’Σ“ΣΘΚ÷«ΡήΙΙΫ®Ζ÷Ή”ΒΡΙΛ≥Χ…ηΦΤ «…ηΦΤ÷«Ρή“©ΈοΒίΥΆœΒΆ≥ΒΡΙΊΦϋΜυ¥ΓΓΘΉςΈΣ“Μ÷÷–¬–ΥΒΡΗ¥‘”ΒίΥΆœΒΆ≥≤Ώ¬‘Θ§κΡ“©Έο≈ΦΝΣΈο (PDC) ΒΡ«Ω¥σΙΠΡήΙι“ρ”Ύ÷«ΡήΫ”ΆΖΚΆΕύΙΠΡήκΡ”ρΓΘκΡ±μœ÷≥ωΙψΖΚΒΡΙΠΡήΚΆΧΊ–‘Θ§Αϋά®“Ή”ΎΜ·―ßΚœ≥…ΚΆΕύ÷÷–ό ΈΓΔΩ…ΒςΫΎΒΡ…ζΈοœύ»ί–‘ΓΔΕύ―υΜ·ΒΡΉ‘ΉιΉΑΡ…ΟΉΫαΙΙΓΔΧΊ“λ–‘ Ε±π/ΫαΚœ“‘ΦΑΕ‘œΗΑϊΡΛ/œΗΑϊΆβΜυ÷ ΒΡ…νΕ»…χΆΗΓΘ¥ΥΆβΘ§Ης÷÷άύ–ΆΒΡΫ”ΆΖ Ι PDC ΡήΙΜΗυΨίΦ≤≤ΓΈΔΜΖΨ≥Μρ÷ΈΝΤ–η«σœλ”Π–‘ΒΊ ΆΖ≈“©ΈοΓΘ”…”Ύ’β–©ΙΧ”–ΒΡ”≈ ΤΘ§PDC “―±ΜΙψΖΚ”Ο”Ύ“©ΈοΒίΥΆΓΘ‘Ύ¥ΥΘ§Ήέ ωΝΥΙΙΫ®÷«ΡήPDCs≥Θ”ΟΒΡΙΠΡήκΡΚΆΝ§Ϋ”Ή”ΒΡΉν–¬Ϋχ’ΙΘ§÷Φ‘Ύ“ΐΤπ»ΥΟ«Ε‘PDC…ηΦΤ≤Ώ¬‘ΦΑΤδΕ‘ΩΙΜςΗςάύΦ≤≤ΓΙ±œΉΒΡΙψΖΚΙΊΉΔΘ§≤ΔΈΣ÷«ΡήPDC“©ΈοΒίΥΆœΒΆ≥ΒΡ―–ΨΩΧαΙ©÷ΗΒΦΓΘ
1 Ϋι…ή
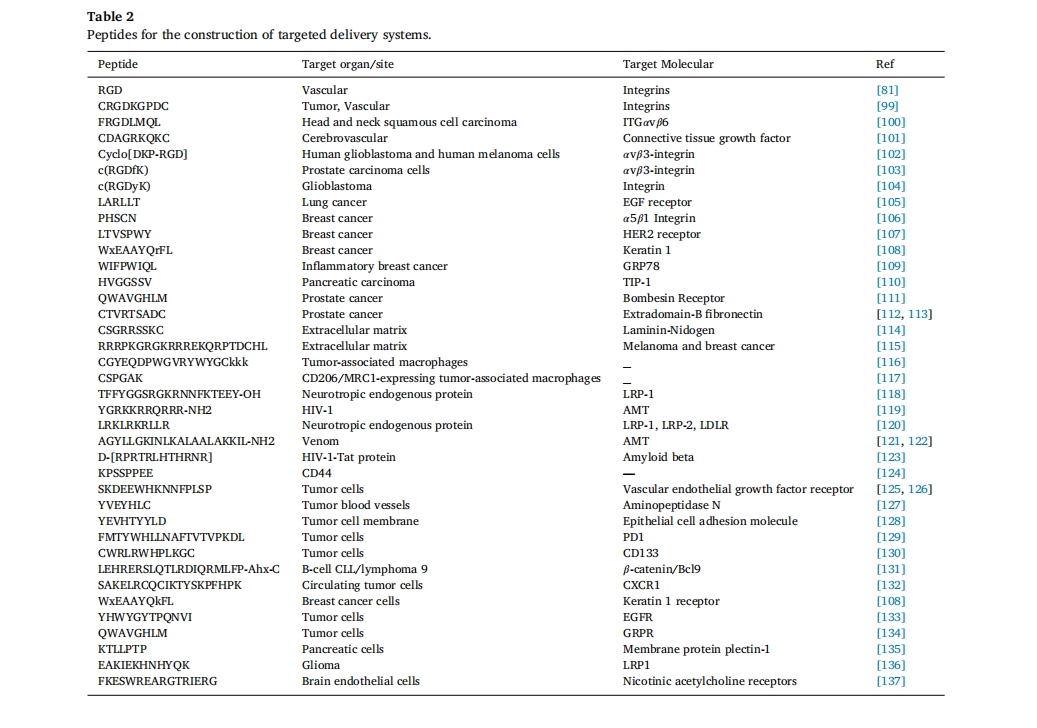
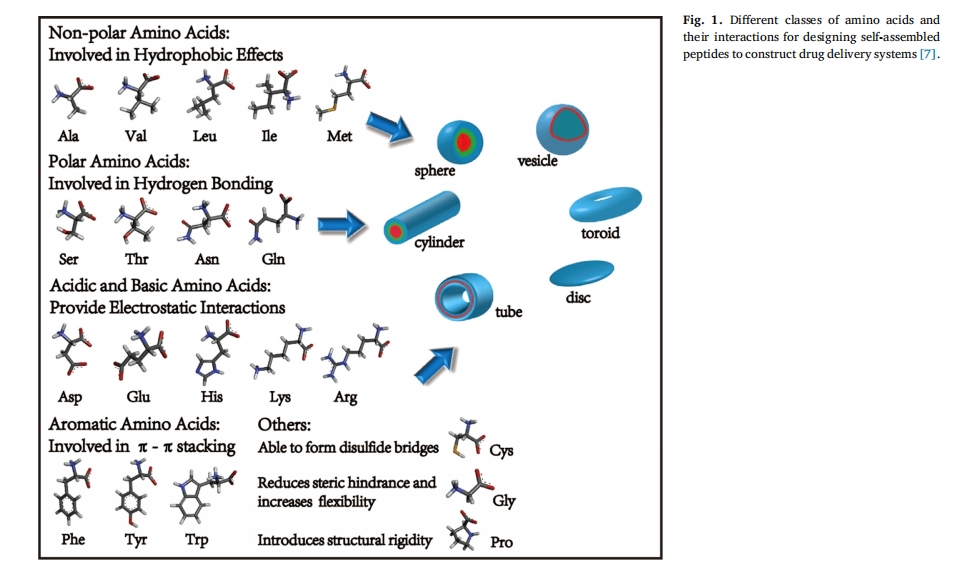
ΨΪœΗ÷«Ρή“©ΈοΒίΥΆœΒΆ≥ΒΡ―–ΨΩ÷Φ‘Ύ±Θ÷Λ“©ΈοΑ≤»ΪΓΔΩ…ΩΊΒΊ ΆΖ≈Θ§“‘¥οΒΫΉνΦ―ΒΡ“©–ßΚΆΉν–ΓΒΡΗ±Ής”ΟΓΘΕύκΡ”…”ΎΨΏ”–ΝΦΚΟΒΡ…ζΈοœύ»ί–‘ΓΔ…ζΈοΫΒΫβ–‘ΓΔΉ‘ΉιΉΑ–‘ΚΆ≤ΜΆ§ΒΡΜ·―ß±δ“λ–‘Θ§‘ΎΙΙΫ®÷«ΡήΡ…ΟΉΒίΥΆœΒΆ≥ΖΫΟφ“ΐΤπΝΥ»ΥΟ«ΒΡΦΪ¥σ–Υ»ΛΓΘ”Ο”ΎΙΙΫ®Ρ…ΟΉ“©ΈοΒΡΕύκΡΆ®≥Θ≥ΛΕ»ΈΣ2-30ΗωΑ±ΜυΥα≤–ΜυΘ§Ά®Ιΐ…ηΦΤ20ΗωΑ±ΜυΥαΦΑΤδ―ή…ζΈοΒΡ–ρΝ–ΚΆΫαΙΙΩ…“‘ΜώΒΟ ΐ«ß÷÷ΙΠΡήΚΆΫαΙΙΓΘΫΪΙΠΡή–‘ΕύκΡ”κ≤ΜΆ§ΒΡ“©Έο“‘Ι≤ΦέΜρΖ«Ι≤ΦέΖΫ ΫΫαΚœΘ§ΙΙΫ®Μυ”ΎΕύκΡΒΡΒίΥΆœΒΆ≥Θ§Ω…“‘≤ζ…ζΝΦΚΟΒΡ÷ΈΝΤ–ßΙϊΓΘ“ρ¥ΥΘ§ΕύκΡ‘ΎΜυ“ρΒίΥΆΖΫΟφΨΏ”–”≈ ΤΓΘ[1], Ι―ΚΥή’Υα[2]ΓΔΒΑΑΉ÷ [3]ΓΔ–ΓΖ÷Ή”Μ·ΝΤ“©Έο[4]ΓΔΈόΜζ“©Έο[5]Θ§œΗΑϊ[6]ΚΆΤδΥϊ÷ΈΝΤΈο÷ ά¥÷ΈΝΤΗς÷÷Φ≤≤ΓΓΘΗς÷÷Ης―υΒΡΑ±ΜυΥα≤–ΜυΘ§»γ«ΉΥ°–‘ΓΔ ηΥ°–‘ΓΔΖΦœψ–‘ΓΔΥα–‘ΚΆΦν–‘Α±ΜυΥαΘ§ΕΦΩ…”Ο”ΎΙΙΫ®ΙΠΡή–‘κΡ–ρΝ–ΓΘ’β–©≤ΜΆ§ΒΡΑ±ΜυΥαΈΣκΡΧαΙ©ΝΥΫαΙΙΜυ¥ΓΘ§“‘≤ζ…ζΗς÷÷œύΜΞΉς”ΟΝΠΓΘ¥ΥΆβΘ§‘Ύ–≠Ά§Ζ«Ι≤ΦέœύΜΞΉς”ΟΒΡ«ΐΕ·œ¬Θ§κΡΩ…“‘Ή‘ΖΔ–Έ≥…Ος»ΖΒΡΡ…ΟΉΫαΙΙΘ§Αϋά®Ρ…ΟΉΑτΓΔΙήΓΔœΥΈ§ΓΔ«ρΧεΚΆΧπΧπ»Π“‘ΦΑΫΚ χΘ®ΆΦ 1Θ©[7]
κΡάύ≥ΐΝΥΨΏ”–Η¥‘”ΓΔΩ…ΒςΩΊΒΡΉ‘ΉιΉΑΫαΙΙΆβΘ§ΜΙΨΏ”–Εύ÷÷Ζ¥”ΠΜυΆ≈Θ§Αϋά®Ζ÷ΫβΜυΆ≈ΓΔΑ±ΜυΜυΆ≈ΓΔέœΜυΜυΆ≈Β»Θ§ ΙΒΟΆ®ΙΐΙ≤ΦέΦϋΝ§Ϋ”“©Έο‘Υ δΗϋΦ”»ί“ΉΚΆΩ…––ΓΘ¥ΥΆβΘ§κΡάύΜΙ «“Μ÷÷≤Έ”κ…ζΈοΧεΡΎΗς÷÷œΗΑϊΙΠΡήΒΡ…ζΈοΜν–‘Έο÷ Θ§ΤδΙΠΡή «Ά®ΙΐΡΘΖ¬ΧΊΕ®ΒΡΒΑΑΉ÷ ΙΠΡή”ρ Βœ÷ΒΡΓΘ–μΕύκΡάύΩ…ΨΏ”–Εύ÷÷ΙΠΡήΘ§»γΑ–œρ–‘ΓΔΙι≥≤–‘ΓΔœΗΑϊ¥©ΆΗ–‘ΚΆ―ΣΡ‘ΤΝ’œ¥©ΆΗ–‘ΓΘ
