ժҪ������ͨ������������Ȼ����Ӧ��ɱ����֢ϸ��������, ��������������Ϊ��֢���Ƶ��·���. �봫ͳ���Ʒ������, ���������ܹ��յ����ڵ���������Ԥ����֢������ת��, ���и����Ŀ���Ч���Լ���С�ĸ�����. Ȼ���������и����ﰲȫ�Ժ�����Ӧ�������IJ�����Ȼ������ս��. ����������Ȼ���������ƵĽṹ����, ���ķ�������ͨ��ֱ����������Ӧ������ҩ�����Ч���������Щ��ս. �����ܽ��˹�ȥʮ���ڷ��ֵ����ö��ķ�����Ϊ��������ҩ������ƽ̨�İ�֢�����Ʒ�. �Ӽ�Ҫ���ܰ�֢�������ƿ�ʼ, ���Ǹ����˶��ķ��ӵ��ض�ҩ�﹦��, �������������Ƽ������翹ԭ������. ������ؽ����˻��ڶ������ṹ��Ϊ����ƽ̨, ����ҩ�������ͻ����ҩ�ﹲͬ��������ǿ����ԭ�ԵĽ�չ. ���Ի��ڶ��ĵİ�֢�����������ٵ���ս�Լ�δ���ķ�չ���ƽ�����չ��.
��֢����в��������Ҫ����֮һ�����������г������ƺͷ������Ƶȴ�ͳ���Ʋ���ȡ���˽�չ�����������������ų�����Ƶ���ս[1��2]����������Ϊ����Ʒ�����������Ч������ҩ���Ѱи�����С�����ƣ��ڹ�ȥ��ʮ���ܵ��㷺��ע��[3��4]����ֱ�ӹ�����ϸ����ͬ�������Ʒ�������������Ȼ���߷�Ӧ���Ӷ�ɱ����ϸ�����Ե�һ�����ëϸ����Ѫ���������Ʒ��������������ְ�֢�����ѱ�������Ӧ���ڰ�֢��������[5]�����������ڼ������߷�Ӧ���²��ԣ���֢��������ȡ����ͻ���Խ�չͨ����Բ�ͬ������ϸ��[6]ԭ���ϣ�����ϵͳ����������ϵͳ����Ӧ������ϵͳ��ɣ�ǰ�������ڲ��������ϸ����������������ϵͳ�е���ϸ��������ϸ����������ϸ������ͻ״ϸ����������������Ӧ������ϵͳ�е�T��B�ܰ�ϸ����������������ƣ���Բ�ͬ���߷�Ӧ������ҩ��ɷ�Ϊ��֢���硢����������ϸ�����ӡ�������ϼ����̻�Tϸ���Լ������������Ŀǰ�����������������[7��8]�Լ����ù��� T ϸ�����й�����������[9]���ٴ�������ʹ�õ�������ϣ���IJ��ԡ�������ϸ������ 1 (PD-1) �������� PD-L1 ��ϸ������ T �ܰ�ϸ����ԭ 4 (CTLA-4) �ļ����ǰ��������С����ҩ�����ڵ���ϼ�����������е㡣����ת�ƹ��̻� T ϸ������Ƕ�Ͽ�ԭ���� T (CAR-T) ϸ������ȡ����Ȼ T ϸ������Ҳ����ֱ���������߷�Ӧ[10�C13]�����ܾ��о������DZ��������������ԭ�Եͺ����صIJ��������ã���ϸ�������ͷ��ۺ����������������ٴ�����ʧ�ܣ��������谭�����һ����չ����ˣ���Ҫ��Ч�������߷�Ӧ�������Ƽ��ͼ��ٲ�����Ӧ�����������߸����ã������͵���֧�ܡ�
����������Ȼ���������Ƶİ�������ɵģ��ѹ㷺Ӧ������֯������ҩ�����͵�ҩ���������ϵĿ���[14�C16]. ���������о���Դ����Ȼ�����ʻ�ģ�µ����ʵ���ṹ������DZ����������˿��ܱ��ֳ���������ԭ���ϸ����ϵ��������������Ʒ������磬�ܿ�����������������߷�Ӧ������������ͨ���������������������������������������Է���ϸ�������[17��18]�������ӵ�������ȣ������Ʒ��ڸ�ҩ���������������ֳ��������������ơ����̵����������ںϳɣ�ҲDZ�ڵ�����������ʵ��������֯�С����ĵĽṹҲ���Խ��о�ȷ�������Է�ֹ��ҩ����Ⱦ��������κι������������߷�Ӧ�����ĵ�����Ч������Ҫ�ȶ������幹��������ĵ����ʿ��壬����ڿ������Ч������Ҫ�������µ����ʿ����ҩ����ս�����⣬�������Ʒ��ķǹ�������ÿ��Դٽ�������װ������ҩ�������ҩ��ͨ�����ֳ��ɿص�ҩ������ѧ����һ���棬��ۺ���ϵͳ��ȣ����γɵ����ṹ����Ϊ����ƽ̨ʱ�����ض���������Ȼ�ĵ���Ȼ�ɷ�ʹ����г�ɫ��ϸ�������ԣ����ĵĺ�����ƿ��Ծ�ȷ�������ǵĽ�ϣ��Ӷ��������пɿ���̬�����ṹ��������ڼ�����Ϻ����ƵĹ�����������ϴӶ��������пɿ���ò�����ṹ����������ڼ�����Ϻ����ƵĹ�����������ϴӶ��������пɿ���ò�����ṹ����������ڼ�����Ϻ����ƵĹ������������[19��20]. �����������з�Ӧ������װ�壬�ر��Ƕ������־���з�Ӧ������װ�壬�Ǹ�Ч�������������֧��[21�C23]������Եؽ���[24��25]�����㷺Ӧ���ڴ�ͳ��֢����[26�C28]. �����Щ���ƺ�DZ�ڵĿ�Ĥ���������ڰ�֢���������о��о�DZ��������Ϊ���Ƽ������֧��[29�C31]��
����Ϊֹ���ϳɶ��ļ�����װ�������ڲ�ͬ���͵İ�֢�������ƣ�ͼ 1��[32��33]. ������Ȼ�������ڽṹ����ı�λ�����ܹ����������Ӧ������ϸ����ϸ���д��ڵ���������ã��Ӷ������������������ϵͳ�����ƹ���[34��35]�����⣬���ӽ������������ٴ������Բ������ƻ�����ϵͳ��������ģ��Ӷ�Ϊ�����������Ʒ��ṩ����һ�ַ���������Ϊֹ�������൱�����Ķ�����Ϊ������ϼ�[36��37]����֢����[38��39]������[40]�Ѿ�������������һ���棬����װ���ѱ��㷺�����������壬���ڽ��������Ƽ�����ϸ���Ϳ���ȴ����嵽Сҩ�������͵�������λ[41��42]��������װ�����̬ת�䣬���������廹������������ҩ���ѭ��ʱ��[43�C45]. ���������װ���ڴ�ͳ���ƺ��������Ʒ����Ľ�����������ر�Ĺ�ע[46�C50]����Ϊ���Ǿ��й�ͬװ�ض��ֻ���Ķ���ԡ��������ڰ�֢���������о��о�DZ�����������ĵIJ��ԵĽ�չ��δ�ܽᡣ
���������������ƹ����е����÷����ܽ��˻����ĵİ�֢�������Ʋ��ԡ��������ȸ�������������Ϊ�����Ʒ��Ļ����ĵ������Ʒ���������������ṹ��Ϊ���Ƽ�����֧�ܵ�Ӧ�á������ر�ǿ����Эͬ�����Ʒ����漰���滯�ƺ���Ʒ�����������Ʒ���ֵ��ע����ǣ����������������ö�����Ϊ���Ƽ������֧�ܵİ�֢�����Ʒ��������漰�漰�����ʺͶ��ĵȳ����еIJ��ԡ����⣬���������ij������м�ʮ�꣬����Ϊֹ�ѿ������������ڲ�ͬ���Ʒ�ʽ�Ͱ�֢�������ġ�Ϊ��ȷ˵�������ǵIJο����ף������ܽ�����������Ĵ���ǹ�ȥʮ���ڷ��ֵģ������ĵ���ƽ̨�����Ϊ���˵��������Ʒ�ʽ���������Ϻ���������Ʒ��������ġ������ĵĸ������ԣ��ܽ�����ĵİ�֢�������Ʋ��Կ��Ը����������������е�Ӧ����״�������ܴ̼������²����������Ч���Ӷ����ܴٽ�δ����֢�������Ƶ��ٴ�ת����
�ļ�����ϼ�
��������ָ����ϵͳ�д��ڵĸ����ڼ�������ά������ƽ�Ⲣ��ֹ�������߲�������ع���ϸ��[51]Ȼ�������������Ҳ�����ڰ�ϸ���б���������������Ŀ��������߷�Ӧ���Ӷ����°�ϸ�����ݺ�����[52��53]���������������ֹ��ϸ���������ӱܣ����̼�ϸ������Tϸ��������ϸ���Ļ�����������ϸ�����ⱻ��Ϊ��һ����ǰ;����Ч�İ�֢�������Ʋ���[54]ԭ���ϣ��Tϸ���������Ҫ�������������PD-1[55], ����ϸ����ѧ��ԭ-4[56], �ܰ�ϸ������� 3�ͺ�-3 [57�C59]��Tϸ��������3��TIM-3��[60�C62]�Լ� T ϸ�������� ITIM �ṹ�� (TIGIT)[63�C65]��ˣ���Щ������Ϊ���������㣬���Կ������������߷�Ӧ�����Ƽ����統ǰ�ٴ�ǰ�о����ٴ������й㷺ʹ�õ� PD-1 �� CTLA-4��ͼ 2��[63]��
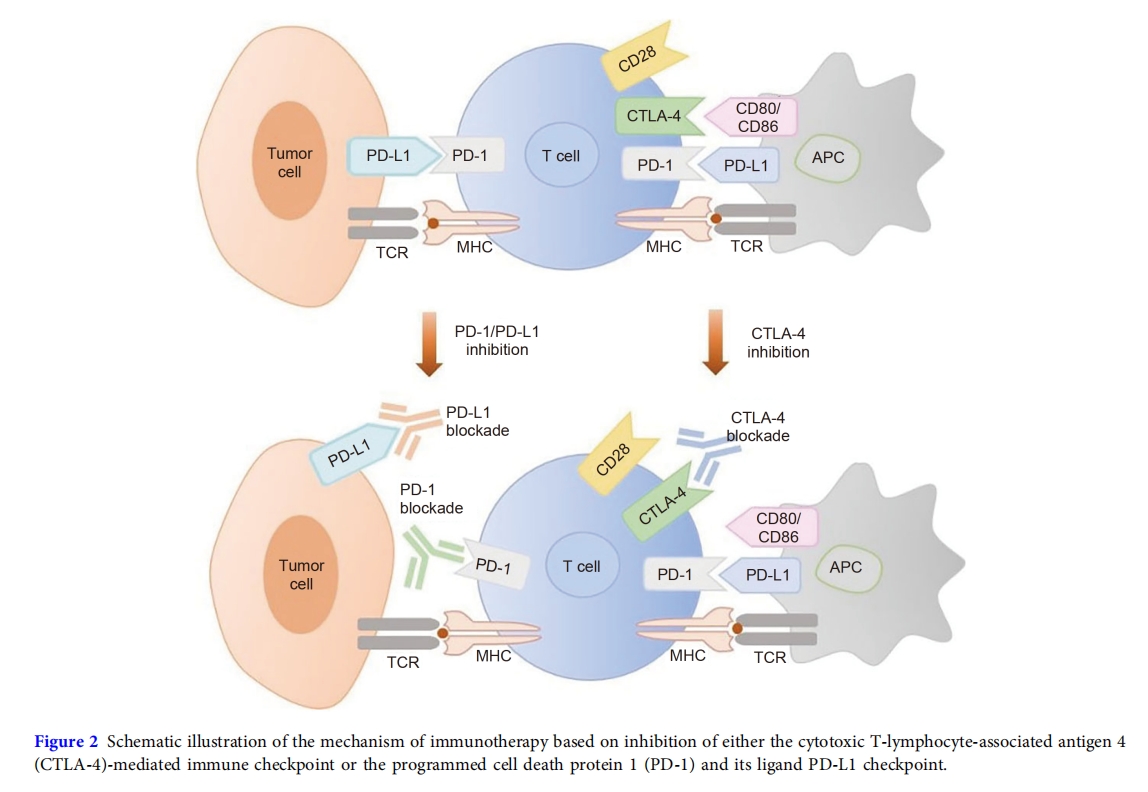
CTLA-4 ��һ�ֿ�Ĥ�ǵ��ף��ڵ����Ի���Tϸ���ϸ߶ȱ���[66]CTLA-4��Tϸ������Ĺ��̼��������壨CD28�����и߶�ͬԴ�ԣ��ܹ���B7����ϣ���CD80��B7-1����CD86��B7-2��������������CD28�߳�Լ20����ͼ 2��ʹ�� CTLA-4 �ܹ��� CD28 ���� B7 ��ϣ��Ӷ���ֹ CD28-B7 ���̼��źŵ��ͷŲ����� T ϸ���[67�C69]��Ŀǰ���о�������CTLA-4 ����ͨ���źŻ���ź�ͨ·���� T ϸ�����߷�Ӧ���ź�ͨ·���� CTLA-4 ��������ø��ʹ T ϸ������ (TCR) ���ź�ȥ���ữ�����ź�ͨ·���� CTLA-4 ����ͨ��ת���̹��̴ӿ�ԭ�ʵ�ϸ�� (APC) ��Ĥ�ϲ���ȥ�� CD80 �� CD86 ���ף��Ӷ����� CD28 ����[70��71]PD-1 ����һ��������չ B7/CD28 ����������㣬�ڻ�� T ϸ����B ϸ������Ȼɱ��ϸ������ͻ״ϸ����������ؾ���ϸ���и߶ȱ���[72]PD-1 �����ɰ���ṹ��Ĥ�ṹ��Ͱ��ڽṹ����ɡ���Ӱ������ T ϸ���� CTLA-4 ��ͬ��PD-1 ͨ�������ڳ��� T ϸ���ϣ����������������ڵ�ЧӦ T ϸ�����ԣ�ͼ 2��[73]. PD-1 ��������� PD-L1��B7-H1 �� CD 274���� PD-L2��B7-DC �� CD 273������ APC ������ϸ������[74��75]PD-1���������ϵ���Tϸ����øʧ���TCR�ź�ȥ���ữ���Ӷ����ռ�������ϸ�����ӵIJ���������Tϸ������[76��77]��ˣ����PD-1/PD-L1ͨ·����ά������������Tϸ���Ļ��ԣ�ʹ����ϵͳ����ʶ���������ϸ�����Ӷ���ֹ����ϸ���������ӱܡ�
��ĿǰΪֹ����Ȼ������CTLA-4�������Ƽ��ǿ��壬�� PD-1 �� PD-L1 ����ȷ���м��临�ӽṹ�������Ǻ�������ܹ���Խ PD-1 �� PD-L1 �����Ķ̷����ģ���1)���ܵ���¡������ΪPD-1/PD-L1�������Ƽ����ٴ�������ȡ����һ����չ���������ʵ������֯���Բ�ȶ��Բ�����ɱ��ߡ���ҩ;�����ޡ�ҩ������ѧ�ɿ��Խϲ�����������谭�����ٴ�ת��[78��79]�����⣬�������Ƽ�ͨ����Ȼɱ��ϸ���;���ϸ������ϸ���������߷�Ӧ���������ޡ���Ϊ PD-1/PD-L1 ������ϼ����Ļ�����С�л������ǽ������¡������Щ��������ҩ����ǵ����ĵķǷ����������Ժ����ںϳɣ������ĵ� PD-1/PD-L1 ���Ƽ��ڰ�֢���������������˹㷺��ע��������Ҫ������ PD-1/PD-L1 ���Ƽ��Ŀ��������ܽ����ٴ�������ٴ�ǰ�����еĿ������С�
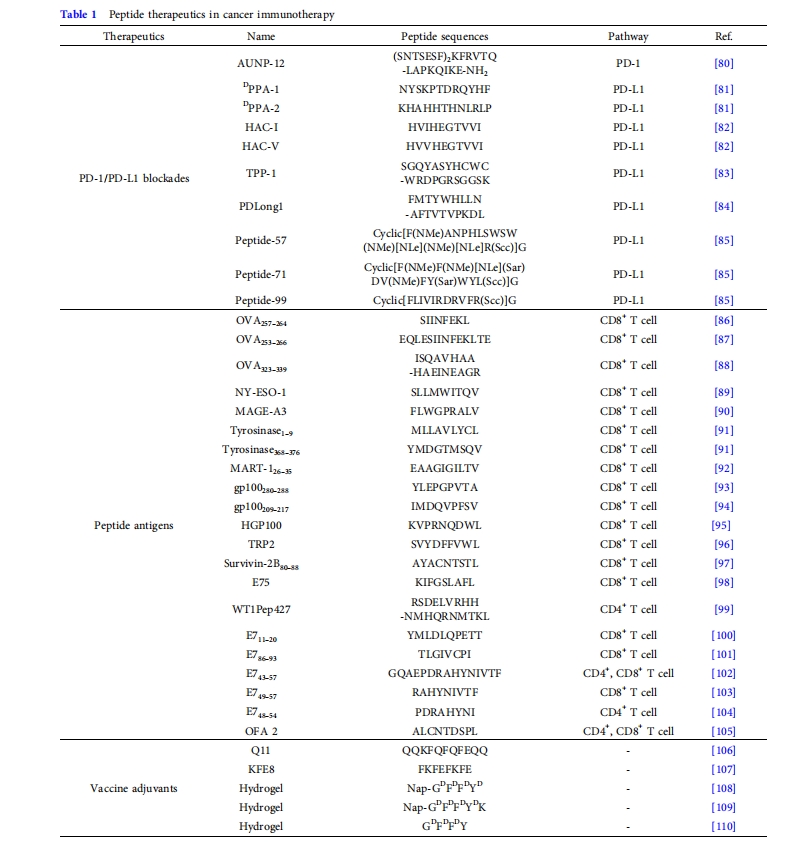
AUNP-12��������PD-1/PD-L1���Ƽ�֮һ���� Aurigene Discovery Technologies �� Pierre Fabre ʵ���ҷ��֣��뿹�����Ƽ���Ⱦ���ҩ������ѧ���ƣ������������� PD-1 �������� 7 �� 30 ������ķ��������[80]���ٴ�ǰ�о��У�AUNP-12���ֳ����������ƶ�����������������������B16F10С���ɫ����ϸ����С�����ٰ�4T1ϸ���������о��������鵼�����ئã�IFN-�ã��IJ�����AUNP-12���ƻ��Ե�DZ���ź�ͨ·����AUNP-12��Ч��ϵ���о�������ɾ��Cĩ��8���л�������Nĩ��˿����л����ᵼ�����ƻ��Ե�ɥʧ����ȥ����֧�ṹ�������Cĩ��������л������˻��ԡ���������AUNP-12�ĵĶ���ģ�����AUNP-12��ȣ����ڰ�֢���������б��ֳ����õĻ��ԡ��ڴ��о�֮��Aurigene���ڴ˸������һϵ����ģ����PD-1/PD-L1���Ƽ���
