ժҪ�����ܹ������ӵĹ���������������ҩ�����ϵͳ�Ĺؼ���������Ϊһ�����˵ĸ��ӵ���ϵͳ���ԣ���ҩ��ż���� (PDC) ��ǿ���ܹ��������ܽ�ͷ�Ͷ�������ı��ֳ��㷺�Ĺ��ܺ����ԣ��������ڻ�ѧ�ϳɺͶ������Ρ��ɵ��ڵ����������ԡ�������������װ���ṹ��������ʶ��/����Լ���ϸ��Ĥ/ϸ������ʵ�����������⣬�������͵Ľ�ͷʹ PDC �ܹ����ݼ�������������������Ӧ�Ե��ͷ�ҩ�������Щ���е����ƣ�PDC �ѱ��㷺����ҩ����͡��ڴˣ������˹�������PDCs���õĹ����ĺ������ӵ����½�չ��ּ���������Ƕ�PDC��Ʋ��Լ���Կ������༲�����Ĺ㷺��ע����Ϊ����PDCҩ�����ϵͳ���о��ṩָ����
1 ����
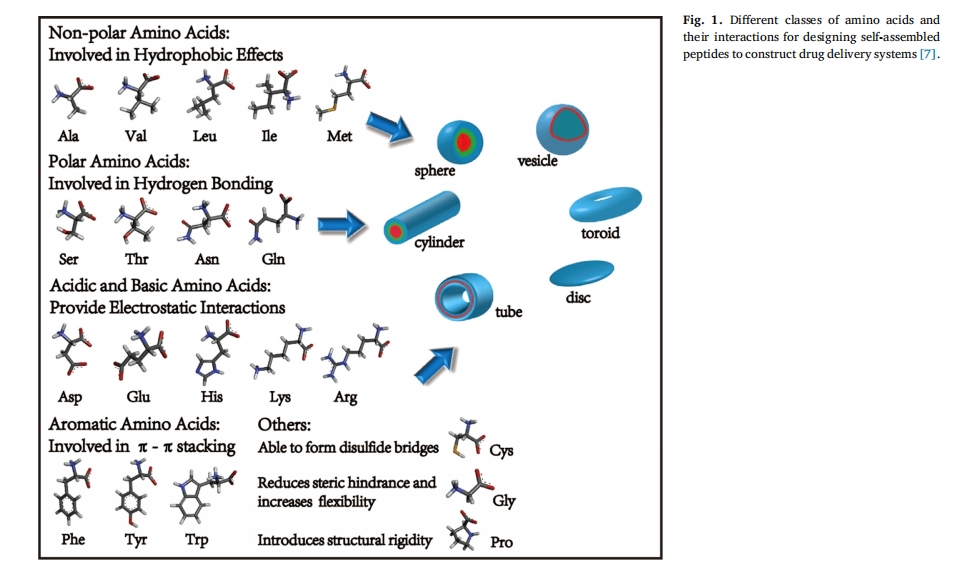
��ϸ����ҩ�����ϵͳ���о�ּ�ڱ�֤ҩ�ﰲȫ���ɿص��ͷţ��Դﵽ��ѵ�ҩЧ����С�ĸ����á��������ھ������õ����������ԡ����オ���ԡ�����װ�ԺͲ�ͬ�Ļ�ѧ�����ԣ��ڹ�������������ϵͳ�������������ǵļ�����Ȥ�����ڹ�������ҩ��Ķ���ͨ������Ϊ2-30��������л���ͨ�����20�������ἰ������������кͽṹ���Ի����ǧ�ֹ��ܺͽṹ���������Զ����벻ͬ��ҩ���Թ��ۻ�ǹ��۷�ʽ��ϣ��������ڶ��ĵĵ���ϵͳ�����Բ������õ�����Ч������ˣ������ڻ�����ͷ���������ơ�[1], �Ѻ�����[2]��������[3]��С���ӻ���ҩ��[4]����ҩ��[5]��ϸ��[6]�������������������Ƹ��ּ��������ָ����İ�����л�������ˮ�ԡ���ˮ�ԡ������ԡ����Ժͼ������ᣬ�������ڹ��������������С���Щ��ͬ�İ�����Ϊ���ṩ�˽ṹ�������Բ�������������������⣬��Эͬ�ǹ�������õ������£��Ŀ����Է��γ���ȷ�����ṹ���������װ����ܡ���ά�����������Ȧ�Լ�������ͼ 1��[7]
������˾��и��ӡ��ɵ��ص�����װ�ṹ�⣬�����ж��ַ�Ӧ���ţ������ֽ���š��������š��ϻ����ŵȣ�ʹ��ͨ�����ۼ�����ҩ������������Ϳ��С����⣬�����һ�ֲ����������ڸ���ϸ�����ܵ�����������ʣ��书����ͨ��ģ���ض��ĵ����ʹ�����ʵ�ֵġ���������ɾ��ж��ֹ��ܣ�������ԡ��鳲�ԡ�ϸ�����Ժ�Ѫ�����ϴ��ԡ�
��ҩ��ż���PDC����һ�����͵�ǰ��ҩ����ԣ�������Ĺ�����Ҫ�鹦�ڶ��������Ľṹ�����Ӧ�Խ�ͷ��PDC ��Ҫ������������ɣ��������Ľṹ����ͽ�ͷƬ�Σ�ͼ 2��

���������о���Ա����PDCs��������Ժ�����װDZ�ܣ��ɹ�����˲�ͬ�����ṹ���������ҩ�ﻯ���������Ч������ҩ��������װ�����н�Ϻ���Ƭ�ο��Դ���ҩ����װ��һ����ˮ���γ��ȶ������ṹ��������Ϊ������װ���⣬����ʵ�ְ��鳲��������Ӧ���ͷŵ�Ŀ�ġ��ڷ���ˮƽ�ϲٿ�PDCs����ζ�ſ��Ծ�ȷȷ��ҩ��ż����Ӧλ���������λ�ã��Ӷ�����PDCs�Ľṹ�������漰PDCs��ǰҩ���Կ�����������ҩ�������ڵ����ա��ֲ�����л���ͷš��ٽ�ҩ�������ڵĴ�л����й��ADME�����Ӷ���߸���ҩ�����Ч����������ҩ�������������ԡ�
�������ĵ�һ���ֱ����˹������ļ��乹����PDCs���о���չ���ڶ������ص�����������廯ѧ��ƺͺϳɵ����½�չ�����������ص��о��������ǿ������ǿ����Ӧ��ǿ�İ���/�����ͷ����ܵ���ϵͳ�����IJ�ͬ�����Լ����ڿ������ٴ�����ε�PDCs�����ǵ�Ŀ���ǰ����������ջ����ĵ�����ҩ�����ϵͳ�Ķ���ԡ���ˣ����Ľ����漰�������ĵ�������
2 �����Ľṹ��
PDC �Ķ��ֹ�����Ҫȡ�������������ԡ����磬ˮ����������������ˮ��ҩ���������ܽ�Ⱥ�ҩ���ҩ�ԣ�����װ���γ� PDC ���ṹ�������ĸ��ư�λ�ֲ���������ǿ����Ĥ���ԡ��Ѿ��о���ʹ��һϵ�й����ĸ��� PDC �����Ч������Щ���ӽ��ں������ۡ�
2.1 ����װ��
����װ��������һ�ֻ�����������ϸ��Ĥ���γ�[8]���������۵�[9]�Լ� DNA ˫�����ṹ[10]���������������ɲ���֮һ������������ӣ������ģ����Ƕ�ܵĹ����飬���γ���ȷ����ṹ������װ����Ƶ��ص����˽���ַ��Ӽ�����ã������������������á���-�жѻ������»�����ɫɢ��������-���帴�����������������ˮ�Ժ��ų���������[11��12] ���⣬ͨ�����װ�����������Ҳ������������������װ[13]����Ҫ���ǣ�������Щ������Խ��Ļ�PDCs��װ�ɾ���Ŀ��ṹ�ĵ���ϵͳ�ǹ�����������װ�ĵĵ���ϵͳ�ľ����Բ��衣�ڱ����У����DZ���������װ�ĵ�һ���Ͷ����ṹ�������о����ܽ�������װ���о��Ľ�չ���������˵���PDCs��̬�IJ��ԡ�
2.1.1 ����װ�ĵĽṹ����
