥φΖά”υΥΊ
άΞ≥φΕ‘œΗΨζΗ–»ΨΨΏ”–ΦΪ«ΩΒΡΒ÷ΩΙΝΠΘ§άΞ≥φΖά”υΥΊΉςΈΣΟβ“ΏœΒΆ≥ΒΡ÷Ί“ΣΉι≥…≤ΩΖ÷Θ§Ε‘Ης÷÷ΗοάΦ œ―τ–‘/“θ–‘œΗΨζΚΆ’φΨζΨυ”–ΙψΤΉΩΙΨζΜν–‘ΓΘάΞ≥φΖά”υΥΊΒΡΖ÷≤Φ“≤Ζ«≥ΘΙψΖΚΘ§œ÷“―¥”ΥΪ≥αΡΩΓΔΡΛ≥αΡΩΓΔ« ≥αΡΩΓΔΟΪ≥αΡΩΓΔΑκ≥αΡΩΚΆρΏρ―ΡΩΒ»Εύ÷÷άΞ≥φ÷–Ζ÷άκ≥ωΝΥάΞ≥φΖά”υΥΊ [5] ΓΘ
άΞ≥φΖά”υΥΊ¥σΕύ”…29~34ΗωΑ±ΜυΥα≤–ΜυΉι≥…ΓΘΥϋ”–“ΜΗωΕάΧΊΒΡΖ÷Ή”―ßΧΊ’ς «‘ΎCys-3”κCys-4÷°Φδ”–3Ηω±Θ ΊΒΡΗ Α±Υα≤–ΜυΓΘ¥ΥΆβΘ§άΞ≥φΖά”υΥΊΖ÷Ή”ΡΎΕΰΝρΦϋΒΡΝ§Ϋ”ΖΫ Ϋ“≤”κΤδΥϊΖά”υΥΊ≤ΜΆ§Θ§Ζ÷±πΈΣCys1-Cys4ΓΔCys2-Cys5ΓΔCys3-Cys6ΓΘ÷ΒΒΟ“ΜΧαΒΡ «Θ§“―¥”άΞ≥φ÷–Ζ÷άκΧα»Γ≥ωΚ§”–4ΗωΕΰΝρΦϋΒΡΖά”υΥΊΓΘ
2 Ζά”υΥΊΒΡ…ζΈοΜν–‘ΦΑΤδΉς”ΟΜζ÷Τ
2.1 ΩΙΨζΉς”Ο
Ζά”υΥΊΡήΙΜΆ®ΙΐΕύ÷÷Μζ÷Τ…±ΟπœΗΨζΜρ“÷÷ΤœΗΨζ…ζ≥ΛΓΘΧεΆβ Β―ιœ‘ ΨΘ§Ζά”υΥΊΨΏ”–ΙψΤΉΒΡΩΙΨζΜν–‘Θ§Ε‘ΗοάΦ œ―τ–‘ΨζΓΔΗοάΦ œ“θ–‘ΨζΓΔ―α―θΨζΓΔΡ≥–©’φΨζΓΔΟΖΕΨ¬ί–ΐΧεΓΔΙ≥ΕΥ¬ί–ΐΧεΚΆΖ÷÷ßΗΥΨζΒ»ΨυΨΏ”–Κή«ΩΒΡΩΙΨζΜν–‘ΓΘάΞ≥φΖά”υΥΊΕ‘ΗοάΦ œ―τ–‘œΗΨζΒΡΉς”Ο–ßΙϊΫœ«ΩΘ§ΒΪΕ‘ΗοάΦ œ“θ–‘œΗΨζΒΡΉς”Ο–ßΙϊΫœ≤ν [5] ΓΘΕχ÷≤ΈοΖά”υΥΊ‘ΎΩΙ’φΨζΉς”ΟΖΫΟφΗϋ”–”≈ ΤΘ§“ρΈΣΕ‘÷≤ΈοΕχ―‘Θ§’φΨζ «Ήν≥ΘΦϊΒΡΈΔ…ζΈοΆΰ–≤ [11] ΓΘΖά”υΥΊΒΡΩΙΨζΜζ÷Τ”κœ÷”–ΒΡΩΙ…ζΥΊΒΡΩΙΨζΜζ÷Τ≤ΜΆ§Θ§Ζά”υΥΊ÷ς“ΣΆ®ΙΐΉς”Ο”ΎœΗΨζœΗΑϊΡΛΓΔ÷–ΚΆΖ÷ΟΎΕΨΥΊ“‘Φθ…ΌΗ–»ΨΓΔΉηΕœœΗΨζΫχ»κœΗΑϊ“‘ΦΑ¥ΌΫχΙ≤…ζΨζΕ®÷≤ΥΡΗωΖΫΟφΖΔΜ”ΩΙΨζΉς”ΟΓΘ
2.1.1 Ζά”υΥΊΕ‘œΗΨζœΗΑϊΡΛΒΡΉς”Ο
Ζά”υΥΊΩΙΨζΜν–‘ΒΡΉς”ΟΜζ÷Τ―–ΨΩ÷ς“ΣΜυ”ΎΖά”υΥΊ¥χ’ΐΒγΚ…ΒΡΧΊ–‘ΓΘ“ΜΑψ»œΈΣΘ§Ζά”υΥΊΒΡΩΙΨζΜν–‘÷ς“ΣΧεœ÷‘ΎΤδ”κ≤Γ‘≠ΧεœΗΑϊΡΛΒΡœύΜΞΉς”ΟΘ§’β÷÷œύΜΞΉς”ΟΒΡΚΥ–Ρ «―τάκΉ”Ζά”υΥΊΖ÷Ή”Έϋ“ΐΆβΡΛ”–ΗΚΒγΚ…≥…Ζ÷ΒΡœΗΨζΓΘ’β–©¥χΗΚΒγΚ…ΒΡ≥…Ζ÷÷ς“Σ «ΗοάΦ œ―τ–‘œΗΨζ±μΟφΒΡΝΉ÷§ΥαΚΆΗοάΦ œ“θ–‘œΗΨζΡΛ…œΒΡ÷§ΕύΧ«ΚΆΝΉ÷§ΓΘ»ΥΠ¬-Ζά”υΥΊ3(human Π¬-defensin 3Θ§ hBD3)ΒΡΆΜ±δ Β―ι“≤±μΟςΘ§ΨΜ’ΐΒγΚ…ΒΡ‘ωΕύ ΙΖά”υΥΊ”Β”–ΗϋΗΏΒΡΩΙΨζΜν–‘ [12] ΓΘΆ§ ±Θ§―–ΨΩ’Ώ”ΟΠΝ-ΚΆΠ¬-Ζά”υΥΊΒγΚ…»± ßΆΜ±δΧε―–ΨΩΝΥΖά”υΥΊΫαΙΙ”κΙΠΡήΒΡΙΊœΒΘ§“≤÷Λ ΒΝΥ’β–©Ψ≤ΒγœύΜΞΉς”Ο‘ΎΖά”υΥΊΒΡΩΙΨζΉς”Ο÷–ΒΡ÷Ί“Σ–‘ [13] ΓΘ
