
еЊвЊЃКУтвпЮЂЛЗОГдкЙЧШБЫ№аоИДжаЦ№зХжСЙиживЊЕФзїгУЁЃЮЊСЫДДдьДйНјГЩЙЧЕФУтвпЮЂЛЗОГЃЌбаОПШЫдБе§дкЬНЫїдіЧПM2аЭОоЪЩЯИАћЗжЛЏЕФЗНЗЈЁЃЙІФмадыФвбБЛЗЂЯжПЩвдгааЇИФЩЦетвЛЙ§ГЬЃЌЕЋЫќУЧЪмЕНаЇТЪЕЭКЭНЕНтЫйЖШПьЕФЯожЦЬхФкЁЃЮЊСЫПЫЗўетаЉЮЪЬтЃЌЩшМЦСЫОпгаM2ЕїНкКЭздзщзАФЃПщЕФыФзїЮЊЙЙНЈГЌЩљЯьгІФЩУзЯЫЮЌЫЎФ§НКЕФЙЙНЈПщЁЃетаЉФЩУзЯЫЮЌПЩвддкГЌЩљДЬМЄЯТвдЪБМфвРРЕадЕФЗНЪНДгЫЎФ§НКжаЪЭЗХГіРДЃЌМЄЛюЯпСЃЬхЕФЬЧНЭНтДњаЛКЭШ§єШЫсбЛЗЃЌвжжЦЛюадбѕЕФВњЩњВЂдіЧП M2 ОоЪЩЯИАћЕФМЋЛЏЁЃИУЫЎФ§НКЭЈЙ§ДЅЗЂ M2 ОоЪЩЯИАћЗжУк BMP-2 КЭ IGF-IЃЌМгЫйЙЧЫшМфГфжЪИЩЯИАћ (BMSCs) ЯђГЩЙЧЯИАћЕФЗжЛЏЃЌеЙЯжГігУгкЙЧдйЩњЕФЯШНјжЮСЦЧБСІЁЃвђДЫЃЌФЃПщЛЏЩшМЦЕФЗТЩњГЌЖЬыФФЩУзЯЫЮЌЫЎФ§НКЮЊжиНЈЙЧаоИДЕФГЩЙЧУтвпЮЂЛЗОГЬсЙЉСЫвЛжжаТВпТдЁЃ
1 НщЩм
ЪЕЯжРэЯыЕФЙЧШБЫ№аоИДЖдгкДДЩЫЁЂжзСіЧаГ§ЁЂЮЎЫѕадЙЧВЛСЌЕШЛМепЖјбдШдШЛЪЧвЛЯюжиДѓЬєеНЁЃ[1] ЙЧШБЫ№ЕФдйЩњЮЂЛЗОГвРРЕгкИЩЯИАћЁЂЯИАћЭтЛљжЪЃЈExtracellular Matrix, ECMЃЉЁЂЗжУкЕФЩњЮяЛюадвђзг[2]ЃЌЦфЯрЛЅзїгУаЮГЩЙЧЬивьадЮЂЛЗОГ[3]ЃЌЫцзХЙЧУтвпбЇЕФЗЂеЙЃЌдНРДдНЖрЕФбаОПБэУїЃЌЙЧдйЩњЙ§ГЬВЂЗЧМђЕЅЕФЙЧаЮГЩКЭЙЧЮќЪеЙ§ГЬЃЌЖјЪЧЖрИіЯЕЭГУмЧаЯрЛЅзїгУЕФНсЙћЃЌАќРЈЙЧїРЯЕЭГЁЂУтвпЯЕЭГЕШ[4] УтвпЯИАћЗжУквђзгРДНЈСЂЕїНкГЩЙЧЯИАћКЭЦЦЙЧЯИАћЗжЛЏЕФЮЂЛЗОГ[5]ЃЌдкЙЧаЮГЩКЭЙЧЮќЪежаЦ№зХжСЙиживЊЕФЕїНкзїгУЁЃОоЪЩЯИАћЪЧЯШЬьУтвпЯЕЭГЕФживЊзщГЩВПЗжЃЌОпгаЯджјЕФСщЛюадЃЌПЩвдИљОнЮЂЛЗОГжаДцдкЕФЗжзгНщжЪзЊЛЏЮЊДйбзад M1 ОоЪЩЯИАћЛђаоИДад M2 ОоЪЩЯИАћ [6]ЃЌM2аЭОоЪЩЯИАћзюжеЭЈЙ§ЪЭЗХХдЗжУкЯИАћвђзгШчBMP2ЁЂTGF-ІТЁЂIGF-IКЭЭтУкЬхЗЂЛгживЊЕФПЙбзЕїНкзїгУЃЌЗжУкЯИАћвђзгДйНјИЩЯИАћЗжЛЏКЭзщжЏдйЩњ[7]ЃЌвђДЫЃЌЕїПиM2аЭОоЪЩЯИАћМЋЛЏНЈСЂгаРћгкЫ№ЩЫаоИДЕФУтвпЮЂЛЗОГвбГЩЮЊЙЧдйЩњСьгђЕФЙиМќЬєеНЁЃ
ФПЧАгУгкЕїПиM2ОоЪЩЯИАћМЋЛЏЕФЗНЗЈжївЊАќРЈЩњЮяЛюадМСЁЂУтвпЕїНкМСКЭЧїЛЏвђзг[7b-d]ЃЌОЁЙмШчДЫЃЌОоЪЩЯИАћЕФГжајЕїНкЖдгкЙЧШБЫ№аоИДжСЙиживЊЃЌЕЋгЩгкШБЗІПЩвдГЄЦкЪЙгУЕФАВШЋгааЇЕФЙІФмЗжзгвдМАФбвдЪЕЯжПижЦЕФОжВПЪЭЗХЃЌетвЛФПБъЩаЮДЪЕЯжЁЃ[4aЈC8 ]ЁЃвђДЫЃЌЦШЧаашвЊбАеваТЕФЙІФмВпТдЃЌгааЇЕїНкM2аЭОоЪЩЯИАћМЋЛЏЃЌДгЖјИФБфУтвпЮЂЛЗОГЃЌДЬМЄЙЧдйЩњЁЃгыЦфЫћЩњЮяЗжзгЯрБШЃЌГЌЖЬыФОпгаИќгХдНЕФЩњЮяЙІФмЁЂИќИпЕФАВШЋадКЭИќЕЭЕФКЯГЩГЩБО[9]ЁЃШЛЖјЃЌЬьШЛыФПЩФмВЂВЛзмЪЧЪЪКЯзїЮЊжЮСЦМСЃЌвђЮЊЫќУЧОпгаЙЬгаЕФШѕЕуЃЌР§ШчАыЫЅЦкЖЬЁЂЮяРэЛЏбЇаджЪВЛЮШЖЈКЭЫЎНтбИЫйЁЃвђДЫЃЌПЊЗЂОпгаСйДВзЊЛЏЧБСІЕФУтвпЕїНкыФЃЌАќРЈгХвьЕФвЉЮяаЮГЩФмСІЁЂПЙНЕНтадФмКЭЩњЮяАВШЋадЃЌжСЙиживЊЁЃВЛГЌЙ§АЫИіЕФЙбыФЭЈГЃБЛГЦЮЊГЌЖЬыФЃЌзюНќЪмЕНЙуЗКЙизЂЁЃвђДЫЃЌЮвУЧЕФФПБъЪЧПЊЗЂвЛжжФмЙЛЕїНкЙЧШБЫ№УтвпЮЂЛЗОГЕФГЌЖЬыФЯЕЭГЃЌвдМгЫйГЩЙЧВЂДйНјЙЧЫ№ЩЫдйЩњЁЃ
НќФъРДЃЌЩњЮяЛюадВФСЯзїЮЊЙІФмЗжзгв§Ц№СЫЙуЗКЙизЂЃЌВЂж№НЅБЛгУгкЩњЮявНбЇгУЭОЃЌАќРЈжЮСЦУтвпШБЯнЯрЙиМВВЁЁЂМгЫйМфГфжЪИЩЯИАћЕФГЩЙЧЗжЛЏЁЂИФЩЦзщжЏбЊЙмЛЏЁЂДйНјЩёОдйЩњЕШ[10]ЃЌБОбаОПЩшМЦСЫвЛжжОпгаДйНјM2аЭОоЪЩЯИАћМЋЛЏМАBMSCЗжЛЏФмСІЕФГЌЖЬыФ(Ser-Glu-Ser-Ser-Glu,SESSE) (ЗНАИ 1 ЃЉвддіЧПЬхФкЮШЖЈадЃЌЮвУЧФЃПщЛЏЩшМЦСЫ SESSE ыФЃЌОпгазщзАФЃПщЃЈCFFЃЌЛЦЩЋЃЉКЭ M2 МЋЛЏФЃПщЃЈSESSEЃЌТЬЩЋЃЉЃЌПЩвдздзщзАГЩФЩУзЯЫЮЌЁЃДЫЭтЃЌЮвУЧКЯГЩСЫГЌЩљЯьгІЫЎФ§НКОЭЕиВЩгУздзщзАГЌЖЬыФФЩУзЯЫЮЌЃЈМЧЮЊUPN@ЫЎФ§НКЃЉаоИДЙЧЫ№ЩЫв§Ц№ЕФШ§ЮЌПеМфШБЫ№ЁЃГЌЩљв§ЗЂЕФЫЎФ§НКЪЧгЩКЃдхЫсбЮЕФєШЛљгыИЦРызгХфЮЛаЮГЩЕФ[11]ЃЌГЌЩљДІРэЦЦЛЕСЫИЦРызггыєШЛљжЎМфЕФХфЮЛЃЌЕМжТЫЎФ§НКНЕНтЃЌДгЖјДйНјСЫЗтзАФЩУзЯЫЮЌЕФЪЭЗХ[12]ЁЃЭЌЪБЃЌUPN@ЫЎФ§НКзїЮЊЮШЖЈЁЂПЩБфаЮЕФШ§ЮЌжЇМмЬюГфЙЧШБЫ№ЧјгђЃЌЭЈЙ§ЛюадГЌЖЬыФФЩУзЯЫЮЌЕФПижЦЪЭЗХЃЌдіЧПГЩЙЧУтвпЮЂЛЗОГЃЌДйНјЙЧаоИДЃЌЮЊСйДВЙЧШБЫ№аоИДЬсЙЉвЛжжаТВпТдЁЃ
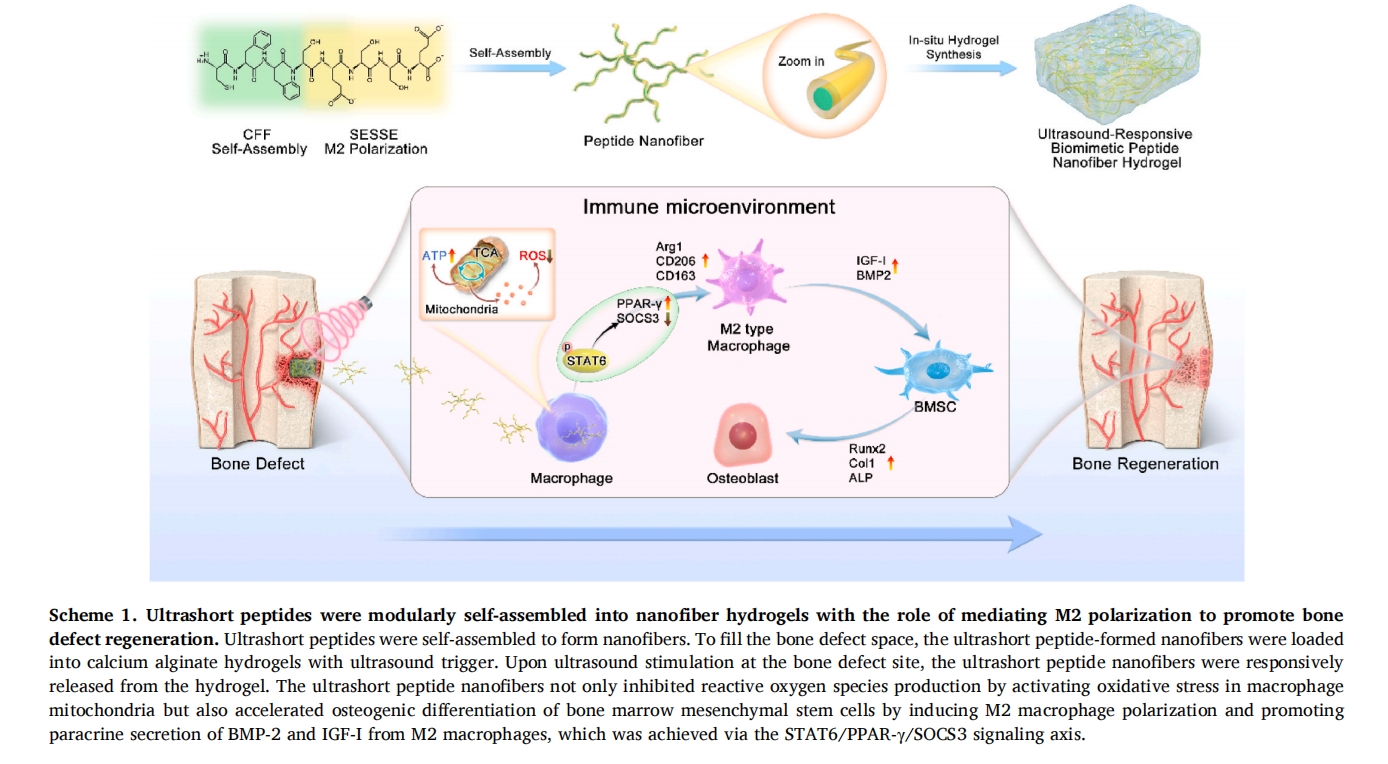
2 НсЙћ
2.1 UPN@ЫЎФ§НКЕФКЯГЩМАБэеї
ЪзЯШЃЌЭЈЙ§ CFF-SESSE здзщзАжЦБИыФФЩУзЯЫЮЌЃЌШчЭМЫљЪОЗНАИ 1. ШчЭМЫљЪОЭМ 1
дк SEMЃЈЩЈУшЕчзгЯдЮЂОЕЃЉЭМЯёжаЙлВьЕННЛСЊФЩУзЯЫЮЌЭјТчЁЃШЛКѓЃЌКЯГЩСЫКЃдхЫсИЦЫЎФ§НКОЭЕигыФЩУзЯЫЮЌвЛЦ№жЦГЩUPN@ЫЎФ§НКЁЃЮЊСЫбщжЄДЫРрыФФЩУзЯЫЮЌЫЎФ§НКЕФГЩЙІКЯГЩЃЌЬсЧАжЦзїСЫFITCБъМЧЕФыФФЩУзЯЫЮЌЁЃШчЭМЫљЪОЭМ 1B жаЙлВьЕНТЬЩЋгЋЙтЕФОљдШЗжВМЃЌжЄЪЕСЫыФФЩУзЯЫЮЌдкЫЎФ§НКФкЕФОљдШЗжВМЁЃДЫЭтЃЌFITR НсЙћЃЈЭМ S1) ЕФЮќЪеЗхДг 1495.5 cm−1вЦЕНСЫ 1485.8 cm−1ЃЌБэУїКЃдхЫсбЮЕФєШЛљгыИЦРызгЗЂЩњСЫХфЮЛЃЌДгЖјбщжЄСЫКЃдхЫсИЦЫЎФ§НКЕФаЮГЩ[13].ЭЌЪБЃЌыФФЩУзЯЫЮЌдкЫЎФ§НКжаЕФИКдиаЇТЪКЭИКдиСПЗжБ№ЮЊ45.2ЁР1.4%КЭ10.5ЁР0.5%ЃЈЭМ S2)ЁЃДЫЭтЃЌЛЙгІгУСїБфбЇЗжЮіРДШЗШЯЫЎФ§НКЕФСїБфадФмЃЌНсЙћЯдЪОДЂФмФЃСП (G') КЭЫ№КФФЃСП (GЁБ) жЕЖМКмИпЃЌБэУїМсгВЕФЫЎФ§НКЪЪКЯгУгкЙЧШБЫ№ЬюГф (ЭМ S3ЃЉВЂВтСПСЫВЛЭЌКЌСПыФФЩУзЯЫЮЌЫЎФ§НКЕФгІСІ-гІБфЧњЯпЃЈЭМ S4)ЃЌБэУїИКдиыФФЩУзЯЫЮЌЕФЫЎФ§НКЕФПЙбЙЧПЖШгыКЃдхЫсИЦЫЎФ§НКЯрБШНЯШѕЃЌвВЪЪКЯЬюГфЙЧШБЫ№ВПЮЛЁЃ