κΡ“©Έο≈ΦΝΣΈοΘ®PDCΘ© «“Μ÷÷–¬–ΆΒΡ«ΑΧε“©Έο≤Ώ¬‘Θ§Τδ”≈“λΒΡΙΠΡή÷ς“ΣΙιΙΠ”ΎΕύ―υΜ·ΒΡκΡΫαΙΙ”ρΚΆœλ”Π–‘Ϋ”ΆΖΓΘPDC ÷ς“Σ”…»ΐΗω≤ΩΖ÷Ήι≥…ΘΚΙΠΡή–‘κΡΫαΙΙ”ρΓΔΜθΈοΚΆΫ”ΆΖΤ§ΕΈΘ®ΆΦ 2Θ©

ΫϋΡξά¥Θ§―–ΨΩ»Υ‘±άϊ”ΟPDCsΒΡ…ζΈοΜν–‘ΚΆΉ‘ΉιΉΑ«±ΡήΘ§≥…ΙΠΜώΒΟΝΥ≤ΜΆ§ΒΡΡ…ΟΉΫαΙΙΘ§”Ο”ΎΧαΗΏ“©ΈοΜ·ΚœΈοΒΡ÷ΈΝΤ–ßΙϊΓΘΫΪ“©Έο”κΉ‘ΉιΉΑκΡ–ρΝ–ΫαΚœΚσΘ§κΡΤ§ΕΈΩ…“‘¥χΕ·“©ΈοΉιΉΑ‘Ύ“ΜΤπΘ§‘ΎΥ°÷––Έ≥…Έ»Ε®ΒΡΡ…ΟΉΫαΙΙΘ§≥ΐΝΥΉςΈΣΙΠΡήΉιΉΑΩιΆβΘ§ΜΙΡή Βœ÷Α–œρΓΔΙι≥≤ΓΔ¥©ΆΗΚΆœλ”Π–‘ ΆΖ≈ΒΡΡΩΒΡΓΘ‘ΎΖ÷Ή”Υ°ΤΫ…œ≤ΌΩΊPDCsΜΙ“βΈΕΉ≈Ω…“‘ΨΪ»Ζ»ΖΕ®“©Έο≈ΦΝΣΖ¥”ΠΈΜΒψΒΡ ΐΝΩΚΆΈΜ÷ΟΘ§¥”ΕχΩΊ÷ΤPDCsΒΡΫαΙΙΓΘ’β÷÷…φΦΑPDCsΒΡ«Α“©≤Ώ¬‘Ω…“‘œ‘÷χΗΡ…Τ“©Έο‘ΎΧεΡΎΒΡΈϋ ’ΓΔΖ÷≤ΦΓΔ¥ζ–ΜΚΆ ΆΖ≈ΓΘ¥ΌΫχ“©Έο‘ΎΧεΡΎΒΡ¥ζ–ΜΚΆ≈≈–ΙΘ®ADMEΘ©Θ§¥”ΕχΧαΗΏΗΟάύ“©ΈοΒΡΝΤ–ßΘ§’β «άμœκ“©ΈοΒίΥΆ‘ΊΧεΒΡΧΊ–‘ΓΘ
±ΨΉέ ωΒΡΒΎ“Μ≤ΩΖ÷±®ΒάΝΥΙΠΡή–‘κΡΦΑΤδΙΙΫ®ΒΡPDCsΒΡ―–ΨΩΫχ’ΙΓΘΒΎΕΰ≤ΩΖ÷÷ΊΒψΫι…ήΝΥΝ§Ϋ”ΧεΜ·―ß…ηΦΤΚΆΚœ≥…ΒΡΉν–¬Ϋχ’ΙΓΘΒΎ»ΐ≤ΩΖ÷÷ΊΒψ―–ΨΩΝΥ’κΕ‘–‘«ΩΓΔ¥©ΆΗ–‘«ΩΓΔœλ”Π–‘«ΩΒΡΑ¥–η/¥ΞΖΔ ΆΖ≈÷«ΡήΒίΥΆœΒΆ≥ΙΙΫ®ΒΡ≤ΜΆ§ΜθΈο“‘ΦΑ¥Π”ΎΩΣΖΔΚΆΝΌ¥≤ ‘―ιΫΉΕΈΒΡPDCsΓΘΈ“Ο«ΒΡΡΩ±ξ «Αο÷ζΕΝ’Ώ’ΤΈ’Μυ”ΎκΡΒΡ÷«Ρή“©ΈοΒίΥΆœΒΆ≥ΒΡΕύΙΠΡή–‘ΓΘ“ρ¥ΥΘ§±ΨΈΡΫΪ≤Μ…φΦΑ÷ΈΝΤ–‘κΡΒΡΉέ ωΓΘ
2 ΙΠΡήκΡΫαΙΙ”ρ
PDC ΒΡΕύ÷÷ΙΠΡή÷ς“Σ»ΓΨω”ΎκΡ«χΒΡΧΊ–‘ΓΘάΐ»γΘ§Υ°»ή–‘κΡ”Ο”ΎΝ§Ϋ” ηΥ°–‘“©Έο“‘‘ωΦ”»ήΫβΕ»ΚΆ“©ΈοΩ…“©–‘Θ§Ή‘ΉιΉΑκΡ–Έ≥… PDC Ρ…ΟΉΫαΙΙΘΜΑ–œρκΡΗΡ…ΤΑ–ΈΜΖ÷≤ΦΘ§¥©ΆΗκΡ‘ω«Ω…ζΈοΡΛ…χΆΗ–‘ΓΘ“―Ψ≠―–ΨΩΝΥ Ι”Ο“ΜœΒΝ–ΙΠΡήκΡΗ≥”η PDC Υυ–ηΒΡ–ßΙϊΘ§’β–©άΐΉ”ΫΪ‘ΎΚσΟφΧ÷¬έΓΘ
2.1 Ή‘ΉιΉΑκΡ
Ή‘ΉιΉΑ «…ζΟϋΒΡ“Μ÷÷Μυ±Ψœ÷œσΘ§άΐ»γœΗΑϊΡΛΒΡ–Έ≥…[8]ΓΔΒΑΑΉ÷ ’έΒΰ[9]“‘ΦΑ DNA ΥΪ¬ί–ΐΫαΙΙ[10] «…ζΟϋΉνΜυ±ΨΒΡΉι≥…≤ΩΖ÷÷°“ΜΓΘΥυ”–…ζΈοΖ÷Ή”Θ§Αϋά®κΡΘ§ΕΦ «ΕύΙΠΡήΒΡΙΙΫ®ΩιΘ§Ω…–Έ≥…Ος»ΖΕ®“εΫαΙΙΓΘΉ‘ΉιΉΑκΡ…ηΦΤΒΡ÷ΊΒψ «ΝΥΫβΗς÷÷Ζ÷Ή”ΦδœύΜΞΉς”ΟΘ§Αϋά®«βΦϋΓΔΨ≤ΒγœύΜΞΉς”ΟΓΔΠ–-Π–Ε―ΜΐΓΔΖΕΒ¬ΜΣΝΠΚΆ…Ϊ…ΔΝΠΓΔΫπ τ-≈δΧεΗ¥ΚœΈοΚΆλΊΝΠΘ§άΐ»γ ηΥ°–‘ΚΆ≈≈≥βΧεΜΐœύΜΞΉς”Ο[11Θ§12] ¥ΥΆβΘ§Ά®ΙΐΑκκΉΑ±Υα“ΐ»κΕΰΝρΦϋ“≤±Μ”Ο”Ύ«ΐΕ·≥§Ζ÷Ή”ΉιΉΑ[13]Ηϋ÷Ί“ΣΒΡ «Θ§ΒςΫΎ’β–©œύΜΞΉς”Ο“‘ΫΪκΡΜρPDCsΉιΉΑ≥…ΨΏ”–ΡΩ±ξΫαΙΙΒΡΒίΥΆœΒΆ≥ «ΙΙΫ®Μυ”ΎΉ‘ΉιΉΑκΡΒΡΒίΥΆœΒΆ≥ΒΡΨωΕ®–‘≤Ϋ÷ηΓΘ‘Ύ±ΨΫΎ÷–Θ§Έ“Ο«±®ΗφΝΥΉ‘ΉιΉΑκΡΒΡ“ΜΦΕΚΆΕΰΦΕΫαΙΙΧΊ’ςΒΡ―–ΨΩΘ§ΉήΫαΝΥΉ‘ΉιΉΑΝΠ―–ΨΩΒΡΫχ’ΙΘ§≤ΔΫι…ήΝΥΒςΩΊPDCs–ΈΧ§ΒΡ≤Ώ¬‘ΓΘ
2.1.1 Ή‘ΉιΉΑκΡΒΡΫαΙΙΧΊ’ς
Ή‘ΉιΉΑκΡΒΡΫαΙΙΑϋά®κΡΒΡΑ±ΜυΥα–ρΝ–ΒΡ“ΜΦΕΫαΙΙΘΜœύΝΎΜρΫτΟήœύΝΎΑ±ΜυΥα÷°ΦδΒΡΖ÷Ή”ΡΎœύΜΞΉς”ΟΘ§≤ζ…ζΒΡ¬ί–ΐΓΔΠ¬-’έΒΰΓΔΖ¥ ΫΜρΈό–ρΙΙœσΒ»ΕΰΦΕΫαΙΙΘΜ“‘ΦΑΨΏ”–ΧΊΕ®’έΒΰΫαΙΙΒΡΖ÷Ή”ΦδΉιΉΑ[14]Costantini Β»»ΥΉήΫαΝΥ PDBselect ΐΨίΦ·÷–Α±ΜυΥαΤΒ¬ ΦΑΕΰΦΕΫαΙΙΉ¥Χ§«ς ΤΘ®±μ 1Θ©[15]
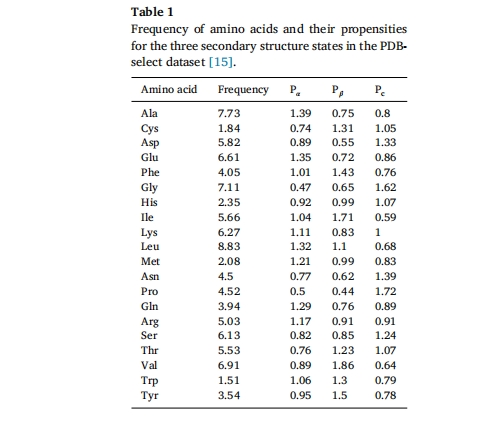
ΕύκΡΒΡΑ±ΜυΥαάύ–ΆΚΆ–ρΝ–ΧΊ’ς «ΤδΉιΉΑΡήΝΠΒΡΜυ¥ΓΘ§ΨΏ”–ΉιΉΑΡήΝΠΒΡΕύκΡΖ÷Ή”ΫαΙΙΆυΆυΨΏ”–Οςœ‘ΒΡΧΊ’ςΘ§»γΝΫ«Ή–‘ΓΔΖΦœψ–‘ΓΔΖ÷…ΔΒγΚ…ΓΔ≤ΜΆ§ΒγΚ…ΫΜΧφ≈≈Ν–ΦΑΕ‘≥Τ–‘Β»ΓΘΝΫ«Ή–‘ΕύκΡΉςΈΣ“ΜάύΧΊ βΒΡΕύκΡΘ§ΨΏ”–”κ¥ΪΆ≥±μΟφΜν–‘ΦΝœύΥΤΒΡ–‘÷ Θ§ΝΫ«Ή–‘ΕύκΡΆ§ ±Κ§”–«ΉΥ°≤ΩΖ÷ΚΆ ηΥ°≤ΩΖ÷Θ§Ε‘ΝΫœύΨυ”–«ΉΚΆΝΠΓΘΗυΨί ηΥ°Τ§ΕΈΒΡΉι≥…Ω…Ζ÷ΈΣ ηΥ°Τ§ΕΈ»Ϊ≤Ω”…Α±ΜυΥα≤–ΜυΉι≥…ΒΡ±μΟφΜν–‘κΡΘ®άύ±μΟφΜν–‘ΦΝκΡΘ©ΚΆ ηΥ°Ν¥”…≥ΛΆιΜυΧΦΝ¥Ήι≥…ΒΡΝΫ«Ή–‘ΕύκΡΓΘ±μΟφΜν–‘κΡΒΡ ηΥ°Τ§ΕΈ“ΜΑψ”…»τΗ…ΗωΝ§–χΒΡ ηΥ°Α±ΜυΥα≤–ΜυΉι≥…Θ§«ΉΥ°Τ§ΕΈΚ§”–“ΜΗωΜρΝΫΗω¥χ“ΜΗωΜρΝΫΗω’ΐΒγΚ…ΜρΗΚΒγΚ…ΒΡΑ±ΜυΥα≤–ΜυΘ§ΤδΫαΙΙΚΆ¥σ–Γ”κΧλ»ΜΝΉ÷§ °Ζ÷œύΥΤΓΘΕ‘”ΎΝΫ«Ή–‘κΡΕχ―‘Θ§ΨωΕ®Ή‘ΉιΉΑ–ΈΧ§ΒΡΙΊΦϋ‘Ύ”Ύ«ΉΥ°Τ§ΕΈΚΆ ηΥ°Τ§ΕΈΒΡΤΫΚβΘ§Ά®ΙΐΒςΫΎΑ±ΜυΥαΒΡ÷÷άύΓΔ ΐΝΩΚΆœύΕ‘ΈΜ÷Οά¥ΗΡ±δΤδΝΫ«Ή–‘÷ Θ§¥”ΕχΩΊ÷ΤΉιΉΑΧεΒΡΉν÷’ΫαΙΙΤδΫαΙΙΚΆ¥σ–Γ”κΧλ»ΜΝΉ÷§ °Ζ÷œύΥΤΓΘΕ‘”ΎΝΫ«Ή–‘κΡΕχ―‘Θ§ΨωΕ®Ή‘ΉιΉΑ–ΈΧ§ΒΡΙΊΦϋ‘Ύ”Ύ«ΉΥ°Τ§ΕΈΚΆ ηΥ°Τ§ΕΈΒΡΤΫΚβΓΘΆ®ΙΐΒςΫΎΑ±ΜυΥαΒΡ÷÷άύΓΔ ΐΝΩΚΆœύΕ‘ΈΜ÷Οά¥ΗΡ±δΤδΝΫ«Ή–‘Θ§Ω…“‘ΩΊ÷ΤΉιΉΑΧεΒΡΉν÷’ΫαΙΙΤδΫαΙΙΚΆ¥σ–Γ”κΧλ»ΜΝΉ÷§ °Ζ÷œύΥΤΓΘΕ‘”ΎΝΫ«Ή–‘κΡΕχ―‘Θ§ΨωΕ®Ή‘ΉιΉΑ–ΈΧ§ΒΡΙΊΦϋ‘Ύ”Ύ«ΉΥ°Τ§ΕΈΚΆ ηΥ°Τ§ΕΈΒΡΤΫΚβΓΘΆ®ΙΐΒςΫΎΑ±ΜυΥαΒΡ÷÷άύΓΔ ΐΝΩΚΆœύΕ‘ΈΜ÷Οά¥ΗΡ±δΤδΝΫ«Ή–‘Θ§Ω…“‘ΩΊ÷ΤΉιΉΑΧεΒΡΉν÷’ΫαΙΙ[16®C18]ΖΦœψκΡ÷ς“ΣΑϋά®“ΜάύΚ§”–ΖΦœψΉεΑ±ΜυΥαΒΡΕύκΡΘ§»γPheΓΔTyrΓΔTrpΒ»Θ§ΥϋΟ«ΒΡΖΦœψ–‘≤ΜΫω«ΐΕ·ΕύκΡΉ‘ΉιΉΑΚΆΡΐΫΚΜ·Θ§Εχ«“Ω…“‘ΒςΫΎΧεœΒΒΡ ηΥ°–‘Θ§ΖΦœψ–‘ΉςΈΣ“Μ÷÷«ΩœύΜΞΉς”ΟΘ§Ω…“‘ΧαΗΏΕύκΡΖ÷Ή”ΒΡΉ‘ΉιΉΑΡήΝΠ[19]÷ΛΟςΝΥΕ‘≥Τ–‘“≤ «Ή‘ΉιΉΑΖ÷Ή”ΒΡ“ΜΗωΙΊΦϋ“ρΥΊΘ§“ρΈΣΕ‘≥Τ–‘Φθ…ΌΝΥΉ‘ΉιΉΑΖ÷Ή”Τ¥Ϋ” ±ΒΡΫγΟφάύ–Ά[20Θ§21]ΓΘ
2.1.2 Ή‘ΉιΉΑΝΠ
ΕύκΡΩ…“‘‘ΎΕύ÷÷Ζ«Ι≤ΦέΦϋΝΠΒΡΙ≤Ά§«ΐΕ·œ¬Θ§»γ ηΥ°Ής”ΟΓΔΠ–-Π–Ε―ΜΐΓΔΨ≤ΒγΝΠΓΔ«βΦϋΒ»Ή‘ΉιΉΑ–Έ≥…≤ΜΆ§–ΈΟ≤ΚΆΙΠΡήΒΡΉιΉΑΧεΓΘΥδ»Μ’β–©Ζ«Ι≤ΦέΦϋΒΡΉς”ΟΝΠœύΕ‘Ϋœ»θΘ§ΒΪΥϋΟ«Ι≤Ά§Ής”Ο ±Θ§ΤδΉς”Ο«ΩΕ»≤Μ»ίΚω ”ΓΘ…ν»κΧΫΨΩΕύκΡΒΡΉ‘ΉιΉΑ––ΈΣΦΑΉιΉΑΧε–Έ≥…ΒΡΜζ÷ΤΘ§ Ή“Σ»ΈΈώ «ΝΥΫβΗς÷÷Ζ«Ι≤ΦέΦϋΝΠ‘ΎΉιΉΑΙΐ≥Χ÷–ΒΡΉς”ΟΘ§ΈΣΕ®œρΒςΩΊΉιΉΑΧεΒΡ–ΈΟ≤ΚΆ≥Ώ¥γΧαΙ©“άΨίΓΘ
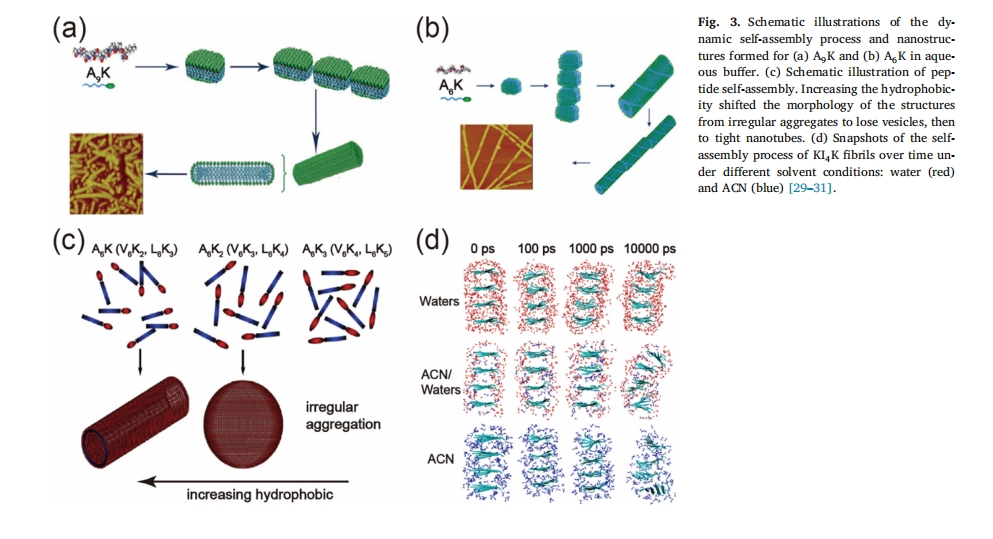
ηΥ°œύΜΞΉς”Ο «κΡΉ‘ΉιΉΑΒΡ÷Ί“Σ«ΐΕ·ΝΠ[16], ηΥ°ΝΠΒΡ±Ψ÷ «λΊ–ß”Π[22]Θ§ ηΥ°œύΜΞΉς”ΟΒΡΡήΝΩœύΕ‘Ϋœ–ΓΘ§ΤδΡήΝΩ‘ΦΈΣ4®C8 kJ/mol[23],ΕύκΡ÷– ηΥ°≤–ΜυΘ®±ϊΑ±ΥαΓΔγ”Α±ΥαΓΔΝΝΑ±ΥαΓΔ“λΝΝΑ±ΥαΓΔά“Α±ΥαΓΔ±Ϋ±ϊΑ±ΥαΚΆ…ΪΑ±ΥαΘ©ΒΡ±»άΐΚΆ ΐΝΩΩ…œ‘÷χ”ΑœλΕύκΡΉιΉΑΧεΒΡΡ…ΟΉΫαΙΙΓΘ“ΜΑψά¥ΥΒΘ§ΦΪ–‘Α±ΜυΥαΚΆΖ«ΦΪ–‘Α±ΜυΥαΉι≥…ΒΡΕύκΡ–ρΝ–Ηϋ»ί“Ή–Έ≥…«ρ–ΈΫαΙΙΓΘΖ«ΦΪ–‘Α±ΜυΥα‘Ύ ηΥ°ΝΠΒΡΉς”Οœ¬ΨέΦ·–Έ≥… ηΥ°ΚΥ–ΡΘ§«ΉΥ°ΦΪ–‘Α±ΜυΥα‘ρ≈≈Ν–‘ΎΆβ≤ύ”κΥ°Ϋ”¥ΞΓΘ‘ΎΗΏ≥ΛΩμ±»ΫαΙΙΘ®œΥΈ§ΓΔ±ΓΡΛΘ©ΒΡ–Έ≥…Ιΐ≥Χ÷–Θ§ ηΥ°ΝΠ «¥ΌΫχΠ¬-’έΒΰΚαœρΕ―ΜΐΒΡ«ΑΧαΘ§ΕχΠ¬-’έΒΰΚαœρΕ―ΜΐΕ‘”ΎΉιΉΑΧεΩμΕ»ΒΡΫχ“Μ≤ΫΜΐάέ÷ΝΙΊ÷Ί“Σ[24Θ§25],GrootΒ»±»ΫœΝΥIle-Phe-ΚΆVal-PheΝΫ÷÷Ή‘ΉιΉΑΕΧκΡΒΡΉ‘ΉιΉΑ––ΈΣΘ§ΖΔœ÷«Α’ΏΒΡΉ‘ΉιΉΑΡήΝΠ»θ”ΎΚσ’ΏΘ§≤Δ÷Λ Β’β «”…”ΎIle-≤–Μυ…ΌΝΥ“ΜΗωCH3±»Val≤–ΜυΗΏΘ§ ηΥ°Ής”ΟΫΒΒΆΘ§Ήν÷’ΒΦ÷¬κΡΉ‘ΉιΉΑΡήΝΠœ¬ΫΒ[26],‘ΎΉ‘ΉιΉΑκΡΖ÷Ή”ΒΡCΕΥΚΆNΕΥ“ΐ»κ÷§ΖΨΝ¥Ω…“‘”––ßΧαΗΏΉ‘ΉιΉΑΫαΙΙΒΡΈ»Ε®–‘[27]LöwikΒ»»ΥΖΔœ÷Θ§“ΐ»κΒΡ÷§ΖΨΝ¥ΒΡ≥ΛΕ»Μα”Αœλ ηΥ°œύΜΞΉς”ΟΒΡ«ΩΕ»Θ§¥”ΕχΈ»Ε®ΚΆ‘ω«ΩΉ‘ΉιΉΑΒΡΡήΝΠ≤ΜΆ§[28]”–―–ΨΩ÷ΛΟςΘ§Ά®Ιΐ‘ωΦ”κΡΒΡ ηΥ°–‘ΜρΦθ…Ό«ΉΥ°–‘Α±ΜυΥαΒΡ ΐΝΩΘ§Ή‘ΉιΉΑΚσ–Έ≥…ΒΡΡ…ΟΉΫαΙΙΩ…“‘¥”Ρ…ΟΉΡ“≈ίΉΣ±δΈΣΡ…ΟΉΙήΚΆΡ…ΟΉ¥χΓΘWangΒ»[29]. Ι”ΟΝΫ«ΉκΡ A6KΚΆA9K ΧΫΧ÷ΝΥ ηΥ°œύΜΞΉς”ΟΕ‘κΡΖ÷Ή”Ή‘ΉιΉΑΫαΙΙΒΡ”ΑœλΓΘΥϊΟ«Ιέ≤λΒΫ A9K Ω…“‘Ή‘ΉιΉΑ≥…ΒΆΉίΚα±»ΫαΙΙΘ§άