
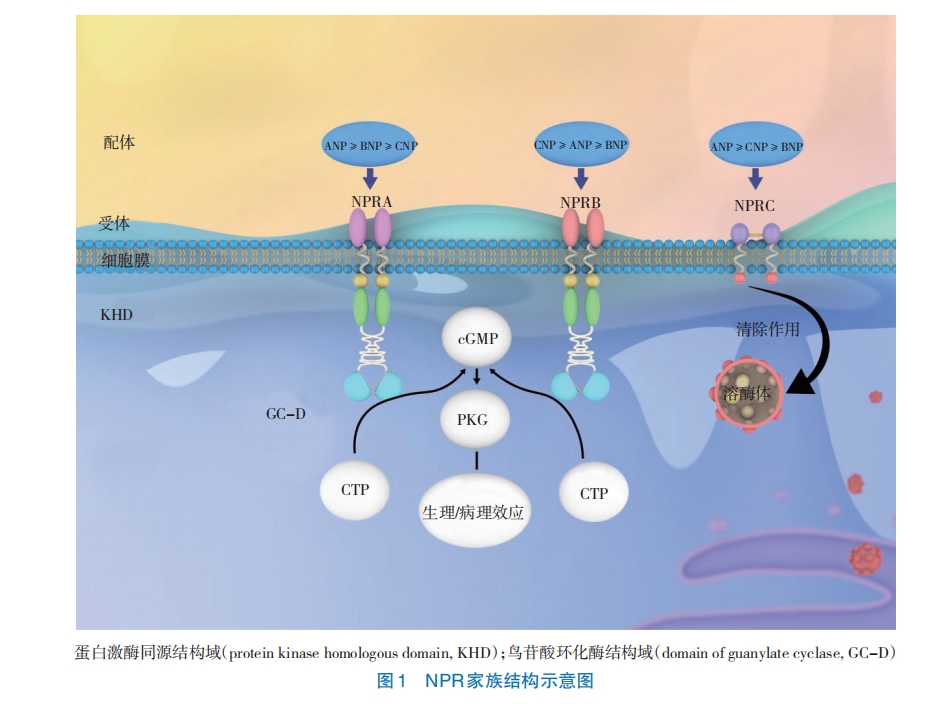
’Σ“ΣΘΚάϊΡΤκΡ ήΧεΘ®NPRΘ©Φ“ΉεΑϋά®άϊΡΤκΡ ήΧεAΘ®NPRAΘ©ΓΔάϊΡΤκΡ ήΧεBΘ®NPRBΘ©ΦΑάϊΡΤκΡ ήΧεCΘ®NPRCΘ©ΓΘ≥ΐNPRCΓΔNPRAΚΆNPRB τ”ΎΡώή’ΥαΜΖΜ·ΟΗ ήΧεΦ“ΉεΒΡ≥…‘±ΓΘNPRΦ“ΉεΙψΖΚΖ÷≤Φ”Ύ»ΥΧε÷–Θ§ΨΏ”–Εύ÷÷…ζάμΙΠΡήΓΘNPRΦ“Ήε‘Ύ–μΕύΕώ–‘÷ΉΝω÷–±μ¥οΘ§‘Ύ÷ΉΝωΒΡΖΔ…ζΖΔ’Ι÷–ΤπΉ≈÷Ί“ΣΒςΩΊΉς”ΟΓΘNPRΦ“Ήε‘ΎΕώ–‘÷ΉΝω÷–ΒΡ±μ¥οΚΆΙΠΡή“―‘Ύ–μΕύΕ·ΈοΡΘ–ΆΚΆ―–ΨΩ÷–ΒΟΒΫ÷Λ ΒΘ§ΒΪNPRΦ“Ήε±μ¥οΒΡΜζ÷Τ…–≤Μ«ε≥ΰΘ§Ε‘ΤδΜζ÷ΤΒΡ―–ΨΩ…–Έ¥”Π”Ο”Ύ÷ΉΝωΒΡΝΌ¥≤÷ΈΝΤΓΘΗΟΈΡΉέ ωΝΥNPRΦ“Ήε‘ΎΕώ–‘÷ΉΝωΖΔ…ζΓΔΖΔ’Ι÷–ΒΡΉς”ΟΦΑΤδ«±‘ΎΒΡΝΌ¥≤Φέ÷ΒΓΘ
άϊΡΤκΡ ήΧεΘ®natriuretic peptide receptor, NPRΘ©“‘ΤδΙψΖΚΒΡ…ζάμΙΠΡήΕχΈ≈ΟϊΘ§≥ΐΝΥΈ»Ε®–Ρ―ΣΙήœΒΆ≥ΆβΘ§ΤδΜΙ”κΟβ“ΏΓΔ―Ή÷ΔΦΑΑ©÷Δ”–ΙΊ[1-3]ΓΘΡΩ«ΑΈΣ÷ΙΘ§“―ΖΔœ÷ΝΥ–ΡΖΩΡΤΡρκΡΘ®atrial natriuretic peptide, ANPΘ©ΓΔΡ‘άϊΡΤκΡΘ®brain natriuretic peptide, BNPΘ©ΓΔC–ΆάϊΡΤκΡΘ®C-type natriuretic peptide, CNPΘ©Β»8÷÷άϊΡΤκΡΦΛΥΊ[4]ΓΘANP÷ς“Σ≤ζ…ζ”Ύ–ΡΖΩΘ§BNPΉν≥θ «‘Ύ÷μΡ‘Χα»ΓΈο÷–ΖΔœ÷ΒΡΘ§¥φ‘Ύ”Ύ»ΥΡ‘Θ§ΒΪΗϋΕύ¥φ‘Ύ”Ύ–Ρ “ΓΘΫϋΤΎ―–ΨΩ±μΟςΘ§―Σ«εBNP”κ–¬…ζΕυ≥÷–χ–‘ΖΈΕ·¬ωΗΏ―ΙΦΑάœΡξΖΈ≤ΩΗ–»ΨΒΡ≤Γ«ι≥ΧΕ»ΨΏ”–œύΙΊ–‘[5-6]ΓΘCNP «Φ“ΉεΒΡΒΎ3Ηω≥…‘±Θ§÷ς“ΣΖ÷≤Φ”Ύ÷– ύ…ώΨ≠œΒΆ≥ΓΔ¥ΙΧε«Α“ΕΓΔ…ω‘ύΓΔ―ΣΙήΡΎΤΛœΗΑϊ[7]ΓΘ’β–©κΡ”–3ΗωΡΛ ήΧεΘ§Φ¥άϊΡΤκΡ ήΧεAΘ®natriuretic peptide receptor A, NPRAΘ©ΓΔάϊΡΤκΡ ήΧεBΘ®natriuretic peptide receptor B, NPRBΘ©ΓΔάϊΡΤκΡ ήΧεCΘ®natriuretic peptide receptor C, NPRCΘ©ΓΘNPRΦ“Ήε÷ς“ΣΆ®Ιΐ ήΧεΫαΚœΒςΩΊΜζΧεΗς÷÷¥ζ–ΜΚΆ…ζάμΙΠΡήΓΘNPRAΚΆNPRB¥”Τδ≈δΧεΦΛΥΊ÷–Μώ»Γ–≈œΔ≤ΔΦΛΜνœ¬”Έ–≈Κ≈Ά®¬ΖΓΘNPRC÷ς“Σ «“Μ÷÷«ε≥ΐ ήΧεΘ§÷ς“Σ≤Έ”κ’β–©ΦΛΥΊΒΡ«ε≥ΐΜρΫΒΫβΓΘ±ΨΈΡΨΆNPRΦ“ΉεΦΑΤδ ήΧεΫαΚœ‘ΎΕώ–‘÷ΉΝωΖΔ…ζΓΔΖΔ’Ι÷–ΒΡΉς”ΟΦΑ«±‘ΎΒΡΝΌ¥≤Φέ÷ΒΫχ––Ήέ ωΓΘ
1 NPRΦ“ΉεΒΡΫαΙΙΚΆΙΠΡή
1.1ΓΓNPRA
NPRA“≤±Μ≥ΤΈΣNPR1ΜρGC-AΓΘ»ΥάύNPRAΜυ“ρ≥ΛΕ»‘Φ16 kbΘ§ΈΜ”Ύ1q21ΓΪ22…œΘ§±ύ¬κ1 061ΗωΑ±ΜυΥακΡΘ§Αϋά®21ΗωΆβœ‘Ή”ΚΆ21ΗωΡΎΚ§Ή”[8]ΓΘΗΟΜυ“ρ Ήœ»±Μ3Ηω≤ΜΆ§ΒΡΉι‘Ύ3Ηω≤ΜΆ§ΒΡΡΘ–Ά÷–ΩΥ¬Γ≥ωά¥Θ§»γ–Γ σΓΔ¥σ σΚΆ÷μ[9]ΓΘNPRAΜυ“ρΤτΕ·Ή”÷–Κ§”–SP1ΓΔSP3ΓΔcGMPΖ¥”Π‘ΣΦΰΫαΚœΒΑΑΉΘ®CREBPΘ©ΚΆΈ§…ζΥΊD ήΧεΘ®VDRΘ©Θ§’β–©ΉΣ¬Φ“ρΉ”ΒςΩΊNPRAΒΡ±μ¥οΓΘNPRA «“Μ÷÷135 kDaΒΡΒΞΩγΡΛκΡΆ§‘¥ΕΰΨέΧεΘ§ΑϋΚ§÷Ν…Ό4Ηω≤ΜΆ§ΒΡΫαΙΙ”ρΘ§Αϋά®450ΗωΑ±ΜυΥαΑϊΆβ≈δΧεΫαΚœΫαΙΙ”ρΓΔ21Ηω≤–Μυ ηΥ°ΩγΡΛΫαΙΙ”ρΚΆ566Μρ568ΗωΑ±ΜυΥαΑϊΡΎΫαΙΙ”ρΓΘΚσ’ΏΫχ“Μ≤ΫΜ°Ζ÷ΈΣ1Ηω250ΗωΑ±ΜυΥαΒΡΒΑΑΉΦΛΟΗΆ§‘¥ΫαΙΙ”ρΓΔ1Ηω41ΗωΑ±ΜυΥαΒΡΝΫ–‘¬ί–ΐΫ¬Ν¥«χΚΆ1Ηω250ΗωΑ±ΜυΥαΒΡΡώή’ΥαΜΖΜ·ΟΗcΕΥ¥ΏΜ·ΫαΙΙ”ρΘ§≥ ’ΐΕΰΨέΧεΫαΙΙ[10]ΓΘNPRA «Ρώή’ΥαΜΖΜ·ΟΗœΗΑϊ±μΟφ ήΧεΦ“ΉεΒΡ≥…‘±[11]Θ§NPRA±Μ…ζάμ≈®Ε»ΒΡANPΚΆBNPΥυΦΛΜν[12]ΓΘNPRAΒΡΦΛΜνΥ≥–ρΈΣANPΓίBNP>CNP[13]ΓΘNPRA‘Ύ–Ρ‘ύΓΔ…ω‘ύΓΔΖΈΓΔΗΈ‘ύΓΔ÷§ΖΨœΗΑϊΓΔΗς÷÷ΤςΙΌΦΑ“Μ–©Α©÷Δ÷–ΕΦ”–±μ¥ο[1Θ§13]Θ§ΤδΕ‘–Ρ―ΣΙήΈ»Ε®ΓΔ―Ή÷ΔΦΑΑ©÷ΔΤπΙΊΦϋΒςΫΎΉς”Ο[14-15]Θ§≤Έ”κΝΥΗς÷÷…ζάμΚΆ≤ΓάμΖ¥”ΠΓΘ
1.2ΓΓNPRB
NPRB“≤±Μ≥ΤΈΣNPR2ΜρGC-BΓΘ»ΥάύNPRBΜυ“ρ≥ΛΕ»‘ΦΈΣ16.5 kbΘ§ΈΜ”Ύ9p21ΓΪ12…œΘ§ΑϋΚ§22ΗωΆβœ‘Ή”[16]ΓΘ1989ΡξΘ§SCHULZ”κΥϊΒΡΆ≈Ε” Ή¥ΈΩΥ¬ΓΝΥ¥σ σΒΡNPRBΜυ“ρ[17]ΓΘNPRBΒΡΆΊΤΥΫαΙΙ”κNPRAœύΥΤΘ§Τδ τ”ΎΡώή’ΥαΜΖΜ·ΟΗΒΡœΗΑϊ±μΟφ ήΧεΦ“Ήε[13]Θ§NPRB±ΜCNPΧΊ“λ–‘ΦΛΜν[12]Θ§NPRBΦΛΜνΒΡΥ≥–ρΈΣCNP>ANPΓίBNP[13]ΓΘNPRB‘Ύ»μΙ«ΚΆΙ«–ΈΧ§ΖΔ…ζ÷–ΤπΙΊΦϋΉς”ΟΘ§NPRBΜυ“ρ¥ΩΚœΉ”ΆΜ±δΒΡ»± ßΩ…ΒΦ÷¬ΕΧ÷ΪΌΣ»ε÷ΔΘ§“≤≥ΤΈΣ÷ΪΕΥΖΔ”ΐ≤ΜΝΦ[18]ΓΘΤδ‘ΎΈ§≥÷–Ρ¬ ΚΆώΦΖΩΫαΙΠΡήΖΫΟφ“≤ΤπΒΫ÷Ί“ΣΉς”Ο[19]ΓΘNPRB‘ΎΕύ÷÷ΤςΙΌ÷–±μ¥οΘ§Αϋά®–Ρ‘ύΓΔ¥σΡ‘ΓΔΉ”Ι§ΓΔ¬―≥≤ΓΔ…ωΓΔΖΈΓΔΗΈ‘ύΦΑ÷§ΖΨœΗΑϊΘ§«“‘Ύ“Μ–©Α©÷Δ÷–“≤”–±μ¥ο[17Θ§20-21]ΓΘNPRBΒΡΙψΖΚ±μ¥ο±μΟςΤδ‘Ύ–μΕύ…ζάμΓΔ≤ΓάμΙΐ≥Χ÷–ΖΔΜ”Ής”ΟΓΘ
1.3ΓΓNPRC
NPRC“≤±Μ≥ΤΈΣNPR3ΓΘ»ΥάύNPRCΜυ“ρ≥ΛΕ»‘ΦΈΣ65 kbΘ§ΈΜ”Ύ5p14ΓΪp13…œΘ§ΑϋΚ§8ΗωΆβœ‘Ή”ΚΆ7ΗωΡΎΚ§Ή”[22]ΓΘ–Γ σNPRCΜυ“ρΈΜ”Ύ15Κ≈»Ψ…ΪΧε…œΘ§”…YANAKAΒ» Ή¥Έ‘Ύ–Γ σ÷–ΩΥ¬Γ[23]ΓΘNPRCΒΡΑϊΆβΫαΙΙ”ρ”κNPRAΚΆNPRB‘Φ30%œύΆ§Θ§’β”κΜΖΜ·ΟΗΝ§Ϋ”ΒΡ ήΧε≤ΜΆ§Θ§ΤδΫωΫωΚ§37ΗωΑϊΡΎΑ±ΜυΥαΘ§ΟΜ”–Ρώή’ΥαΜΖΜ·ΟΗΜν–‘[24]ΓΘ”κNPRAΚΆNPRB≤ΜΆ§Θ§NPRC «“Μ÷÷”…ΕΰΝρΦϋΝ§Ϋ”ΒΡΆ§–ΆΕΰΨέΧε[25]ΓΘNPRCΤτΕ·Ή”ΨΏ”–SP1ΓΔAP-2ΓΔYY-1ΓΔE2FΚΆCREBΉΣ¬Φ“ρΉ”ΒΡΫαΚœΈΜΒψ[23]ΓΘMAACKΒ»[26]Χα≥ωΗΟ ήΧε‘Ύ…ζΈο―ß…œ «≥ΝΡ§ΒΡΘ§≤ΔΉςΈΣάϊΡΤκΡΒΡΧΊ“λ–‘«ε≥ΐΫαΚœΈΜΒψΘ§’β÷÷ ήΧε±Μ≥ΤΈΣ«ε≥ΐ ήΧεΓΘΒΪΫϋΡξά¥―–ΨΩ[27]ΖΔœ÷Θ§NPRCΕ‘Έ§≥÷–Ρ―ΣΙήœΒΆ≥Έ»Ε®”–Ή≈ΙΊΦϋΉς”ΟΓΘBNPΚΆCNPΩ…“‘”κNPRCΫαΚœ≤ΔΫχ––«ε≥ΐΚΆΫΒΫβΓΘNPRCΕ‘άϊΡΤκΡΦΛΥΊΒΡ«ΉΚΆΝΠΥ≥–ρΈΣANP>CNP>BNP[13]ΓΘNPRCΒΡ±μ¥ο ήΕύ÷÷…ζάμ“ρΥΊΒΡ”ΑœλΓΘΨί±®ΒάΘ§NPRC‘ΎΗΏ―Σ―ΙΓΔΧ«Ρρ≤ΓΚΆΖ ≈÷ΜΦ’Ώ÷–ΒΡ±μ¥οΖ«≥ΘΗΏΘ§»±ΖΠΧεΝΠΜνΕ·Μα‘ωΦ”NPRCΒΡ±μ¥οΘ§ΕχΕ®ΤΎΒΡΧε”ΐΕΆΝΕΜρΜνΕ·Μα“÷÷ΤNPRCΒΡ±μ¥ο[28]ΓΘNPRC“≤ΙψΖΚ±μ¥ο”Ύ–Ρ‘ύΓΔΖΈΓΔ…ω…œœΌΓΔ¥σΡ‘ΤΛ÷ ΓΔ–ΓΡ‘ΓΔΗΈ‘ύΓΔ÷§ΖΨœΗΑϊΦΑ“Μ–©Α©÷Δ÷–[28]ΓΘ
NPRAΚΆNPRB τ”ΎΡώή’ΥαΜΖΜ·ΟΗ ήΧεΘ§ΕχNPRC τ”Ύ«ε≥ΐ ήΧεΘ§άϊΡΤκΡΆ®Ιΐ”κ3÷÷ ήΧεNPRAΓΔNPRBΦΑNPRCΒΡΑϊΆβΫαΙΙ”ρΫαΚœΕχΤπΉς”ΟΘ§ΤδΉς”ΟΜζ÷Τ“≤”–Υυ≤ΜΆ§Θ®ΦϊΆΦ1Θ©ΓΘ
2 NPRΦ“Ήε‘ΎΕώ–‘÷ΉΝω÷–ΒΡΉς”Ο
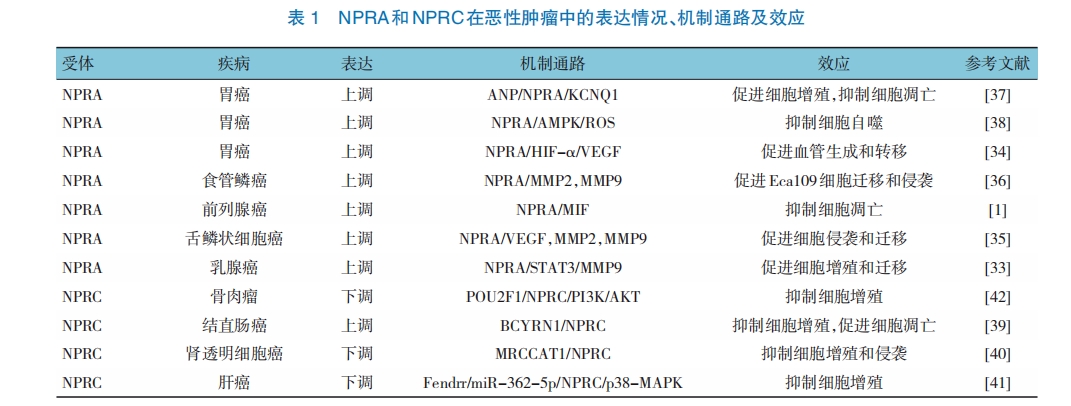
NPRAΚΆNPRC‘ΎΕώ–‘÷ΉΝω÷–ΒΡΉς”Ο“―Ψ≠”–ΚήΕύΒΡ±®ΒάΘ§ΒΪΙΊ”ΎNPRBΒΡ±®ΒάΫœ…ΌΓΘ±ΨΈΡ÷ς“ΣΧ÷¬έNPRAΚΆNPRC‘ΎΑ©÷ΔΖΔ…ζΖΔ’Ι÷–ΒΡΉς”ΟΘ®Φϊ±μ1Θ©ΓΘ
2.1ΓΓNPRA
NPRA‘ΎΑ©÷Δ÷–ΒΡ―–ΨΩΩ…“‘ΉΖΥίΒΫ1993ΡξΓΘOHSAKIΒ»[29] Ή¥Έ‘Ύ»Υ–ΓœΗΑϊΖΈΑ©œΗΑϊœΒΚΆHeLaœΗΑϊ÷–ΖΔœ÷ΝΥΙΠΡή–‘NPRAΓΘVESELYΒ»[30]Ϋχ“Μ≤Ϋ―–ΨΩΝΥάϊΡΤκΡ ήΧε”κΑ©œΗΑϊ‘ω÷≥ΒΡΙΊœΒΓΘ―–ΨΩ±μΟςΘ§NPRA‘ΎΕύ÷÷»ΥάύΑ©œΗΑϊ÷–±μ¥οΘ§Τδ÷–Αϋά®»ιœΌΑ©œΗΑϊ[30]ΓΔ«ΑΝ–œΌΑ©œΗΑϊΒ»[1]ΓΘNPRA”…”ΎΤδΙψΖΚΒΡ…ζάμΙΠΡήΘ§‘Ύ–μΕύ÷ΉΝωΒΡΖΔ…ζΓΔΖΔ’Ι÷–ΤπΉ≈÷Ί“ΣΒΡΉς”ΟΓΘKONGΒ»[31] Ήœ»―–ΨΩΝΥNPRA‘Ύ÷ΉΝωΖΔ…ζ÷–ΒΡ÷±Ϋ”Ής”ΟΓΘΕ·ΈοΡΘ–ΆΫαΙϊœ‘ ΨΘ§»±ΖΠNPRAΩ…“‘±ΘΜΛC57BL/6–Γ σΟβ ήΖΈΑ©ΓΔΤΛΖτΑ©ΚΆ¬―≥≤Α©ΒΡ”ΑœλΓΘ‘ΎΦ≤≤ΓΡΘ–Ά÷–Θ§œ¬ΒςNPRAΩ…“‘“÷÷Τ÷ΉΝωΒΡ…ζ≥ΛΚΆΉΣ“ΤΘ§Χα ΨNPRAΩ…Ρή≥…ΈΣ“Μ÷÷–¬ΒΡΩΙΑ©÷ΈΝΤΑ–ΒψΘΜΤδ―–ΨΩΜΙ±μΟςΘ§NPRA‘Ύ÷ΉΝωœΗΑϊΘ®A549ΓΔB16Β»Θ©÷–Ιΐ±μ¥οΓΘNPRAΩ…“‘Ά®ΙΐΒςΫΎΗ…œΗΑϊΒΡΡΦΦ·ΚΆ―ΣΙή…ζ≥…ά¥ΒΦ÷¬÷ΉΝωΒΡΫχ’Ι[32]ΓΘ‘Ύ»ιœΌΑ©[33]ΓΔΈΗΑ©[34]ΓΔ…ύΝέΉ¥œΗΑϊΑ©[35]ΚΆ ≥ΙήΑ©[36]÷–Θ§NPRAΩ…Ά®Ιΐ≤ΜΆ§ΒΡΆΨΨΕΘ®MMP2ΓΔMMP9ΓΔHIF-1ΠΝΒ»Θ©ά¥ΧαΗΏVEGFΥ°ΤΫΘ§¥ΌΫχ―ΣΙή…ζ≥…Θ§¥”Εχ¥ΌΫχ÷ΉΝωΒΡ«÷œ°ΚΆΉΣ“ΤΓΘ‘Ύ«ΑΝ–œΌΑ©[1]ΚΆΈΗΑ©[37]Θ§NPRAΩ…Ά®Ιΐ“÷÷Τ÷ΉΝωœΗΑϊΒρΆωά¥¥ΌΫχ÷ΉΝωΫχ’ΙΓΘ”–―–ΨΩ±μΟςΘ§‘ΎΈΗΑ©÷–[38]Θ§“÷÷ΤNPRAΒΡ±μ¥οΩ…“‘Ά®ΙΐJNKΚΆp38Ά®¬ΖΘ§¥ΌΫχΈΗΑ©œΗΑϊ÷ήΤΎΉη÷ΆΓΔœΗΑϊΥάΆωΚΆΒρΆωΓΘ
2.2ΓΓNPRC
NPRC‘ΎΑ©÷Δ÷–ΒΡ―–ΨΩΫϋΡξά¥≤≈±ΜΧαΦΑΘ§ΒΪNPRCΉςΈΣ“ΜΗω–¬–ΥΒΡΑ–ΒψΘ§‘ΎΑ©÷ΔΖΔ…ζΓΔΖΔ’ΙΒΡΜζ÷Τ―–ΨΩ÷–ΨΏ”–Ψό¥σΒΡ«±ΝΠΓΘNPRCΉςΈΣ“Μ÷÷«ε≥ΐ ήΧεΘ§“ΜΑψΤπΉ≈“÷Α©ΒΡΉς”ΟΓΘ‘ΎΫα÷±≥ΠΑ©[39]Θ§…ωΆΗΟςœΗΑϊΑ©[40]ΚΆΗΈΑ©[41]Θ§NPRCΩ…±Μ…œ”Έ≥ΛΝ¥Ζ«±ύ¬κRNAΒςΩΊΓΘΙΐ±μ¥οNPRCΩ…“÷÷ΤœΗΑϊ‘ω÷≥Θ§¥ΌΫχœΗΑϊΒρΆωΘ§¥”ΕχΖΔΜ”ΩΙΑ©Ής”ΟΓΘ‘ΎΙ«»βΝω÷–[42]Θ§NPRCΩ…Ά®ΙΐΉηΕœPI3K/AktΆ®¬Ζά¥“÷÷ΤΙ«»βΝωœΗΑϊΒΡ…ζ≥ΛΓΘ
3 NPRΦ“Ήε‘ΎΕώ–‘÷ΉΝω’οΝΤ÷–ΒΡΝΌ¥≤Φέ÷Β
3.1ΓΓNPRΦ“Ήε‘ΎΕώ–‘÷ΉΝω’οΕœ÷–ΒΡΦέ÷Β
NPRΦ“Ήε‘Ύ–μΕύΕώ–‘÷ΉΝωΉι÷·÷–±μ¥οΘ§”κ÷ΉΝωΒΡ’οΕœΓΔΖ÷ΤΎΦΑ‘ΛΚσΟή«–œύΙΊΓΘ‘Ύ ≥ΙήΝέΉ¥œΗΑϊΑ©ΚΆΈΗΑ©÷–Θ§NPRAΒΡ±μ¥ο”κΝήΑΆΫαΉΣ“ΤΚΆΘ®TNMΘ©Ζ÷ΤΎœύΙΊ[36Θ§38]Θ§Ε‘‘Λ≤β÷ΉΝωΖΔ’Ι”–÷Ί“ΣΉς”ΟΘΜNPRA”κ…ύΝέΉ¥œΗΑϊΑ©÷–VEGFΒΡ±μ¥οΦΑ÷ΉΝωœΗΑϊΒΡ«÷œ°ΚΆΉΣ“Τ”–ΙΊ[35]Θ§’βΩ…Ρή «“ΜΗω‘ΛΚσ“ρΥΊΘΜNPRAΒΡ±μ¥ο“≤”κ«ΑΝ–œΌΑ©ΒΡΝΌ¥≤Ζ÷ΤΎ≥ ’ΐœύΙΊ[1]ΓΘNPRBΜυ“ρ‘Ύ«ΑΝ–œΌΑ©÷–ΒΡ±μ¥ο”κ÷ΉΝωΧεΜΐΚΆΕώ–‘≥ΧΕ»”–ΙΊΘ§NPRBΩ…≥…ΈΣΦχ±πΝΦΓΔΕώ–‘Ήι÷·ΒΡ“Μ÷÷ ÷ΕΈ[43]ΓΘ‘Ύ«ΑΝ–œΌΑ©ΡΘ–Ά÷–Θ§NPRCΩ…≥…ΈΣ«ΑΝ–œΌΑ©‘γΤΎ’οΕœΒΡΑ–Βψ[44]ΓΘ
3.2ΓΓNPRA‘ΎΕώ–‘÷ΉΝω÷ΈΝΤ÷–ΒΡΦέ÷Β
NPRA“―±Μ±®ΒάΈΣΑ©÷Δ¥ΌΫχΑ–ΒψΓΘ–μΕύ Β―ι±μΟςΘ§œ¬ΒςNPRAΒΡ±μ¥οΩ…“‘Ά®Ιΐ≤ΜΆ§ΒΡΆΨΨΕ”’ΒΦ÷ΉΝωœΗΑϊΒρΆωΘ§“÷÷ΤœΗΑϊ‘ω÷≥ΚΆ«÷œ°[33Θ§35-36]ΓΘΤδΜΙΩ…“‘Ά®Ιΐœ¬ΒςVEGFΓΔCXCΚΆCXCR4“÷÷Τ―ΣΙή…ζ≥…Θ§¥”Εχ“÷÷Τ÷ΉΝωœΗΑϊΒΡ«®“Τ[32]ΓΘ
Isatin «“Μ÷÷Χλ»ΜΒΡ‘”ΜΖή’―ή…ζΈοΘ§Ω…œ¬ΒςNPRAΒΡ±μ¥οΘ§“÷÷ΤΤχΒά―Ή÷Δ[45]ΓΘIsatinΕ‘Α©÷ΔΒΡ÷ΈΝΤΚή”–«ΑΨΑΘ§“λΙΙΥΊΤ’±ι¥φ‘Ύ”Ύ»ΥΧε÷–Θ§Τδ≤ΜΫω «Ρώή’ΥαΜΖΜ·ΟΗΒΡ“÷÷ΤΦΝΘ§“≤ «–μΕύΤδΥϊΟΗΚΆ ήΧεΒΡ“÷÷ΤΦΝΘ§άΐ»γΉιΒΑΑΉ»Ξ““θΘΜ·ΟΗΓΔΧΦΥατϊΟΗ[46]ΓΘ“λΥΊ‘”ΫΜΦΝ‘Ύ–μΕύΕώ–‘÷ΉΝω÷–ΖΔΜ”ΩΙΑ©Ής”ΟΘ§ΒΪΤδΡώή’ΥαΜΖΜ·ΟΗ“÷÷ΤΦΝΆΨΨΕΒΡΩΙΑ©Μζ÷Τ…–Έ¥≤ϊΟς[47]ΓΘ¥ΥΆβΘ§Isatin hybridsΨΏ”–ΝΦΚΟΒΡΩΙΑ©Μν–‘Θ§άΐ»γΘ§SemoxindΚΆ φΡαΧφΡαΩ…”Ο”Ύ÷ΈΝΤ–μΕύΑ©÷Δ[47-48]ΓΘ
Ζ÷Ή””ΓΦΘΨέΚœΈοΡ…ΟΉΩ≈ΝΘΘ®MIPNPSΘ©Ε‘ANPΨΏ”–«ΉΚΆΝΠΚΆ―Γ‘ώ–‘Θ§Ω…ΫΒΒΆΑ©÷Δ÷–ANPΒΡΥ°ΤΫΘ§“÷÷ΤANP/NPRA–≈Κ≈Ά®¬Ζ[49]ΓΘ±ύ¬κNP73-102ΒΡ÷ ΝΘ[«Α–ΡΖΩάϊΡΤκΡΒΡNHΘ®2Θ©Ρ©ΕΥκΡ]ΒΡ“λΈΜ±μ¥οœ¬ΒςΝΥNPRAΒΡ±μ¥οΘ§“÷÷ΤΝΥA549œΗΑϊΒΡΖΈΉΣ“ΤΚΆBALB/c–Γ σΒΡ÷ΉΝωΖΔ…ζΘ§Ιΐ±μ¥οNP73-102Ω…œ‘÷χ”’ΒΦΚΎ…ΪΥΊΝωœΗΑϊΘ®B16Θ©ΒΡΒρΆω[31]ΓΘ―ΣΙήΫτ’≈ΥΊΔρΩ…“÷÷ΤNPR1Μυ“ρΒΡΉΣ¬ΦΘ§œ¬Βς–Ρ‘ύ÷–NPRAΒΡ±μ¥ο[50]ΓΘ«ΉΜΖΥΊA «Οβ“Ώ“÷÷ΤΦΝΜΖφΏΥΊAΒΡ ήΧεΘ§Ω…”κNPRAΫαΚœΘ§¥ΏΜ·Pro 822ΓΔ902Μρ958ΒΡΥ≥Ζ¥“λΙΙΜ·Θ§¥”Εχ ΙNPRA ßΜνΘ§“―≥…ΈΣΑ©÷Δ÷ΈΝΤΒΡΑ–Βψ[51]ΓΘΈΔ…ζΈοΕύΧ«HS-142-1 «“Μ÷÷άϊΡΤκΡόΉΩΙΦΝΘ§Ω…“÷÷ΤNPRAΚΆNPRBΒΡΦΛΜνΚΆΫαΚœΘ§ΒΪΕ‘NPRCΈό”Αœλ[52]ΓΘ
3.3ΓΓNPRC‘ΎΕώ–‘÷ΉΝω÷ΈΝΤ÷–ΒΡΦέ÷Β
‘Ύ…ώΨ≠ΡΗœΗΑϊΝωœΗΑϊœΒ÷–Θ§NPRCάύΥΤΈοΖΔΜ”ΩΙ‘ω÷≥Ής”ΟΘ§ΕχNPRAΚΆNPRB¥ΌΫχ…ώΨ≠ΡΗœΗΑϊΝωΒΡ‘ω÷≥[53]ΓΘ“ΜΑψά¥ΥΒΘ§NPRAΚΆNPRBΨΏ”–¥ΌΑ©Ής”ΟΘ§ΕχNPRCΨΏ”–“÷Α©Ής”ΟΓΘNPRCΉςΈΣ“Μ÷÷«ε≥ΐ ήΧεΘ§Ω…”κANPΓΔBNPΚΆCNPΫαΚœΘ§≤ΔΫχ––«ε≥ΐΚΆΫΒΫβΓΘNPRC «ΖώΩ…“‘“÷÷ΤNPRA–≈Κ≈Ά®¬ΖΜΙΈ¥”–―–ΨΩ÷ΛΟςΘ§»γΙϊ¥φ‘Ύ¥σΝΩΒΡNPRCΘ§Τδ «ΖώΩ…“‘“÷÷ΤάϊΡΤκΡΦΛΥΊ”κNPRAΚΆNPRBΒΡΫαΚœΘ§NPRCΉςΈΣ’β–©–≈Κ≈Ά®¬ΖΒΡΤπΒψΘ§ΤδΉςΈΣ“÷Α©Α–ΒψΒΡΉς”ΟΜζ÷ΤΜΙ–η“ΣΗϋΕύΒΡ―–ΨΩ»Ξ÷ΛΟςΓΘ
4 Ϋα¬έ”κ’ΙΆϊ
±ΨΈΡΨΆNPRΦ“Ήε‘Ύ–μΕύΕώ–‘÷ΉΝωΒΡΖΔ…ζΓΔΖΔ’Ι÷–ΤπΉ≈÷Ί“ΣΉς”ΟΫχ––Ήέ ωΓΘ–μΕύΕ·ΈοΡΘ Β―ι“―Ψ≠÷ΛΟςΘ§NPRAΦ“Ήε‘Ύ–μΕύΕώ–‘÷ΉΝωΒΡΖΔ…ζΓΔΖΔ’Ι÷–Τπ÷Ί“ΣΉς”ΟΓΘ»ΜΕχΘ§”…”Ύ»±ΖΠΚœ ΒΡNPRAΩΙΧεΘ§NPRA‘Ύ»ΥΧεΉι÷·÷–±μ¥οΒΡ―–ΨΩ”–œόΓΘΡΩ«ΑΘ§NPRΦ“Ήε÷ς“Σ”–3Ηω”Π”ΟΝλ”ρΓΘΔΌNPRA‘ΎΕύ÷÷Α©÷Δ÷–ΒΡ¥ΌΑ©Ής”ΟΘ§Ω… ΙΤδ“÷÷ΤΉς”ΟΒΡ―–ΨΩ”»ΈΣ÷Ί“ΣΓΘΔΎNPRΦ“Ήε‘Ύ÷ΉΝω÷–ΒΡ±μ¥ο”κ÷ΉΝωΒΡΧεΜΐΓΔΖ÷ΤΎΓΔΉΣ“ΤΚΆΖ÷Μ·Οή«–œύΙΊΘ§Ω…ΉςΈΣ’οΕœΚΆ‘ΛΚσΒΡ≤ΈΩΦ÷Η±ξΓΘΔέNPRCΉςΈΣ“Μ÷÷«ε≥ΐ ήΧεΘ§Ά®≥ΘΖΔΜ”ΩΙΑ©Ής”ΟΘ§ΤδΩ…“‘”κNPRAΨΚ’υ≈δΧεΘ§“÷÷ΤΤδ–≈Κ≈Ά®¬ΖΓΘNPRΦ“ΉεΉςΈΣ“Μ÷÷ΩΙΑ©Α–ΒψΘ§…–Έ¥”Π”Ο”ΎΝΌ¥≤Θ§ΡΩ«Α»‘¥Π”Ύ―–ΨΩΫΉΕΈΓΘ»ΜΕχΘ§”…”ΎNPRΦ“Ήε‘Ύ»ΥΧε÷–ΒΡΙψΖΚΉς”ΟΘ§Ε‘Τδ…ν»κ―–ΨΩ”»ΈΣ÷Ί“ΣΘ§≤ΜΫωΈΣΑ©÷ΔΒΡ’οΕœΚΆ÷ΈΝΤΧαΙ©–¬ΒΡΥΦ¬ΖΘ§Εχ«“‘ΎάϊΡΤκΡΦΛΥΊΒΡ―–ΨΩ÷–“≤ «÷Ί¥σΆΜΤΤΓΘ
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷ΨΓΘ