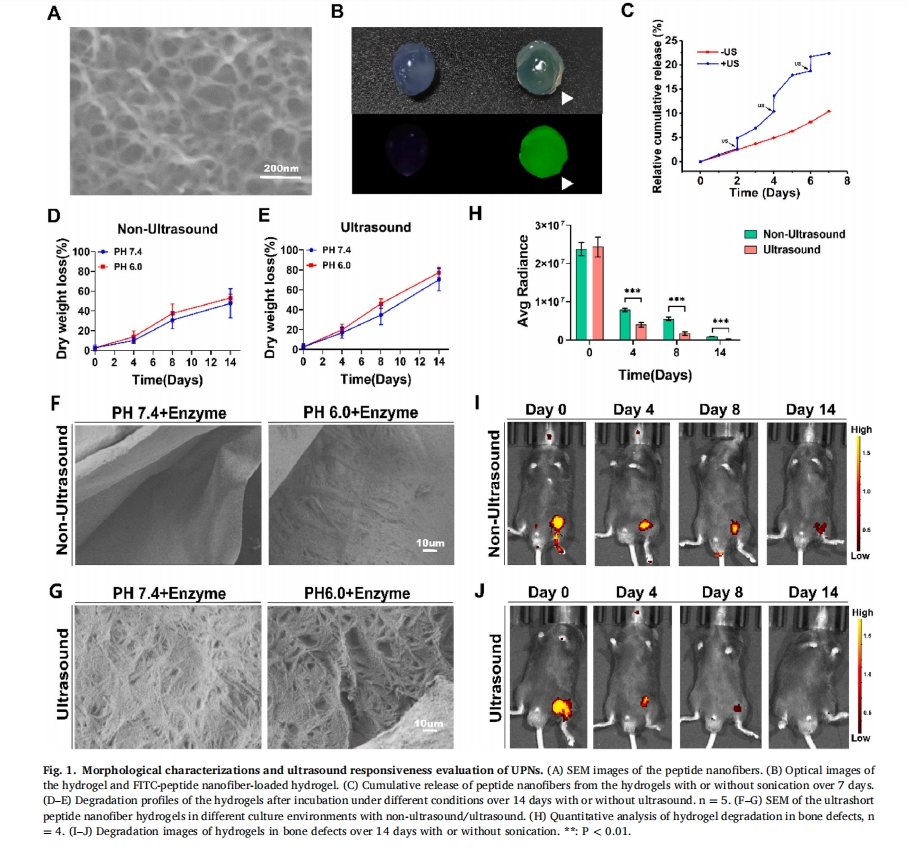
2.2 ЦРЙР UPN@ЫЎФ§НКЕФГЌЩљЯьгІад
ЪзЯШбаОПСЫВЛЭЌГЌЩљЧПЖШЯТЫЎФ§НКЕФНЕНтааЮЊЁЃДЫЭтЃЌЛЙЙлВьЕНГЌЩљв§ЗЂUPN@ЫЎФ§НКЕФМгЫйЪЭЗХЃЌШчЭМЫљЪОЭМ 1C. жежЙГЌЩљДЬМЄКѓЃЌЫЎФ§НКМЬајГжајЪЭЗХыФФЩУзЯЫЮЌЁЃДЫЭтЃЌНЋUPN@ЫЎФ§НК НўШыPBSШмвКЃЈpH 6.0КЭ7.4ЃЉжавдФЃФтЙЧШБЫ№ЧјгђЕФЫсадЮЂЛЗОГЃЌГЌЩљЗДгІШдШЛМгЫйЫЎФ§НКжиСПЕФЫ№ЪЇЃЈЭМ1DКЭEЃЉЁЃДЫЭтЃЌЛЙгІгУСЫИїжжГЌЩљВЈВЮЪ§ЃКВЛНјааГЌЩљДІРэЁЂ1 W/cm2ЁЂ1.75 W/cm2КЭ 2.5 W/cm2ГЌЩљДІРэ 2 ЗжжгЁЃНсЙћБэУїЃЌЫцзХГЌЩљЧПЖШЕФдіМгЃЌЫЎФ§НКЕФНЕНтЫйЖШвВМгПьЃЈЭМS5ЃЉЭЌбљЃЌдкЪЉМгГЌЩљВЈКѓЙлВьЕНСЫИќЖрЕФ UPN@ЫЎФ§НК ПеЧЛЃЈЭМ1FКЭG)ЁЃДЫЭтЃЌКЌгаКЭВЛКЌвШЕААзУИЕФЫЎФ§НКжЎМфУЛгаЯджјВювьЃЌБэУїЖдвШЕААзУИОпгаЫљашЕФЮШЖЈадЃЈЭМ S6ЃЉЁЃГ§СЫЬхЭтбЇЯАЃЌЬхФкЫЎФ§НКдкЙЧШБЫ№жаЕФНЕНтЖЏСІбЇЃЈЭМ 1ЮвЃЉЛђгыЃЈЭМ 1J) ЛЙЭЈЙ§ШЋЩэгЋЙтГЩЯёбаОПСЫГЌЩљЃЌЖЈСПБэУїГЌЩљЯьгІМгЫйСЫЫЎФ§НКЕФНЕНтЃЈЭМ 1ЃЈHЃЉЁЃ
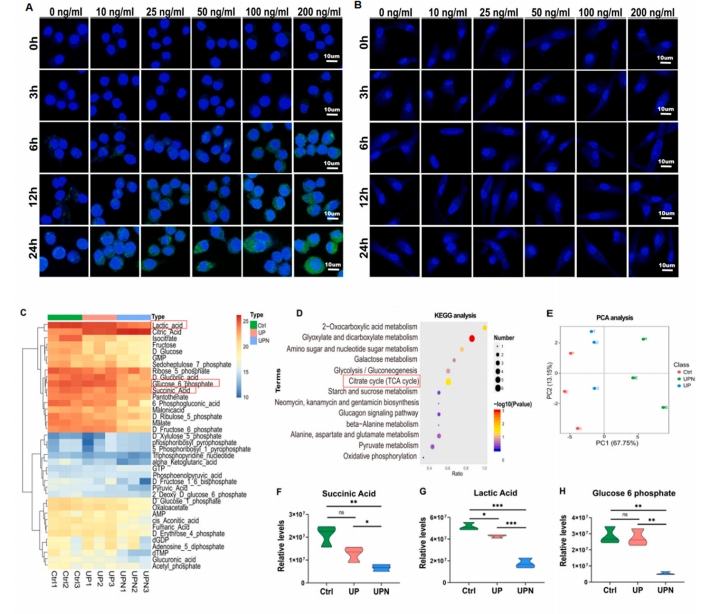
2.3 UPN ЕїНкОоЪЩЯИАћЯпСЃЬхДњаЛКЭбѕЛЏгІМЄ
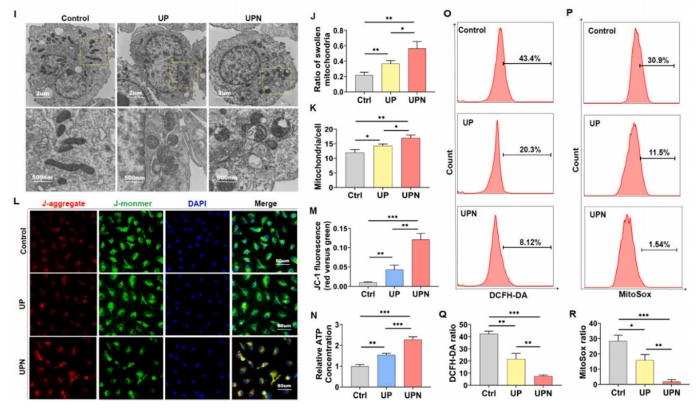
ЪзДЮРћгУУтвпгЋЙтЗЈбаОПСЫОоЪЩЯИАћЖдГЌЖЬыФФЩУзЯЫЮЌЕФЭЬЪЩзїгУЃЌНсЙћБэУїЃЌUPNдк6 hЪБПЊЪМБЛОоЪЩЯИАћЭЬЪЩЃЌЫцзХЗѕг§ЪБМфЕФбгГЄКЭUPNДЬМЄХЈЖШЕФдіМгЃЌUPNБЛОоЪЩЯИАћЭЬЪЩЕФЯжЯѓШевцУїЯдЃЈЭМ 2A).ШЛЖјЃЌГЌЖЬыФФЩУзЯЫЮЌВЂУЛгаБЛBMSCsУїЯдЭЬЪЩЃЈЭМ 2B). ЯпСЃЬхДњаЛЪЧОоЪЩЯИАћЙІФмзЊЛЏЕФЙиМќПижЦЦїЃЌВЂОіЖЈЦфМЋЛЏзДЬЌ[14]ЁЃОоЪЩЯИАћДњаЛзщбЇЗжЮіЯдЪОЃЌгыUPNХрбјЕФОоЪЩЯИАћгыЬЧНЭНтЃЈEMPЃЉКЭШ§єШЫсбЛЗЃЈTCAЃЉУмЧаЯрЙиЃЌЬсЪОUPNПЩвдМЄЛюОоЪЩЯИАћЕФЬЧНЭНтКЭШ§єШЫсбЛЗЃЈЭМ 2CЈCEЃЉЁЃTCAбЛЗжачњчъЫсЕФЖЈСПЗжЮіБэУїUPNвжжЦСЫчњчъЫсДњаЛЃЈЭМ 2F). ЭЌЪБЃЌЖдEMPДњаЛжаШщЫсКЭЦЯЬбЬЧ-6СзЫсЕФЖЈСПЗжЮіБэУїЃЌUPNНЕЕЭСЫШщЫсЕФВњЩњЃЈЭМ 2G) КЭОоЪЩЯИАћжаЕФЦЯЬбЬЧ 6 СзЫс (ЭМ 2H).ЭИЩфЕчОЕМьВтОоЪЩЯИАћФкЯпСЃЬхЕФаЮЬЌЃЌНсЙћЯдЪОUPNsгеЕМЯпСЃЬхЕФаЮЬЌгЩдРДЕФАєЧђзДБфЮЊХђеЭЕФЭждВаЮЃЈЭМ 2I).НЋUPsКЭUPNЗѕг§ОоЪЩЯИАћКѓЗЂЯжЃЌUPNзщЯИАћЯпСЃЬхжзеЭБШР§ЮЊ53%ЃЌUPзщЯИАћЯпСЃЬхжзеЭБШР§ЮЊ38%ЃЌCtrlзщЯИАћЯпСЃЬхжзеЭБШР§НгНќ20%ЃЌЧвUPNзщЯИАћЯпСЃЬхжзеЭБШР§НЯCtrlзщНјвЛВНдіМгЃЈЭМ 2J).ДЫЭтЃЌЯпСЃЬх/ЯИАћБШР§НсЙћЯдЪОЃЌUPNзщгыCtrlзщЯрБШБШР§НјвЛВНЩ§Ип(ЭМ 2K)ЁЃЭЈЙ§ЯпСЃЬхФЄЕчЮЛКЭЛюадбѕ(Reactive Oxygen Species, ROS)ЪЭЗХЗжЮіUPNЖдОоЪЩЯИАћЯпСЃЬхНсЙЙЕФгАЯьЁЃгУJC-1МьВтЯпСЃЬхФЄЕчЮЛJC-1ЁЃУтвпгЋЙтНсЙћЯдЪОUPNПЩЪЙJC-1ОлКЯЮяЕФБэДяСПгЩЖдеезщЕФ0.02гЋЙтЧПЖШдіжСUPNзщЕФ0.13(ЭМ 2LM)ЁЃATPМьВтЯдЪОЃЌUPNsДйНјСЫОоЪЩЯИАћЯпСЃЬхШ§єШЫсбЛЗЃЌБэУїОоЪЩЯИАћФмСПДњаЛБЛМЄЛю(ЭМ 2N). СїЪНЯИАћЪѕЗжЮіжаЃЌОоЪЩЯИАћФкзмROSгУDCGH-DAБъМЧЃЈЭМ 2O)ЃЌВЂгУ MitoSox (ЭМ 2P)ЁЃНсЙћБэУїЃЌUPN ВЛНіФмЙЛНЕЕЭОоЪЩЯИАћжазм ROS ЕФЪЭЗХЃЌЖјЧвЛЙФмЙЛНЕЕЭОоЪЩЯИАћЯпСЃЬхжа ROS ЕФЪЭЗХЃЈЭМ 2QR)ЁЃзмжЎЃЌUPNПЩвдЕїНкЯпСЃЬхДњаЛЃЌдіЧПMMPЛюадЃЌМѕЩйROSЪЭЗХЁЃ

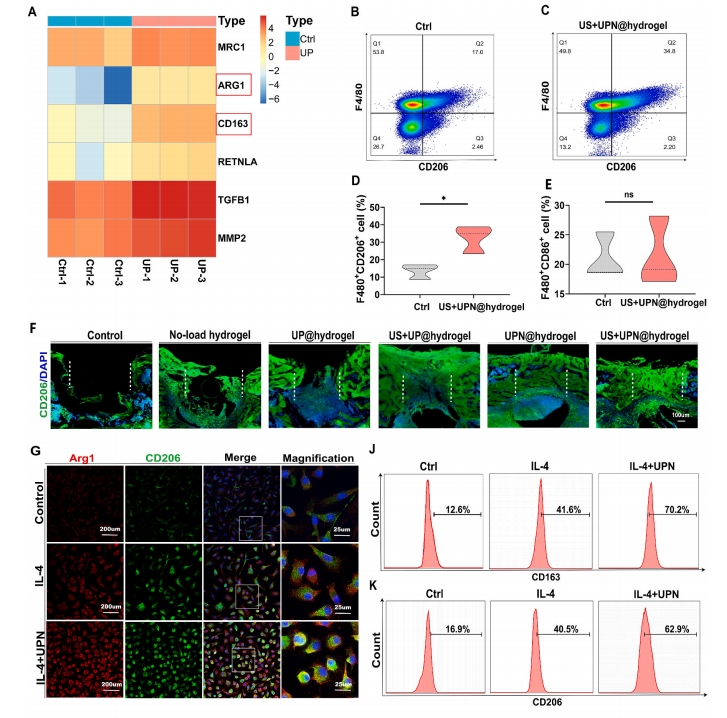
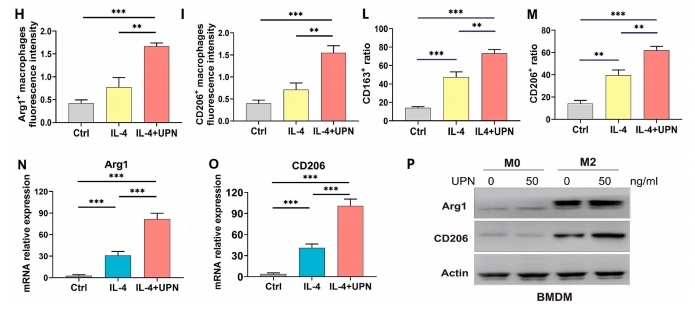
2.4 UPN ДйНј M2 аЭОоЪЩЯИАћМЋЛЏ
ОнБЈЕРЃЌM1ДйбзОоЪЩЯИАћКЭM2аоИДОоЪЩЯИАћЛсгАЯьЫ№ЩЫКѓЕФзщжЏдйЩњЁЃ15]ЁЃArg1ЁЂCD206КЭCD163ЪЧM2аЭОоЪЩЯИАћБэДяЕФЛљвђЁЃЖдгУUPsжЮСЦ14ЬьЕФаЁЪѓЙЧШБЫ№НјааЛљвђзЊТМВтађЗжЮіЯдЪОЃЌArg1КЭCD163ЕФБэДяЯджјдіМгЃЈЭМ3A).гУГЌЖЬыФФЩУзЯЫЮЌЫЎФ§НКЃЈМЧЮЊUS+UPN@hydrogelЃЉдкЙЧШБЫ№ЧјгђНјаа14ЬьЕФГЌЩљДІРэКѓЃЌM2аЭОоЪЩЯИАћБЛF4/80КЭCD206БъМЧЃЈЭМ3BКЭCЃЉЁЃСїЪНЯИАћЪѕЯдЪОCD206БъМЧЕФM2аЭОоЪЩЯИАћЪ§СПеМОоЪЩЯИАћзмЪ§ЕФАйЗжБШдіМгЃЈЭМ3D)ЃЌЖјCD86БъМЧЕФM1аЭОоЪЩЯИАћЮоУїЯдВювьЃЈЭМ3E).УтвпгЋЙтНсЙћЯдЪОUS+UPN@hydrogelФмЙЛдіМгЙЧШБЫ№ДІCD206ЕФБэДя(ЭМ3F).ЮЊСЫНвЪОЗТЩњыФФЩУзЯЫЮЌгыОоЪЩЯИАћжЎМфЕФЯрЛЅзїгУЃЌЮвУЧНЋIL-4гыBMDMвЛЦ№Зѕг§ЃЌвдбаОПM2аЭОоЪЩЯИАћЕФМЋЛЏЬхЭтУтвпгЋЙтЯдЪОЃЌUPNФмЙЛНјвЛВНДйНјIL-4НщЕМЕФM2аЭОоЪЩЯИАћБъжОЮяArg1КЭCD206ЕФБэДяЃЈЭМ3GЈCI).СїЪНЯИАћЪѕМьВтCD163КЭCD206ЕФБэДяЃЈЭМ3JКЭK)ЃЌНсЙћЯдЪОЃЌIL-4+UPNзщCD163БэДяТЪЮЊ70.6%ЃЌIL-4зщЮЊ50.3%ЃЌCtrlзщдМЮЊ17.2%ЃЈЭМ3L)ЃЌIL-4+UPNзщCD206БэДяТЪЮЊ61.8%ЃЌIL-4зщЮЊ40.9%ЃЌCtrlзщдМЮЊ18.1%ЃЈЭМ3M).RT-qPCRНсЙћБэУїUPNsЯджјдіМгСЫArg1КЭCD206ЕФБэДя(ЭМ3ЭЌбљЃЌWesternгЁМЃНсЙћЯдЪОUPNsУїЯдДйНјArg1КЭCD206ЕААзБэДяЃЈЭМ3P).злЩЯЫљЪі,БОбаОПЬсЪОUPN@ЫЎФ§НКжЮСЦКѓЙЧШБЫ№аоИДЙ§ГЬжа,M2аоИДОоЪЩЯИАћЦ№жїЕМзїгУ,ЖјГЌЩљДЅЗЂНјвЛВНДйНјСЫM2аЭОоЪЩЯИАћЕФМЋЛЏЁЃ

2.5 ЭЈЙ§upnЕїПиSTAT6/PPAR-ІУ/SOCS3жсгеЕМm2аЭОоЪЩЯИАћЕФМЋЛЏ
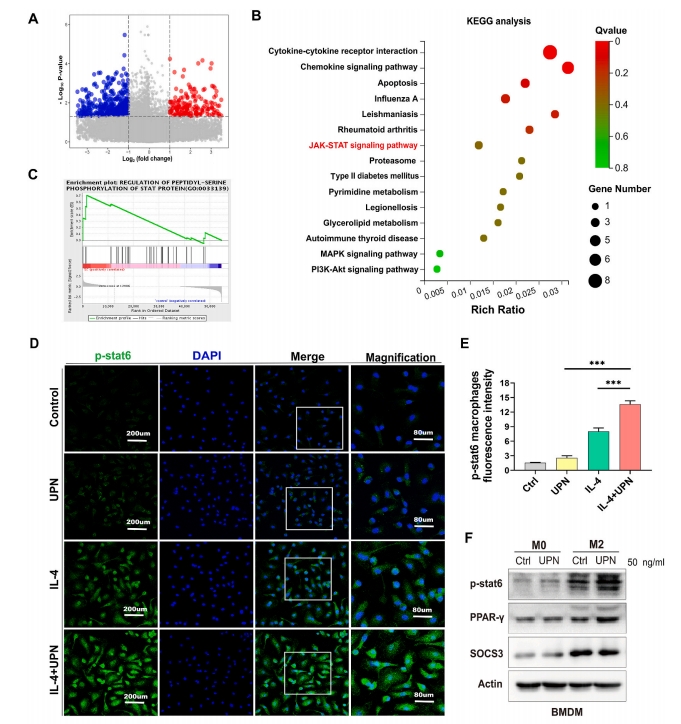
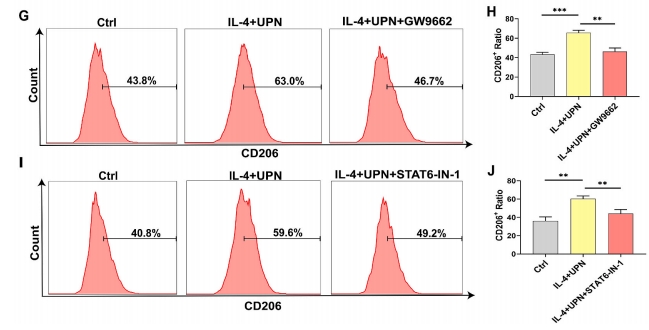
баОПЭХЖгЕФГѕВНбаОПНсЙћЯдЪОЃЌаЁЪѓЙЧШБЫ№ВПЮЛЭЈЙ§ЮВОВТізЂЩфUPs 14ЬьНјаажЮСЦЃЌЭЈЙ§ЛљвђБОЬхТлЗжЮіЁЂОЉЖМЛљвђгыЛљвђзщАйПЦШЋЪщЗжЮіКЭЛљвђМЏИЛМЏЗжЮіЗЂЯжЃЌUPsгыJAK/STATаХКХЭЈТЗУмЧаЯрЙиЃЈЭМ 4AЈCC). ЮЊжЄЪЕUPNгеЕМM2аЭОоЪЩЯИАћМЋЛЏЕФЗжзгЛњжЦЃЌЮвУЧгУIL-4КЭUPNгеЕМM2аЭОоЪЩЯИАћ24аЁЪБЃЌгыCtrlзщЯрБШЃЌЦфp-STAT6гЋЙтЧПЖШЯджјдіМгЃЈЭМ 4D).АыЖЈСПЗжЮіЯдЪОЃЌIL-4+UPNзщЃЈ13.5%ЃЉМгШыUPNКѓЃЌp-STAT6гЋЙтЧПЖШНЯIL-4зщЃЈ7.8%ЃЉКЭCtrlзщЃЈ1.7%ЃЉУїЯддіЧПЃЈЭМ 4E).Western blottingНсЙћжЄЪЕЃЌдкM2аЭОоЪЩЯИАћжаМгШыUPNКѓЃЌp-STAT6КЭPPAR-ІУЕААзЫЎЦНЩЯЩ§ЃЌSOCS3ЕААзЫЎЦНЯТНЕЃЌЬсЪОUPNЭЈЙ§діЧПSTAT6КЭPPAR-ІУЕААзНјвЛВНДйНјM2аЭОоЪЩЯИАћМЋЛЏЃЌЖјUPNНщЕМSOCS3ЕААзЯТНЕЃЌДгЖјДйНјM2аЭОоЪЩЯИАћМЋЛЏЃЈЭМ 4FЃЉIL-4+UPNзщp-STAT6КЭPPAR-ІУЕААзХЈЖШУїЯдЩ§ИпЃЌSOCS3ЕААзХЈЖШУїЯдНЕЕЭЃЌЖјUPNзщгыCtrlзщжЎМфSOCS3ЁЂp-STAT6КЭPPAR-ІУБэДяЮоУїЯдВювьЁЃвдЩЯНсЙћЬсЪОUPNЭЈЙ§STAT6/PPAR-ІУ/SOCS3аХКХжсНјвЛВНгеЕМM2аЭОоЪЩЯИАћМЋЛЏЃЈЭМ 4F).СїЪНЯИАћЪѕНсЙћЯдЪОЃЌGW9662ЃЈPPAR-ІУвжжЦМСЃЉвжжЦСЫUPNгеЕМЕФM2аЭОоЪЩЯИАћБъжОЮяCD206ЕФБэДяЃЈЭМ 4G).СїЪНЯИАћЪѕНсЙћЯдЪОSTAT6-IN-1ЃЈSTAT6вжжЦМСЃЉвжжЦUPNгеЕМЕФM2аЭОоЪЩЯИАћБъжОЮяCD206ЕФБэДяЃЈЭМ 4ЃЈHЃЉЁЃ

2.6 UPNДйНјBMSCsГЩЙЧЗжЛЏКЭЧЈвЦ
BMSCs ЪЧФмЙЛаоИДЙЧЫ№ЩЫЕФжжзгЯИАћ [16] баОПБэУїЃЌОоЪЩЯИАћгЩДйбзадM1бЧаЭЯђПЙбзадM2бЧаЭЕФзЊБфдкЙЧЫ№ЩЫаоИДЙ§ГЬжаЦ№зХжСЙиживЊЕФзїгУЃЌвђЮЊОоЪЩЯИАћЗжУкЕФЯИАћвђзгЖдИЩЯИАћЕФФММЏЁЂдіжГКЭЗжЛЏгаЯджјЕФгАЯьЁЃ[17]ЁЃЮЊСЫНјвЛВНбщжЄM2ОоЪЩЯИАћХдЗжУкЩњГЄвђзгЪЧЗёгАЯьBMSCЕФааЮЊКЭУќдЫЃЌЮвУЧЪЙгУЙВХрбјЯЕЭГРДЦРЙРBMDMКЭBMSCжЎМфЕФЯрЛЅзїгУЁЃUPNДІРэЕФBMDMКЭBMSCЙВХрбјЕФЪОвтЭМШчЭМЫљЪОЃЈЭМ 5A).МюадСзЫсУИШОЩЋМьВтBMSCsГЩЙЧПѓЛЏЧщПіЃЌЗЂЯжIL-4+UPNзщBMSCМюадСзЫсУИЛюаддкЕк7ЬьНЯCtrlзщУїЯдЩ§ИпЃЈЭМ 5B КЭ C) КЭ 14 (ЭМ 5DКЭE)ЃЌмчЫиКьШОЩЋНсЙћЯдЪОЃЌЙВХрбјЬхЯЕжаЕк21ЬьIL-4+UPNзщBMSCsПѓЛЏНсНкЪ§СПНЯCtrlзщдіМг(ЭМ 5FЁЂGЃЉЩЯЪіНсЙћжЄУїЃЌГЌЖЬыФФЩУзЯЫЮЌПЩвдНјвЛВНгеЕМОоЪЩЯИАћЯђM2бЧаЭМЋЛЏЃЌвђДЫЃЌUPNгеЕМM2ОоЪЩЯИАћМЋЛЏКѓЃЌM2ОоЪЩЯИАћЩњГЄвђзгЕФХдЗжУкзїгУДйНјСЫBMSCsЕФГЩЙЧЗжЛЏЁЃ
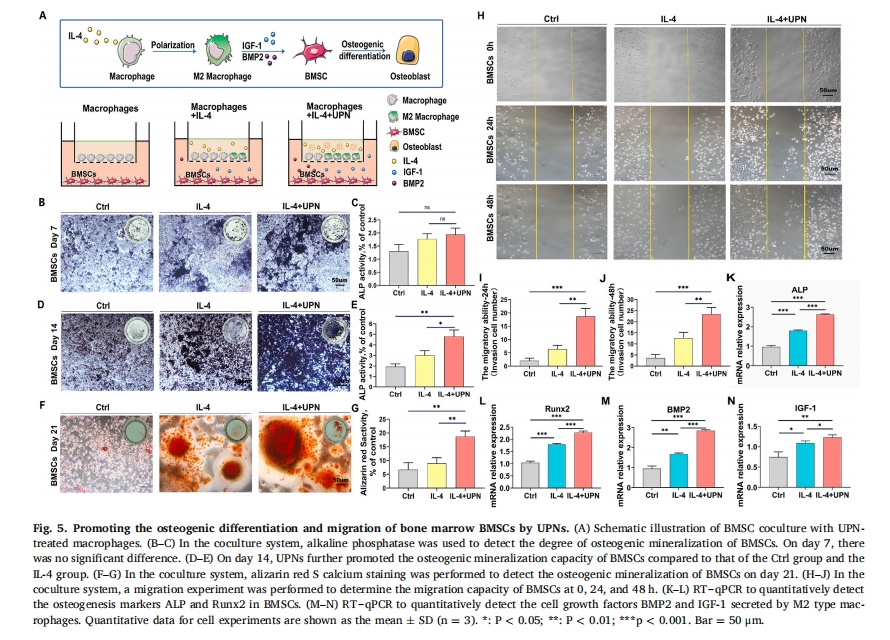
ЮЊСЫНјвЛВНЬНЬжM2бЧаЭОоЪЩЯИАћЖдBMSCsЕФХдЗжУкзїгУЃЌЮвУЧдкЙВХрбјЬхЯЕжаЦРЙРСЫBMSCsЕФЧЈвЦФмСІЁЃTranswellЧЈвЦЪЕбщЯдЪОЃЌ0 hЪБCtrlзщЁЂIL-4зщКЭIL-4+UPNзщBMSCsжЎМфЮоУїЯдВювьЃЈЭМ 5H). 24 hЪБIL-4+UPNзщЧЈвЦЯИАћБШР§ЮЊ18.5%, IL-4зщЧЈвЦЯИАћБШР§ЮЊ7.2%, ЖдеезщЧЈвЦЯИАћБШР§НгНќ2% (ЭМ 5I). IL-4+UPNзщЃЈ23.6%ЃЉМгШыUPNКѓЃЌ48 hЪБЧЈвЦЯИАћЪ§ЯджјИпгкIL-4зщЃЈ13.7%ЃЉКЭЖдеезщЃЈ4.8%ЃЉЃЈЭМ 5J).вдЩЯНсЙћЬсЪОUPNsФмгааЇДйНјBMSCsЕФЧЈвЦЃЌRT-qPCRМьВтBMSCsжаГЩЙЧЯрЙиЛљвђЃЌАќРЈМюадСзЫсУИЃЈALPЃЉКЭRuntЯрЙизЊТМвђзг2ЃЈRunx2ЃЉЃЌНсЙћЯдЪОГЌЖЬыФФЩУзЯЫЮЌФмЯджјДйНјBMSCsЕФГЩЙЧЗжЛЏЃЈЭМ 5KL)ЁЃДЫЭтЃЌЙЧаЮЬЌЗЂЩњЕААз-2 (BMP-2) КЭвШЕКЫибљЩњГЄвђзг (IGF-1) вВЪЧОоЪЩЯИАћЗжУкЕФЩњГЄвђзг[18ЮвУЧМйЩш UPN ПЩвдЭЈЙ§ХдЗжУк IGF-1 ЩњГЄвђзгзїгУгк BMSCsЃЈЭМ 5M−NЃЉЁЃ
2.7 ГЌЩљЯьгІUPN@ЫЎФ§НКдкЬхФкЙЧШБЫ№жЮСЦжаЕФзїгУ
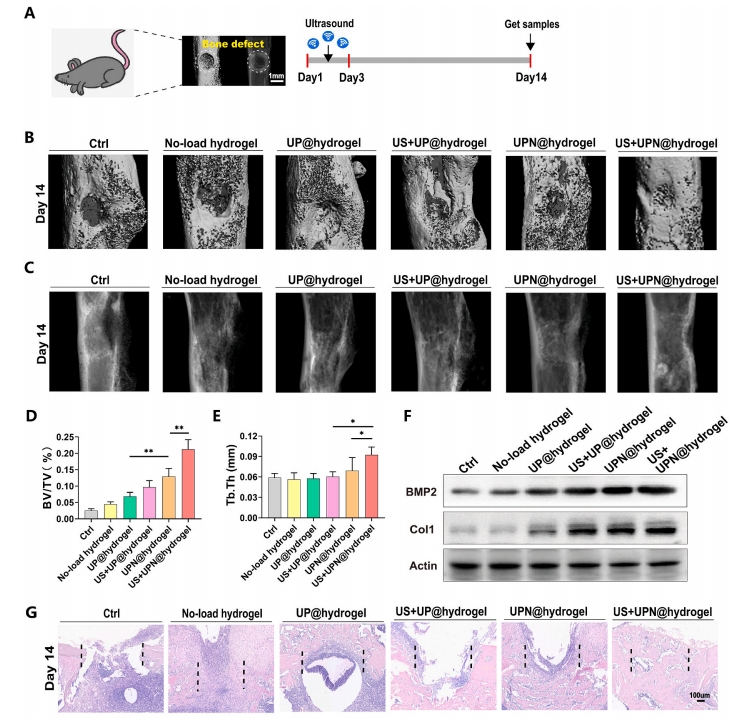
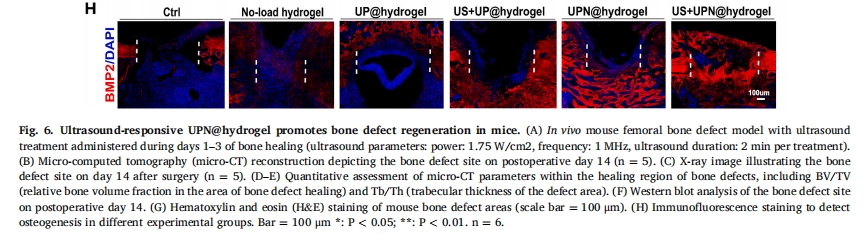
НЈСЂSPFМЖ8жмСфC57BL/6JаЁЪѓЙЧШБЫ№ФЃаЭЃЌВЩгУГЌЩљДЅЗЂUPN@ЫЎФ§НКЃЈЭМ 6A). ЮЊСЫжБЙлЙлВьгњКЯЙ§ГЬЃЌВЩгУmicro-CTГЩЯёМьВщВЛЭЌзщЙЧШБЫ№ЕФгњКЯаЇЙћЁЃЪѕКѓ14ЬьЃЌmicro-CTжиНЈЭМЯёЃЈЭМ 6B) ЙЧШБЫ№ВПЮЛЃЌвдМА X ЩфЯпЭМЯёЃЈЭМ 6C)ЃЌЭЈЙ§Micro-CTЩЈУшЖЈСПЗжЮіЙЧШБЫ№гњКЯЧјЙЧЬхЛ§/зщжЏЬхЛ§(BV/TV)МАЙЧаЁСККёЖШ(Tb.Th)ЃЌНсЙћЯдЪОUS+UPN@hydrogelзщЙЧгњКЯаЇЙћНЯКУ(ЭМ 6D КЭ EЃЉЁЃДЫЭтЃЌДгЙЧШБЫ№ВПЮЛЕФзщжЏжаЬсШЁЕААзжЪВЂНјааЕААзжЪгЁМЃЗжЮіБэУїЃЌUS + UPN@ ЫЎФ§НКзщЕФГЩЙЧЕААзБъжОЮя BMP2 КЭ Col1 КЌСПзюИпЃЈЭМ 6F). H&EШОЩЋЦРМлаЁЪѓЙЧШБЫ№ЕФаоИДФмСІЃЌНсЙћЯдЪОаЁЪѓЙЧШБЫ№ЧјгђаТЩњЙЧаЁСКж№НЅБЛаТЩњЙЧЬцДњЃЈЭМ 6G).ГЩЙЧЛљвђБъжОЮяBMP2УтвпгЋЙтМьВтГЪКьЩЋгЋЙтЃЌЧвUS+UPN@hydrogelзщгЋЙтЧПЖШзюЧПгкCtrlзщЃЈЭМ 6H).вдЩЯНсЙћжЄЪЕСЫгІгУГЌЩљв§ЗЂЕФГЌЖЬыФФЩУзЯЫЮЌЫЎФ§НКВФСЯПЩвдгааЇИФЩЦЙЧШБЫ№ЕФдйЩњЁЃ