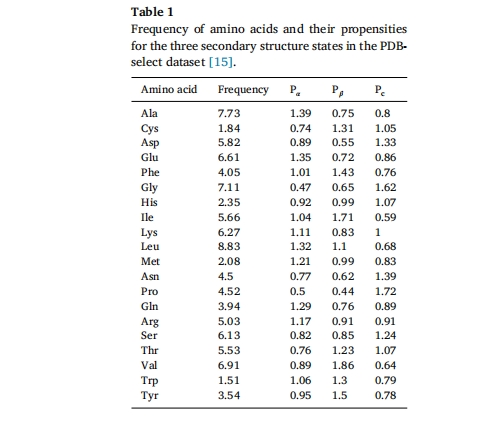
����װ�ĵĽṹ�����ĵİ��������е�һ���ṹ�����ڻ�������ڰ�����֮��ķ���������ã���������������-�۵�����ʽ��������ȶ����ṹ���Լ������ض��۵��ṹ�ķ��Ӽ���װ[14]Costantini �����ܽ��� PDBselect ���ݼ��а�����Ƶ�ʼ������ṹ״̬���ƣ��� 1��[15]
���ĵİ��������ͺ���������������װ�����Ļ�����������װ�����Ķ��ķ��ӽṹ�����������Ե��������������ԡ������ԡ���ɢ��ɡ���ͬ��ɽ������м��Գ��Եȡ������Զ�����Ϊһ������Ķ��ģ������봫ͳ������Լ����Ƶ����ʣ������Զ���ͬʱ������ˮ���ֺ���ˮ���֣����������������������ˮƬ�ε���ɿɷ�Ϊ��ˮƬ��ȫ���ɰ�����л���ɵı�������ģ��������Լ��ģ�����ˮ���ɳ����̼����ɵ������Զ��ġ���������ĵ���ˮƬ��һ�������ɸ���������ˮ������л���ɣ���ˮƬ�κ���һ����������һ������������ɻ�ɵİ�����л�����ṹ�ʹ�С����Ȼ��֬ʮ�����ơ������������Ķ��ԣ���������װ��̬�Ĺؼ�������ˮƬ�κ���ˮƬ�ε�ƽ�⣬ͨ�����ڰ���������ࡢ���������λ�����ı����������ʣ��Ӷ�������װ������սṹ��ṹ�ʹ�С����Ȼ��֬ʮ�����ơ������������Ķ��ԣ���������װ��̬�Ĺؼ�������ˮƬ�κ���ˮƬ�ε�ƽ�⡣ͨ�����ڰ���������ࡢ���������λ�����ı��������ԣ����Կ�����װ������սṹ��ṹ�ʹ�С����Ȼ��֬ʮ�����ơ������������Ķ��ԣ���������װ��̬�Ĺؼ�������ˮƬ�κ���ˮƬ�ε�ƽ�⡣ͨ�����ڰ���������ࡢ���������λ�����ı��������ԣ����Կ�����װ������սṹ[16�C18]��������Ҫ����һ�ຬ�з����就����Ķ��ģ���Phe��Tyr��Trp�ȣ����ǵķ����Բ���������������װ�������������ҿ��Ե�����ϵ����ˮ�ԣ���������Ϊһ��ǿ����ã�������߶��ķ��ӵ�����װ����[19]֤���˶Գ���Ҳ������װ���ӵ�һ���ؼ����أ���Ϊ�Գ��Լ���������װ����ƴ��ʱ�Ľ�������[20��21]��
2.1.2 ����װ��
���Ŀ����ڶ��ַǹ��ۼ����Ĺ�ͬ�����£�����ˮ���á���-�жѻ��������������������װ�γɲ�ͬ��ò���ܵ���װ�塣��Ȼ��Щ�ǹ��ۼ�����������Խ����������ǹ�ͬ����ʱ��������ǿ�Ȳ��ݺ��ӡ�����̽�����ĵ�����װ��Ϊ����װ���γɵĻ��ƣ���Ҫ�������˽���ַǹ��ۼ�������װ�����е����ã�Ϊ���������װ�����ò�ͳߴ��ṩ���ݡ�
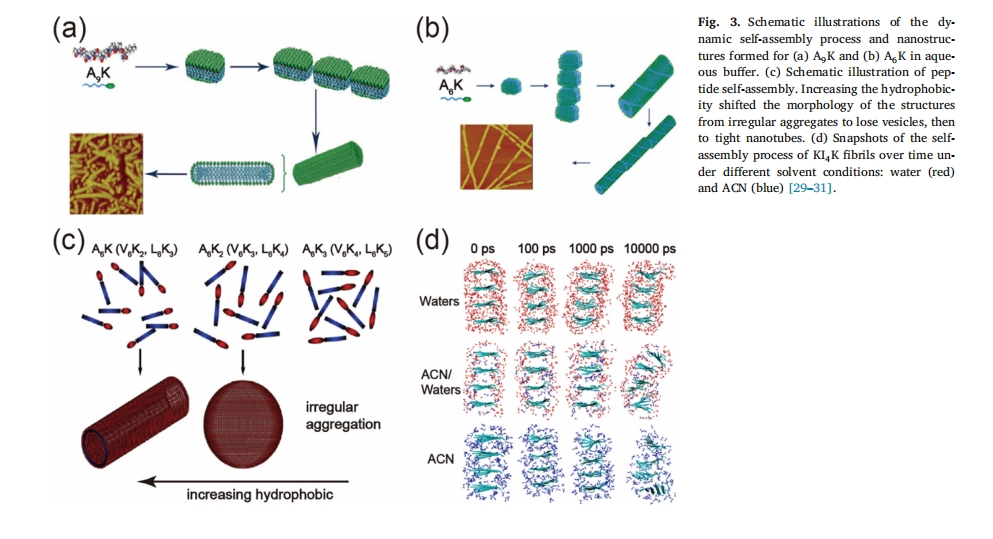
��ˮ�������������װ����Ҫ������[16],��ˮ���ı�������ЧӦ[22]����ˮ����õ�������Խ�С��������ԼΪ4�C8 kJ/mol[23],��������ˮ�л��������ᡢ�Ӱ��ᡢ�����ᡢ�������ᡢ�Ұ��ᡢ���������ɫ���ᣩ�ı���������������Ӱ�������װ������ṹ��һ����˵����������ͷǼ���������ɵĶ������и������γ����νṹ���Ǽ�����������ˮ���������¾ۼ��γ���ˮ���ģ���ˮ���������������������ˮ�Ӵ����ڸ߳����Ƚṹ����ά����Ĥ�����γɹ����У���ˮ���Ǵٽ���-�۵�����ѻ���ǰ�ᣬ����-�۵�����ѻ�������װ����ȵĽ�һ������������Ҫ[24��25],Groot�ȱȽ���Ile-Phe-��Val-Phe��������װ���ĵ�����װ��Ϊ������ǰ�ߵ�����װ�������ں��ߣ���֤ʵ��������Ile-�л�����һ��CH3��Val�л��ߣ���ˮ���ý��ͣ����յ���������װ�����½�[26],������װ�ķ��ӵ�C�˺�N������֬����������Ч�������װ�ṹ���ȶ���[27]Löwik���˷��֣������֬�����ij��Ȼ�Ӱ����ˮ����õ�ǿ�ȣ��Ӷ��ȶ�����ǿ����װ��������ͬ[28]���о�֤����ͨ�������ĵ���ˮ�Ի������ˮ�����������������װ���γɵ����ṹ���Դ���������ת��Ϊ���ܺ����״���Wang��[29]. ʹ�������� A6K��A9K ̽������ˮ����ö��ķ�������װ�ṹ��Ӱ�졣���ǹ۲쵽 A9K ��������װ�ɵ��ݺ�Ƚṹ�����������������װ���ͼ 3a).Ȼ����������ˮ�ȵĽ��ͣ�����ͨ��A�γɸ��ݺ�ȵ���ά�ṹ6�أ�ͼ 3b) ����������ˮ������ʱ������װ�������ھۼ��γ����νṹ�����֮�£�Meng ����[30]. ������ˮ����õ����ӻ�ı���װ�ṹ����̬���Ӳ�����ľۼ����Ϊ�������ݣ�����Ϊ���ܽṹ��ͼ 3c), ��һ����ì�ܵĽ��ۿ���������ͬ�ֵ�ɼ���ڽ�ǿ���ų���������Ϊ��ˮ��ֵ�K��������Խ�࣬�������ų�����Խ����ˣ��ĵ�����װ��ò������ˮ����ú;����ų�֮���ƽ������ġ�Zhao et al[31]. �۲쵽���� (ACN) �ƻ���ˮ����ã����������������װ��Ӱ�졣��ͼ��ʾͼ 3d, KI ��ԭ��ά4K���ĸ���-�۵���ɣ�����I4�� �۵�Ƭ����ˮ���ĵ����á���ˮ�У��۲쵽�ķ��ӽ��ܶѻ��γ�ԭ��ά����ά�ڲ��䵱��ˮ���ģ�ˮ���������ڴ����� �� �۵�Ƭ֮���ݳ���Ȼ����������/ˮ����1:1 Ħ���ȣ��������У��ķ��Ӷѻ���̫���ܣ��ܼ�����Ҳ�ֲ�����ˮ�����С����Ӷ���ѧģ���һ��������������ӿ��Խ����ĵ���ˮ���ֲ�������ˮ����ã��Ӷ��ƻ������۵�Ƭ֮���������[32]��Щ���صĽṹ����������Ϊ֧�ܹ���������������������������װ��ҩ�ʵ��ҩ��Ŀ����ͷš�33��34]��
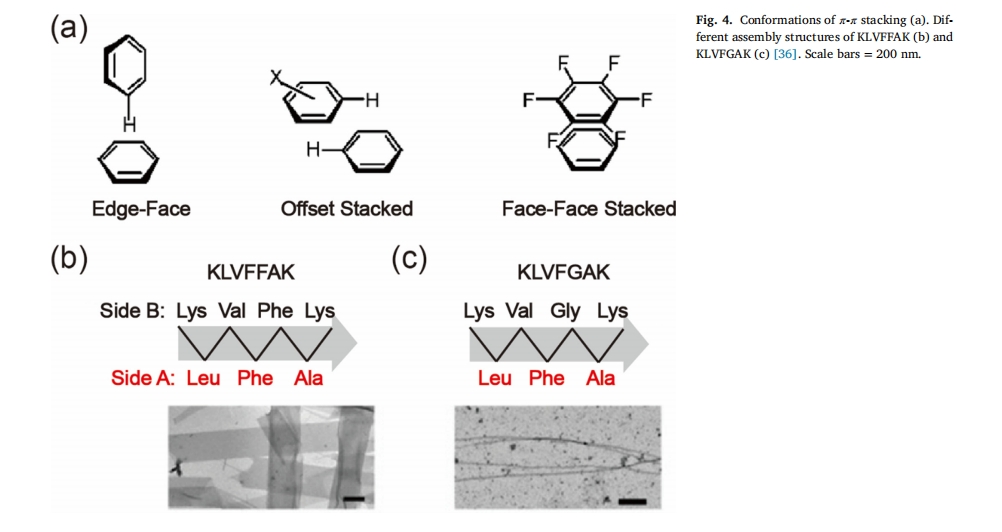
��-�жѻ�������ֳƷ�������ã���Ҫ�����ڦ�-�й�����ϵ�У�������Ϊ8�C12 kJ/mol[35],��һ�����ˮ����ò�ͬ����-�л�������þ��з����ԣ��漰�����ض��ռ����е���ˮ����á�ͨ�������ֻ�����ʽ�������߶�������á�ƫ�ƶѻ�����������ѻ����νṹ��ͼ 4a��[36],��-�жѻ���Ҫ���ɷ����就����л����籽������[37��38], ɫ����[39]���Ұ���[40]) ���������С�����������ŵ�����[41]������ Fmoc ����[42]�Ͷ�ï��[43]��Ҳ�ܲ�����-�жѻ����ں��з��㻷���Ľṹ�У����ֻ�������ѻ���������װ��Ķ��������������γɸ��ӽ�����������ṹ[44��45],��-�еĻ��۲���������ķ��ӵ�����װ����[46��47] ������һЩ�����Ե��������ۼ����������ž���������[48��49],���磬�������߿�Ķѻ����ٴ��жϰ����ĺ�Ĭ����AD���Ľ��[50],��ʹ�����������Ծۼ�����Ҫ�����ǵ������з����就����л�֮���-�м��Ļ��ۣ���ˣ��ƻ������������Ӽ�Ħ�-������ó����������Ƶ������߿���γɣ���̽��AD�����Ʋ��ԡ�KLVFFAK��KLVFFAE A�£�16�C22����һ��ͻ���壬��KLVFFAK�Ķ��еĵڶ����������ᱻ�ʰ���ȡ����KLVFGAK����-�жѻ����ñ�KLVFFAK���ö࣬��˺������������������Լ���������װ�ɽϿ��Ĵ�״�ṹ��Ϊ��խ��������ά�ṹ��ͼ 4b, c)[51],���⣬��-�жѻ����Խ�������װ�ĵ��ٽ�ۼ�Ũ�ȣ������װ�ṹ���ȶ��Ժ�������ѧ����[52��53]��
�����Եľ���������ͬ��������������ų⡣��������Դ���������е����Ի�������ᡣ����������Ȼ�Ƚ���������Ϊ0.5��4kJ/mol������������װ��Ӱ��ϴ����Ծ�����Ϊ�������Ľṹ�У�ͬ��ɲ����ľ����ų�����������Ϊ������װ���������ȶ���������Ƭ��һ��Ϊ��ˮ������ˮ���ͣ���ˮƬ����ˮ̮�������ۼ��γ��ںˡ����γɵĽ����ijߴ�ȡ������ˮ�˵ı�����������ˮ��������Ƭ�εľ����ų�֮���ƽ�⡣��ͬ�ֵ���ų��෴������ɺ����֮��ľ����������Կ����ķ��ӵľۼ���Ϊ����ʹ����������ɺ���ɲл��˴˽ӽ��ķ�ʽ����װ��Ramachandran ���֣��ھ��������������£����෴��ɵ��Ŀ����Է��ۼ�����װ��һ���γ��ܹ�����ҩ����ܵ���������������ɼ�ľ����������Կ����ķ��ӵľۼ���Ϊ��ʹ������װ��ʹ������ʹ�����IJл��������Ramachandran���֣��ھ��������������£����෴��ɵ��Ŀ����Է��ۼ���װ��һ���γ��ܹ�����ҩ����ܵ���������������ɼ�ľ����������Կ����ķ��ӵľۼ���Ϊ��ʹ������װ��ʹ������ʹ�����IJл��������Ramachandran���֣��ھ��������������£����෴��ɵ��Ŀ����Է��ۼ���װ��һ���γ��ܹ�����ҩ����ܵ���������[54]�����෴��ɵİ�����л��ֲ��������е�����ʱ����������װ�����������ڲ��÷���ƽ�еķ�������[55��56],Stupp���о�֤ʵ�����෴��ɵİ�����л��侲�������������γɵ�������ά���и�ǿ�����ȶ���[57]. �������ؿ���ģ���о���������װ�����о�������ö�ά����װ��ò����Ҫ�ԡ����������������ά��ˮ������簱����л�֮��ľ�������ÿ����ȶ���ά�ṹ[58]ͨ��̽�����Ӿ���ɶ�����װ��Ӱ�죬�������徻���Ϊ1������������װ��ά���γ�[59]��
����װ��ϵ��pHֵ��Ӱ�쾲��ЧӦ����Ҫ���أ�����pHֵ�ᵼ�¶��ĵ�ɵı仯�������ı���ĵ����з�ʽ����������װ����ά�����ܵ�ת�䡣���ڴ˹��ɣ���Ƴ��������װ���ӻ����ķ���KFFE[60]. Honggang Cui ����֤������ EF4E �� KF4��һ�ֵ�ɵ� K ֻ����ͬ�ֵ���ų��������γ�խ��Ť������ϸ��ά����Ȥ���ǣ������෴��ɵ� E �� K �л������뵽ͬһ�� EF ��ʱ4K�ģ����������кͣ������˷��Ӽ�ľ����ų⣬��װ����������������γ�һ������������ǰ���ᵽ�IJ�ͬ������Ӧ��װ����γɻ�����ͼ��ʾͼ 5a[61]��
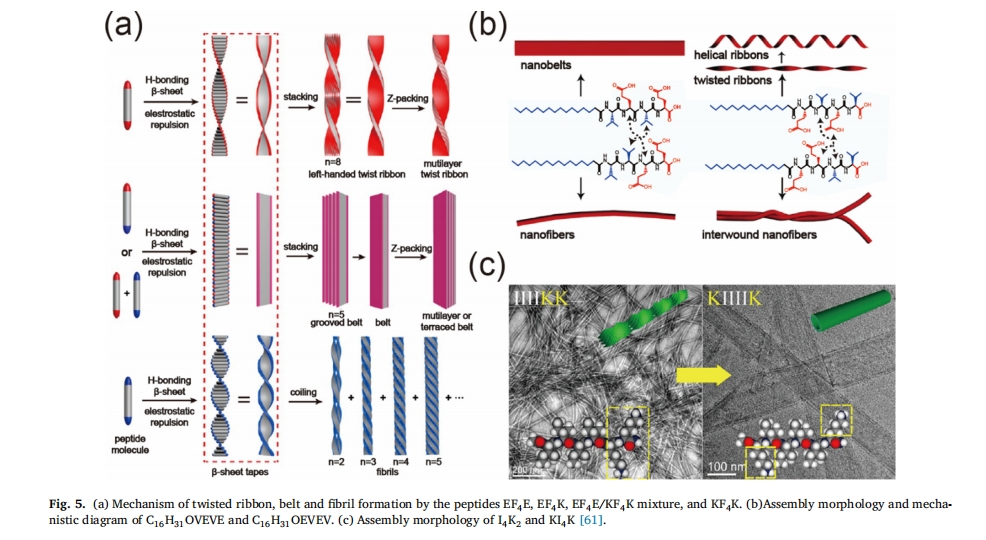
�ĵİ�����������������װ�ṹ�������[32��62],ͨ���ı��İ�����л������У����Եõ���ȫ��ͬ��һά���ṹ[62]. ��ͼ��ʾͼ 5b����C16H31 OVEVE�;��н�����ˮ����ˮ������л���C16H31 OEVEV�����γɸ����Ĵ������֮�£���C16H31 OVVEE �Ͳ��������е�C16H31 OEEVVֻ���γɽ�ϸ����ά��������ΪVEVE��EVEV�Ľ������п��Ժܺõط�ɢ��ɣ�����EE֮����ų�������EEVV��VVEE�����������У��������簱����EE�˴˿����������ų�����ǿ��ͬ�����о������֣�����I4K2�� KI4K ������ͬ����������İ�����, ����װ��̬�������Բ��� (ͼ 5c). ����I4K2������һ�˴������K�л�������ϵ����ǿ�ҵľ����ų⣬�Ӷ�������-Ƭ��֮�����ˮ���������Ӷ��γ�ϸ������ά���෴��KI����л�4Kλ�ڷ������ˣ��ԳƵĽṹ�������˵�K����ľ��������������������ˮ������ܾ���ЧӦ��Ӱ���С��
�����һ������ķ��»��������з����Ժͱ����ԡ�ͨ������£����ǿ�ȴ��ڷ��»�����С�ڹ��ۼ���������֮��������ԼΪ4�C12 kJ/mol[23]�����Ľṹ�к��н϶��C=O��NH���ţ������γɽ϶�������N-H��O���������Ӱ���Ķ����ṹ����Ҫ���أ���Ϊ����������ͷ��Ӽ�������봫ͳ������Լ��Ľṹ��ȣ��������������������ڷ��ӹǼ��д��ڴ����������ʹ��װ���س��᷽����������Ҳ��Ϊʲô������Լ�һ���������γ����λ��״�ṹ�������������γ���ά״�ṹ��ԭ��Zhu et al[63]֤�������ʹ���н�����ˮ�Ժ���ˮ������л������γɦ�-�۵��ṹ���ýṹ��ֱ����ά�ij������У��Ӷ�������ά��Ť������������ˣ�����������Ħ�-�۵����γɵ����ķ�������װ�;ۼ����γɾ��и߳����ȵ�Բ����������ά�����״�������[63�C67],Pahrump ���˷���������ά�������ĸ�������л�֮��ķ��Ӽ��������ʹ���������γɴ�ֱ��������ά����� �� Ƭ��ṹ���ɴ˽�ʾ�˸�������ķ����������γɸ��ݺ�ȵ���ά�ṹ[68],���֮�£�Զ����ĵİ�����л�������װ��ά����ά�ȶ��Ժͽṹ���Է�������ý�����������ƻ���ı䵼��Բ����������ά�Ľṹ�����仯�����糤��ά�ļ�����״�����ı䣬�������ν�������������װ����̬��ʧ[69]��
�ĵ�����װ���ɶ��ַǹ��ۼ��������ģ���Щ���������ķ�����ͬ�����������Ӷ��γɲ�ͬ��ò����װ�塣ֻ���ڳ�����������ǹ��ۼ����Ļ����ϣ�����ͨ�����ڸ��ַǹ��ۼ�������Դ�С����Ч������װ�����ò�ͳߴ硣
2.2 ����PDCs�Ŀ�ת������ҩ��
��ҩ��������װ�Ľ�Ͽ��γɸ���������ϵͳ������Ӧ�����ԡ����������Ժ����ҩ���ȶ��ԡ����⣬��������̬��������ϵͳ������ʡ���Ч�ʡ�����������ϸ�����ֲ����൱���Ӱ��[70]���磬Լ100��������������������ǿ���Ժͱ���ЧӦ��EPR����Ѫ������[71],������������������֯��С�������ӣ�<20���ף���������������֯����Ĥͨ�Ժõ����ƣ�������̫С���ױ�������������������������[72]�������ο�����ȣ����и߳����ȵ�������ά�Ͱ�״��״��������λ������ϸ����ȡ���������������[73]���֮�£��ϴ����ά״�Ͱ�״���ṹ�����ױ���״��Ƥϵͳ��������ҩ�����˨������[74]����ˣ�Ѱ��һ�ּ��ɶ�����̬���Ƶ�������̬ת������ϵͳ�����˹㷺���о���Ȥ����������װ��PDCs������ͨ����������װ��ʵ����̬ת��������PDCs��Խ������Ч����
�����Խ��Խ����о���Ȥת�����PDC����װ���������ض��������Ƶ���Ҫ��������װ�ĵ����ṹ������������װ���ṹ���ݼ�����������Ӧ�Եط�����̬�仯������Ԥ�ں�֤ʵ�ɴﵽ���õ�Ч����������̬��ҩ��ADME��Ӱ�죬�о���Ա�����ڿ������ֿɱ����������ӣ�NPs����������Ӧ�����ѱ�����������PDCs�ijߴ����״�仯���������ڽ�����Ƿ���[75]��
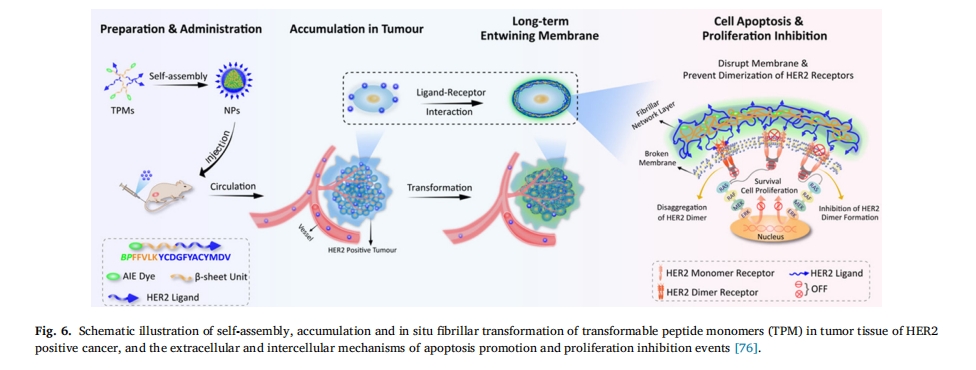
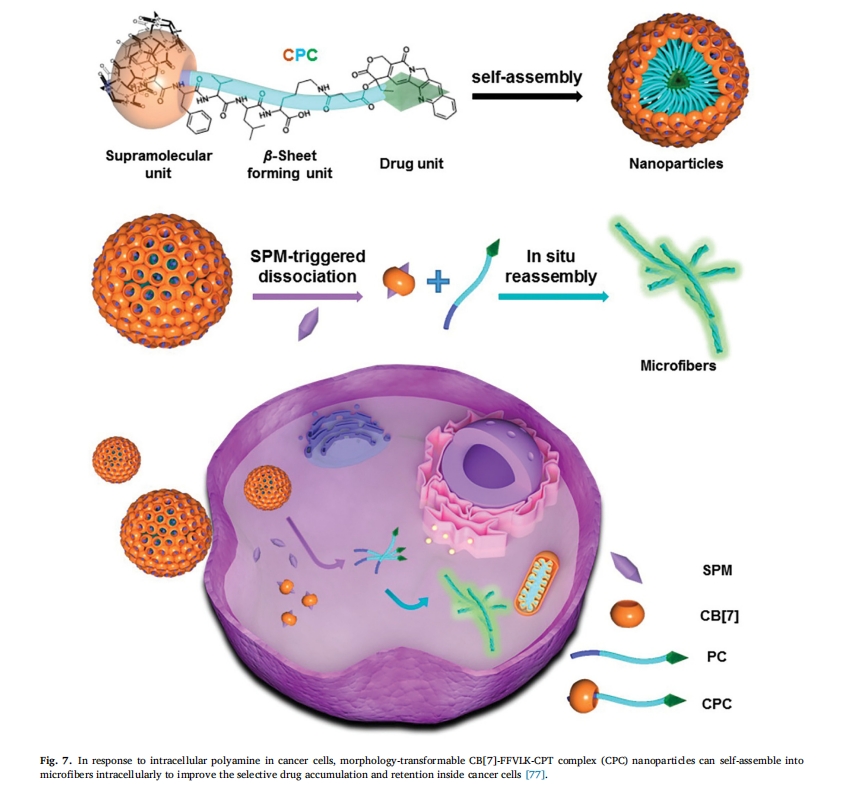
���磬Zhang������Ƶ�һ������ת����[76]����Щ���ܹ���ˮ��Һ������װ�ɿ�ע������ν�����ע���һ�������ϰ�ϸ���ϵ� HER2�����Ǿͻύ���γ�������ά���ڰ�ϸ�������γ���Ӧ��������ά�ĸ�����Ч���ƻ��� HER2 ���ۻ������°�ϸ��������ͼ 6). Chen �ȱ������ɶష��������̬��ת��ij�����������ҩ����ɳ����ӵ�Ԫ����«[7]�� (CB[7])������-Ƭ���γɵ�Ԫ���� FFVLK����ҩ�ﵥԪ��ϲ���� (CPT)����ɡ�FFVLK-CPT ������ͨ�� CB[7]-Phe-������������� CB[7] �ǹ��۸��ϣ��������װ�� NP���� CB[7] ͨ����ϸ���й������ϸ���ھ�������ʱ��NP ����ת��Ϊ��ά��������ǿ CPT �ڰ�ϸ���еĻ��ۺͱ���������������ϵͳ���ԣ�ͼ 7��[77]. ������[78]���������Ľ�ϣ���Ƴ�һ�־��пɸı���̬�ĸ�Ч���������硣�����������������յ��������緢����������̬�仯����������ֱ��Լ 100 ���ף�������Ƭ�����Ȼ����Ϊ���ף����������װ����Ի�е���ƻ�����Ĥ����ԭ�ı�ֱ�����͵�ϸ�����С�������װ������Ƭ��ͨ�������ض�����֢;������ǿ������������
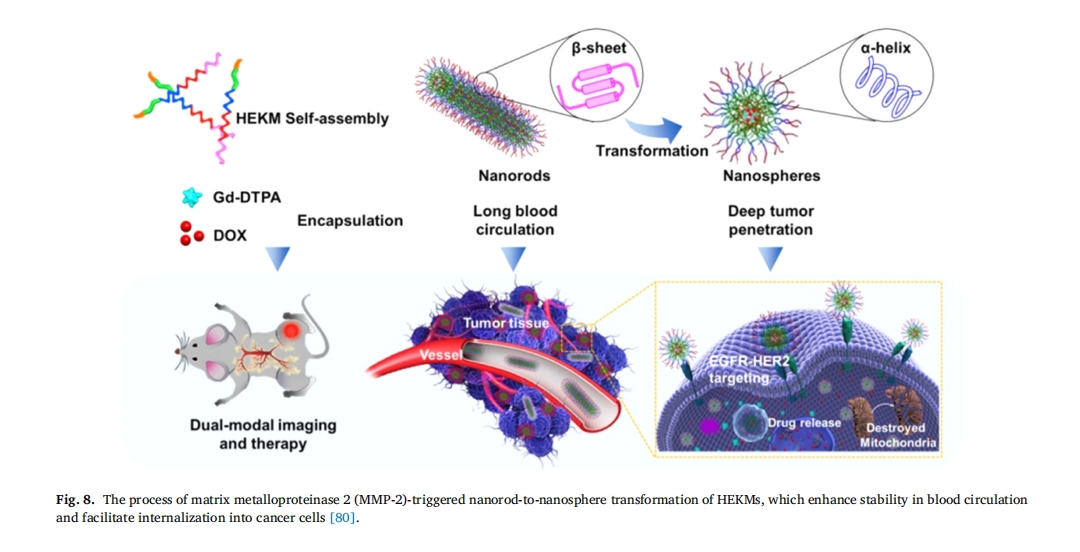
�����о���������״��������ѭ��ʱ��ϳ���������ҩ���ڲ����ڳ���������������������Ӿ������ϸ��������[79]����ˣ�Wang ����[80]. ������һ�ֻ����������ص����ܽ���ϵͳ����ΪHEKM����ϵͳ�ɰ�״������ɣ���Щ�����������������ڶ�ת��Ϊ��С�Ľ������������ν����㹻С�������������оۼ���ʵ������������ְ���ת����������ϵͳ��ǿ�˰�ù�أ�DOX����ϸ�������ԡ��ڻ��Ժ������������������������Ч����ͼ 8����
2.3 ������
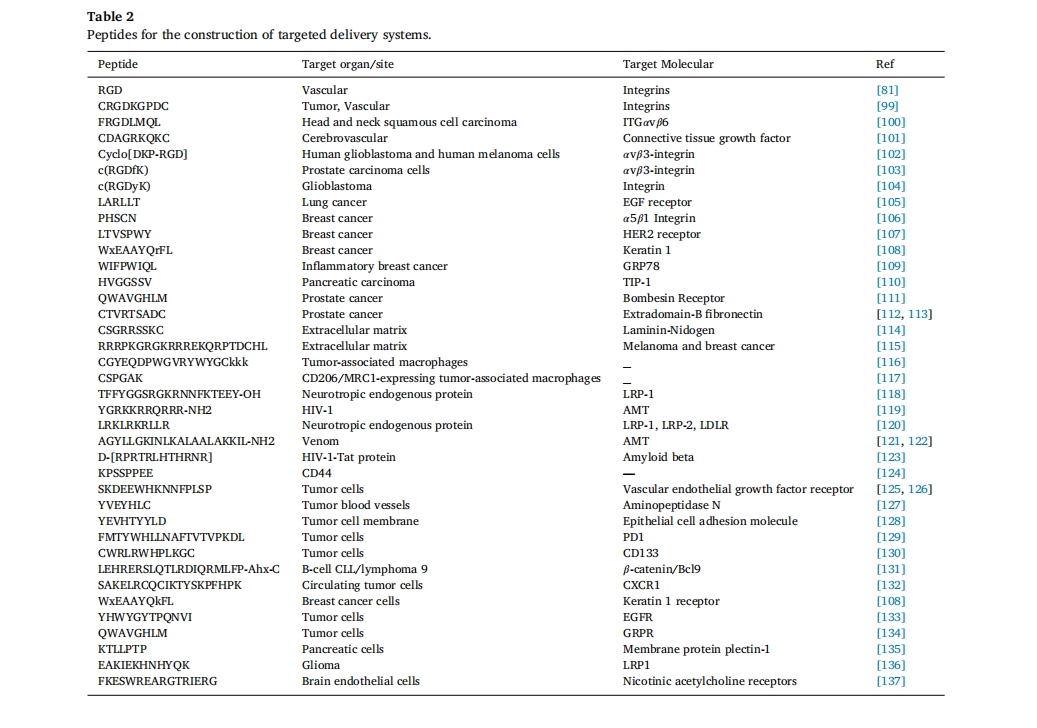
PDCs ��Ӧ��Ŀ����ͨ���ӳ��϶̵�ѭ����˥�ں���߰���Ч������ǿ����ҩ�����Ч�����õİ��������ͨ�� EPR ЧӦ��������������֯���Լ����ð�λ�߱���������Ա��������ʵ�ְ���ҩ����͡��봫ͳ����ȸ߷��������ӽṹ�İ���������ȣ�С���Ӱ������ܵ�Խ��Խ����о���ע������������Ѫ���Ϲ㷺������������ؼ����Ա������͵������鳲�Ļ����� RGD[81]. ��������[82]�ȱ�����5��1�����ؽ�ϼ�������RGD����״�ı��ֳ������Խ�ϸ�ǿ�Ľ��������������������������������RGD���У���ΪiRGD����i�������ڻ���[83]���ڹ�ȥ�ļ�ʮ���iRGD ����������ģ������ǿ��ҩ��Ļ��ۺ���Ч[84��85].2018�꣬iRGD��Ϊ����������abraxane�����ٰ����Ӽ�����I���ٴ����飨�ٴ�����ţ�NCT03517176����
��һ��������������ϸ������� (ECM) �����ģ���Ϊ ECM �ijɷ��ڼ�������ʵ�������ж����ȱ������ ECM �����ĵı������������������ B (EDB-FN) �ͼ��쵰��-C ����� [86��87], ��ճ������-��������������[88]�����ʽ�������ø�����ġ������֯�������� (CTGF/CCN2) �����[89]�Լ�Դ��̥���������� (PlGF-2) �� ECM �����[90]Hubbell ����ͬ��ͨ��ģ����Ȼ���忪���� ECM ����ģ���Щ���ڰ�֢���Ʒ�����������˷ܵ�Ӧ�á���һ�����о�DZ���������� Bicycle Therapeutics ��������-ҩ��ż���� (BT1718)[91]BT1718��˫��������ɣ�ͨ����������ѽ��������������ľ��ż�����ɰ���ECM�еĻ��ʽ�������ø14��MMP-14����BT1718��I/IIa���������ڻ�������ʵ�������ߣ����������ٰ��ͷ�Сϸ���ΰ����ٴ������ţ�NCT03486730�����С�
���⣬���䲿λ�����Ա���IJ�ͬ����Ҳ�ǰ����ĵ���Ҫ�е㡣���磬���ٰ����ѳ������ӹ���Ĥ����ǰ���ٰ��Ȱ�֢�й������LHRH���壬Ϊ����ҩ�������ṩ�˼��õIJ��ԡ���DOX��2-������-DOX��LHRH�����Ľ�ϣ��������Ӱ�ϸ���İ����ԣ�����ҩ�ﶾ�ԡ�Lindgren��[92]. ���� YTA2�� YTA4����װ����ʽ���γ�PDC��������װ�������ȣ���ϸ�����ʹ��ø����Ҷ�ỹԭø������������15-20����Դ��ϸ�����浰��ϸ����ճ������-1��ICAM-1�������п��Ա���Ѫ��Tϸ�������Ե����̡�ICAM-1�Ŀ���������ҩ�����ӣ��Ӷ��������ҩ���ڰ�֢���������������е���Ч�ԡ����ּ���������Ч�������˻��Ƶĸ����á�Jeffrey���˷���10��20������춬����Ȱ����ڰ�����۷���dz���Ч�����ǵ�ʵ���������������Ķ̹��Ŀ��Խ�����Ч�غɶ�λ�����۴�91���ȶ��շǰ�������Ч�غ���Ч 9 ��[93]. ����������Ľ���������о�������ѡ���Եؽ�����ҩ�����͵��������������ء������ء�С���� RNA (siRNA) �ͻ�������[94�C98]�� 2��
�ܽ������ڹ����������ϵͳ���ĵ������о���
2.4 ϸ������
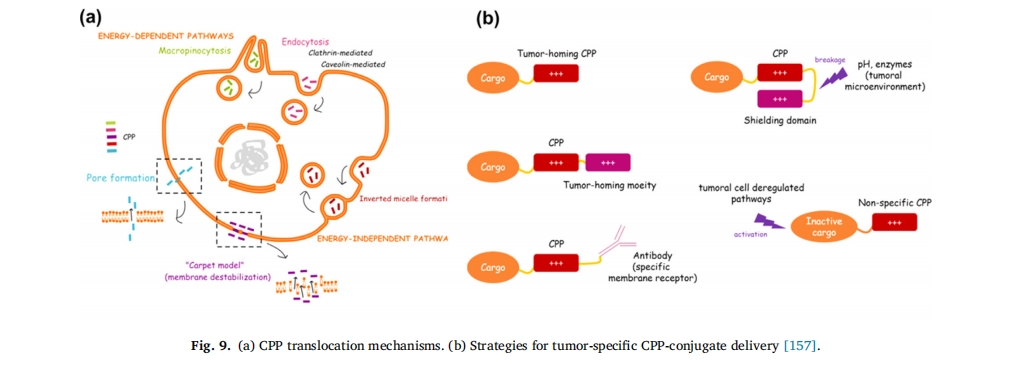