Ζά”υΥΊΖ÷Ή”‘ΎœΗΨζœΗΑϊΡΛ…œΨέΦ·÷°ΚσΘ§Ω…ΒΦ÷¬œΗΑϊΡΛΤΤΝ―ΓΔœΗΑϊ≥…Ζ÷–Ι¬©Θ§ΫχΕχ“÷÷ΤœΗΨζΒΡ…ζΟϋΙΐ≥ΧΘ§Ήν÷’ΒΦ÷¬œΗΨζΥάΆωΓΘΡΩ«ΑΘ§÷Ν…Ό”–ΥΡ÷÷ΡΘ–ΆΟη ωΖά”υΥΊΤΤΜΒœΗΨζœΗΑϊΡΛΒΡΙΐ≥ΧΘΚΓΑΨέΦ·Γ±ΡΘ–ΆΓΔΓΑΜΖ–ΈΩΉœΕΓ±ΡΘ–ΆΓΔΓΑΡΨΆΑΑεΓ±ΡΘ–ΆΚΆΓΑΒΊΧΚΓ±ΡΘ–Ά [5] ΓΘ’βΥΡ÷÷ΡΘ–ΆΧα ΨΘ§Ζά”υΥΊΕ‘œΗΨζœΗΑϊΡΛΒΡΤΤΜΒΜζ÷ΤΩ…Ρή“ρ≤ΜΆ§ΒΡΖά”υΥΊΚΆœΗΨζάύ–ΆΕχ”–Υυ≤ΜΆ§ΓΘ
2.1.2 ÷–ΚΆΖ÷ΟΎΕΨΥΊΦθ…ΌœΗΨζΗ–»Ψ
‘Ύ―–ΨΩ»ΥάύΖά”υΥΊ”κΕύ÷÷≤ΜΆ§œΗΨζ≤ζ…ζΒΡœΗΨζΕΨΥΊœύΜΞΉς”ΟΒΡ Β―ι÷–Θ§KudryashovaΒ» [14] ΖΔœ÷Θ§≤ΜΆ§Ζά”υΥΊ ΙœΗΨζΕΨΥΊ ßΜνΒΡΜζ÷Τ”–Οςœ‘ΒΡœύΥΤ–‘Θ§Αϋά®ΕύΦέœύΜΞΉς”ΟΓΔΕ‘œΗΨζΕΨΥΊΒΡΖ«ΨΚ’υ–‘ΜρΨΚ’υ–‘“÷÷ΤΉς”ΟΘ§“‘ΦΑ»Ξ’έΒΰΉς”ΟΓΘΒΪ ”Ο”ΎΥυ”–Ζά”υΥΊΑ–œρΟπΜνœΗΨζΕΨΥΊΒΡΜζ÷ΤΜΙ”–¥ΐΫχ“Μ≤Ϋ―–ΨΩΓΘ
2.1.3 ΉηΕœœΗΨζΫχ»κœΗΑϊ
―–ΨΩΖΔœ÷Θ§Ρ≥–©Ζά”υΥΊ≤ΔΟΜ”–Κή«ΩΒΡ…±Ψζ“÷ΨζΜν–‘Θ§ΒΪ «ΥϋΡήάϊ”ΟΫœ«ΩΒΡΜζ–ΒΖά”υΜν–‘ΤπΒΫΖά÷ΙœΗΨζ«÷»κ»ΥΧεΒΡΉς”ΟΘ§»γ–Γ≥Π“ΰΈ―ΒΉ≤Ω≈Υ œœΗΑϊ≤ζ…ζΒΡHD6ΓΘHD6‘ΎΥφΜζΫαΚœΒΫœΗΨζ±μΟφΒΑΑΉ÷ (»γΨζΟΪΓΔ±όΟΪ)ΚσΘ§Ψ≠άζ”––ρΒΡΉ‘ΉιΉΑΘ§–Έ≥…≤χ»ΤœΗΨζΒΡ‘≠œΥΈ§ΚΆΡ…ΟΉΆχΘ§¥”ΕχΖΔΜ”Ζά÷Ι≥ΠΒά≤Γ‘≠Χε»κ«÷ΒΡΉς”ΟΘ§Τδ÷–Θ§HD6ΉιΑ±Υα-27≤–ΜυΒΡΏδΏρΜΖΚΆΝΝΑ±Υα-32ΒΡCΕΥτ»Μυ÷°ΦδΒΡΨ≤ΒγœύΜΞΉς”Ο≤Έ”κΫιΒΦΝΥ’β÷÷Ή‘ΉιΉΑ [15] ΓΘ“≤”–“Μ–© Β―ιΫαΙϊ±μΟςΘ§HD6Ζά÷ΙœΗΨζ»κ«÷ΒΡΡήΝΠ «ΙψΤΉΒΡΘ§Ε‘ΗοάΦ œ“θ–‘ΜρΗοάΦ œ―τ–‘œΗΨζΟΜ”–Οςœ‘ΒΡ―Γ‘ώ–‘Θ§’β“≤¥”Νμ“ΜΖΫΟφ÷Λ ΒΝΥHD6Ε‘”ΎœΗΨζΒΡΜζ–ΒΖά”υΜν–‘ [16] ΓΘ
2.1.4 ¥ΌΫχΙ≤…ζΨζΕ®÷≤