���⣬Chang���ˡ�[81]���ھ����ɾ���չʾ��������������������ˮ���D��PD-1/PD-L1 ��������� ( DPPA-1��NYSKPTDRQYHF)�������ɾ���չʾ��������ʹ�û�ѧ�ϳɵ� D ���ն�ɸѡ��L�б��ϵ�D ��¡�������������ɱ� (Ig-V) �ṹ��ʼ������������۵�IgV�ṹ��� D�汾 ( D IgV PD-L1 9)�����������ɾ���չʾ�е��ն��ġ�ͨ��ɸѡ M13 �ɾ�����չʾ��ʮ�������Ŀ⣬����ѡ��������D���� ( DPPA 1: NYSKPTDRQYHF��DPPA 2: KHAHHTHNLRLP) ��Ƶ����ߡ�����������干��������˽�ϳ��� (��D PPA 1 ��D PPA 2 ���� PD-1�ı���Ϊ 0.51������1.13��mol��L −1����ʽϸ����ʵ���������D PPA 2��ȣ� D PPA 1 ���ֳ������������ PD-1/PD-L1 ����õ�����������ʵ������� D PPA 1��ҩ������36ֻBalb/cС������ֲ���CT26ϸ��������������������ڼ����˿���������ϵͳ���¡���ˣ���ˮ��D�ľ�����Ϊ��֢��������С����ҩ���DZ����Ŀǰ���ڶ����ٴ�ǰ�о��еõ�Ӧ�á�
�ڽ�ĸ����չʾ�����Ļ����ϣ�Maute���ˡ�[82]����˫�Ŀ���ԣ��������� PD-L1 ���и������ľ��������������� HAC-I (HVIHEGTVVI) �� HAC-V (HVVHEGTVVI)�������ַ����У�Դ���� PD-L1 ����ý�������� PD-1 �ṹ��ĵ�һ���Ŀ�����ʶ����Ƹ�������ͻ��л������ڶ����Ŀ�ȷ���л��������ϡ��������������������λ�� 41 ������������Ӱ���л���ͬ�� HAC �����У��������õ� HAC �����ܹ��� PD-L1 ��ϼ�DֵԼΪ100 pmol L −1�������о�������HAC �ľ�������С�ͺʹ����������������ر��ǣ��� HAC �Ľ��з����Ա�Ǻ�ͨ�������ӷ���ϲ�ɨ�������ʾ�������Ƿ���� PD-L1���Ӷ�ʵ��ֱ��������ϡ�
���ڽ�������;۽��Ŀ�ɸѡ���Ե�ϸ������չʾ������Zhu ����[83]������һ�ְ���PD-L1�� (TPP-1, SGQYASYHCWCWRDPGRSGGSK)������ PD-L1 ���и߽����������������PD-1/PD-L1����á�����ͻ�����ʾ���� TPP-1 ���������ɼ��� T ϸ�����������߷�Ӧ���Ӷ�֤�����ڰ�֢�����Ʒ��е�����Ч����Andersen ����ͬ��[84]��Ʋ��ϳ���һ��Դ��PD-L1��Tϸ����λ����ΪPDLong-1��FMTYWHLLNAFTVTVPKDL�������а���һ��PD-L1������CD8 + Tϸ����λ��PDL1 15�C23��LLNAFTVTV�������߷��֣���PDLong-1���̼�������ͻ״ϸ����DC����������ֿ���������Tϸ����������һ���ֱ�����PD-L1���Tϸ�������¼����������ֱ�ӵ���DC������ֵ�����ԭ�ԡ����⣬�Ӱ�ʱ��ʩ�����Ĵ���PD-L1���Ƽ�������Magiera-Mularz���ˡ�[85]�о�����-57����-71����-99������������PD-L1�Ľ�����������߷������������ľ�������PD-1/PD-L1����ã�����������PD-L1����������-71��˳���������>��-57>��-99����Щ�������ӱ������������˵ļ�����ϼ����ڵ�������ϵͳ������к�ǿ��Ч�����������Ƽ����������������ϣ�����ʵ��һ����ϣ���ĵ����ð�֢�������Ʋ��ԡ�
�����ĵİ�֢����
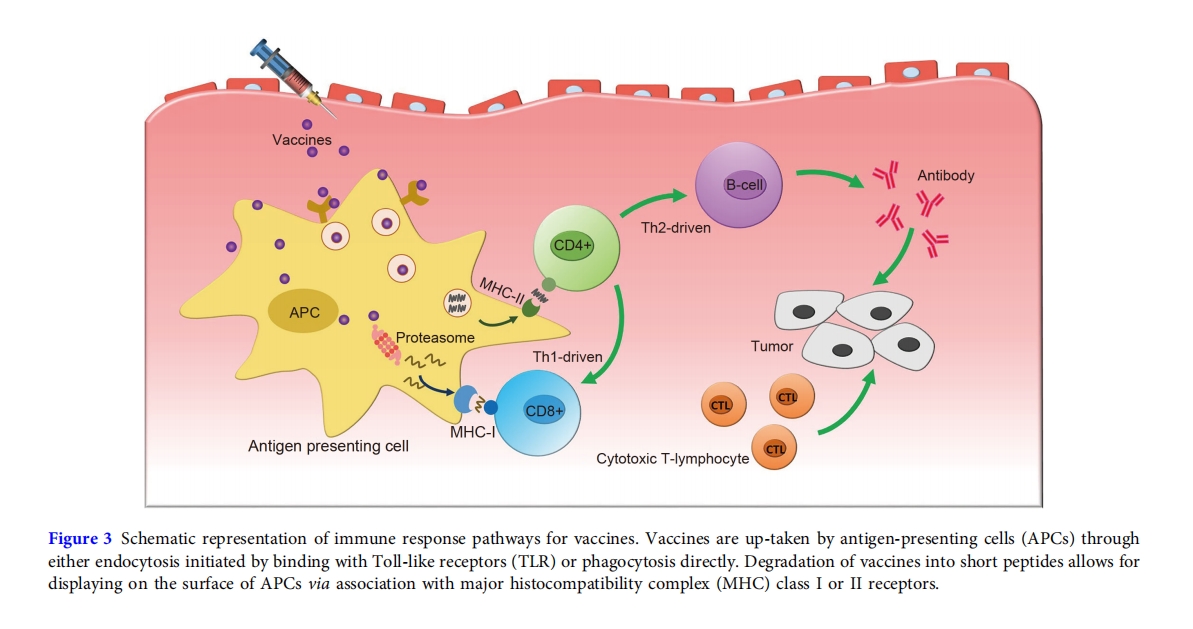
��Ϊ�������Ƶ���һ�ִ����Է�������֢����ּ�ڼ�����������ϵͳ����������ϸ����ؿ�ԭ���ֿ���ϸ�������ڿ�ԭ���������߷�Ӧ�ͳ��������䣬��֢�����ڰ�֢���������б��ֳ����DZ����������ֿ����������������Ʒ������ӹŴ����ִ�ҽѧ���õ��˹㷺��Ӧ��[111]���ڴ�ͳ�İ�֢�����У��Ӳ�ͬ���͵İ�ϸ�����ռ�������ϸ����ؿ�ԭ���ڴ̼� B ϸ���� T ϸ��������ϸ�����ڽ��������ڼ䣬ע����������ͨ���� Toll ������ (TLR) ����������������û�ֱ���������ñ���ԭ�ʵ�ϸ�����գ�ͼ 3��[112]��APCϸ���У����յĿ�ԭ�ڵ���ø��鵼�Ĺ����н���Ϊ���ġ�����������չʾ��APCϸ������ͨ������Ҫ��֯�����Ը����� (MHC) I ��� II �������Ͽɼ������߷�Ӧ���� MHC I ��ͨ·�У���ԭ�� MHC �� T ϸ������ (TCR) ��Ͽɲ��� CD8 +ϸ��������ϸ������ T �ܰ�ϸ�� (CTL) ϸ����ϸ�����߷�Ӧ[113]. Ȼ������ԭ�� MHC II �������ϻἤ�� T ����ϸ������������ B ϸ���Խ�����Һ���ߣ������ CTL ϸ���Խ���ϸ������[114��115]��ͳ�İ�֢�����Ǽ���������Լ��Ӳ����з�����Ļ��Ե����ǻ���������ijЩ��֢������ȡ���˳ɹ�������ͳ�������Ʒ���Ȼ������ս�ԣ���Ҫԭ��������Ⱦ����ص����߷��ա���ԭ�ȶ��Բ���������䵽�ܰͽ������Լ��Ѱ�����IJ����������ߵ�[116]��
�Ŀ�ԭ
�����������С��ԭ��λ����������ϵĶ��������侫ȷ�Ľṹ������������������Ϊ�������[117��118]��ԭ��TLR��MHC��������㹻���������ڴٽ�CD4 +��CD8 + Tϸ���IJ����Խ������߷�Ӧ������Ҫ�����ڶ��Ŀ�ԭ������ԭ�����ޣ�������������Э�������Ŀ�ԭ�����߷�Ӧ��һ�ֵ��͵IJ��ԣ���ʵ�ϣ�����Ϊֹ�Ѿ����������������������ڼ�Ҫ�ܽ��������磬�����漰����20�����������е��Ŀ�ԭ����ơ������������о�ʱ��ϳ����й�������ĸ���ϸ�����Ѳο������ط�[111��112]��
�������������������Ȼ�����ʵ����У���Щ�����ʵ��ǻ�������ƶ�����ΪDZ�ڿ�ԭ�����յ��Բ��������źͳ������������� 1���ݴ̼�;������Ƶ���MHC I��������صġ����ڼ���CD8 + Tϸ�����Ŀ�ԭͨ����8�C10����������ɣ�����MHC II�������CD4 + Tϸ�����صĿ�ԭͨ������13�C18�������ᣬ���ܶ��ij���û���ϸ������[117]���ڸ��廼�ߵ�MHC������������ԣ��ٴ������й۲쵽���Ŀ�ԭ�����������ԡ���������������߷�Ӧ�е��������������ϣ�ʹ���Ŀ�ԭ�ڰ�֢������������Ȼ����ǰ��[119]���������ܽ��˹�ȥʮ���ڿ������Ŀ�ԭ���ص��ע�ĵ�ԭ���Ժ�DZ�ڵ�����ԭ�Ի��ơ�
�������ѹ㷺Ӧ���ں�ɫ������������[120]. �������ɫ�����Ŀ�ԭ���Ժ�ɫ��ϸ���ֻ����ף������Ұ���ø��MART-1��Melan-A�����ǵ��� 100��gp100��[121]����Ҫ�ٽ� CTL �IJ����Բ��������������磬�Ұ���ø�Ǻ�ɫ�غϳ��е�����ø����������������ԭ���ģ�IE���Ұ���ø1�C9���Ұ���ø368�C376 [91]. �Ұ���ø368�C376�ṹ��� 3 λ�춬�����л����춬����ȡ������֤ʵ���зǷ�������ԭ�ԡ�ԭʼ MART-1 26�C35�ṹ��[92��122]���ٴ�������Ҳ��������ԭ�����ƺ�ɫ�������ߡ����⣬������ gp100 �е�gp100 280�C288�ṹ��[93]��ɫ�����ͽ�����ɫ��ϸ��������� CTL ���´ֻ��߲�������ԭ�ԣ����� CTL �������ϵ͡������������� gp100 209�C217�����������ϸ������ 2 (IL-2) ��Ͽɸ��ƶ�ϸ�������߷�Ӧ����ˮ�Ա�λ HGP100 25�C33���Ұ�����ص��� 2 (TRP2 180�C188 ) �ڵ���ˮ�Ա�λ����������ĺ�ɫ����������ԭ��Guo���ˡ�[95]ͨ�������ɾۣ�D,L-������-����-�Ҵ��������뿹ԭ HGP100 ���ܻ�������������֬�����¶�ǰ�����֬���塣Mirkin ����ͬ��[123]����ԭ HGP100 �ĸ��ŵ����ߴ̼����κ������Կ������硣Wakabayashi���ˡ�[96]�����Ͱ�������ɢ����Ϊ�������壬�����Ϳ�ԭTRP-2�ģ����ľ�����������л����Σ�KKKGSVYDFFVWL������������Ī�£�R-848������ϵͳ�����ƺ��������������ƺ���С���ת�Ʒ�����ֳ���ǿ�����������ԭHGP100��TRP2Ҳͬʱ����װ���пս�����������������У�����Ч�̼���ͻ״ϸ����DC��������죬����һ������������������-����TNF-������IFN-�á�IL-12��IL-4���Դٽ�����[124]��
Դ�����嵰�ı�λ���㷺�����Ŀ�ԭ������ OVA 257�C264��OVA 253�C266 ,OVA 250�C264�� OVA 323�C339����ͨ������ CD8 +ϸ������ T ϸ�����߷�Ӧ�������� 60 ���Dz�����ͪ������ø E2 �ǻ���ɵ������ң�Wang ����[86]������һ�ֲ���ģ������֧�ܣ���֧�ܰ�����ԭ OVA 257�C264�Ѻ��������������-�����ʻ��� (CpG)�����ֶ������ƽ̨���ֳ�Эͬʱ�յ������Ƽ�����ͻ״ϸ�����Ӷ���ǿ CD8 + T ϸ���IJ�������������⣬�� OVA 257�C264��λ�ӳ��������������ŵ� CCYSIINFEKL �����ֳ��Ʊ�ӫ�⿹ԭ-�����״أ���-AuNC��������ֳ���ǿ�����ߴ̼�����[125]ͨ���� AUNC ����ͬʱ���� CpG �������������õ���һ����ߡ����⣬Zhang ����[88]�������Գ�����ͻ״ϸ��Ϊ�е�����������B1��ܻ��ij�С�������������������磬����Ч�ؽ�����OVA 257�C264��OVA 323�C339��HGP100 25�C33���ڵ��Ŀ�ԭ�������ܰͽᡣ
Ĥ����ǵ���ճ���� 1 (MUC1) �ڱ�����Ƥ������ź�ת������ؼ����ã������֢�������ٰ������ٰ�����������ܰ�������ͨ�����ǻ���ͻ������ȱ���Ӷ�ʹ MUC1 ��������ԭ��[126]����һ�����������ܹ��յ� MUC1 ���ϸ������ T �ܰ�ϸ����Ӧ���Ŀ�ԭ����ơ�MUC1 ��һ�� I �Ϳ�Ĥ�ǵ��ף��������ǰ���ṹ���ɿɱ������� 20 ���������ظ����� (PDTRPAPGSTAPPAHGVTSA) ��ɣ�����ÿ�������ظ������е�˿������հ���л����и��ǻ���ˮƽ����ˣ����Ը��� MUC1 ��λ���ǻ�������� MUC1 ����Ŀ�ԭ�����磬Huang���ˡ�[127]��Ʋ��ϳ��˼��ֺ�ѡ����ͨ���� HGVTSAPDTRPAPGSTAPPA ������λ�� 9 �� 16 ���ǻ����հ���л���ϵ���װ�� Q11����Щ���� B ϸ����λ������������������ϸ������ T ϸ�����߷�Ӧ���÷�Ӧ�� I �� T ����ϸ��������⣬Zhao ����ͬ��[128]������˻��ڿ�ԭ��ѡ���Ĵ����ظ�TSAPDTRPAP����װ����Nap-G D F D F D Y D K�������ӵ�MUC1��ؿ��������硣
���˹㷺ʹ�õĺ���ϸ��ͻ�䵰�ס����嵰�� MUC1 �⣬һЩ��������ԭ�Ե���Ҳ��������������硣���磬HER2/neu �����ı�λ E75 ����Ʋ��������ٰ����ƣ�HER2/neu ��һ����������Ƥ���б����ԭ������[98]. Դ�� Wilm �������� (WT1) �� WT1 Pep427 Ҳ������������ԭ�Կ�ԭ�����뵥��̼���ܹ��۽�ϣ����յ����������� IgG ��Ӧ[99]. ŦԼʳ����״ϸ����-1��NY-ESO-1��[89]��һ������ԭ��֢غ�迹ԭ�����������֢����ɫ���������ٰ����и߶ȱ�����ܹ��յ� T ϸ��������ߡ�Gazzinelli ��ͬ��[129]����ԭ NY-ESO-1 ������ CpG DNA ���ӵ�̼���� (CNT) �ϣ�������һ���µĿ�������ƽ̨��Wang ��ͬ��[90]����E2����������ͬʱ������ԭNY-ESO-1��HLA-A2���Կ˷�������ԭ����ԭ�Ե͵����⡣���⣬��������ͷ��������HPV���ı�λ����HPV16 E7 11�C20 [100��101]��E7 86�C93 [101]��E7 43�C57 [102��103��130]��E7 49�C57 [101��103��131]�Լ� E7 48�C54 [103��104]�������߿�ԭ������ OFA 1��OFA 2 �� OFA 3[105]�ȶ����ѱ�������֢�������ƵĿ�ԭ���ɴ��������������һ���Ķ��Ŀ�ԭ�����ۻ��ܶ��Ŀ�ԭ�����ļ�β�ƿ�ԭ����϶��Ŀ�ԭ�Լ����Ի����Ŀ�ԭ���¿�ԭ���ȣ����ٴ������������˹㷺��ע������ʾ����֢���Ƶľ�DZ����
����������
�ö���������翹ԭ����������ȷ�Ľṹ������ϵͳ�ڵ�ѡ���бꡣȻ�����Ŀ�ԭ���������ԭ����Ȼ���㣬��Ҫ���������Ĵ��ڡ����ڶԿ��������߷�Ӧ�������˽⣬���������ѱ��㷺������ǿ��������ڼ�����߷�Ӧ����������Ϊֹ��ʹ�����εȶ��������������������ƣ���Ŀǰ���õ����������������������� TLR ���壬��Ȼ���ڽṹ���������⣬���»��������ҩ����[132��133]���������װ���ѱ�����Ϊ�����Ŀ�ԭ����Ҫ����Ϊ�����������װ�����չʾ��ۿ�ԭ��������
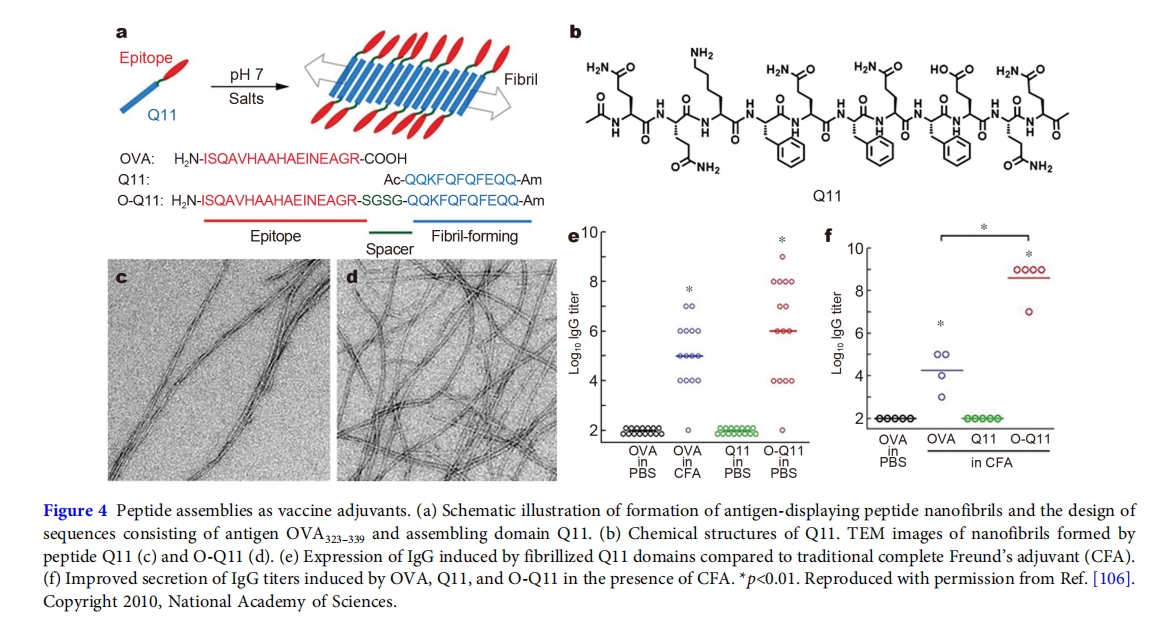
Collier �Ŷӱ����������������Ŀ����Թ���[106]�������߽���ԭ OVA 323�C339���ŵ��ܹ�����װ����ȷ������ά�Ľṹ�� Q11 (QQKFQFQFEQQ) �ϣ��Ӷ������� OVA-Q11 ���º���������װ����ȷ������ά��ͼ 4)������ OVA-Q11 ���� C57BL/6 С����Ѫ���� IgG �ζ�Ⱥ���������ӣ��Ӷ���ǿ������ԭ�ԡ�OVA-Q11 ��������ǿ������ԭ�Կ��ܹ�����ԭ��ά�ϱ�λ�Ķ�۱���չʾ������ϸ���о�������OVA-Q11 �յ���ϸ�����ӷ�Ӧȱ���� IgM ��Ӧ���ߣ��������ڶ����� T ϸ����Ӧ������ԭ�Ի��ơ�Q11 �ṹ���������Ӿ��и����ǻ����հ���л��� MUC1 ������λ���Դӱ���չʾ B ϸ����λ������ԭ��ά�п�����������ԭ��Q11 �ṹ������Ʒ��Collier ����ͬ��[107]������ԭ OVA 323�C339���ӵ���װ���� KFE8 (FKFEFKFE)������ OVA-KFE8 �γ�������ά������������ OVA-Q11 ��ǿ���巴Ӧ�����⣬Rudra ����ͬ��[134]�� Q11 �ṹ���е���Ȼ D-�������Ϊ L-�����ᣬ���о���������ά�� OVA ��λ���߷�Ӧ�Ķ�ӳ��ЧӦ���ڶ�ӳ��������ά�ı��������ϣ����߷����� L-��Ӧ����ȣ��� D-������������ɵ�������ά��ǿ��С��Ŀ��巴Ӧ���ӳ��˿�ԭ�ʵ�ʱ�䣬����� D-������������о��и����ܣ��������廯ѧ�����������ڵ�������ϵͳ����Ҳ���и����ܡ���Щ�����������������װ����Ϊ�����������������Դٽ�������֡�
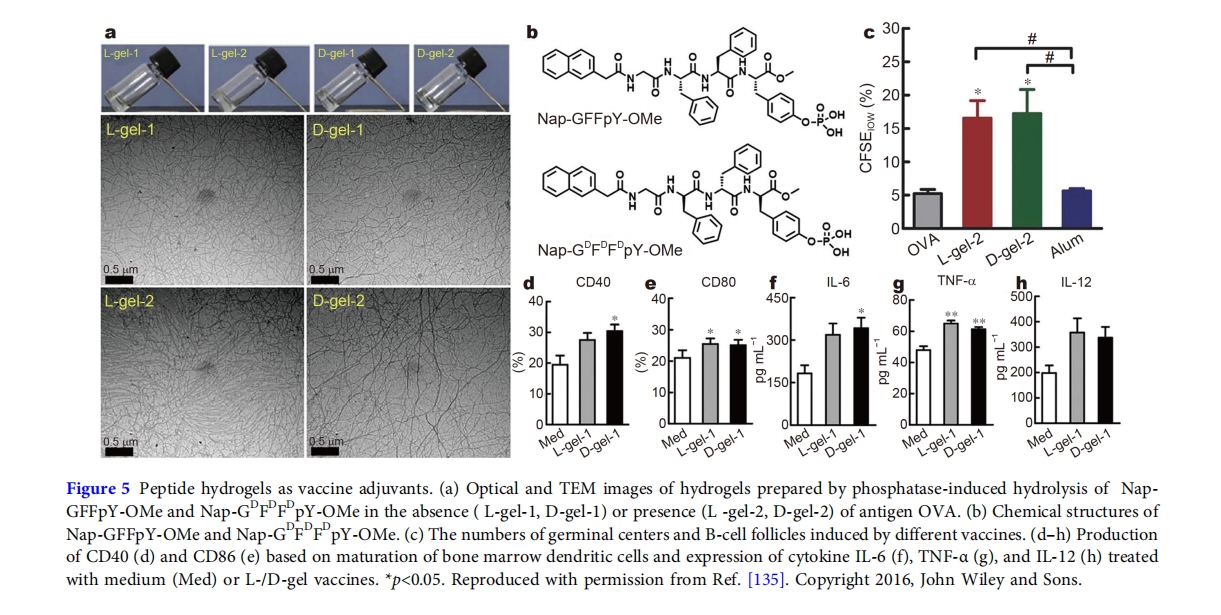
��������װ֮�⣬Yang ����ͬ��[135]����������װ����ɵ�ˮ������������������������ܻ�����������������������ﰲȫ�ԣ�ͼ 5)������ͨ���ϳ��� D �ͻ� L �Ͱ�������ɵ������������� Nap-GFFpY-OMe �� Nap-G D F D F D pY-OMe���о�������ˮ������������ֵĶ�ӳ��ЧӦ����Щ�������ڼ�������ø (ALP) �յ���ȥ���ữ�����·���ˮ���������������� OVA ���Ĺ���װ������ˮ��������Ϊ������ζ��ˮ������ OVA ����Ч���ա������о����������ֶ�ӳ��ˮ��������������Ч�յ����߿���IJ�����ϸ�����ӵķ��ڣ���������ϸ���Կ�ԭ����ȡ��ǿ����ԭ���ܰͽ�Ļ��ۡ���ͻ״ϸ���ij�����������ĵ��γɡ��ر��ǣ����߷����� L ��ˮ������ȣ�D ����ˮ�����ڻ��� OVA ��Ԥ����������������и��õ����ܡ�
Ϊ�˽�һ������øˮ����������������Ʊ����ѣ�����ֱ�ӴӶ�ӳ����������Nap-GFFY��Nap-G D F D F D Y������ˮ��������[108]������ˮ�����Ĵ�������ʹ���ܹ���Ч��װ��ԭOVA�����⣬ˮ��������װ��X����������ϸ������Ϊ��ԭ���Ƽ���ͨ��CD8 + Tϸ���;�����������������ӳ�����С��������ڡ���øָ����ˮ���������ƣ�Nap-G D F D F D Yˮ�����ڼ������߷�Ӧ�����L��ˮ�������ֳ���ǿ�������������ˮ���������ĸ��һϵ��Դ��Nap-GFFY��Nap-���������ѱ����������������������磬�� Nap-G D F D F D Y D ��C �����Ӵ�����ɻ�ɵIJл��������� Nap-G D F D F D Y D K �� Nap-G D F D F D Y D E[109]���ڰ���OVA����ʱ��������Ķ�����ɵ�ˮ�����ȴ�����Ķ���ˮ�������ֳ����õ��յ����߷�Ӧ����������ܹ�����OVA����Ч����������һ�������У������÷����忹��ҩȡ��G D F D F D Y Nĩ�˵�����Ԫ���Ӷ��õ�����ҩ�����ε���������[110]�����ҩ��Ŀ������ԣ�����OVA��ˮ������С���б��ֳ��Ƿ���������������������OVA�����⣬��MUC1��λ���ϵ�Nap-G D F D F D Y D K�����п�����ǿ��ԭMUC1������ԭ��[128]��
��֢���������е�����װ��
������һЩ�ٴ�ǰ�о����ٴ��������������Ʒ����������ȡ���˳ɹ��������ٴ���������Ȼ��������ԭ�Խϵ͵����⡣ԭ���ϣ�����Ȼ�����ʵ��������������ֱ�λ�����������Ķ���Ӧ���������������Ҳ�����˶��ı�ø�ٽ���Ŀ����ԣ����һ���������ĵ�ѭ�����������ܰͽ���Χ�Ļ��ۡ�������Щȱ��֮�⣬���ߵ�APCϸ���������Խ�һ���������Ʒ�����Ч����ԭ�Է������ս��������Щ���ǣ�̽�����ڰ�֢���������е��������ܱ��������Ҫ��
������װ����ȷ�����ṹ������̬��������[136]��������ά[137], nanoribbons [138]������[139]�����ɷǹ�������ã�������ˮ����á��������ú� ��-�� �ѻ�����ã������ķֲ����磬���н����Щ�����ǿ��DZ���������ϣ��������ṹ�ǵ��ͺ�չʾ���Ʒ�������ƽ̨����Ϊ���Ǿ��ж��صĽṹ���ԣ��������������Ժ����オ����[132��140]���������Ƽ����ϵ������ṹ�п����ӳ����Ƽ���ѭ�������������������������������ɶ��ЧӦ����ġ����⣬�����ṹ�ı��������������������������Ƽ���������λ�Ļ��ۡ�����װ�����Щ����ʹ�Ļ�������Ͼ�����Ϊ����������ǿ��ԭ���߷�Ӧ����Ϊ�������Ƽ�����ƽ̨�ľ�DZ�������ڽ����ڰ�֢���������ʹ������װ��İ�֢�����Ʒ������������������������Ƽ�����ϵͳ�������������硢��ԭ��������ϼ��������봫ͳ��֢���Ʒ������ϡ�
��������е�����װ��
���ܰ�֢��������Һ��ϸ�����߷�Ӧ�о��о�DZ���������������ٴ�������Ȼ�ܵ�����ԭ�Ե͵����š�����������ڶ���ԭ����ɵģ������������ơ�T ϸ���������� T ϸ�������͡������ܹ�����ԭ�ʵ�ϸ�����ٽ���ԭ���������͵���֧�ܿ��Ը�������ԭ�ԣ��Ӷ����ܴ����������ٴ����֡��봫ͳ���������ȣ�����װ��ľ�ȷ�ɶ�������ʹ������Ϊ��֢�������ϵͳ������о�DZ��������װ�����Ϊ����ƽ̨�Ĵ�ͳ������չ���������������Ƽ��ľۼ����Ի����Խ������ҵ���ϵͳ����Ϊҩ��������[141]. ҩ�������ӵ���װ���˾����Ĵ���ϵͳ���ŵ��⣬��������ƽ̨���濹ԭ���ܶȣ�������ҩ���Ƽ������óɷֵĺ������Ӷ��������ﰲȫ�ԡ����������ɱ�����ҩ������ͳ���������˷������ϣ������Ĵ���ϵͳ���ڰ�֢���������������ԭ������ϣ���ġ�
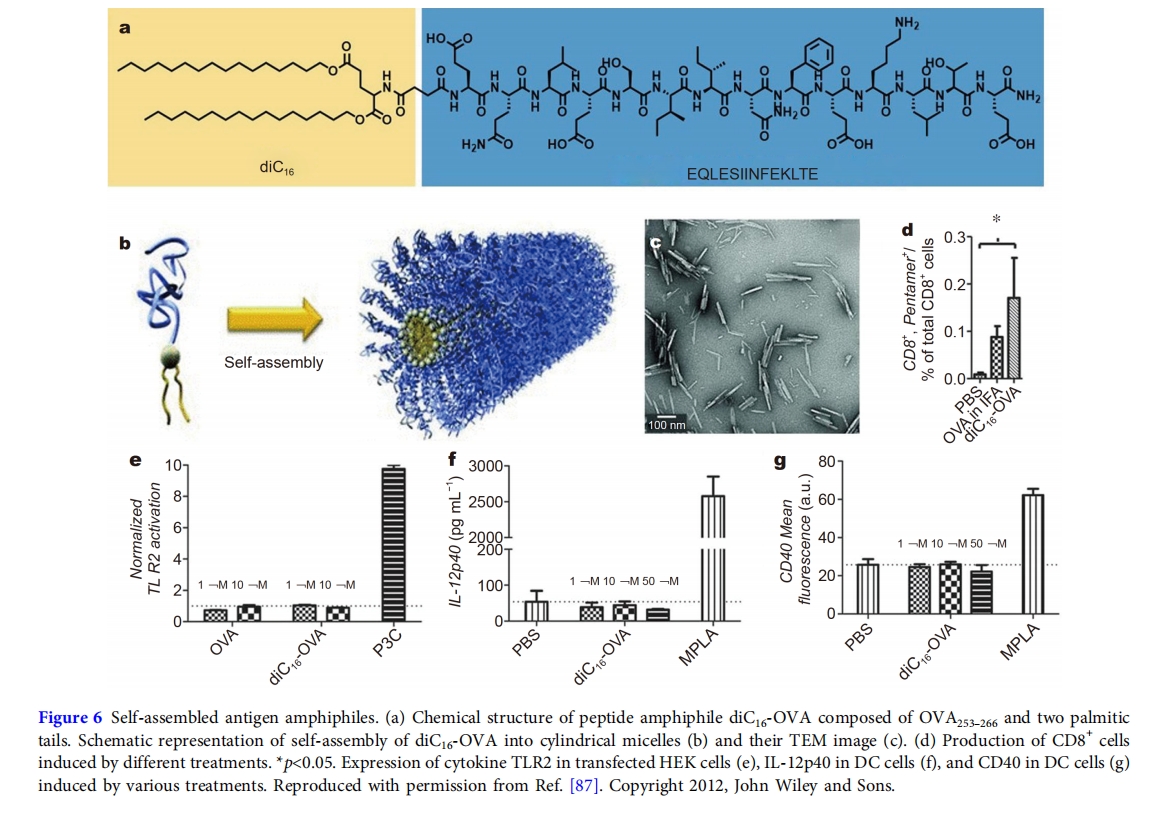
����ҩ�������Tirrell ��ͬ����[87]ͨ������������������ӵ��������嵰��ϸ������Tϸ����λ����OVA 253�C266��SIINFEKL���������˿�ԭ������Ŀ����Թ������Ӷ��γ��˿�ԭ������DiC16-OVA��ͼ 6). DiC16-OVA ����������װ�γ�Բ���ν�����ֱ��ԼΪ8 ���׳��ȷֲ���Ҫ��200��500���ס�����Բ���ν��������β��ɵ���ˮ���ĺ�չʾ��ԭ��λ����ˮ������ɡ������������£���ԭ�����ȶ�������������Сʱ�������ڿ�ԭ����ͻ��۵��ܰͽᡣϸ���ⶨ�������� DiC16-OVA ��װ������·���ϸ�����ᵼ�� DC ϸ���̼��������߷�Ӧ�������о��������Ի���������С��������ƣ���ͨ������ϸ������ T ϸ�����߷�Ӧ�����������������ӳ�С��Ĵ��ʱ�䡣��һ�����������ԭ����������װ�����ṹ��һ����Ч�Ŀ�ԭ���Ͳ��ԣ�Ҳ�ǿ����Ը�����ԭϵͳ����Ч���ԡ�
�������͵�һ��֢�����⣬����װ�廹�����Կɵ����������Ͷ��ֿ�ԭ��Ϊ�ˣ�Collier ����ͬ��[142]������һ�ִ�����������ά�IJ��ԣ��ò����ܹ����϶��ֵ����ʣ�ͬʱ����������Ĺ����������ԣ�ͼ7���������ǩ���� MALKVELEKLKSELVVLHSELHKLKSEL �ںϣ���Ϊ ��Tail������Һ�лᾭ���� �� ������ �� �۵��Ļ�������ת�䡣�� ��Tail �ںϵ��� ��Tail-GFP �������װ��������ά���� Q11 ͬʱ�ܽ⣬��ʹ�ںϵ���Ч���ϵ�������ά�С�������ά�����ϵ����ʵļ����ɸ�����Һ�еĵ�����Ũ�Ⱦ�ȷ�������÷������ڽ���ͬ�ĵ����ʣ��� ��Tail-GFP �� ��Tail-cutinase���Կɿص�Ħ�������ϵ�������ά�У��Ӷ��յ�������������صĿ���ζȡ���ϵ����ںϵ��������Ժ;�ȷ���϶�������ʵ�������������һ�ֿɶ��ƵĶԭ����֧�ܣ���֧�ܵĿ�ԭ���ƿɸ��ݸ���ԭ�ļ����ͼ�ǿ���䷽���п��ơ��ԭ����ƽ̨����ͬʱ�Բ�ͬ��ԭ������������ͶԵ�һ��ԭ����и����������ƣ��ڰ�֢������ַ�����о��DZ����
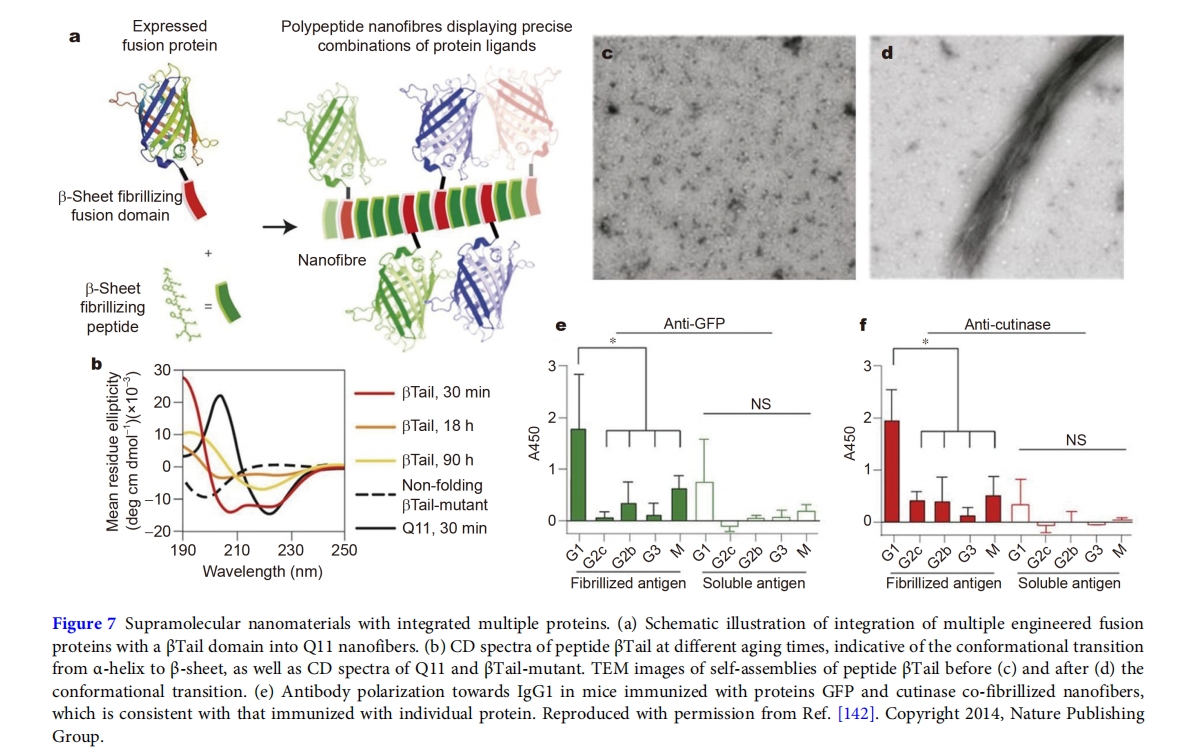
��������Ʒ��е�����װ
�������������������Ƹ��ӣ��Ҹ�����������Դ�һ�������Ʒ����ڴ��������²����Ե������߷�Ӧ����������ԭ�Խϵͣ������˰�֢�����������ٴ������е�Ӧ�á�ͬʱ���ϲ�ͬ�Ʒ����������Ʊ���Ϊ��ͨ������;����ǿ���߷�Ӧ����Ч����[143�C145]���⣬�뻯�ơ����Ƶȴ�ͳ�Ʒ��ļ�ʱ����Ч����ȣ����߷�Ӧ�ij���ʱ����Խϳ��������߷�Ӧͨ����Ҫ������ҩ�����һ�������˰�֢�������������ڰ�֢�����е��ٴ�Ӧ�á��������ij��������������������Ͽ���ǿ���߷�Ӧ����ת����֢�����������ƾ���Эͬ����[146��147]��ˣ�Ϊ����Ч�ӳ����ߵ������ڣ�����Ҳ�������漰��ͳ�Ʒ��������Ʒ�������Ʒ�����������װ����й��ۺͷǹ������ջ���Ĺ�������������ѹ㷺Ӧ������������Ʒ������ڽ������������������װ����Ϊ�����Ʒ���ͬ����ƽ̨������������Ʒ�����ȡ�õĽ�չ��ֵ��ע����ǣ����ϸ�ҩ�����Ʒ���ʹ�ø�ҩ��ǿ���ķ���[38]�������ǹ�ͬ���Ͳ��ԣ����Ľ��������ۡ����⣬ʹ�ö�����Ϊ��������IJ���[50]�����ص��ע�ĵ�����װ�����Ҳ���ᱻ���ǡ�
��������Ʒ�
������װ����ɵ������ṹ��ˮ�����ܹ���װ�����������Ƽ���������ԭ��ϸ���Ϳ���ȴ������ʵ��ĺ�С���Ӻϳ�ҩ���С�����ʡ��ر��ǣ�����װ���ˮ��������Ϊ����ע������Ʊ����������˸�ҩ�ڼ�������λ���Ƽ���ά�֡����⣬����װ��������방֢�����־�������������صĴ̼��½��в��ݣ��Ӷ��п���ʵ�����Ƽ��Ŀռ��ʱ�䰴���ͷš���ˣ���������װ�干ͬ���Ͷ����������Ƽ�������ǿ��֢�������������߷�Ӧ�ľ�DZ�������������ܽ����������ʹ���������ͬ���ݶ��������Ʒ����Ƽ������ӡ�
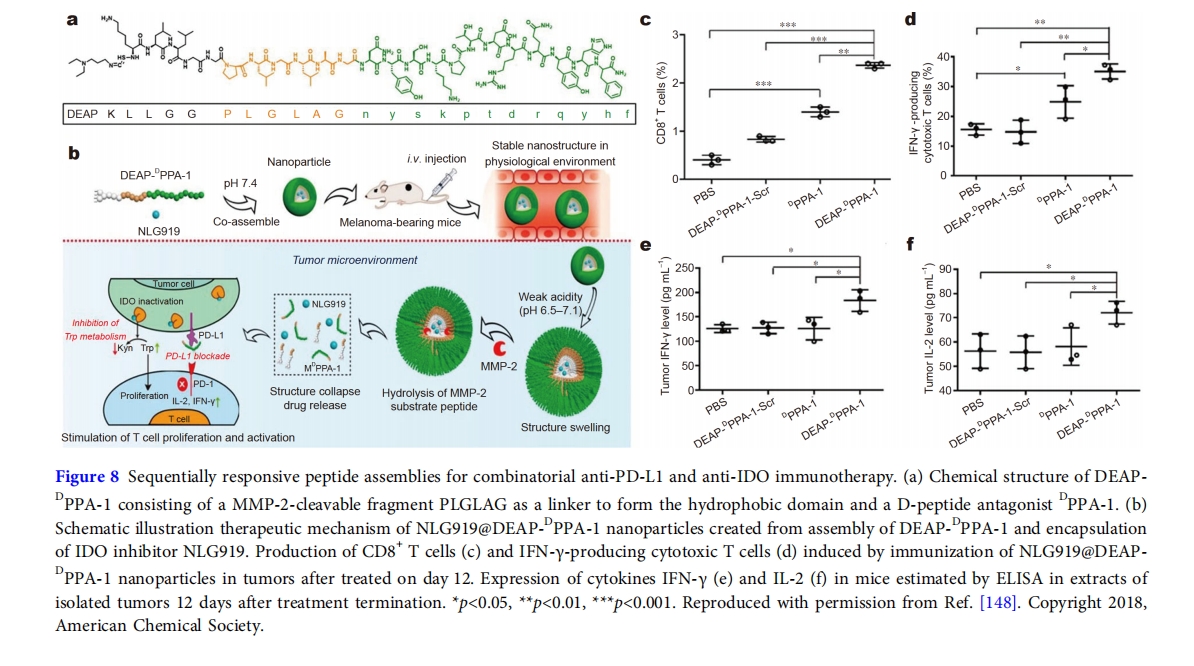
���ڹ����Ͳ��ԣ�����װ�����ڹ����Ͷ��Ŀ�ԭ��С����ҩ�����Ƽ���Nie ����[148]������һ������װ�������ӣ����������ӻ��ں��� 3-���һ������������������� (DEAP) Ƭ�Ρ��ɱ����ʽ�������ø-2 (MMP-2) �ѽ�Ľṹ�� PLGLAG �Ͷ� D ������ ( D PPA-1) ���������ĵĹ���װ������ҩ�� NLG919 �ǵ��Ͱ� 2,3-˫����ø (IDO) �����Ƽ�����ø��һ����������ø����Ϊ���ܹ��� L-ɫ����ˮ��Ϊ L-Ȯ���� (ͼ 8)������DEAP�������ӻ�������ˮ�Խ��ͣ������������յ������������ӣ���ΪNLG919@DEAP- D PPA-1���Ľṹ���ͣ��Ӷ��ٽ�MMP-2�ѽ�PLGLAG�ṹ��ʹ�������ӱ������Ӷ��ͷ����صĻ������ϸ��������MMP-2��ʹ����D PPA-1��ҩ��NLG919��������λ��Χ�ռ��ͷţ����Ƿֱ����PD-L1��IDO����ʽϸ��������ʾ���ѽ��LAG DPPA-1�ṹ������������Nĩ�˲л������Ա��ֳ���PD-L1��ǿ����������������������£���MMP-2�����£�NLG919@DEAP- D PPA-1�����յ���IDO��������������NLG919�൱����NLG919@DEAP- D PPA-1���ƻ��к�ɫ������С���������������CTL��ˮƽ���Ӷ���Ч����������������
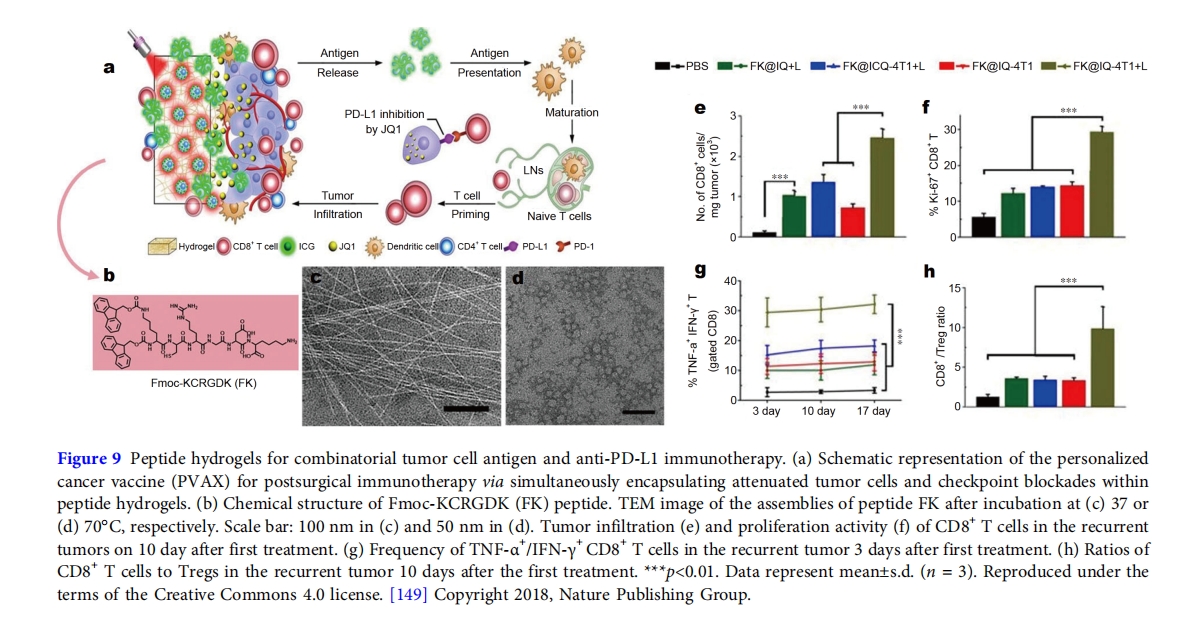
��С�������ƿ�ʼ������װ���������������Ʒ���ͬ��������ϸ�����硣�ڴ˱����£�Li ����ͬ��[149]�����˸��Ի���֢���磨PVAX��ͨ��ͬʱ����������ϸ���ͼ�����ϼ���װ����ˮ�����У��Ӷ��γ� FK@IQ-4T1 ���磨ͼ 9)����С�� 4T1 ��������������ֲ�����ռ���������ϸ�������� Foxp2 �̶���������Һ��������Ȼ�����ˮ������װ��һ��С����ҩ�� JQ 1 ��������ṹ���ĩ���⵰�� BRD4 �����Ƽ�����ͨ������������ PD-L1 �ı��������������ܡ���ˮ������һ������ Fmoc-KCRGDK (FK) �Ʊ����ɣ��������� N ��������л��Ϻ������� Fmoc ���ţ����յ��ĵ�����װ��ͨ�� ��,��-�ѻ�������ȶ�ˮ�������� RGD Ƭ�����������Ƽ�������������͡�Ϊ�˴ٽ����Ƽ����ͷţ����ü�����������ЧӦ���������װ����̬ת�䣬�����ؾ��и߹���ת��Ч�ʵ�ӫ��Ⱦ�� ICG ���ٽ� 4T1 �� JQ1 ���ͷš���ʽϸ������ø��������������������ʾ��FK@IQ-4T1 ����ٽ��������DC ���죬���� CD8 + CTL ���߷�Ӧ����� PD-1/PD-L1 ����ͨ������ BRD4 �������ʵ���һ��֤����PVAX������Ԥ����������������ת�Ʒ��������ͨ�������������߷�Ӧ����������������������������ơ�
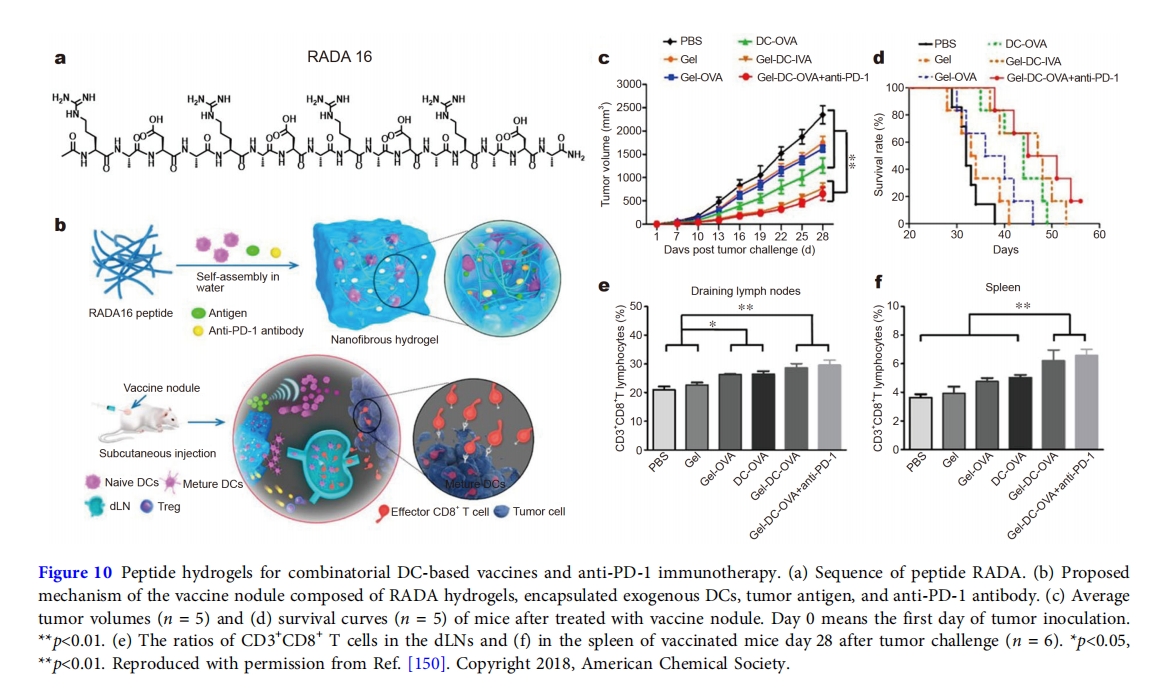
���⣬�����������ˮ������Ϊ���壬���� [150]�γ�������ͨ��ͬʱ����Դ��DCϸ����OVA��ԭ����PD-1�����װ��RADA16��ˮ�����У�ͼ 10)���� RADA16 ��һ�ֽ������ˮ����ˮ���У��ѱ�֤����һ����Ч��ˮ�������������γɼ�̵���ˮ����[14]���� DC ϸ����װ��ˮ�����п��Ա��� DC ϸ����ϸ���������ӳ�����ע�䲿λ�ij���ʱ�䣬���ٽ������䵽�ܰͽᡣDC ϸ���Ϳ�ԭ�����������Դ�Ժ������� DC ������߷�Ӧ���Ӷ���ǿ��ԭ������ T ϸ�����ߡ����� PD-1 ��������װ��ˮ�����У�ͨ����ֹ�� PD-1/PD-L1 ��������� MHC I �µ����ٽ������� CD8 + T ϸ������ֳ�������Ȼ���������ʵ��֤ʵ��Gel-DC-OVA�����ܹ��յ�DCϸ������ʹ̼���ԭ������ЧӦTϸ������Gel-DC-OVA+anti-PD-1����С������ӳ���������������ڲ���������������������Gel-DC-OVA+anti-PD-1�յ��ij�ǿ����ԭ�Թ�����CD8 + Tϸ�����ܰͽ�Ľ���Ͷ�������Tregϸ�������ơ�
���ϳ������ƺ���������
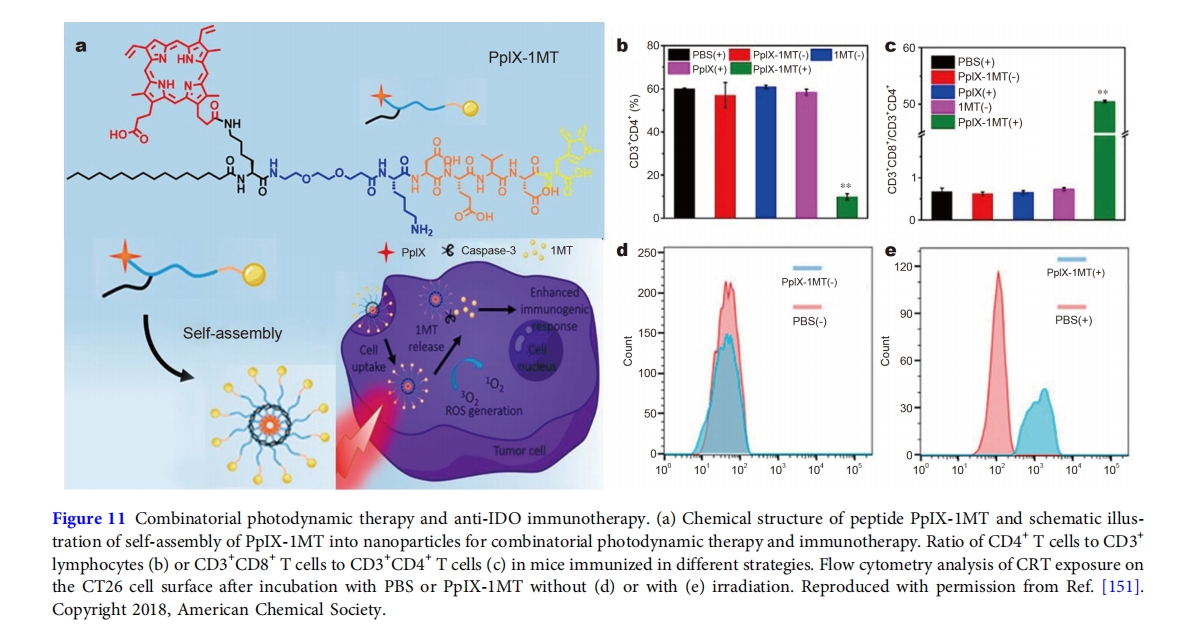
�������ϲ�ͬ�������Ʒ�������װ�廹��ʾ���ڳ������ƺ��������������ϸ�ҩ�ľ�DZ�������磬����Ʒ��ͻ��ƶ�����Ч����ԭ����������������ͬʱҲ������������ԭ��ϸ�����������������߷�Ӧ����ǿ����������������ˣ������������������Ƶ�����Ӧ�ñ��ֳ�Эͬ�������ã�������ٴ�ǰ�о��������˹㷺��ע���ڴ˱����£��ε��� [151]������һ����Ƕ������ɵ��������ӣ���ΪPpIX-1MT��������Ϲ���Ʒ��������Ʒ���ͼ 11). ��PpIX-1MT����ˮ����ɣ����������β������PpIX����ˮ�ΰ�����PEG����ͨ��caspase-3���ѽ��DEVD�����������������Ƽ�1-��ɫ���ᣨ1MT�����ӡ������������£���PpIX-1MT�ۼ�����������ƽ��ֱ��ԼΪ128.5 ������Ҫ����ˮ�����������ͨ����ǿ���Ժ��������ð�������ϸ����������PpIX�����Ļ�������ROS���յ���ϸ��������caspase-3���յ������и�DEVD���У��Ӷ��ͷ�1MT���ӣ�IDO���Ƽ������Է�ֹCTLϸ�����µ���Tregϸ��������������ơ���ʽϸ����֤ʵ�˹���Ʒ����ڸ���������ϸ������ı�¶���������ԭ��ϸ���������Լ�CD8 + Tϸ�����߷�Ӧ�ļ�������о����֣�������Ʒ��ٽ��İ�ϸ����������������IDO������������߷�Ӧ��ǿ���ϣ����Դﵽ���ԭ���������������ת������Ŀ�ġ������о�����Ʒ��֣�����Ʒ�������ԭ��������������������CD8 + Tϸ�����߷�Ӧ������˷�ת�������Ӷ�������һ�ּ���Эͬ���Ʋ��ԡ�
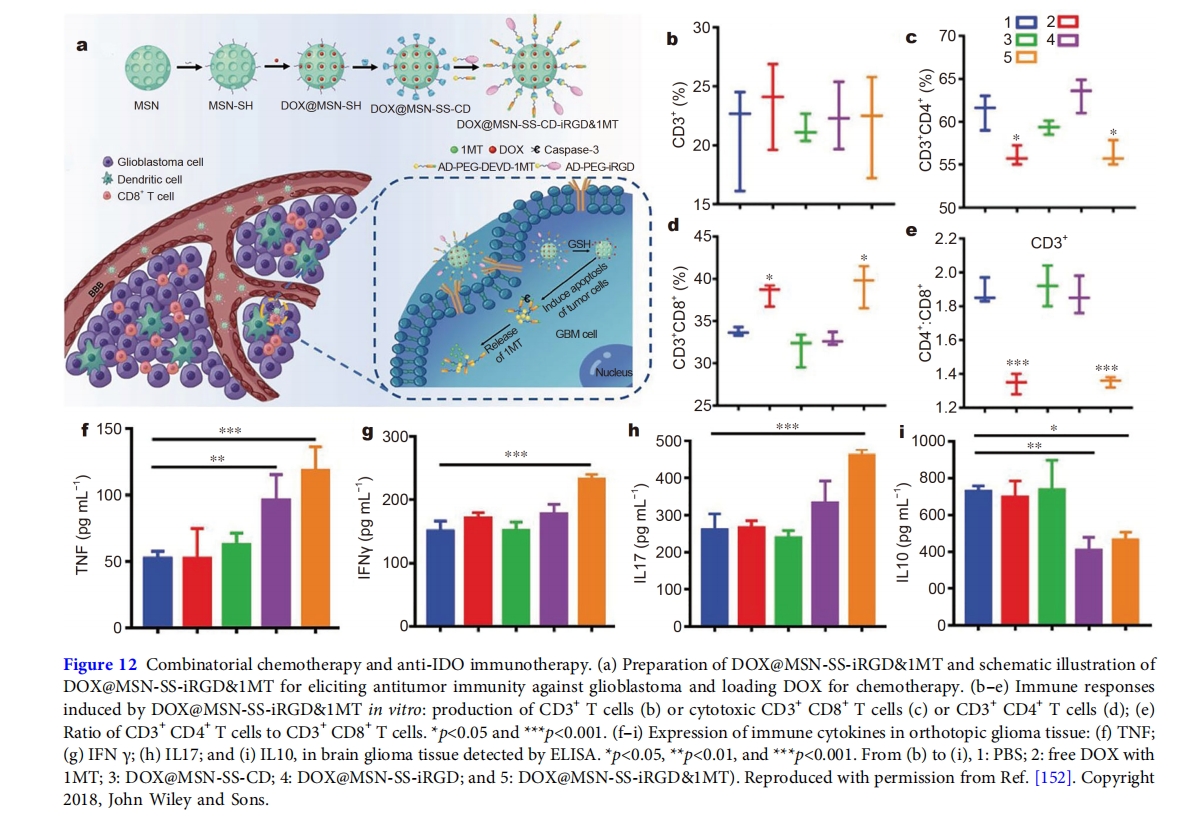
���⣬�Ž���ʵ���һ������˻���ͬʱ��ԭλ������ע�뻯��ҩ�ﰢù�أ�DOX�������������Ƽ�1MT�����ϻ��ƺ������Ʒ����ƽ���ĸϸ������ͼ 12�� [152]��������ϵͳ��Ϊ DOX@MSN-SS-iRGD&1MT��������ҩ�� DOX �Ľ���������������� (MSN) ��ɣ����ڱ�����ʾ��������ϼ� 1MT ������ϸ�������λ iRGD�������� DOX �� MSN �� ��-CD ����ͨ�������������չʾ����ͨ�� ��-CD-������Ϸǹ������ӡ��������Ƽ� 1MT �ͽ����ͨ��DEVD �ṹ�������ڱ�¶�� caspase-3 ʱ�ͷ����Ƽ����� GSH ��ԭ��������� ��-CD ��ȥ���ٽ��� DOX �� MSN �е��ͷš�����ʵ���������DOX@MSN-SS-iRGD&1MT�������������յ�������ϸ���������������������߷�Ӧ�������о�֤��DOX@MSN-SS-iRGD&1MT�ܹ���Ѫ�����ϣ���������λ���пռ���ͺ��ͷ�DOX��1MT��ͨ�����ƺ��������Ƶ�Эͬ���ã��շ�CTL���߷�Ӧ������Tregϸ���Ļ���Ӷ��ӳ�����������С��������ڣ�����������������
�ܽ���չ��
��֢�����Ʒ������������������߷�Ӧ�Ա����ֲ���ϸ����DZ�ڵ��յ������������Է�ֹ��֢������ת�Ʒ�������ƣ������������о��й�����Ӧ��ǰ�������������Ļ�����װ������ƹ��ܼ���������߷�Ӧ�Ļ��Ʒ����ܽ��˻����ĵİ�֢�������Ʋ��ԡ������ĵķǷ����������Ժ��䵰���������Ľṹ��������Ȼ�����ѱ�����������ϼ�����ԭ���������������Ƽ���������װ�����Կɿط�ʽ������ͻ�ͬ�������Ƽ�������ֳ����Ƚ�������������Ϊֹ�������ٴ�ǰ�о����������Ʒ����е������߷�Ӧ��������������������������֤�������������Ʒ����ٴ������еľ�DZ����
�����ڹ�ȥʮ����ȡ���˽�չ���������ĵİ�֢�����Ʒ����ٴ�Ӧ����Ȼ������ս�ԣ�����ֻ������ʵ�����������Ҫ����ս���ڴ�������������ԭ����Խϵͣ�������������[153]�뿹�������ȴ���ӵ�����ȣ�Դ�Ե����ʵı�λͨ�����ض��б���ֳ��ϵ͵�ѡ�������������⣬�����аб������ı��������Կ��ܻ��һ���������ֹ����������ٴ�ǰ�����й۲쵽�����������ú��ۺ������Ļ������Ʒ�����һ���ش���ս���ڸ�ҩ��ȫ�ԡ���Щ�����ú��ۺ�������Դ�����ƻ����ϵͳ�����������Բ���Ƶ��Ѱе����Լ��ɴ˲������������ߵȡ����������ĵ���ϵͳ�Ѿ���������Ƶİ����ͷ�Ч��������������λ�����ͷŻ�����Ȼ������ս��[154]��
���ǵ�������ս���������и�����ԭ�ԺͿɽ��ܵĸ�ҩ��ȫ�Ե��Ƽ�����Ϊ���������Ʒ���δ����չ���������Ʒ��Ŀ���������ļ�����ϼ�����ԭ���¿�ԭ�����[155]��һ�ֶ�ܲ��ԣ���������Ƽ���б������������Ӷ��������������߷�Ӧ[156]��ͨ�������µĵ���ϵͳ�����������ܰͽ��л T ϸ���Ľ���ͻ��ۣ������Ͼ���Эͬ���õĶ��������Ʒ�������DZ�ڵظ�������ԭ�ԡ��ر��ǣ������������־����Ƶ�ҩ���ͷźͽ��������������λ�������ϵͳ���ܻ��ֹ���Ƽ����Ѱ��ͷš����⣬���������ݵ�ҩ������������װҲ�ѱ�����Ϊһ���µ�ҩ����Ͳ��ԣ������䷽�м����������������ʣ���˿���������尲ȫ�ԡ������Щ�뷨�������ĵİ�֢�����Ʒ���δ����֢���Ƶ�һ����ǰ;�IJ��ԡ�
��������������Ϊ��ҵ����ѧϰ����Ȩ��ԭ����ԭ��־���У�������Ȩ������ϵɾ�������±�ע���������³����������Ķ�ԭ�ļ��ο����ף����Ķ�ԭ��־��