ϸ��Ĥ��һ����Ҫ���������ϣ����������൰���ʡ�������������Ӻ��������壨֬���塢������������������Ĥ���䣬������ҩ����ϸ��Ĥ��Ч����Ϊ��Ҫ��ϸ�����ģ�CPPs����Դ��ת�˵���ת���ṹ����һ���ܴ�ϸ��Ĥ��С���Ӷ��ģ�һ����5~30��������л���ɡ����ĸ��������ᡢ������ȼ������ᣬ�������������´�����ɡ������ԡ���ϸ��Ĥ����ǿ���ص㡣�����������ں�������ٵ���ҩ�P�������Խϸ��Ĥ���ϣ�������ǰ;��ҩ����������֮һ[138]��1994��Fawell�ȱ�����һ����Դ��HIV-1���ף�RKKKRRQRRR����ϸ������Tat�������ѷ��ֶ���ϸ�����ģ�����TAT��MAP��transportan����Arg/Lys�л����ź�ת����VP22��CPP�ڻ��Ķ���ģ���ѱ��������CPP��Ĥ��صı��㷺�ϿɵĻ�����Ҫ������������ֱ�ӿ�Ĥ���ƣ�����������ģ��[139]����϶�γ�[140]�Լ�����̺��ģ��[141��142]������һ���������������������ã�����С�Ѻ�����������������;��[143�C145]�;ް�������[146]��CPP �ڻ�ģ�͵�ժҪ���£�ͼ 9a��
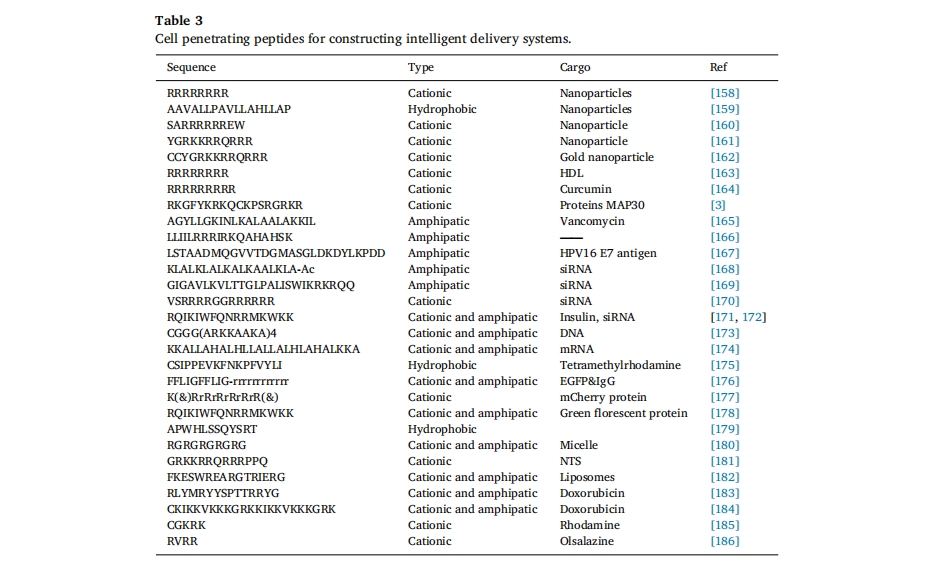
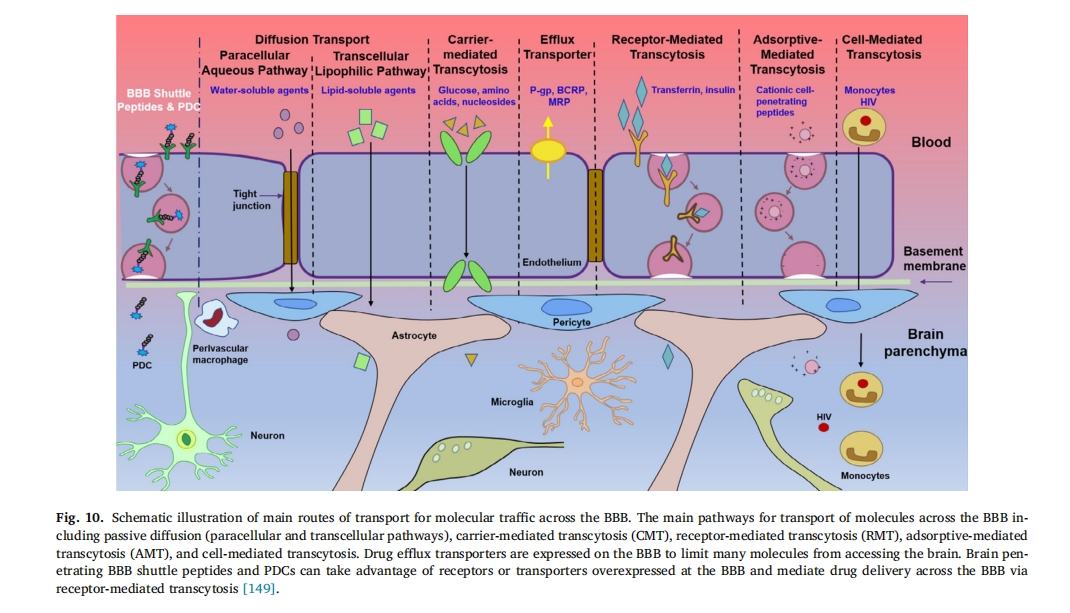
����CPPs���û�������������������Ч�Ĺ������������б��ϳ�[147]����������Ȼ�� CPP �о����ص㣬���������� CPP �������Ͳ�������ͼ 9b. ���⣬��ԽѪ�������� CPP ��������һ����ϣ���Ҿ�����ս�Ե������ѷ��ָ��� CPP �ܹ���������ҩ�����͵���ʵ���С���Щ CPP �������弴�ɴ�Ѫ�����ϣ��ɶ̵������Ժ�/��������������ɣ�����ͨ�������鵼��ת���̻��ƿ�Խϸ��Ĥ������[148]���⣬���� CPP ����������Ҳ�õ��˷�չ������ǿ�� BBB �������������ض������ǿ�����ͽ鵼ת�������õ�����[149]��ͼ 10)��Kwon���������һ�ֻ��ڴ���TAT-�춬����ø�����������ܰ�ϸ����Ѫ������Ч����ϵͳ[150]. ������ǿ��������֮�⣬CPP �������ڿڷ��ȵ��صij��ԡ��ݱ�����R8 �ı����������ȵ��� NPs �Կ˷�ճҺ����Ƥ���ϣ�ʹ�ȵ����������ö���� 1.9 ���������ʾ���ڷ���/�����ʵ��͵ľ�DZ��[151]��֮ǰ�Ŀ����Թ���������CPPsite 2.0 ���ݿ������Լ 1850 �� CPP ���У�Ԥ����һ���ֽ���������[152]. ���� 25 �� CPP ���ҩ�������ٴ������У�����������֢����ʹ����֢�����ಡ��˥�ϵȶ��ּ���[153�C156]�� 3��
�ܽ��˽���ʮ���������Ĵ���ϸ�����ĵ��о�����Щ���ѳɹ�Ӧ������ǿ�����ʡ������С����ҩ��Ĺ�Ч��
2.5 ��Ӧ��
��Ӧ���Ľ��ܵ�ϵͳ/�ⲿ�����仯��Ӱ�죬�Դ̼�������װ��ҩ���ͷţ������¶ȡ�pH��ø��GSH ������Ũ�ȵȣ�Ϊ��ҩ������ϵͳ�Ĺ����ṩ�ɿ��ԡ��¶���Ӧ����ͨ�����ڹ�����ע�������Ӧˮ����������ҩ�����ͺ�ϸ����װ�����磬���Ե��Ե��� (ELP) ���������� VPGXG �ĹѾ��ظ������ڵ��� 35��C ���¶����ܽ⣬������ע��������� (37��C) ���γ�ˮ��������ˣ�ELP ���Ŀ�ʹϸ�����������Ƽ�������������Һ�е�������ϣ����������¶����γ�����״���ʣ�������֯���̺�����ҽ��Ӧ�á��м��ּ����ᵼ�� pH ƽ��ı䣬������֢�Ͱ�֢����Ҳ���Ժ����������Ӧ���ԣ�Ȼ���ͷű�����ķ��ӣ�����ҩ������ʺͺ��ᡣ��ϸ���ľֲ�ϸ���� pH ֵ���ܱȽ���ϸ���� 0.5-1 �� pH ��λ��������ʹ�� pH ��Ӧ�Ķ����Ի�����֯�����ǽ�����֯���и��ߵİ�����������Ҳ���Ժ�����ƣ�ʹ�������������Ӧ��Ȼ���ͷű�����ķ��ӣ���ҩ������ʺͺ��ᡣ��ϸ���ľֲ�ϸ���� pH ֵ���ܱȽ���ϸ���� 0.5-1 �� pH ��λ����ˣ�ͨ��ʹ�� pH ��Ӧ�ģ����Զ����Ի�����֯�������ǽ�����֯��ʵ�ָ��ߵİ�����������Ҳ���Ժ�����ƣ�ʹ�������������Ӧ��Ȼ���ͷű�����ķ��ӣ���ҩ������ʺͺ��ᡣ��ϸ���ľֲ�ϸ���� pH ֵ���ܱȽ���ϸ���� 0.5-1 �� pH ��λ����ˣ�ͨ��ʹ�� pH ��Ӧ�ģ����Զ����Ի�����֯�������ǽ�����֯��ʵ�ָ��ߵİ���������[187]. ������[188]�ɹ��Ʊ���һ������ pH ��Ӧ������������װ�壻��������Ķ����ṹ���Դ� pH 7.4 ʱ�� �� Ƭ���Ϊ pH 5.0 ʱ�� �� �������Ӷ�������̬���������ݱ�Ϊ������ά��Matthew ���˱�����һ�� pH ��Ӧ�Ļ�������ά��װ�壬����װ����������������ø (GOx) ͨ�������γɵ����������ǡ�ȼ�ϡ������;ֲ� pH ֵ���Ӷ�������ƽ����ˮ�������硣�ڵ�Ѫ�Ƿ����ڼ䣬Ѫ���½����� GOx ����������������٣����������е� pH ֵ���ߡ�pH ��Ӧ��ˮ�������� pH ֵ�����߶��ܽⲢ�ͷ��ȸ�Ѫ���أ��ṩ������-pH ��Ӧ������άˮ���������������ȸ�Ѫ�������Կ���Ѫ��[189]��
һЩ��Ӧ���Ŀ���Ϊø�����ҩ��������Ի��ܵ�����ø��Ӱ�졣�ݱ���������ø��������λ��ȱѪ����ۼ���Ƶ�ʸ��ߣ����� MMP����֯����ø�ͼ�������ø��Jin Zhang ��[190]. ������MMP-2��ӦǶ���ģ����Ŀ�������������������װ������NP����øMMP-2��������λ������ʱ��ͨ������CACֵת��Ϊ��������ά�����ε���ά��ת��������������еĻ���Gd 3+Ũ�ȣ��Ӷ�����MRI�źš����⣬Jinseong Kim�����Ʊ���ǰҩNP[191].������װ�ĵ���øB�����Կ��ѽ��ģ�FRRG����DOX�������ɣ�ͨ����ϸ���й��ȱ���ĵ���øB�鵼��ø�ٽ����������ͷ�DOX��
3 ����Ƭ��
���ӻ����ӳ�PDCѭ��ʱ��Ϳ��ƻ�����Ӧ���ͷŵĹؼ���һ��ѡ�����ĩ�ˣ��������ĵ�N�˻�C�ˣ��Ͱ��װ���˻���Ϊ����λ�㣬����ĩ�����ӶԶ��Ĺ���Ӱ�첻���������ǰ�-���װ���Michael�ӳɷ�Ӧ���з�Ӧ����ѧ�졢ѡ���Ըߡ���Ӧ�����º͵��ص㣬���㷺����PDC�ϳɷ�Ӧλ��[192]�����⣬�����������������н�����һ�����װ���л���Ϊ��Ӧλ�㣬��˿������ż��Ч�ʡ������ӱ�����ѭ���б��ֳ��ȶ��ԣ��Է�ֹҩ�����ͷ��������ͷţ����PDCs��������������е��ȶ�������������Ҫ�����á������ӵ����ȡ�����漰PDCs�Ļ��ƺͼ������Ƶ���Ҫ�������ӵı仯�������ϵͳ���ֹ��ܣ��⽫������ϵͳ�����ܺ�ҩ�︺�ز����ش�Ӱ�졣����PDCs�����Ӽ����Ͷ��ֶ������ɷ�Ϊ���ѽ�Ͳ����ѽ���ʽ[193]���ѽ�����Ӽ�����ø��ѧ�鵼���ѽ��Լ��漰 pH �����Ĺ����� (GSH) ��������Ӧ�ѽ��4��
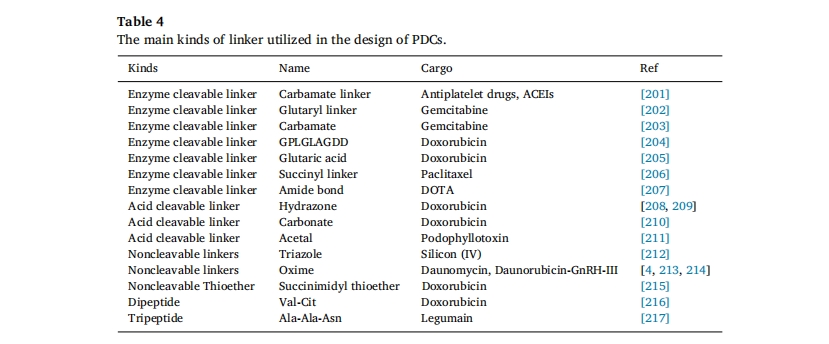
�ܽ���PDCs�������Ҫʹ�õ����������ͣ������������ڵ��ȶ����ѵõ�֤ʵ���Ӵ�С��˳��Ϊ������>����������>��>̼����[194]��
������֯ϸ����pHֵΪ7.4����ʵ��������֯��������pHֵ����������֯��195��196]�����������������º����ԣ�����pH���еĻ�ѧ�������ڿ�������ҩ��Ӱ�λPDCs�е��ͷš���Щ��������ѭ��pH�����ȶ��ģ�������������pH��ˮ�⣬������ȩ���ǰ����º�������Ӧ����Ƭ�Ρ�GSH�����ѽ�һϵ�пɲ����ͷ�Ļ�ѧ�������������������/������/�ڡ���������/���ڻ����ï���ͽ�����ͷ�������е�GSHˮƽ�ܸߡ���ҩ����ͷ��棬�ֲ����ӵ�GSHŨ�������ڿ�����Ч�غɵ��ͷš����ߵ�GSHŨ���Դٽ���ͷ���ѽⱻ���ڶ�������������ԡ�ø�������������� PDC ����һ�ֳ������ԡ����� Val-Ala �� Val-Cit �ǹ㷺ʹ�õ�ø������������ֶ���������ѭ���ж����ֳ����õ��ȶ��ԣ����ҽ�����֯����ø��������ø 1 �����²Żᱻ�и�[193]MMP �㷺���������ڣ��ѳ�Ϊ�ڶ��о������⡣���磬Tang ����[197]������MMP-2������(PVGLIG)��Ϊҩ����ɼ������ż������������У������µĸ�ҩϵͳ���Կ�����ɼ�����ͷţ��Ӷ��ڽ����������л�ø��õ�����Ч��������о��˶�MMP-2���еİ���(GPLGIAGQ)��������������λ�ͷŽ�����siRNA�ȣ�����ҩ���ڷǰ���֯���ͷ�[198��199���⣬������ͨ����ͷ�����������ҩ�︺��[200]��
4 ���ڿ������ٴ�����ε� PDC
4.1 ����
������Щ��ѡҩ�����ת��ΪPDCs�ı���Ҫ������ǿѭ���ȶ��Ժ����Ч�ʵ�֤�ݡ����ѽ��������ͷŵĻ��������һ�����е����ӵ㡣һ����˵����ѡҩ���IC50�ϵ�50�ܽ��Լ���[218].PDC ��ʹ�õĻ���ʾ������ DOX[219]����ɼ����PTX��[220�C222], ���ù��[223]�ͼ�������[224]�漰��Щ�������Ƽ�����Ҫ�ϰ���ˮ��Һ�е��ܽ��Բ��������֯�����Բ��Լ��Խ���ϸ���IJ���Ӱ���йء�Ϊ�˽����Щ���⣬��ͳ�ϻ�ʹ�ø��μ����ܽⲻ����ҩ������ֲ��Ի������һ�������⣬���������ܼ��Ķ������á����磬�������������⣬Kolliphor EL �����ڡ��ܽ⡱��ɼ�� (Taxol) ��Ϊ����������������Ӧ����Χ�����й�[225]������ҩ������ˮ���Ľ�ϲ������Ի��������ܽ�����⣬�������ṩ�������ܵĻ���[226]��
Ϊ�˿˷�����ҩ��DOXѭ��ʱ�������Ѱж��Եľ����ԣ�Chilkoti����[209]. ������һ�ְ����������ABD-DOX��PDCs��ʹ������λDOX�ĸ���Ũ������4�������ɽ�һ����DOX��ҩЧ����ʱ���ӳ�120����ˮ���Բ�Ͷ��Դ������˹��ʶ��أ�PPT�����ٴ���֢�����е�Ӧ�á��ܵ�[227]�����һ�ֻ����ĵ�PPT��������ҩ����ܽ�ȣ����������PPT����Ѫ���ԣ���������˿�����Ч�����������Ŀ���Ѫ�����⡢ȫ�����Ժ���ҩ�Դ�������������ò���������������Ч����������ĸĽ����滯�Ƶķ���֮һ�ǻ���PDCs������ҩ��ֱ�Ӱ���ϸ�����������˼�������������ָ�������⣬�����������������絰���ʺͺ����Լ��������壨֬���塢����������������˵��ϸ��Ĥ����Լ���ǿ�Ĥ�������Ҫ�������ϡ���ˣ����ҩ���ϸ��Ĥ��Ч��������Ҫ������ˮ��ҩ������ˮ������Ͽ��Ըı���Ӽ��ԣ���Ч���Ĥ������������ϸ�������ѱ��ɹ�������ǿ�����ʡ������С����ҩ��Ĺ�Ч�����磬������ɵ�ϸ�������ѱ��������Ӵ�����ɵĺ������ӣ��Թ���������ϵͳ�����Ƹ����Ŵ������������������ߺ������������ڵ��ȶ��ԺͿ�Ĥ���������ഩĤ���ѱ��ɹ�������ǿ�����ʡ������С����ҩ���ҩЧ�����磬������ɵĴ�Ĥ���ѱ��������Ӵ�����ɵĺ������ӣ�����������ϵͳ�����Ƹ����Ŵ������������������ߺ������������ڵ��ȶ��ԺͿ�Ĥ���������ഩĤ���ѱ��ɹ�������ǿ�����ʡ������С����ҩ���ҩЧ�����磬������ɵĴ�Ĥ���ѱ��������Ӵ�����ɵĺ������ӣ�����������ϵͳ�����Ƹ����Ŵ������������������ߺ������������ڵ��ȶ��ԺͿ�Ĥ������228��229���ϵ������һ�־���ǿĤ���ԺͿ��������ԵĴ���-PTXż�����ǿ��PTX�Ŀ��������ԣ�ͬʱ�˷���PTX�ܽ��Բ��������ҩ�Ե�����[230]��
4.2 �ٴ������е� PDC
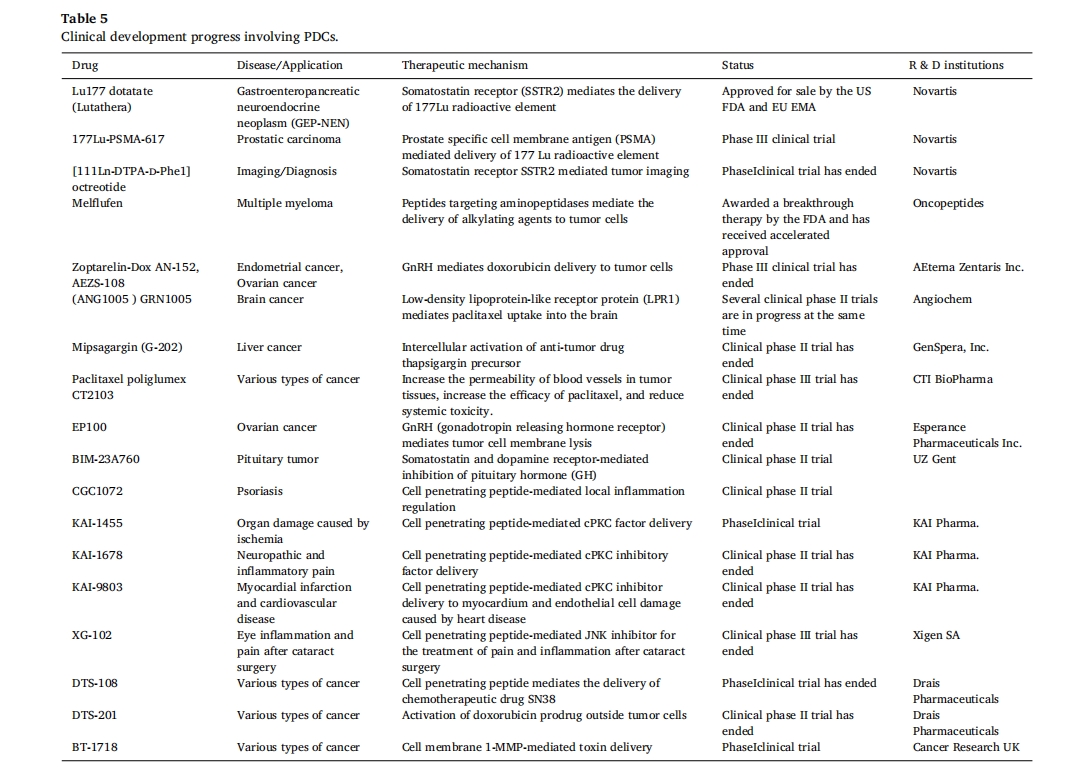
���� PDC �Ľṹ�뿹��-ҩ��ż���� (ADC) ���ƣ�����Щ��������ż����� ADC ���ж������ƣ�����ѡ���ԡ�������������������������������������PDC ҩ�� Lu-dotatate177 �ѻ������ʳƷҩƷ�ල������ (FDA) ����������θ�����������ڷ�������[231]��Ȼ�������и����PDCs���ڿ����Σ�����TH1902��һ��װ�ض���������PDC�������������������ٰ����ѳ�����TH1904��һ��װ��DOX��PDC�������������ѳ�����������PDC������֢��sortilin 1 (SORT1)����Ĺ��ȱ���������������ٰ����ѳ������ΰ�����ֱ������Ƥ���������ٰ�[232]��һ�ֻ��ڳ�Ϊ���г��ĵ������� PDC ҩ��Ҳ�dz��ܻ�ӭ�����г���ͨ������Ϊ 9-20 �������ᣬ���������������װ���л�����Щ���װ���л���С���ӽ�ͷ��Ӧ����������Ϊ���Թ������г����ؽ���������ҩ����ӵ�ת�˵��ס�ҩ�︽�������г����ϣ���ȷ���������谭[233]. �������г��Ʒ��ѽ�������ض��������ٴ�����[234]�� 5��
�ܽ�������FDA��վ�Ϸ������漰PDCs���ٴ��о���չ���棨https://www.fda.gov/����
5 ���ۺ�δ��չ��
������ǿ����PDC��Ϊ�����Ľ�������ҩ������ϵͳ�Ķ��������ߵ�DZ�������������ںϳɡ����ܶ�䡢ҩ�︺�ع㷺����̬�����ԺͿɵ��ԡ���ADC��ȣ���������������Ƭ�εij�ɫ�����ƣ�PDC���и���Ļ���Ϳ����ԡ����ܾ�����Щ���ƣ�PDC�ڽ����ٴ�ʱ������������ս����Ҫ��ս֮һ�����ȶ��Բ����������ٶȿ졣PDC������ѭ���б����ȶ����Է�ֹϸ��������Ч�غɵ�Ԥ���ͷź���֮������ȫ����¶��Ϊ������ʵ������ͨ����Ҫ������PDCѭ��ʱ�䣬�Ա����㹻��ʱ�佫PDCs������������֯������NP��װ��PDCs��PEG���������ӳ�PDCs�����ڰ�˥�ڵ����������IJ��ԡ����˴�ͳ�ļ��ͺ�ҩ;�����羲������ע�䣩�⣬��Ӧ̽���µĸ�ҩ���ԣ����ǻ������PDC��ҩ���Ա���PDC���⡣�ڿ���������Ȼ����ƺ��Ż�����Ч�Ŀ����Ļ�ҩ�﷽��ȡ����Խ��Խ��ijɾͣ������PDCs��ѡ���Եذ����ض������ϸ�������ǹ���ϸ����Ȼ��һ���ؼ���ս������PDC�о�����һ��ؼ�������ȱ������PDC���Ե�ר��ҩ������ѧ������������Ч��PDCs��Ҫ��Ƴ����ж��ֹ��ܵĹ����ģ������˹�����ϵͳ�ĺ������ݷ���������̽����װ��ò�백�������ͺ�����֮��Ĺ�ϵ��ʵ��PDCs����װ�ṹ��ҩ���ͷŵľ����ء��ٽ�PDCs�Ŀ�ѧת��������Ҫ�ںϳɼ���������Ŭ���������������ʡ���֤ҩ����ȶ��Եȡ�ʵ��PDC����װ�ṹ��ҩ���ͷŵľ����ء��ٽ�PDCs�Ŀ�ѧת��������Ҫ�ںϳɼ��������¹������������ʡ���֤ҩ����ȶ��Եȡ�ʵ��PDC����װ�ṹ��ҩ���ͷŵľ����ء��ٽ�PDCs�Ŀ�ѧת��������Ҫ�ںϳɼ��������¹������������ʡ���֤ҩ����ȶ��Եȡ�
��������������Ϊ��ҵ����ѧϰ����Ȩ��ԭ����ԭ��־���У�������Ȩ������ϵɾ�������±�ע���������³����������Ķ�ԭ�ļ��ο����ף����Ķ�ԭ��־��