Ζά”υΥΊ≤ΜΫωΨΏ”–ΩΙΨζΉς”ΟΘ§ΜΙ”–¥ΌΫχΙ≤…ζΨζΕ®÷≤ΒΡΉς”ΟΓΘOuΒ» [17] ΖΔœ÷Θ§≥ΠΒά÷–ΒΡΠΝ-Ζά”υΥΊΕ‘Ι≤…ζάύΗΥΨζΟΜ”–»ΈΚΈ…±ΨζΜν–‘Θ§Ζ¥ΕχΩ…Ά®Ιΐ‘ω«ΩœΗΨζΕ‘≥ΠΒάπΛΡΛΒΡπΛΗΫΉς”ΟΘ§¥ΌΫχΙ≤…ζάύΗΥΨζΕ®÷≤Θ§“‘Έ§≥÷≥ΠΒάΈΔ…ζΈοΒΡΕύ―υ–‘Θ§¥”ΕχΒ÷”υΆβά¥≤Γ‘≠ΧεΒΡ»κ«÷ΓΘ
Ήέ…œΥυ ωΘ§Ζά”υΥΊ≤ΜΫω”–Ζ«≥ΘΙψΤΉΒΡΩΙΨζΉς”ΟΘ§ΤδΩΙΨζΜζ÷Τ“≤Εύ÷÷Εύ―υΓΘ»γΫώΘ§”…”ΎΩΙ…ζΥΊΒΡάΡ”ΟΘ§ΚήΕύΡΆ“©Ψζ÷ξ÷πΫΞ≥ωœ÷Θ§ΗχΝΌ¥≤÷ΈΝΤ¥χά¥ΝΥΚή¥σΒΡΡ―ΧβΓΘΒΪΖά”υΥΊΕάΧΊΕύ―υΒΡΩΙΨζΜζ÷ΤΘ§ΡήΙΜΚήΚΟΒΊΫβΨωœ÷”–ΒΡœΗΨζΡΆ“©Έ ΧβΘ§”–ΆϊΧφ¥ζœ÷”–ΒΡΩΙ…ζΥΊ≥…ΈΣ–¬“Μ¥ζΒΡΩΙΨζ“©ΈοΓΘΕχΖά”υΥΊΒΡΡΘΡβΈοBrilacidin(PMX-30063)ΓΔLytixar(LTX109)Β»“≤Ε‘Εύ÷÷œΗΨζ”–ΩΙ–‘Θ§‘Ύ’κΕ‘Ρ≥–©Η–»Ψ–‘Φ≤≤ΓΒΡ÷ΈΝΤ―–ΨΩ÷–Θ§Ζά”υΥΊΡΘΡβΈο…θ÷Ν“―Ψ≠Ϋχ»κΝΥΝΌ¥≤―–ΨΩΫΉΕΈΘ§≤Δ»ΓΒΟΝΥ≥θ≤ΫΒΡ≥…ΙΠΘ§ «Ζ«≥Θ”–«ΑΨΑΒΡΩΙ…ζΥΊΚρ―Γ“©Έο [ 18, 19] ΓΘ
2.2 ΩΙ≤ΓΕΨΉς”Ο
Ζά”υΥΊΕ‘ΑϋΡΛ≤ΓΕΨΚΆΖ«ΑϋΡΛ≤ΓΕΨΕΦ”–ΩΙ≤ΓΕΨΜν–‘ΓΘΤδΩΙ≤ΓΕΨΜζ÷Τ≥ΐΝΥ“÷÷Τ≤ΓΕΨ»ΎΚœΘ§÷±Ϋ”’κΕ‘≤ΓΕΨΑϋΡΛΓΔΧ«ΒΑΑΉΚΆ“¬Ω«ΆβΘ§ΜΙΡήΫαΚœ≤ΔΒςΫΎΥό÷ςœΗΑϊ±μΟφ ήΧεΓΔΤΤΜΒœΗΑϊΡΎ–≈Κ≈¥ΪΒΦΘ§¥”Εχ“÷÷Τ≤ΓΕΨΗ¥÷Τ [6] ΓΘ»ΜΕχΘ§“≤”–―–ΨΩ±μΟςΘ§Ζά”υΥΊ‘ΎΖΔΜ”ΩΙ≤ΓΕΨΉς”ΟΒΡΆ§ ±Θ§“≤Ρή¥ΌΫχΡ≥–©≤ΓΕΨΕ‘ΜζΧεΒΡΗ–»ΨΓΘ±»»γΘ§HD1“ρΤδΩΙHIVΜν–‘Εχ±ΜΙψΖΚ―–ΨΩΘ§ΒΪΆ§ ±HD1“≤Ρή‘ωΦ”…œΤΛœΗΑϊΒΡΆ®ΆΗ–‘Θ§¥ΌΫχHIVΩγ‘Ϋ…œΤΛΤΝ’œΘ§¥”Εχ¥ΌΫχ≤ΓΕΨΗ–»ΨΚΆ¥Ϊ≤Ξ [20] ΓΘ
2.2.1 Ζά”υΥΊΚΆ≤ΓΕΨ÷°ΦδΒΡ÷±Ϋ”œύΜΞΉς”Ο
Ζά”υΥΊ”κ≤ΓΕΨΩ≈ΝΘΒ»≈δΧεΫαΚœ”–Εύ÷÷ΡΘ ΫΓΘ Ήœ»Θ§¥χ’ΐΒγΚ…ΒΡΖά”υΥΊΡή”κΑϋΙϋ≤ΓΕΨΒΡ÷§÷ ΥΪΖ÷Ή”≤ψ÷±Ϋ”Ής”ΟΘ§¥”ΕχΤΤΜΒ≤ΓΕΨΘ§ ΙΤδ≤ΜΨΏ¥Ϊ»Ψ–‘ΓΘ»ΜΕχΖά”υΥΊΕ‘÷§÷ ΥΪΖ÷Ή”≤ψΒΡΤΤΜΒ»ΓΨω”ΎΑϋΡΛ÷§÷ ≥…Ζ÷ΒΡ≤ν“λΘ§ΗΚΒγΚ…ΝΉ÷§Ε‘Ζά”υΥΊΟτΗ–Θ§Εχ÷––‘ΥΪ≤ψ÷§ΡΛΕ‘Ζά”υΥΊΜυ±Ψ≤ΜΟτΗ–ΓΘ≤Δ«“”…”ΎΖά”υΥΊ «ΝΫ–‘Ζ÷Ή”Θ§Ζά”υΥΊ“≤ΡήΆ®Ιΐ«Ή÷§–‘”κ≤ΓΕΨΩ≈ΝΘœύΜΞΉς”Ο [6] ΓΘΒΎΕΰΘ§Ζά”υΥΊΨΏ”–”κ≤ΓΕΨ±μΟφΧ«ΒΑΑΉΫαΚœΒΡΜν–‘ [ 21, 22] ΓΘ”κ≤ΓΕΨΗΫΉ≈ΒΑΑΉΫαΚœΒΡΖά”υΥΊΩ…ΤΤΜΒ≤ΓΕΨ”κœΗΑϊΡΎΙΊΦϋ ήΧεΒΡœύΜΞΉς”ΟΓΘHD5Ω…ΫαΚœ”κΒΞ¥Ωπε’ν≤ΓΕΨ-2«÷»κΚΆπΛΗΫΡήΝΠœύΙΊΒΡ≤ΓΕΨ±μΟφΧ«ΒΑΑΉDΘ§ΫχΕχΦθ…Ό≤ΓΕΨΕ‘œΗΑϊΒΡπΛΗΫ [23] ΓΘ≥ΐ¥Υ÷°ΆβΘ§HD1ΓΔHD2ΚΆHD5Ω…ΫαΚœHIV-1±μΟφΒΡgp120“‘ΦΑœΗΑϊ±μΟφ ήΧεCD4Θ§¥”ΕχΦθ…ΌHIVΕ‘ΜζΧεΟβ“ΏœΗΑϊΒΡΤΤΜΒ [24] ΓΘΉή÷°Θ§Ζά”υΥΊΩ…÷±Ϋ”Η…»≈HIV-1ΚΆΒΞ¥Ωπε’ν≤ΓΕΨ-2”κœΗΑϊΒΡΫαΚœΚΆΗΫΉ≈ΓΘΆ§ ±Θ§”κ≤ΓΕΨΩ≈ΝΘΫαΚœΒΡΖά”υΥΊΖ÷Ή”ΡήΙΜ”’ΒΦ≤ΓΕΨΩ≈ΝΘΒΡΨέΦ·Θ§¥ΌΫχΩΙ‘≠≥ ΒίœΗΑϊΕ‘≤ΓΕΨΩ≈ΝΘΒΡ…ψ»ΓΓΔ¥ΠάμΘ§“≤ΡήΆ®Ιΐ÷±Ϋ”ΉηΑ≠≤ΓΕΨ”κœΗΑϊΫαΚœΜρ Ι≥…Ά≈ΒΡ≤ΓΕΨΫχ»κΫœ…ΌΒΡœΗΑϊ¥”ΕχΫΒΒΆ≤ΓΕΨΒΡ¥Ϊ»Ψ–‘ΓΘ≥ΐ¥Υ÷°ΆβΘ§Ε‘–¬ΙΎ≤ΓΕΨΒΡ―–ΨΩΖΔœ÷Θ§HD1ΚΆΠ»-Ζά”υΥΊΡήΙΜ”’ΒΦ–¬ΙΎ≤ΓΕΨΦ§ΆΜΒΑΑΉΒΡ»Ξ’έΒΰΚΆ≥ΝΒμΘ§≤Δ“÷÷ΤΤδ”κ―ΣΙήΫτ’≈ΥΊΉΣΜ·ΟΗ2ΒΡœύΜΞΉς”ΟΘ§¥”ΕχΦθ…Ό–¬ΙΎ≤ΓΕΨΕ‘»ΥΧεΒΡΗ–»Ψ [25] ΓΘ
‘Ό’ΏΘ§Ζά”υΥΊΜΙΡήΆ®ΙΐΉη÷Ι≤ΓΕΨΆ―Ω«¥”ΕχΖΔΜ”ΩΙ≤ΓΕΨΜν–‘ΓΘ―–ΨΩΖΔœ÷Θ§»ΥΧε÷–ΠΝ-Ζά”υΥΊΧΊ±π «HD5Ε‘“÷÷Τ»Υ»ιΆΖΉ¥Νω≤ΓΕΨΗ–»ΨΖ«≥Θ”––ß [26] ΓΘ»Υ»ιΆΖΉ¥Νω≤ΓΕΨΒΡ“¬Ω«ΒΑΑΉΚΆHD5œύΜΞΉς”ΟΘ§ Ι≤ΓΕΨ“¬Ω«ΗϋΦ”Έ»Ε®Θ§÷±Ϋ”Ήη÷ΙΝΥ≤ΓΕΨΒΡΆ―Ω«Θ§≤ΔΫΪ≤ΓΕΨΩ≈ΝΘΉΣ“ΤΒΫ»ήΟΗΧε÷–Θ§¥”Εχ÷–ΕœΝΥ≤ΓΕΨΒΡΗ¥÷ΤΓΘ
≤ΓΕΨΫω¥©ΆΗΥό÷ςœΗΑϊΡΛΜΙ≤ΜΡήΆξ≥…Η–»ΨΘ§≤ΓΕΨΒΡΉΣ¬ΦΓΔΒΑΑΉ÷ ΒΡ≤ζ…ζΓΔ≤ΓΕΨΒΡΉιΉΑΚΆ ΆΖ≈ΕΦ±Ί–κΖΔ…ζ≤≈ΡήΆξ≥…“ΜΗωΗ¥÷Τ÷ήΤΎΓΘ’β–©≤Ϋ÷ηΈΣΖά”υΥΊΧαΙ©ΝΥΉηΕœ≤ΓΕΨΗ–»ΨΒΡΜζΜαΓΘ
2.2.2 ’κΕ‘Υό÷ςœΗΑϊΒΡΩΙ≤ΓΕΨΜζ÷Τ
Ε‘”ΎΥό÷ςœΗΑϊά¥ΥΒΘ§Ζά”υΥΊΡήΙΜ”κΥό÷ςœΗΑϊ±μΟφΒΡΒΑΑΉ÷ ΫαΚœΘ§¥”Εχ“÷÷Τ≤ΓΕΨ”κΥό÷ςœΗΑϊΒΡ»ΎΚœΓΘΕ‘”ΎHIV-1ΒΡ»κ«÷Θ§HD1ΡήΆ®Ιΐ”κœΗΑϊΡΛ±μΟφ ήΧεΫαΚœΫΒΒΆœΗΑϊΡΛ±μΟφCD4ΓΔCCR5ΚΆCXCR4 ήΧεΒΡΝςΕ·–‘Θ§¥”Εχ“÷÷Τ≤ΓΕΨ”κœΗΑϊΡΛΒΡ»ΎΚœ [27] ΓΘΕ‘”Ύ–¬–ΆΙΎΉ¥≤ΓΕΨSARS-CoV-2Θ§HD5ΡήΙΜΖβ±’œΗΑϊ±μΟφΒΡ―ΣΙήΫτ’≈ΥΊΉΣΜ·ΟΗ2 ήΧεΘ§¥”ΕχΦθ…Ό≤ΓΕΨΕ‘»ΥΧεΒΡΗ–»Ψ [28] ΓΘ¥ΥΆβΘ§–μΕύ≤ΓΕΨ(»γHIV-1ΓΔΚτΈϋΒάΚœΑϊ≤ΓΕΨΒ»)‘ΎΗ–»ΨΥό÷ςœΗΑϊΒΡΙΐ≥Χ÷–ΕΦ”–ΒΑΑΉΦΛΟΗCΒΡ≤Έ”κ [ 29, 30] ΓΘ”–―–ΨΩΖΔœ÷Θ§Ρ≥–©Ζά”υΥΊΡήΙΜΆ®Ιΐ“÷÷ΤΒΑΑΉΦΛΟΗCΒΡΜν–‘≤Έ”κ“÷÷Τ≤ΓΕΨΕ‘»ΥΧεΒΡΗ–»ΨΓΘ’β“ΜΖΔœ÷±μΟςΘ§ΗΡ±δΜρ“÷÷ΤœΗΑϊΡΎ–≈Κ≈¥ΪΒΦΆ®¬ΖΩ…Ρή «Ζά”υΥΊΩΙ≤ΓΕΨΜν–‘ΒΡΝμ“Μ÷÷Μζ÷Τ [ 31, 32] ΓΘΉέ…œΥυ ωΘ§Ζά”υΥΊΕ‘≤ΓΕΨΒΡ“÷…±Ής”Ο…φΦΑΕύΖΫΟφΜζ÷ΤΓΘΖά”υΥΊΖΔΜ”ΩΙ≤ΓΕΨΜν–‘…φΦΑ≤ΓΕΨ¥”¥©‘ΫœΗΑϊΡΛΩΣ Φ÷Ν”κœΗΑϊΡΎΒΡΑ–±ξœύΜΞΉς”ΟΒΡ»ΪΙΐ≥ΧΓΘ
―–ΨΩΖΔœ÷Θ§ΚήΕύ≤ΓΕΨΗ–»ΨΡήΙΜ“ΐΤπΜζΧεΖά”υΥΊ±μ¥οΒΡ‘ωΦ”Θ§ΕχΧα«Α‘Ύ–Γ σΧεΡΎΉΔ…δΙΐΝΩΒΡΖά”υΥΊΡήΙΜΦθ…Ό≤ΓΕΨΗ–»ΨΒΈΕ» [33] ΓΘ’β–©ΫαΙϊΧα ΨΘ§‘Ύ»ΥΧεΡΎ‘ΛΖά–‘ΒΊ Ι”Ο¥σΝΩΖά”υΥΊ «Ζώ“≤ΡήΙΜΦθ…Ό≤ΓΕΨΗ–»ΨΒΈΕ»Θ§¥”ΕχΤπΒΫ‘ΛΖάΗ–»ΨΒΡΉς”Ο? ΒΪ «Θ§‘Ύ»ΥΧεΡΎ‘ΛΖά–‘ΒΊ Ι”Ο¥σΝΩΖά”υΥΊΒΡάϊ±ΉΜΙ–η“ΣΗϋ…ν»κΒΡΧΫΥςΚΆΖ÷ΈωΓΘ“ρΈΣ―–ΨΩΖΔœ÷Θ§ΗΏ≈®Ε»ΒΡΖά”υΥΊ”–œΗΑϊΕΨ–‘Ής”Ο≤Δ«“Ρή¥ΌΫχΡ≥–©Φ≤≤ΓΒΡΖΔ…ζΖΔ’Ι [ 34, 35] ΓΘ“ρ¥ΥΘ§‘ΎΈ¥ά¥ Ι”ΟΖά”υΥΊ‘ΛΖάΗς÷÷≤ΓΕΨΗ–»Ψ ±Θ§ΜΙ”ΠΉέΚœΩΦ¬«Ζά”υΥΊΗχΜζΧε¥χά¥ΒΡΗς÷÷Η±Ής”ΟΓΘ‘θ―υΙφ±ή’β–©Η±Ής”ΟΘ§Μρ’Ώ‘θ―υ’“ΒΫ“ΜΗω Ι»ΥάύΜώ“φΉν¥σΜ·ΒΡΤΫΚβΒψΘ§ΜΙ–η“ΣΫχ––ΗϋΦ”…ν»κΒΡ―–ΨΩΓΘΕχΕ‘”ΎΡΩ«Α―œ÷ΊΆΰ–≤»ΥάύΫΓΩΒΒΡ–¬–ΆΙΎΉ¥≤ΓΕΨά¥ΥΒΘ§“―”–―–ΨΩ±μΟςΘ§Ζά”υΥΊΡΘΡβΈοBrilacidin(PMX-30063)ΡήΙΜΆ§ ±Ής”Ο”ΎΑ–œΗΑϊΚΆ≤ΓΕΨ¥”ΕχΦθ…Ό≤ΓΕΨΫχ»κœΗΑϊΘΜΝμΆβΘ§Brilacidin(PMX-30063)”κ»πΒ¬ΈςΈΛΒΡΝΣΚœ Ι”Ο“≤±μœ÷≥ω–≠Ά§ΒΡΩΙ≤ΓΕΨΜν–‘ [36] ΓΘ
2.3 ΩΙ÷ΉΝω–ß”Π
Τπ‘¥”Ύ…œΤΛΉι÷·ΒΡΑ© «ΡΩ«ΑΉν≥ΘΦϊΒΡΕώ–‘÷ΉΝωΘ§ΧΊ±π «”……œΤΛœΗΑϊΖ÷ΟΎΒΡΠ¬-Ζά”υΥΊ≤Έ”κΝΥΚήΕύ…œΤΛ‘¥–‘÷ΉΝωΒΡΖΔ…ζ”κΖΔ’ΙΘ§ ήΒΫΝΥΙψΖΚΙΊΉΔΓΘ―–ΨΩΖΔœ÷Θ§ hBD1‘Ύ«ΑΝ–œΌΑ©Β»Α©÷Δ÷–±μ¥οœ¬ΒςΘ§≤Δ«“Ω…ΡήΉςΈΣ÷ΉΝω“÷÷Τ“ρΉ”ΖΔΜ”Ής”Ο [37] ΓΘ‘Ύ±ϊΗΈ≤ΓΕΨΗ–»Ψ“ΐΤπΒΡΗΈœΗΑϊΑ©÷–Θ§ hBD1ΒΡœ¬ΒςΡήΙΜ ΙE-ΗΤπΛΒΑΑΉœ¬ΒςΓΔΗΈœΗΑϊ…ζ≥Λ“ρΉ”ΒςΫΎΒΡά“Α±ΥαΦΛΟΗΒΉΈο…œΒςΘ§“÷÷ΤΑ©œΗΑϊΒΡ…ζ≥ΛΚΆ«®“Τ [38] ΓΘΕχ hBD3»¥‘ΎΆΖΨ±≤Ω÷ΉΝω“‘ΦΑΙ§Ψ±Α©Β»Εύ÷÷Α©÷Δ÷–ΕΦ”–Ιΐ±μ¥ο [39] ΓΘ≤Δ«“Θ§œ÷“―ΖΔœ÷÷ΉΝωœΗΑϊΖ÷ΟΎΒΡ hBD3Ρή¥ΧΦΛ÷ΉΝω…ζ≥ΛΚΆ«®“ΤΘ§ Ι÷ΉΝωœΗΑϊΕ‘ΒρΆω≤ζ…ζΒ÷ΩΙΝΠΘ§≤Δ”–÷ζ”Ύ’–ΡΦ”κ÷ΉΝωΫχ’ΙœύΙΊΒΡΨό …œΗΑϊΓΔΒΞΚΥœΗΑϊΓΔΈ¥≥… λ ςΆΜΉ¥œΗΑϊΓΔΦ«“δTœΗΑϊΚΆΖ ¥σœΗΑϊ [ 39, 40] ΓΘ
Ζά”υΥΊΒΡ±μ¥οΝΩ‘ΎΚήΕύάύ–ΆΒΡ÷ΉΝωΉι÷·÷–ΕΦ”–Οςœ‘ΒΡ±δΜ·Θ§ΧΊ±π «”……œΤΛœΗΑϊΖ÷ΟΎΒΡΠ¬-Ζά”υΥΊ≤Έ”κΝΥΚήΕύ…œΤΛ‘¥–‘÷ΉΝωΒΡΖΔ…ζ”κΖΔ’ΙΘ§“Μ–©Ζά”υΥΊΕ‘÷ΉΝω”–“÷÷ΤΉς”ΟΘ§Νμ“Μ–©Ζά”υΥΊ‘ρ‘Ύ÷ΉΝω÷–Ιΐ±μ¥ο≤Δ¥ΌΫχ÷ΉΝωΈΔΜΖΨ≥ΒΡ–Έ≥…Θ§ΒΪ’β≤Δ≤ΜΖΝΑ≠Ζά”υΥΊ‘ΎΈ¥ά¥ΒΡΝΌ¥≤ΙΛΉς÷–Ω…ΡήΉςΈΣ÷ΉΝωΒΡ…ζΈο―ß±ξ÷ΨΚΆ÷ΈΝΤΑ–ΒψΓΘΡΩ«Α“―”–―–ΨΩ±μΟςΘ§HD6ΒΡΥ°ΤΫ”κ¥σ≥ΠΑ©ΜΦ’ΏΒΡΉήΧε…ζ¥φ¬ ≥ ΗΚœύΙΊΘ§ «¥σ≥ΠΑ©ΒΡΕάΝΔ‘ΛΚσ±ξ÷ΨΈοΘ§≤Δ«“HD6‘Ύ¥σ≥ΠΑ©÷–Ρή¥ΌΫχΑ©œΗΑϊΒΡ‘ω÷≥ΓΔ«®“ΤΚΆ«÷œ°Θ§ «ΗΟΦ≤≤ΓΒΡΝΦΚΟ÷ΈΝΤΑ–Βψ [41] ΓΘ
2.4 ―Ή÷ΔΒςΫΎΉς”Ο
≤ΜΆ§Ζά”υΥΊΆ®Ιΐ≤ΜΆ§ΒΡΜζ÷ΤΉς”Ο”ΎΧΊΕ®œΗΑϊΘ§Φ»Ρή”’ΒΦ―Ή÷ΔΘ§”÷Ρή“÷÷Τ―Ή÷ΔΓΘΖά”υΥΊΒΡΩΙ―ΉΉς”ΟΚΆ¥Ό―ΉΉς”Ο”…Τδ±μ¥οΥ°ΤΫΨωΕ®Θ§Ά§ ±“≤»ΓΨω”ΎΦ≤≤ΓΉ¥Χ§ΚΆ≤Γ‘≠Χ屩¬ΕΉ¥Χ§ΓΘ‘Ύ¥Ό―ΉΖΫΟφΘ§Εύœν Β―ι±μΟςΘ§Π¬-Ζά”υΥΊΩ…Ά®Ιΐ”κΕύΗωœΗΑϊ±μΟφ ήΧεΫαΚœ¥ΌΫχ¥Ό―Ή–‘œΗΑϊ“ρΉ”ΚΆ«ςΜ·“ρΉ”ΒΡ≤ζ…ζΘ§‘ω«ΩΟβ“ΏΖ¥”ΠΓΘΠ¬-Ζά”υΥΊ≤ΜΫωΡήΆ®ΙΐCCR2ΫιΒΦΒΞΚΥ/Ψό …œΗΑϊΒΡΖ¥”Π–‘«®“ΤΘ§“≤Ω…“‘Ά®Ιΐ”κCCR6ΫαΚœΘ§”’ΒΦCD4 +Φ«“δTœΗΑϊΚΆΈ¥≥… λ ςΆΜΉ¥œΗΑϊΒΡ«ςΜ·Ής”Ο [40] ΓΘ»ΥΒΡΡΎ‘¥–‘Ζά”υΥΊ hBD2Ρή‘ω«ΩΚΥή’ΥαΙ―ΨέΜ·ΫαΙΙ”ρ―υ ήΧεΚΆΈ§ΦΉΥα”’ΒΦΜυ“ρΔώ ήΧεΒΡ±μ¥ο≤ΔΡήΙΜ”’ΒΦΤδ“ΉΈΜΘ§¥”Εχ÷±Ϋ”ΦΛΜν¥”ΗΙΡΛΖ÷άκ≥ωΒΡΖ ¥σœΗΑϊΒΡ¥Ό―ΉΚΆ«®“ΤΖ¥”ΠΓΘhBD2ΜΙΡή‘ω«ΩΒΞΚΥœΗΑϊΕ‘ΚΥή’ΥαΙ―ΨέΜ·ΫαΙΙ”ρ―υ ήΧεΚΆΈ§ΦΉΥα”’ΒΦΜυ“ρΔώ ήΧεœύ”Π≈δΧεΒΡΟτΗ––‘Θ§¥”Εχ‘ω«ΩΒΞΚΥœΗΑϊ‘Ύ―Ή÷Δ÷–ΒΡΉς”Ο [42] ΓΘ