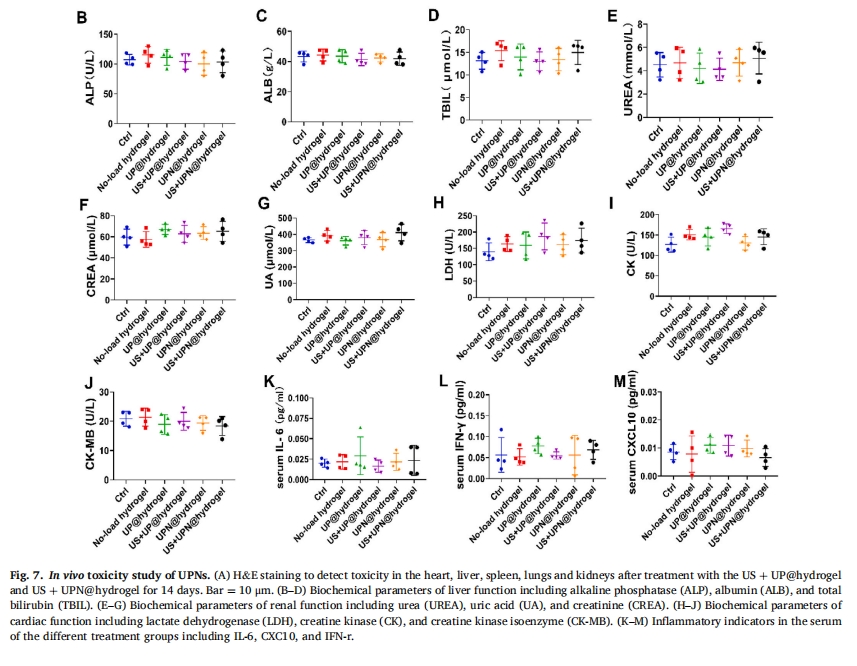
2.8 UPN@hydrogelЕФЬхФкЖОадбаОП
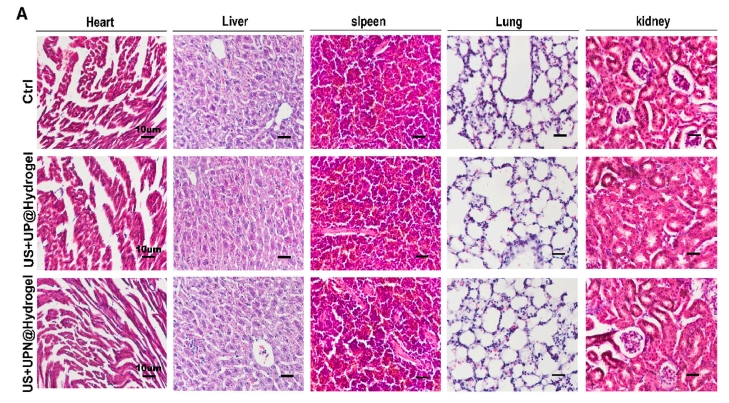
БОбаОПНЋUP@hydrogelЃЈМЧЮЊUS+UP@hydrogelЃЉКЭUPN@hydrogelЃЈМЧЮЊUS+UPN@hydrogelЃЉОжВПжВШыаЁЪѓЙЧШБЫ№ВПЮЛЁЃUP@hydrogelЮЊЕЅЬхыФЫЎФ§НКЃЌUPN@hydrogelЮЊзщзАыФФЩУзЯЫЮЌЫЎФ§НКЁЃРћгУГЌЩљДЬМЄв§ЗЂЫЎФ§НКЕФжЮСЦЗДгІЁЃжЮСЦ14ЬьКѓЃЌгУH&EШОЩЋМьВщаФдрЁЂИЮдрЁЂЦЂдрЁЂЗЮдрКЭЩідрЕФвЉЮяЖОадЃЌНсЙћЯдЪОетаЉЦїЙйУЛгабзжЂНўШѓЛђВЁРэИФБфЃЈЭМ 7A). дкЮвУЧЕФбаОПжаЃЌЮвУЧНјааСЫЬхФкЦРЙРвдЦРЙР UPN@hydrogel ЕФАВШЋадЁЃзаЯИЬсШЁбЊЧхбљБОЃЌвдбаОПИЮдрЁЂЩідрКЭаФдрЙІФмвдМАбзжЂЗДгІЕФЧБдкБфЛЏЁЃЮЊСЫШЋУцЗжЮі UPN@hydrogel ЕФАВШЋадЃЌЮвУЧЖдЬсШЁЕФбЊЧхбљБОНјааСЫГЙЕзЕФЦРЙРЁЃЮвУЧЕФжиЕуЪЧМрВт UPN@hydrogel в§Шыв§Ц№ЕФИЮдрЁЂЩідрКЭаФдрЙІФмЕФШЮКЮБфЛЏЁЃДЫЭтЃЌЮвУЧЛЙзаЯИбаОПСЫЫЎФ§НКгІгУв§Ц№ЕФбзжЂЗДгІЁЃЮвУЧВтЪдСЫИЮЙІФмжИБъЃЌАќРЈМюадСзЫсУИ (ALP)ЁЂАзЕААз (ALB) КЭзмЕЈКьЫи (TBIL) (ЭМ 7BЈCDЃЉЁЃЩіЙІФмжИБъАќРЈФђЫиЃЈUREAЃЉЁЂФђЫсЃЈUAЃЉЁЂМЁєћЃЈCREAЃЉЃЈЭМ 7EЈCGЃЉЁЃаФдрЙІФмжИБъАќРЈШщЫсЭбЧтУИЃЈLDHЃЉЁЂМЁЫсМЄУИЃЈCKЃЉЁЂМЁЫсМЄУИЭЌЙЄУИЃЈCK-MBЃЉЃЈЭМ 7HЈCJ)ЁЃНсЙћЯдЪОжЮСЦКѓИЮ/Щі/аФдрЙІФмдке§ГЃЗЖЮЇФкЁЃЮвУЧМьВтЕФбзжЂвђзгАќРЈIL-6ЁЂCXC10ЁЂIFN-rЃЈЭМ 7KM)ЁЃЯдЮЂОЕЯТЙлВь800 ІЬg/ml UPКЭUPNзїгУ0ЁЂ48ЁЂ72 hЪБОоЪЩЯИАћЕФЛюадЧщПі,ПЩМћЯИАћзДЬЌСМКУ,ЫЕУїUPКЭUPNЮоУїЯдЖОад(ЭМ S7 AC),ЫцКѓВЩгУCCK-8ЗЈМьВт0~800 ІЬg/ml UPN@hydrogelДІРэBMDMsКЭBMSCs 0ЁЂ48ЁЂ72 hКѓЕФЛюСІ,НсЙћЯдЪОUPNЖдЯИАћЛюадУЛгагАЯь(ЭМ S7 DFЃЉЁЃ
3 ЬжТл
МЬајЬНЫїФмЙЛИФБфОоЪЩЯИАћМЋЛЏзДЬЌЁЂНЈСЂгаРћгкЙЧдйЩњЕФУтвпЮЂЛЗОГЕФУтвпЕїНкЗжзгКЭЩњЮяВФСЯжСЙиживЊЁЃ[19]ДЫЭтЃЌзюНќЕФбаОПБэУїЃЌM2аЭОоЪЩЯИАћПЩвдЗжУкЯИАћвђзгКЭЭтУкЬхЃЌАяжњЕїНкЙЧЫшМфГфжЪИЩЯИАћЕФГЩЙЧЗжЛЏЁЃ[20]ЁЃИФЩЦДйНј BMSCs ГЩЙЧЗжЛЏЕФУтвпЮЂЛЗОГЖдгкЙЧШБЫ№ЕФаоИДжСЙиживЊЁЃECM ЕААздкГЩЙЧЯИАћЗжЛЏКЭЙЧдйЩњжаЦ№зХжСЙиживЊЕФзїгУ[21]бРБОжЪЛљжЪЕААз 1 (DMP1) ЪЧвЛжждкЙЧїРжаИпЖШБэДяЕФЙиМќ ECM ЕААзЃЌПЩЕїНкЙЧПѓЛЏ[22],дкDMP1ЛљвђЭЛБфЕФвХДЋаЁЪѓФЃаЭжаЃЌЛљвђзЊТМзщВтађЗжЮіНсЙћЯдЪОЃЌDMP1ЕФШБЪЇгыЙЧїРжаУтвпаХКХЭЈТЗЕФИФБфУмЧаЯрЙи[23]ЁЃгаШЄЕФЪЧЃЌDMP1АБЛљЫсађСаИЛКЌАБЛљЫсSЛђEЃЌS:EБШР§НгНќ3:2ЁЃЮЊСЫДйНјЙЧжЪЪшЫЩФЃаЭжаЕФBMSCЗжЛЏЃЌЮвУЧЩшМЦВЂКЯГЩСЫГЌЖЬыФSESSE[24]ЁЃзюНќЃЌЮвУЧЕФбаОПЗЂЯжСЫвЛИіживЊЗЂЯжЃЌМДDMP1МАЦфбмЩњыФЃЌАќРЈSESSEЃЌОпгаЕїНкM2аЭОоЪЩЯИАћМЋЛЏЕФФмСІЃЈЮДЗЂБэЪ§ОнЃЉЁЃШЛЖјЃЌSESSEыФЪЧЗёФмгааЇЕїНкУтвпЮЂЛЗОГДйНјЙЧдйЩњШдВЛЧхГўЁЃДЫЭтЃЌSESSEЕФЮШЖЈадКЭЪйУќгаЯоЃЌЖдЦфСйДВзЊЛЏНсЙћЙЙГЩЬєеНЁЃетаЉЗЂЯжЭЙЯдСЫНјвЛВНбаОПЕФБивЊадЃЌвдШЗЖЈSESSEдкДйНјЙЧдйЩњЗНУцЕФжЮСЦЧБСІЃЌВЂНтОіЮШЖЈадЮЪЬтЃЌвдГЩЙІгІгУгкСйДВЁЃ
дкетЯюбаОПжаЃЌЮвУЧПЊЗЂСЫвЛжжаТЗНЗЈРДдіЧПЬхФкВЩгУФЃПщЛЏГЌЗжзгздзщзАИХФюЃЌЬсИпГЌЖЬыФ (UP) ЕФЮШЖЈадЁЃОпЬхРДЫЕЃЌЮвУЧЩшМЦСЫвЛжжУћЮЊ SESSE ЕФыФЃЌЫќОпгаздзщзАФмСІЃЌПЩвдаЮГЩГЌЗжзгзщзАЬхЃЌМД UPNЃЈГЌЖЬыФФЩУзЯЫЮЌЃЉЁЃUPN ЕФаЮГЩЯджјЬсИпСЫ SESSE ыФдкЩњЮяЛЗОГжаЕФЮШЖЈадКЭЪйУќЁЃздзщзАыФФЩУзЯЫЮЌЕЅЬхАќКЌСНИіЙІФмгђЃКЃЈ1ЃЉздзщзАгђ CFFЃЌдДзд ІТ ЕэЗлбљыФЃЌПЩаЮГЩыФађСаВЂЬсЙЉНЛСЊЮЛЕуЃЛ[24] КЭЃЈ2ЃЉжЮСЦНсЙЙгђSESSEЁЃдкЫЎадЬѕМўЯТЃЌFFНсЙЙгђЕФЪшЫЎзїгУКЭІа-ІаЖбЛ§ЪЙыФЕЅЬхздзщзАГЩФЩУзЯЫЮЌЃЌЦфжаFFНсЙЙгђаЮГЩЪшЫЎКЫаФЃЌSESSEыФНсЙЙгђаЮГЩДјИКЕчКЩЕФЧзЫЎКЫаФ[25]ЁЃдкБОбаОПжаЃЌЮвУЧЕФжївЊФПБъЪЧДДдьвЛИігаРћЕФЮЂЛЗОГЃЌдкЙЧЫ№ЩЫЛжИДЕФдчЦкНзЖЮДйНјУтвпЗДгІЃЌДгЖјДйНјЙЧЫшМфГфжЪИЩЯИАћЃЈBMSCЃЉЕФЗжЛЏЁЃЮЊСЫЪЕЯжетвЛФПБъЃЌЮвУЧЬсГіРћгУГЌЩљПижЦЕФЫЎФ§НКЯЕЭГдкЫ№ЩЫЕФГѕЦкПижЦЪЭЗХИпХЈЖШЕФЩњЮяЛюадЙбыФЁЃБОжЪЩЯЃЌЮвУЧЕФбаОПЮЇШЦВћУїЖЬЦкУтвпЕїНкзїгУКЭИЩЯИАћЗжЛЏЬиадеЙПЊЁЃвђДЫЃЌБОбаОПжаЪЙгУЕФЫЎФ§НКВЛашвЊЯрЕБДѓЕФЛњаЕЧПЖШЛђГЄЦкФЭОУадРДДйНјГЩЙЧЁЃШЛЖјЃЌетжжЫЎФ§НКЕФЩњЮяСІбЇадФмКЭЕЏадЮДРДЕФИФНјНЋдкГЄЦкгІгУжаДјРДСМКУЕФЧАОАЁЃ
ДЫРрГЌЖЬыФЪЕЯжУтвпЕїНкЕФЛњжЦдкжЎЧАЕФбаОПжаЩаЮДУїШЗВћУїЁЃ[10a]ОоЪЩЯИАћЭЈЙ§ДњаЛжиБрГЬБЛМЄЛюЃЌдкЙЧдйЩњЙ§ГЬжаЕФУтвпЕїНкжаЦ№зХжСЙиживЊЕФзїгУЁЃ[4aЈC8 ] .ЯпСЃЬхзїЮЊУтвпЗДгІЕФаЕїепЃЌЕїНкОоЪЩЯИАћЕФДњаЛЦНКтЃЌВЂУмЧагАЯьОоЪЩЯИАћЕФБэаЭ[26],БОбаОПЗЂЯжЃЌUPN@hydrogelЭЈЙ§МЄЛюЯпСЃЬхЙІФмЃЌЕїПиОоЪЩЯИАћМЋЛЏНјШыM2БэаЭЁЃJC-1ЪЧгЩЛљжЪН№ЪєЕААзУИЃЈMMPsЃЉдкЯпСЃЬхжаЛ§РлЖјВњЩњЕФЁЃДЫЭтЃЌUPNДЬМЄЯджјдіЧПСЫЯпСЃЬхФЄЕчЮЛБъжОЮяJC-1ОлМЏЬхЕФаХКХЃЌдіМгСЫЯпСЃЬхШ§СзЫсЯймеЕФЪЭЗХЃЌНЕЕЭСЫЯпСЃЬхROSздгЩЛљЕФЪЭЗХЫйТЪЁЃвђДЫЃЌЮвУЧМйЩшЯпСЃЬхПЩФмДЋЕнаХКХВЂвдДњаЛЮяЕФаЮЪНЬсЙЉФмСПРДЮЌГжОоЪЩЯИАћБэаЭЁЃ
ЯпСЃЬхзїЮЊаХКХЯИАћЦїЃЌБЛШЯЮЊЪЧжївЊЕФЩњЮяФмСПЯИАћЦїЃЌЮЊЯИАћЛюЖЏЬсЙЉФмСПЃЌВЂВЮгыЯИАћЬивьадБэаЭЕФЖЏЬЌЕїНкЁЃ[27]ИќживЊЕФЪЧЃЌЯпСЃЬхДњаЛЮядкОоЪЩЯИАћМЋЛЏЕФгеЕМКЭЮЌГжжаЬсЙЉаХКХЃЌВЂзїЮЊдчЦкМьВщЕуЗжзгЗЂЛгживЊзїгУ[28]ЁЃБОбаОПБэУїЃЌUPN@ЫЎФ§НК геЕМОоЪЩЯИАћЕФ TCA бЛЗКЭЬЧНЭНтДњаЛЃЌЦфДњаЛВњЮяЖдОоЪЩЯИАћМЋЛЏжСЙиживЊЁЃчњчъЫсЪЧ TCA бЛЗжаЕФДйбзДњаЛЮяЃЌдкжЌЖрЬЧгеЕМЕФ M1 ОоЪЩЯИАћжаЛ§Рл[15a],ЮвУЧЗЂЯжUPN@hydrogelФмУїЯдНЕЕЭОоЪЩЯИАћжаДйбзадчњчъЫсЁЃШщЫсКЭЦЯЬбЬЧ-6-СзЫсЪЧЬЧНЭНтЕФДйбзВњЮя[29]ЃЌUPN@ЫЎФ§НКМѕШѕСЫОоЪЩЯИАћФмСПДњаЛжабзжЂДњаЛЮяЕФВњЩњЁЃзмЕФРДЫЕЃЌUPN@ЫЎФ§НКДйНјСЫПЙбзДњаЛЮяВЂвжжЦСЫОоЪЩЯИАћTCAбЛЗКЭЬЧНЭНтжаЕФДйбзДњаЛЮяЃЌетПЩФмДйНјM2аЭОоЪЩЯИАћМЋЛЏЁЃДЫЭтЃЌЮвУЧЕФбаОПБэУїЃЌUPNЛЙБэЯжГіЭЈЙ§STAT6/PPAR-ІУ/SOCS3аХКХЭЈТЗЕїНкM2аЭОоЪЩЯИАћМЋЛЏЕФЖюЭтФмСІЁЃОпЬхЖјбдЃЌАзЯИАћНщЫи-4 (IL-4) МЄЛюДЅЗЂзЊТМвђзгSTAT6ЃЌЦфзщзАГЩЛюЛЏЕФзЊТММЄЛювђзгВЂгыЬиЖЈАаЛљвђНсКЯЃЌДгЖјДйНјPPAR-ІУвьЖўОлЛЏЕФгеЕМКЭЫцКѓЕФM2ОоЪЩЯИАћЛюЛЏЁЃДЫЭтЃЌSTAT6вжжЦСЫSOCS3ЕФБэДяЃЌДгЖјДйНјM2аЭОоЪЩЯИАћЕФМЋЛЏ[Ш§ЪЎБОбаОПНсЙћЮЊUPNЭЈЙ§STAT6/PPAR-ІУ/SOCS3аХКХЭЈТЗЖдM2аЭОоЪЩЯИАћМЋЛЏЗЂЛгЕїПизїгУЕФЙлЕуЬсЙЉСЫгаСІЕФжЄОнЁЃ
ЮЊСЫВћУї UPN геЕМЕФ M2 аЭОоЪЩЯИАћКЭ BMSC жЎМфИДдгЕФЯрЛЅзїгУЃЌЮвУЧЩшМЦСЫвЛжжИДдгЕФЙВХрбјЯЕЭГЃЌИУЯЕЭГЩцМАЙЧЫшРДдДЕФОоЪЩЯИАћ (BMDM) КЭ BMSCЁЃЛёЕУЕФНсЙћУїШЗБэУїЃЌUPN геЕМЕФ M2 аЭОоЪЩЯИАћЯджјдіЧПСЫЙВХрбјЮЂЛЗОГжаЩњГЄвђзг BMP2 КЭ IGF-1 ЕФЗжУкЁЃетжждіЧПЗДЙ§РДгжЯджјдіЧПСЫ BMSC ЕФГЩЙЧЗжЛЏКЭЧЈвЦФмСІЃЌДгЖјЮЊ UPN геЕМЕФ M2 аЭОоЪЩЯИАћдкДйНјЙЧдйЩњЗНУцЕФЧБдкзїгУЬсЙЉСЫаТЕФМћНтЁЃЫфШЛЮвУЧжЎЧАЕФбаОПБэУї SESSE жБНгДйНјСЫ BMSC ГЩЙЧЗжЛЏЃЌЕЋашвЊГЌЙ§АЫжмЕФГЄЪБМфБЉТЖЁЃДЫЭтЃЌГЌЩљВЈЛЙгУгкПижЦ SESSE ЕФЪЭЗХЁЃдкЙЧаоИДЕФдчЦкНзЖЮЃЌУтвпЯИАћЦ№зХЙиМќзїгУЃЌЖј BMSC ГЩЙЧЛюадЭЈГЃЗЂЩњдкШ§ЬьКѓЁЃДЫЭтЃЌжЕЕУзЂвтЕФЪЧЃЌдкЬхЭтДЬМЄХЈЖШЮЊ 10 жС 200 ng/ml ЕФ UPN дк 6-24 аЁЪБФкБэЯжГіЖд BMSC ЭЬЪЩЕФЯджјЕжПЙСІЁЃетвЛгаШЄЕФЙлВьНсЙћЪЙЮвУЧЭЦВт SESSE ЫЎФ§НКПижЦ BMSC ЗжЛЏЕФжївЊЛњжЦЪЧЭЈЙ§УтвпЯИАћЕїНкНјааМфНгЕїНкЁЃЭЈЙ§гАЯьУтвпЯИАћШКЃЌSESSE ЫЎФ§НКПЩФмДДдьСЫвЛИігаРћЕФЮЂЛЗОГЃЌДйНј BMSC ЗжЛЏКЭЫцКѓЕФЙЧдйЩњЁЃ
ЮЊСЫЦРЙРГЌЩљДЅЗЂЕФ UPN@hydrogel ДйНјЙЧдйЩњЕФЙІаЇЃЌЮвУЧЪЙгУ 8 жмДѓЕФ C57BL/6J аЁЪѓНЈСЂСЫЙЧШБЫ№ФЃаЭЁЃЫцКѓЃЌгУГЌЩљДЅЗЂЕФ UPN@hydrogel жЮСЦВњЩњЕФЙЧШБЫ№ЃЌЪЙЮвУЧФмЙЛЦРЙРЦфдкДйНјЙЧаоИДЗНУцЕФжЮСЦЧБСІЁЃЪѕКѓ 14 ЬьЃЌНсЙћЯдЪО US + UPN@hydrogel зщЙЧгњКЯаЇЙћИќМбЁЃДЫЭтЃЌШЋУцЕФЩњЮябЇЗжЮіБэУїЃЌUS + UPN@hydrogel зщЕФЙЧаЮЬЌЗЂЩњЕААз 2 (BMP2) КЭ I аЭНКдЕААз (Col1) ЫЎЦНзюИпЃЌетгыЮвУЧДгЬхЭтЕїВщЁЃЪЙгУЫеФООЋКЭвСКь (H&E) ШОЩЋЕФзщжЏбЇМьВщЯдЪОЃЌаЁЪѓЙЧШБЫ№ЧјгђФкаТаЮГЩЕФЙЧаЁСКж№НЅБЛдйЩњЙЧШЁДњЁЃеыЖдГЩЙЧЛљвђБъМЧЮя BMP2 ЕФУтвпгЋЙтМьВтЯдЪОЧПСвЕФКьЩЋгЋЙтЃЌгыЖдее (Ctrl) зщЯрБШЃЌUS + UPN@hydrogel зщЯдЪОГізюЧПЕФгЋЙтЧПЖШЁЃетаЉНсЙћЙВЭЌБэУїЃЌдкЙЧШБЫ№НЈФЃПЊЪМКѓЕк 1-3 ЬьНјааЕФГЌЩљжЮСЦгааЇгеЕМОоЪЩЯИАћзЊЛЏЮЊ M2 бЧаЭЃЌДгЖјЯдзХИФБфЙЧЫ№ЩЫВПЮЛжмЮЇЕФУтвпЮЂЛЗОГЃЌзюжедіЧПЙЧШБЫ№ЕФгњКЯЙ§ГЬЁЃживЊЕФЪЧЃЌГЌЩљВЈЕФгІгУвВгаРћгкздзщзАыФДгЫЎФ§НКжаЪЭЗХГіРДЃЌетЪЧгЩгкГЌЩљВЈМгЫйЃЌДгЖјДйНјГЩЙЧВЂЯджјдіЧПЙЧШБЫ№ЕФаоИДФмСІЁЃ
4 НсТл
ЮвУЧПЊЗЂСЫвЛжжГЌЩљДЅЗЂыФФЩУзЯЫЮЌЫЎФ§НКЃЌПЩгааЇДйНјЙЧдйЩњЁЃГЌЖЬыФ SESSE ЩшМЦгагУгк M2 ОоЪЩЯИАћМЋЛЏКЭздзщзАЕФФЃПщЃЌвдИФБфОоЪЩЯИАћзДЬЌЁЃЪЭЗХЕФыФФЩУзЯЫЮЌМЄЛюОоЪЩЯИАћЯпСЃЬхжаЕФбѕЛЏгІМЄЃЌНјЖјвжжЦЛюадбѕЪЭЗХЃЌЭЌЪБгеЕМ M2 ОоЪЩЯИАћМгЫйЙЧЫшМфГфжЪИЩЯИАћЕФГЩЙЧЗжЛЏЁЃзмжЎЃЌетЯюбаОПЮЊЛљгкГЌЖЬыФЕФЛюадзщжЏЙЄГЬЬсЙЉСЫвЛжжаТЭООЖЃЌЦфжаПЩвдЩшМЦЛљгкГЌЖЬыФЕФЩњЮяЛюадВФСЯРДДйНјзщжЏЫ№ЩЫдйЩњЁЃ
5 ЪЕбщВПЗж/ЗНЗЈ
5.1 ГЌЖЬыФФЩУзЯЫЮЌЕФжЦБИ
УПДЮЪЕбщНЋЖГИЩыФШмНтгкЖўМзЛљбЧэП (DMSO) жаЃЌХЈЖШЮЊ 10.0 mg/mlЃЌжЦБИаТЯЪЕФ CFFSESSE ыФдвКЃЌвдБмУтдЄОлМЏЁЃШЛКѓЃЌНЋ 0.5 ml ыФдвКЕЮШы 2.0 ml ШЅРызгЫЎШмвКжаЃЌОВжУ 5 minЁЃЛьКЯЮядйеёЕД 12 hЃЌЕУЕНздзщзАЕФГЌЖЬыФФЩУзЯЫЮЌЁЃзюКѓЃЌгУШЅРызгЫЎЭИЮі 2 d ШЅГ§гаЛњШмМСЁЃНЋжЦБИКУЕФГЌЖЬыФФЩУзЯЫЮЌЖГИЩБЃДцЁЃ
5.2 ИКдиыФФЩУзЯЫЮЌЕФГЌЩљЯьгІЫЎФ§НКЕФжЦБИ
ЮЊСЫНЋыФФЩУзЯЫЮЌзАШыЫЎФ§НКжаЃЌНЋ 5.0 КСПЫжЦБИКУЕФыФФЩУзЯЫЮЌгы 1 КСЩ§ 2.5% (жиСП/ЬхЛ§) КЃдхЫсФЦЃЌШЛКѓНЋЦфМгШы 1.0 ml 0.1 M ТШЛЏИЦ (CaCl2)ШмвКЃЌОВжУ10ЗжжгЁЃгУШЅРызгЫЎЧхЯДИКдиыФФЩУзЯЫЮЌЕФЫЎФ§НКЁЃ
5.3 ГЌЖЬыФФЩУзЯЫЮЌЕФаЮУВ
гУЩЈУшЕчзгЯдЮЂОЕЃЈZeiss Sigma 300 VP вЧЦїЃЉЦРЙРГЌЖЬыФФЩУзЯЫЮЌЕФаЮЬЌЁЃдк 2 kV ЯТЪЙгУЕчзгФЃЪНСНДЮМЧТМЬиеїЁЃ
5.4 ГЌЖЬыФФЩУзЯЫЮЌЕФадФм
5.4.1зЯЭтПЩМћЙтЦз
зЯЭт-ПЩМћЮќЪеЙтЦзВЩгУзЯЭт-ПЩМћЗжЙтЙтЖШМЦЃЈINESAЃЌжаЙњЃЉдк 25 Ёц ЯТВтСПЁЃВЈГЄЗЖЮЇЩшЖЈЮЊ 200~650 nmЃЌЩЈУшЫйЖШЮЊжаЕШЁЃ
5.4.2 ФЃСП
ЪЙгУ HAKKE СїБфвЧЖдКЃдхЫсИЦЫЎФ§НКНјааСїБфЗжЮіЁЃЪЙгУеёЕДЬѕМўЯТЕФгІБфЩЈУшЪЕбщВтСПЫЎФ§НКЕФеГЕЏадЁЃЩЈУшВтЪд 0.1% жС 10% ЕФгІБфЃЌвдМЧТМДЂФмФЃСП (GЁф) КЭЫ№КФФЃСП (GЁх) жЕЁЃЫљгадЫааОљЪЙгУВЛЭЌЕФбљЦЗзїЮЊжиИДЁЃЪЙгУЕФЦЕТЪЮЊ 1 HzЃЌМфЯЖПэЖШЮЊ 5 mmЁЃ
5.4.3 ЫЎФ§НКНЕНтКЭГЌЩљВЈЯьгІНЕНт
Ф§НКЛЏКѓЃЌдк 37ЁуC ЕФЮТЖШЯТзаЯИГЦСПЫЎФ§НКЃЈW0ЃЉЁЃЫцКѓЃЌНЋЦфНўШы pH жЕЮЊ 7.4 ЕФСзЫсбЮЛКГхбЮЫЎ (PBS) жаЃЌвдДДНЈЪЪЕБЕФЩњРэЯрЙиЛЗОГЁЃШЛКѓЃЌДгбљЦЗжаГ§ШЅЭтВПЫЎЃЌВЂвдЬиЖЈМфИєГЦСПбљЦЗЃЈWSЃЉЁЃЫљгаЪЕбщОљжиИДНјааШ§ДЮЁЃЖдгкГЌЩљЯьгІНЕНтЪЕбщЃЌвдМфИєЪБМфЃЈ20 kHzЁЂ45 WЁЂ3 ЗжжгЃЉЪЙгУГЌЩљВЈДІРэбљЦЗЁЃдкЩЯЪіЪЕбщжаЃЌЫљгабљЦЗОљдкГ§ШЅЭтВПЫЎКѓГЦжиЁЃ
5.4.4 ыФЪЭЗХКЭГЌЩљЯьгІЪЭЗХ
дкгаЛђУЛгаГЌЩљееЩфЕФЧщПіЯТЗжЮіЫЎФ§НКЪЭЗХыФЁЃНЋКЌгаыФЕФЫЎФ§НКНўШы5.0 ml PBSЛКГхвКЃЈpH 7.4ЃЉжаЁЃдкдЄЩшЕФЪБМфМфИєКѓЃЌДгЛьКЯШмвКжаЪеМЏ1.0 mlШмвКЃЌВЂМгШыЯрЭЌЬхЛ§ЕФаТЯЪPBSЁЃШЛКѓЪЙгУгЋЙтЗжЙтЙтЖШМЦЗжЮіыФЕФХЈЖШЁЃдкГЌЩљДЬМЄЕФыФЪЭЗХЪЕбщжаЃЌЪЙгУГЌЩљВЈЃЈ20 kHzЃЌ45 WЃЌ3ЗжжгЃЉвдЬиЖЈМфИєееЩфбљЦЗЁЃШЛКѓЃЌЬсШЁ1.0 mlШмвКВЂЪЙгУгЋЙтЗжЙтЙтЖШМЦНјааЗжЮіЁЃ
5.5 аЁЪѓЙЩЙЧШБЫ№ФЃаЭЕФНЈСЂ
БОЪЕбщОЭЌМУДѓбЇПкЧЛвНбЇдКвНбЇТзРэЮЏдБЛсХњзМЃЈ2019-DW-040ЃЉЁЃЪЕбщЪЙгУ8жмСфаладC57BL/6JаЁЪѓ36жЛЁЃаЁЪѓЫЧбјдкЮоЬиЖЈВЁдЬхЃЈSPFЃЉЛЗОГжаЃЌЮТЖШЃЈ22ЁР1ЁцЃЉКЭЪЊЖШЃЈ40%ЁЋ60%ЃЉПЩПиЃЌЙтее/КкАЕжмЦкЮЊ12аЁЪБЃЌЬсЙЉЪГЮяКЭЫЎЁЃЪжЪѕЦїаЕдкЪѕЧАИпбЙУ№ОњЃЌ10%ЫЎКЯТШШЉЃЈ3 ml/kgЃЉОВТізЂЩфЁЃаЁЪѓТщзэКѓвЦжСЪЕбщЬЈЃЌЪжЪѕВПЮЛгУ3%ЕтєњШмвККЭ75%ОЦОЋЯћЖОЁЃЫцКѓЃЌЧаПЊаЁЪѓЙЩЙЧЦЄЗєЃЌЖладАўРыМЁШтКЭНюФЄЃЌБЉТЖЙЩЙЧЃЌ0.ЪЙгУжБОЖ8 mmЧђаЮзъЭЗДЉЭИвЛВрЙЧЦЄжЪЃЌСЂМДНЋГЌЖЬыФФЩУзЯЫЮЌЫЎФ§НКОжВПжУгкЙЧШБЫ№ВПЮЛЃЌдйгУПЩЮќЪе4-0ЯИЫПЗьКЯЦЄЗєЃЌЙЙНЈЙЩЙЧЫ№ЩЫаоИДФЃаЭЁЃЪѕКѓЪЙгУПЙЩњЫиПижЦИаШОЃЌЕчШШДВЕцЃЈ38.3ЁР0.5ЃЉЁцЮЊЖЏЮяБЃХЏЃЌД§ЖЏЮяЭъШЋДгТщзэжаЫеабКѓЗХЛиКЯЪЪС§ОпЁЃаЁЪѓЙЧШБЫ№ЪжЪѕКѓЧА3ЬьдкШБЫ№ВПЮЛЭПФЈвНгУёюКЯМСЃЌГЌЩљжЮСЦ2 minЃЌЧПЖШЮЊ1.75 W/cmгУПЩЮќЪе4-0ЯИЫПЗьКЯЦЄЗєЃЌЙЙНЈЙЩЙЧЫ№ЩЫаоИДФЃаЭЁЃЪѕКѓЪЙгУПЙЩњЫиПижЦИаШОЃЌЕчШШДВЕцЃЈ38.3ЁР0.5ЃЉЁцЮЊЖЏЮяБЃХЏЁЃД§ЖЏЮяЭъШЋДгТщзэжаЫеабКѓЃЌЗХЛиЯргІС§згЁЃаЁЪѓЙЧШБЫ№ЪжЪѕКѓЧА3ЬьдкШБЫ№ДІЭПФЈвНгУёюКЯМСЃЌГЌЩљжЮСЦ2ЗжжгЃЌ1.75 W/cmгУПЩЮќЪе4-0ЯИЫПЗьКЯЦЄЗєЃЌЙЙНЈЙЩЙЧЫ№ЩЫаоИДФЃаЭЁЃЪѕКѓЪЙгУПЙЩњЫиПижЦИаШОЃЌЕчШШДВЕцЃЈ38.3ЁР0.5ЃЉЁцЮЊЖЏЮяБЃХЏЁЃД§ЖЏЮяЭъШЋДгТщзэжаЫеабКѓЃЌЗХЛиЯргІС§згЁЃаЁЪѓЙЧШБЫ№ЪжЪѕКѓЧА3ЬьдкШБЫ№ДІЭПФЈвНгУёюКЯМСЃЌГЌЩљжЮСЦ2ЗжжгЃЌ1.75 W/cmаЁЪѓЙЧШБЫ№ЪжЪѕКѓЃЌЧАШ§ЬьдкШБЫ№ВПЮЛЭПФЈвНгУёюКЯМСЃЌвд1.75 W/cm2 ЕФЙІТЪНјаа 2 ЗжжгЕФГЌЩљжЮСЦаЁЪѓЙЧШБЫ№ЪжЪѕКѓЃЌЧАШ§ЬьдкШБЫ№ВПЮЛЭПФЈвНгУёюКЯМСЃЌвд1.75 W/cm2 ЕФЙІТЪНјаа 2 ЗжжгЕФГЌЩљжЮСЦ2Эп/ЦНЗНУзЃЈ1 M/HzЃЉЁЃБОДЮЪЕбщжаЕФаЁЪѓБЛЗжЮЊСљзщЃКЃЈ1ЃЉCtrl зщЃКЮДНјааШЮКЮжЮСЦЕФзщЃЈБэЪОЮЊЁАЖдеезщЁБЃЉЃЛЃЈ2ЃЉЮДМгдиЫЎФ§НКзщЃКЮДИКдиКЃдхЫсИЦЫЎФ§НКЃЈМЧЮЊЁАЮДИКдиЫЎФ§НКзщЁБЃЉЃЛЃЈ3ЃЉUP@ЫЎФ§НКзщЃКГЌЖЬыФИКдиКЃдхЫсИЦЫЎФ§НКЃЈМЧЮЊЁАUP@ЫЎФ§НКзщЁБЃЉЃЛЃЈ4ЃЉУРЙњ+UP@ЫЎФ§НКзщЃКГЌЩљДІРэ+ГЌЖЬыФИКдиКЃдхЫсИЦЫЎФ§НКЃЈМЧЮЊЁАUS+UP@ЫЎФ§НКзщЁБЃЉЃЛЃЈ5ЃЉUPN@ЫЎФ§НКзщЃКГЌЖЬыФФЩУзЯЫЮЌИКдиКЃдхЫсИЦЫЎФ§НКЃЈМЧЮЊЁАUPN@ЫЎФ§НКзщЁБЃЉЃЛЃЈ6ЃЉУРЙњ+UPN@hydrogel зщЃКГЌЩљДІРэ+ГЌЖЬыФФЩУзЯЫЮЌИКдиКЃдхЫсИЦЫЎФ§НКЃЈМЧЮЊЁАUS+UPN@ЫЎФ§НКзщЁБЃЉЁЃУПзщ6жЛЪЕбщаЁЪѓЁЃ
5.6 ЮЂаЭМЦЫуЛњЖЯВуЩЈУшЃЈCTЃЉМьВтЙЧШБЫ№аоИД
ЖдеезщКЭЪЕбщзщЙВ36жЛаЁЪѓдкЙЩЙЧЫ№ЩЫКѓ14ЬьБЛАВРжЫРЁЃЪЙгУmicro-CTЯЕЭГЃЈmicro-CT50ЃЌШ№ЪПЃЉЗжЮіЙЧШБЫ№бљБОЁЃМђЖјбджЎЃЌУПИібљБОДгЙЧШБЫ№ДІЩЈУш80ВуЃЌАќРЈећИіЙЧЫ№ЩЫЧјгђЃЌВЂдк70 kVКЭ200 mAЯТСПЛЏбЁЖЈЧјгђФкЫљгаЙЧаЁСКЕФЙЧУмЖШЃЌПеМфЬхЫиДѓаЁЮЊ10ІЬmЁЃДЫЭтЃЌДгmicro-CTЗжЮіжадйЩњвдЯТЙЧУмЖШВЮЪ§ЃКаЁСКЬхЛ§/змЬхЛ§ЃЈBV/TVЃЉКЭЙЧаЁСККёЖШЃЈTb.ThЃЉЁЃ
5.7 ЩњЮяВтЖЈ
5.7.1 аЁЪѓЙЧЫшМфГфжЪИЩЯИАћЃЈBMSCsЃЉЕФЗжРыКЭХрбј
Жд36жЛSPFМЖ4жмСфC57BL/6JаЁЪѓЪЕЪЉЮоЭДАВРжЫРКѓЃЌНЋаЁЪѓНўХндк75ЃЅввДМжа10ЗжжгЁЃдкЙЩЙЧКЭыжЙЧДІЧаПЊЦЄЗєЃЌШЛКѓЮоОњЗжРыЙЩЙЧЃЌВЂНўХндкКЌгаЧрУЙЫи-СДУЙЫиЕФЮоОњPBSШмвКжаЁЃШЛКѓЃЌШЅГ§ЙЩЙЧКЭыжЙЧжмЮЇЕФМЁШтКЭНсЕозщжЏЃЌВЂдкЫЋПЙОњPBSШмвКжаГхЯДШ§ДЮЁЃНгЯТРДЃЌЧаПЊЙЩЙЧКЭыжЙЧЕФСНЖЫЃЌБЉТЖЙЧЫшЧЛЁЃгУ2 mlзЂЩфЦїгУЭъШЋХрбјвКГхЯДбљЦЗШ§ДЮЁЃДЫЭтЃЌЙЧЫшЧЛЛЙвЊГхЯДЪ§ДЮЃЌжБжСЙЧБкбеЩЋБфАзЁЃШЛКѓНЋЯИАћаќвКЭЈЙ§200ФПФсСњЭјжС15 mlРыаФЙмжаЃЌВЂдк2000 rpmЯТРыаФ5ЗжжгЁЃЦњШЅЩЯЧхвКЃЌгУКьЯИАћСбНтвКжиаќЯИАћЃЌ4 ЁцСбНт10 minЃЌМгШыЕШСПХрбјЛљжежЙКьЯИАћСбНтЃЌ2000 rpm РыаФ5 minЃЌМЦЪ§ЯИАћЃЌНгжжгк10 cm ХрбјУѓЃЌ37 ЁцЁЂ5% CO Хрбј2ЪЊЖШБЅКЭЁЃ
5.7.2 аЁЪѓЙЧЫшОоЪЩЯИАћЃЈBMDMЃЉЕФЗжРы
ДгаЁЪѓЙЧЫшЧЛжаЬсШЁЯИАћЕФЙ§ГЬгыаЁЪѓЙЧЫшМфГфжЪИЩЯИАћЕФЙ§ГЬРрЫЦЃЌжБЕНКьЯИАћСбНтЃЌШЛКѓНЋЯИАћжиаТаќИЁдк M-CSF ХрбјЛљЃЈ40 ng/mlЃЉЃЈжизщаЁЪѓ M-CSFЃЛPetroleum TechnologyЃЌRocky MountainЃЌNJЃЉжаЁЃЕк 2 ЬьКЭЕк 3 ЬьВЙГфКЌга M-CSF ЕФХрбјЛљЃЌЕк 5 ЬьЬэМгаТЯЪЕФ M-CSF ХрбјЛљЁЃетЬхЭтЪЕбщАќРЈЫФзщЃКЃЈ1ЃЉCtrl зщЃКЮДНгЪмШЮКЮжЮСЦЕФзщЃЈБэЪОЮЊЁАЖдеезщЁБЃЉЃЛЃЈ2ЃЉIL-4зщЃКгУIL-4геЕМОоЪЩЯИАћЃЈБъМЧЮЊЁАIL-4зщЁБЃЉЃЛЃЈ3ЃЉUPN зщЃКгУГЌЖЬыФФЩУзЯЫЮЌгеЕМОоЪЩЯИАћЃЈМЧЮЊЁАUPNзщЁБЃЉЃЛЃЈ4ЃЉIL-4+UPNзщЃКгУIL-4КЭГЌЖЬыФФЩУзЯЫЮЌгеЕМОоЪЩЯИАћЃЈБэЪОЮЊЁАIL-4+UPNзщЁБЃЉЁЃ
5.7.3 УтвпгЋЙт
УтвпгЋЙтШОЩЋМьВтОоЪЩЯИАћМЋЛЏМАИЩЯИАћГЩЙЧФмСІЁЃСљзщОљдкЙЧЫ№ЩЫжЮСЦКѓ14 dДІЫРЃЌЪеМЏЙЧШБЫ№ДІЕФЙЩЙЧЃЌгУ4%ЖрОлМзШЉЙЬЖЈЃЌЙЬЖЈ48 hКѓНЋЪеМЏЕФзщжЏЗХШы10%EDTAжаЗХжУ4жмЃЌУП2 dИќЛЛвЛДЮЭбИЦвКЁЃЭбИЦБъБОгУPBSЧхЯДЃЌгУ30%есЬЧШмвКЭбЫЎ48 hКѓНўШыTissue-Tek® O⋅CTАќТёЁЃгУЕЭЮТЧаЦЌЛњЃЈЕТЙњLeica MicrosystemsЃЉСЌајЧаИюЙЧШБЫ№ДІЃЌУПВуКёЖШОљдШЮЊ4 ІЬmЃЌЧаИюКѓЕФБъБОЪвЮТЗХжУ30 minКѓЃЌЗХШы4 ЁцБћЭЊжа10 minЁЃКцИЩ30 minКѓЃЌPBSЯДЕг3ДЮЃЌУПДЮ5 minЁЃЫцКѓЃЌгУ10%ЩНбђбЊЧхдкЪвЮТЯТЗтБеЧаЦЌ1 hЃЌНјааУтвпгЋЙтШОЩЋЁЃНЋгУЪЪЕББШР§ЯЁЪЭЕФвЛПЙЙЄзївКЃЈBMP2ЃЌ1:200ЃЌBiossЃЉЕФPBSЕЮМгЕНбљЦЗжаЃЌдк4 ЁцБмЙтЛЗОГжаЗѕг§Й§вЙЁЃгУPBSЕЮЯД3ДЮЃЌУПДЮ10 minЃЌШЛКѓМгШыЪЪЕБЯЁЪЭЕФЖўПЙЙЄзївККЭЪѓдДПЙЬхЃЈ594ЃЌ1:1000ЃЌThermo FisherЃЉЁЃдкЙВОлНЙЯдЮЂОЕЃЈNIS ElementsЃЌNikonЃЌШеБОЃЉЯТЃЌЪЙгУзЈУХЕФЭМЯёШэМўЃЈImageJЃЌNIHЃЌУРЙњЃЉНјааЗжЮіЁЃЧаЦЌгУ10%бђбЊЧхЪвЮТЗтБе1hКѓНјааУтвпгЋЙтШОЩЋЁЃНЋгУЪЪЕББШР§ЯЁЪЭЕФвЛПЙЙЄзївК(BMP2,1:200,Bioss)ЕФPBSЕЮМгЕНбљЦЗжаЃЌ4ЁцБмЙтЗѕг§Й§вЙЁЃгУPBSЕЮЯДШ§ДЮЃЌУПДЮ10minЃЌШЛКѓМгШыЪЪЕБЯЁЪЭЕФЖўПЙЙЄзївККЭЪѓдДПЙЬх(594,1:1000,Thermo Fisher)ЁЃдкЙВОлНЙЯдЮЂОЕ(NIS Elements,Nikon,Japan)ЯТЃЌЪЙгУзЈУХЕФЭМЯёШэМў(ImageJ,NIH,USA)НјааЗжЮіЁЃЧаЦЌгУ10%бђбЊЧхЪвЮТЗтБе1hКѓНјааУтвпгЋЙтШОЩЋЁЃНЋгУЪЪЕББШР§ЯЁЪЭЕФвЛПЙЙЄзївК(BMP2,1:200,Bioss)ЕФPBSЕЮМгЕНбљЦЗжаЃЌ4ЁцБмЙтЗѕг§Й§вЙЁЃгУPBSЕЮЯДШ§ДЮЃЌУПДЮ10minЃЌШЛКѓМгШыЪЪЕБЯЁЪЭЕФЖўПЙЙЄзївККЭЪѓдДПЙЬх(594,1:1000,Thermo Fisher)ЁЃдкЙВОлНЙЯдЮЂОЕ(NIS Elements,Nikon,Japan)ЯТЃЌЪЙгУзЈУХЕФЭМЯёШэМў(ImageJ,NIH,USA)НјааЗжЮіЁЃPBSЕЮЯД3ДЮЃЌУПДЮ10 minЃЌМгШыЪЪЕБЯЁЪЭЕФЖўПЙЙЄзївКМАЪѓдДПЙЬхЃЈ594ЃЌ1:1000ЃЌThermo FisherЃЉЃЌдкЙВОлНЙЯдЮЂОЕЃЈNIS ElementsЃЌШеБОФсПЕЃЉЯТЃЌВЩгУзЈУХЕФЭМЯёШэМўЃЈImageJЃЌУРЙњNIHЃЉНјааЗжЮіЁЃPBSЕЮЯД3ДЮЃЌУПДЮ10 minЃЌМгШыЪЪЕБЯЁЪЭЕФЖўПЙЙЄзївКМАЪѓдДПЙЬхЃЈ594ЃЌ1:1000ЃЌThermo FisherЃЉЃЌдкЙВОлНЙЯдЮЂОЕЃЈNIS ElementsЃЌШеБОФсПЕЃЉЯТЃЌВЩгУзЈУХЕФЭМЯёШэМўЃЈImageJЃЌУРЙњNIHЃЉНјааЗжЮіЁЃ
ЮЊСЫЦРЙРЯИАћЧЈвЦЃЌНЋЯИАћдкЪвЮТЯТгУ 4% ЖрОлМзШЉЙЬЖЈ 20 ЗжжгЃЌгУ PBS ЧхЯДЃЌШЛКѓгУ 10% ЩНбђбЊЧхЗтБе 1 аЁЪБЁЃгУ PBS дйДЮЧхЯДЯИАћ 3 ДЮЃЌУПДЮ 5 ЗжжгЁЃНЋгУЪЪЕББШР§ЕФвЛПЙЙЄзїШмвКЃЈArg1ЃЌ1:200ЃЌBiossЃЛCD206ЃЌ1:200ЃЌProteintechЃЛp-stat6ЃЌ1:100ЃЌCell SignalЃЉЯЁЪЭЕФ PBS ж№ЕЮМгШыбљЦЗжаЃЌВЂдкКкАЕЛЗОГжадк 4 ЁуC ЯТЗѕг§Й§вЙЁЃдкЙВОлНЙЯдЮЂОЕЃЈNIS ElementsЃЌФсПЕЃЌШеБОЃЉЯТЃЌЪЙгУзЈвЕЭМЯёШэМўЃЈImageJЃЌNIHЃЌУРЙњЃЉНјааЗжЮіЁЃ
5.7.4 ЕААзжЪгЁМЃЗЈ
НЋдЄРфЕФСбНтЛКГхвКМгШыЯИАћжаЃЌЕЋдкСбНтЧАЯђЯИАћжаМгШы 50 ІЬl ЕААзУИвжжЦМСКЭ 50 ІЬl СзЫсУИвжжЦМСЁЃНгЯТРДЃЌНЋ 250 ІЬl WB Super RIPA СбНтЛКГхвКМгШыЕН 1 ЁС 10 6ЯИАћЃЌМгШыЕААзУИвжжЦМСКЭСзЫсУИвжжЦМСЛьКЯЮяИї2.5 ІЬlЁЃНЋЛьКЯЮяжУгкБљЩЯЪЙЯИАћСбНт30ЗжжгЃЌШЛКѓдк4ЁуCвд13,000 rpmРыаФ5ЗжжгЁЃЪеМЏЩЯЧхвКЃЌЦфжаКЌгаашвЊЬсШЁЕФЕААзжЪЁЃЪЙгУBCAММЪѕВтЖЈЕААзжЪКЌСПЃЌВЂМЦЫуЫљЕУбљЦЗЕФХЈЖШЁЃгыЪЪЕБЛКГхвКЛьКЯКѓЃЌ5ЁСЪЎЖўЭщЛљСђЫсФЦЃЈSDSЃЉФ§НКдк70ЁуCЯТМгШШ10ЗжжгвдБфадЁЃЪЙгУSDS-PAGEКЭPVDFФЄгааЇЕиЗжРыЕААзжЪЬсШЁЮяЃЌЫљЪіФЄгУДПМзДМБЅКЭ5УыжгЁЃгУПьЫйзшЖЯШмвКЃЈBiotechЃЌNOЁЃC510053ЃЉНјааЕААзжЪзшЖЯЃЌВЂНЋбљЦЗгыЯЁЪЭЕФвЛПЙдк4ЁуCЯТЗѕг§Й§вЙЁЃTBSTЃЈBiotechЃЌNo.ЪЙгУЖўПЙЃЈC006161ЃЉЯДPVDFФЄШ§ДЮЃЌУПДЮ10 minЁЃМгШыЯЁЪЭКѓЕФЖўПЙЃЌЗѕг§1 hЁЃPVDFФЄЯдгАВЩгУЛЏбЇЗЂЙтЪдМСКаЃЈPierceЙЋЫОЃЌУРЙњЃЉЁЃвЛПЙЙЄзївКЮЊЃКBMP2ЃЌ1ЃК1000ЃЈBiossЙЋЫОЃЌжаЙњЃЉЃЛCol1ЃЌ1ЃК1000ЃЈBiossЙЋЫОЃЌжаЙњЃЉЃЛArg1ЃЌ1ЃК1000ЃЈBiossЙЋЫОЃЌжаЙњЃЉЃЛCD206ЃЌ1ЃК1000ЃЈProteintechЙЋЫОЃЌУРЙњЃЉЁЃЖўПЙЙЄзївКЮЊЃКA32790ЃЌ1ЃК1000ЃЛA32729ЃЌ1ЃК1000ЃЈThermo FisherЙЋЫОЃЌжаЙњЃЉЁЃArg1ЃЌ1ЃК1000ЃЈBiossЃЌжаЙњЃЉЃЛCD206ЃЌ1ЃК1000ЃЈProteintechЃЌУРЙњЃЉЃЛЖўПЙЙЄзївКЮЊЃКA32790ЃЌ1ЃК1000ЃЛA32729ЃЌ1ЃК1000ЃЈThermo FisherЃЌжаЙњЃЉЁЃArg1ЃЌ1ЃК1000ЃЈBiossЃЌжаЙњЃЉЃЛCD206ЃЌ1ЃК1000ЃЈProteintechЃЌУРЙњЃЉЃЛЖўПЙЙЄзївКЮЊЃКA32790ЃЌ1ЃК1000ЃЛA32729ЃЌ1ЃК1000ЃЈThermo FisherЃЌжаЙњЃЉЁЃ
5.7.5 ЪЕЪБЖЈСПОлКЯУИСДЪНЗДгІ (RT-qPCR)
ИљОнжЦдьЩЬЕФЫЕУїЃЌЪЙгУ TRIzolЃЈTakaraЃЌжаЙњЃЉЪдМСЬсШЁ RNAЃЌВЂЪЙгУ HiScript II Q RT SuperMix НЋ 1.0 ІЬg RNA ФцзЊТМЮЊ cDNAЁЃЫљгаЪ§ОнОљЛљгкІТ-actin зїЮЊЛљгкЯШЧАбаОПЕФЭГвЛВЮПМ[27].RT-qPCRв§ЮяађСаШчЯТЃК
5.7.6 мчЫиКьШОЩЋ
НЋ BMSC вд 2 ЁС 10 4 ИіЯИАћ/mlЕФУмЖШНгжждк 12 ПзАхЩЯЃЌВЂдкЕк 21 ЬьЯИАћУмЖШДяЕН 80% ЛуКЯЪБгУмчЫиКьШОЩЋЁЃмчЫиКьШОЩЋШмвК (Sigma-Aldrich) ИљОнжЦдьЩЬЕФЫЕУїжЦБИЃЌЕїНк pH жС 8.3 ВЂНЋЦфДЂДцдк 4 ЁуC ЯТвдБИКѓгУЁЃЖдШОЩЋбљЦЗНјааСНДЮ PBS (Promo CellЃЌгЂЙњ) ЧхЯДЃЌУПДЮ 5 ЗжжгЁЃПзФкЕФЯИАћдкЪвЮТЯТгУ 4% ЖрОлМзШЉЙЬЖЈ 30 ЗжжгЃЌгУеєСѓЫЎ (diH2O) Ш§ДЮЃЌгУмчЫиКьШОЩЋвКдк 37ЁуC ЯТШОЩЋ 30 ЗжжгЃЌгУ diH2OВЂИЩдяЁЃЪЙгУжБСЂЙтбЇЯдЮЂОЕЃЈLeicaЃЌгЂЙњЃЉдк20БЖЗХДѓБЖЪ§ЯТЙлВьЯИАћФкЕФКьЩЋПѓЛЏНсНкЁЃ
5.7.7 МюадСзЫсУИЛюадМьВт
ЗжРыХрбј BMSC КѓЃЌвд 6ЁС10 4 ИіЯИАћ/mlЕФУмЖШНгжжгк 12 ПзАхжаЃЌХрбј 7 КЭ 14 ЬьЁЃЕБЯИАћУмЖШДяЕН 80% ЪБНјааМюадСзЫсУИ (ALP) ШОЩЋЁЃМюадСзЫсУИЯдЩЋвКАДеежЦдьЩЬЕФЫЕУїжЦБИЃЈMlbioЃЌЩЯКЃЃЌжаЙњЃЉЁЃЪЙгУ PBSЃЈPromoCellЃЌгЂЙњЃЉГхЯДбљЦЗСНДЮЃЌУПДЮ 5 ЗжжгЁЃНЋПзжаЕФЯИАћдкЪвЮТЯТгУ 4% ЖрОлМзШЉЙЬЖЈ 30 ЗжжгЃЌВЂЪЙгУ diH2O.ШЛКѓЃЌНЋЯИАћдк37ЁуCЯТЗѕг§30ЗжжгЃЌгУdiH2OВЂИЩдяЁЃгУЙтбЇЯдЮЂОЕЃЈLeicaЃЌгЂЙњЃЉдк20БЖЗХДѓБЖЪ§ЯТЖдЯИАћНјааХФееЃЌМюадСзЫсУИШОЩЋГЪЯжРЖЩЋЁЃ
5.7.8 ЯИАћЖОадВтЪдЬхЭт
UPзщКЭUPNзщЬсЧАЗѕг§6 hЃЌХЈЖШЗжБ№ЮЊ6.25 ng/mlЁЂ12.5 ng/mlЁЂ25 ng/mlЁЂ50 ng/mlЁЂ100 ng/mlКЭ200 ng/mlЃЌВЛЭЌХЈЖШЛюадыФБЉТЖЯИАћЕФХрбјЪБМфЮЊ0ЁЂ48КЭ72 hЁЃЪЙгУCell Counting Kit-8ЃЈCCK-8ЃЉМьВтЯИАћдіжГЧщПіЃЌОпЬхВйзїВЮееГЇМвЫЕУїЁЃУП2 mlХрбјЛљМгШы100 ІЬl CCK-8ЃЌЗѕг§2 hЁЃЪЙгУGen5™УИБъвЧЃЈBioTekвЧЦїЃЉдк450 nmДІЖдЯИАћНјааМьВтЃЈ12ПзАх3ЁС3ЕуЃЉЃЌВЂдкЯдЮЂОЕЯТЙлВьЯИАћЕФЛюадзДЬЌЁЃ
5.7.9 ЯИАћЖОадВтЪдЬхФк
ЙЧШБЫ№аЁЪѓЃЌUS+UP@hydrogelзщЗХжУГЌЖЬыФЫЎФ§НКЃЌUS+UPN@hydrogelзщЗХжУГЌЖЬыФФЩУзЯЫЮЌЫЎФ§НКЃЌСНзщОљНјааГЌЩљДІРэЁЃ14ЬьКѓШЁЦфаФдрЁЂЩідрЁЂЦЂдрЁЂЗЮдрКЭИЮдрНјааЫеФООЋКЭвСКьЃЈH&EЃЉШОЩЋЃЌЙлВьетаЉЦїЙйЕФЯИАћНсЙЙКЭаЮЬЌЃЌВЂУмЧаЙлВьаЁЪѓЕФзДЬЌЁЃ
5.8 ЭГМЦЗжЮі
Ъ§ОнвдЦНОљжЕЁРБъзМВюБэЪОЁЃЪЙгУ SPSS 19.0 ШэМўЃЈSPSSЃЌжЅМгИчЃЌILЃЉНјааЭГМЦЗжЮіЁЃВЩгУ Kolmogorov-Smirnov МьбщРДбщжЄЪ§ОнЕФе§ЬЌЗжВМЁЃЪЙгУЕЅвђЫиЗНВюЗжЮі (ANOVA) МьбщБфСПЕФЯджјадгАЯьЁЃЪЙгУЖржиБШНЯ Tukey МьбщдкдЄЩшЕФ alpha ЮЊ 0.05 ЪББШНЯЪ§ОнЁЃP жЕ < 0.05 БЛШЯЮЊОпгаЭГМЦбЇвтвхЁЃ
Утд№ЩљУїЃКБОЮФЮЊаавЕНЛСїбЇЯАЃЌАцШЈЙщдзїепМАддгжОЫљгаЃЌШчгаЧжШЈЃЌПЩСЊЯЕЩОГ§ЁЃЮФеТБъзЂгазїепМАЮФеТГіДІЃЌШчашдФЖСдЮФМАВЮПМЮФЯзЃЌПЩдФЖСддгжОЁЃ