Ά§ ±Θ§Β±≤Γ‘≠Χε±Μ«ε≥ΐΚσΘ§Ζά”υΥΊ“≤Ρή“÷÷Τ―Ή÷ΔΒΡΖΔ’ΙΘ§Φθ…Ό―Ή÷ΔΖ¥”ΠΕ‘Ήι÷·‘λ≥…ΒΡΥπ…ΥΓΘ―ά÷ή―ΉΒΡ÷ΈΝΤ÷–Θ§ hBD3Ω…Ά®ΙΐΩΙ―ΉΉς”ΟΖΔΜ”≥…Ι«¥ΌΫχΦΝΒΡΉς”ΟΘ§¥”Εχ¥ΌΫχ―ά÷ήΉι÷·ΒΡ–όΗ¥‘Ό…ζ [43] ΓΘ‘ΎΦ±–‘ΖΈΥπ…ΥΒΡ Β―ιΡΘ–Ά÷–Θ§―–ΨΩ’ΏΖΔœ÷Θ§ΚψΚ”ΚοΠ»-Ζά”υΥΊΡήΙΜΆ®Ιΐ“÷÷Τ÷––‘ΝΘœΗΑϊ«ςΜ·ΚΆπΛΗΫΘ§Φθ»θΖΈ≈ίΨό …œΗΑϊΒΡ¥Ό―ΉœΗΑϊ“ρΉ”ΚΆΜυ“ρΒΡ±μ¥οΘ§¥”ΕχΜΚΫβΤχΒά―Ή÷Δ [44] ΓΘ
ΠΝΓΔΠ¬-Ζά”υΥΊΆ®≥ΘΚ§”–6ΗωΑκκΉΑ±Υα≤–ΜυΘ§ΥϋΟ«‘ΎΖ÷Ή”ΡΎ–Έ≥…ΕΰΝρΦϋΘ§¥ΥΫαΙΙΧΊ’ςάύΥΤ”ΎΉς”Ο”ΎΦΊΆ®ΒάΒΡΒδ–ΆΕΨΥΊκΡ [45] ΓΘΫϋΡξά¥―–ΨΩΖΔœ÷Θ§HD1Ω…“‘ΉςΈΣ“Μ÷÷ΕΨ–‘ΉηΕœΦΝΚΆ“Μ÷÷–¬ΒΡ–ό ΈΦΝΆ§ ±ΫαΚœΒΫΩγΡΛ¬ί–ΐS1-S2Ν§Ϋ”Ή”ΚΆKv1.3Ά®ΒάΒΡΩΉΒά«χ”ρΘ§ΕχHD5“≤ΡήΉςΈΣ“Μ÷÷ΕΨ–‘ΉηΕœΦΝΫαΚœΒΫKv1.3Ά®ΒάΒΡΩΉΒά«χ”ρ [46] ΓΘΦΊάκΉ”Ά®ΒάKv1.3‘ΎTœΗΑϊΜνΜ·÷–”–ΙΊΦϋ–‘Ής”ΟΘ§ΉςΈΣKv1.3Ά®ΒάΒΡ“Μ÷÷–¬–ΆΡΎ‘¥“÷÷ΤΦΝΘ§»ΥάύΖά”υΥΊ”κTœΗΑϊ÷–ΒΡKv1.3Ά®ΒάΫαΚœΩ…ΡήΜα”ΑœλœΗΑϊ“ρΉ”Ζ÷ΟΎΘ§’βΈΣΖά”υΥΊ‘Ύ ”Π–‘Οβ“Ώ÷–ΒΡΉς”ΟΧαΙ©ΝΥ–¬ΒΡ÷ΛΨί [46] ΓΘ
’β–©―–ΨΩΫαΙϊΧα ΨΘ§Ω…“‘άϊ”Ο≤ΜΆ§Ζά”υΥΊΕ‘ΜζΧε―Ή÷ΔΒΡ≤ΜΆ§ΒςΫΎΉς”ΟΕχΜώ“φΓΘΡΩ«ΑΝΌ¥≤…œΗ–»Ψ’οΕœΒΡΫπ±ξΉΦ «œΗΨζ≈ύ―χ≥ ―τ–‘Θ§ΒΪœΗΨζ≈ύ―χΥυ–η ±Φδ≥ΛΓΔΦΌ―τ/“θ–‘¬ ΗΏΘ§≤Δ«““Ή ήΩΙ…ζΥΊΒΡ”ΑœλΓΘΕχΖά”υΥΊΉςΈΣΜζΧεΟβ“ΏœΒΆ≥ΒΡ÷Ί“ΣΉι≥…≥…Ζ÷Θ§‘Ύ”ωΒΫœΗΨζΒΡ»κ«÷ ±Ω…œ‘÷χ…ΐΗΏΘ§Ω…ΉςΈΣœΗΨζΗ–»ΨΒΡ‘γΤΎ’οΕœ÷Η±ξ÷°“ΜΓΘΡΩ«ΑΘ§ΟάΙζ ≥ΤΖΚΆ“©ΈοΙήάμΨ÷“―Ψ≠≈ζΉΦΝΥΠΝ-Ζά”υΥΊΒΡΦλ―ιΩ…ΉςΈΣΙ«ΩΤΦΌΧε÷ήΈßΗ–»ΨΝΌ¥≤’οΕœΒΡΗ®÷ζ ÷ΕΈ [47] ΓΘΕχ‘Ύ–¬ΙΎΜΦ’Ώ÷–Θ§―Σ«εΖά”υΥΊΥ°ΤΫ“≤”–œ‘÷χ‘ωΗΏ [48] ΓΘ‘Ύœχ¥≠÷ΈΝΤ÷–Θ§÷ΈΝΤ–‘ΒΡ±«ΡΎ”Π”ΟΖά”υΥΊ“≤Ρήœ‘÷χΫΒΒΆœχ¥≠œύΙΊœΗΑϊ“ρΉ”ΑΉœΗΑϊΫιΥΊ-9ΚΆΑΉœΗΑϊΫιΥΊ-13“‘ΦΑΤδΥϊΟβ“ΏΒςΫΎœΗΑϊ“ρΉ”ΒΡΥ°ΤΫΘ§¥”Εχ”––ß“÷÷ΤΤχΒάΗΏΖ¥”Π–‘ [49] ΓΘ≥ΐ¥Υ÷°ΆβΘ§Ζά”υΥΊΜΙ”κΩΥ¬όΕς≤ΓΒ»≥ΠΒάΉ‘…μΟβ“Ώ–‘Φ≤≤ΓœύΙΊ [15] ΓΘ
3 ’ΙΆϊ
Ζά”υΥΊ «…ζΈοΧεœ»ΧλΟβ“ΏœΒΆ≥ΒΡ÷Ί“ΣΉι≥…≤ΩΖ÷Θ§Μυ”ΎΉ‘…μΕάΧΊΒΡΫαΙΙΘ§Ζά”υΥΊΨΏ”–ΚήΕύ…ζΈο―ßΜν–‘Θ§Τδœύ”ΠΖ÷Ή”Ής”ΟΜζ÷Τ“≤ΚήΗ¥‘”Θ§…––ηΫχ“Μ≤ΫΒΡ―–ΨΩΧΫΥςΓΘ
ΡΩ«ΑΘ§Ζά”υΥΊΒΡΜώ»ΓΆΨΨΕ÷ς“Σ”–»ΐ÷÷Θ§Ζ÷±πΈΣ¥”œΗΑϊΜρΧε“Κ÷–Χα»ΓΓΔΜ·―ßΚœ≥…“‘ΦΑΜυ“ρΙΛ≥Χ÷Τ±ΗΓΘΒΪΖά”υΥΊ‘ΎΉι÷·÷–ΒΡ±μ¥οΫœ…ΌΘ§¥ΩΜ·ΙΛ“’Η¥‘”Θ§≥…±Ψ“≤ΫœΗΏΘ§≤ζΝΩ“≤Ϋœ…ΌΘ§“ρ¥ΥΘ§Μυ“ρΙΛ≥Χ÷Τ±ΗΈό“…≥…ΈΣΡΩ«ΑΜώ»ΓΖά”υΥΊΒΡ Ή―ΓΖΫΖ®ΓΘ≤Δ«“Θ§Ζά”υΥΊΒΡΡΘΡβΈο“≤ΨΏ”–άύΖά”υΥΊΒΡ…ζΈο―ßΜν–‘Θ§≤Δ«“‘ΎΝΌ¥≤―–ΨΩ÷–“―»ΓΒΟΫχ’ΙΘ§“≤Ω…ΉςΈΣΈ¥ά¥“©Έο―–ΖΔΫχ“Μ≤Ϋ―–ΨΩΒΡΖΫœρΓΘ
Μυ”ΎΕ‘Ζά”υΥΊœ÷”–ΫαΙΙΒΡ―–ΨΩΘ§»ΥΟ«Ω…“‘ΗυΨίΖά”υΥΊΒΡΜν–‘«χ”ρ”κΙ«Φή…ηΦΤ–¬ΒΡΩΙΨζ“©ΈοΓΘ»γΠ»-Ζά”υΥΊΒΡΜΖ–ΈΙ«ΦήΗ≥”ηΝΥΖ÷Ή”Ηϋ«ΩΒΡΈ»Ε®–‘Θ§Ω…“‘ΉςΈΣ–¬–Ά“©ΈοΒΡΫαΙΙ÷ßΦήΘ§ΧαΗΏ“©ΈοΒΡΈ»Ε®–‘ΓΘΠ¬-Ζά”υΥΊΒΡΠΟ-ΚΥ–Ρ «Τδ÷ς“ΣΒΡΜν–‘«χ”ρΘ§Ω…ΫΪΗΟΜν–‘«χ”ρ”κΤδΥϊ÷÷άύΖά”υΥΊΜρΤδΥϊ“©ΈοΒΡΜν–‘«χ”ρΉιΚœ–Έ≥…ΨΏ”–Εύ÷÷Μν–‘Εύ÷÷Μζ÷ΤΒΡ–¬“©ΈοΓΘ≥ΐ¥Υ÷°ΆβΘ§œ÷“―”–―–ΨΩ±μΟςΘ§ΩΙ…ζΥΊ”κΖά”υΥΊ”––≠Ά§Ής”ΟΘ§“≤Ω…ΗυΨί’β“Μ―–ΨΩΫαΙϊΘ§ΫΪ≤ΜΆ§÷÷άύΒΡΖά”υΥΊΚΆΩΙ…ζΥΊ≈δΈιΘ§ΩΣΖΔΝΣΚœ”Ο“©ΖΫΑΗΘ§ΧαΗΏΝΤ–ß [50] ΓΘ
ΡΩ«ΑΘ§“―”–―–ΨΩ’ΏΖΔœ÷Θ§Ζά”υΥΊΩ…ΉςΈΣ…ζΈο±ξ÷ΨΈο÷°“Μ≤Έ”κ‘Λ≤β≤Γ»ΥΒΡΙβΟτ–‘Θ§¥”ΕχΗ®÷ζœΒΆ≥–‘ΚλΑΏά«¥·ΒΡ’οΕœ [51] ΓΘ”κ¥ΥΆ§ ±Θ§Ζά”υΥΊ”κ―Ή÷Δ–‘≥Π≤ΓΒΡ―–ΨΩ“≤»’“φ‘ωΕύΘ§Ρ«Ο¥ΤδΥϊΒΡΉ‘…μΟβ“Ώ–‘Φ≤≤Γ «Ζώ“≤”κΖά”υΥΊœύΙΊΘ§Ζά”υΥΊΕ‘”Ύ’β–©Ή‘…μΟβ“Ώ–‘Φ≤≤ΓΒΡ’οΕœΚΆ÷ΈΝΤ «Ζώ”–Αο÷ζΘ§“≤ «“ΜΗω÷ΒΒΟ―–ΨΩΒΡΖΫœρΓΘ‘ΎΈ¥ά¥ΒΡΖά”υΥΊ”Π”Ο―–ΨΩ÷–Θ§ΝΌ¥≤ΡΆ“©Έ Χβ“≤÷ΒΒΟΙΊΉΔΓΘΕχ’κΕ‘ΡΩ«ΑΦ§ ÷ΒΡ–¬ΙΎ≤ΓΕΨ“ΏΟγΒΡΩΣΖΔ―–ΨΩΘ§“≤”–―–ΨΩ’ΏΫΪΠ¬-Ζά”υΥΊΉτΦΝ”κ≤ΓΕΨΒΡΕύ±μΈΜκΡΝ§Ϋ”Θ§ ΙΗΟΕύ±μΈΜκΡ±μœ÷≥ωΝΥΫœΗΏΒΡΩΙ‘≠–‘Θ§Ω…”Π”Ο”ΎΒ±«Α–¬ΙΎΖΈ―ΉΒΡ‘ΛΖά [52] ΓΘ
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷ΨΓΘ
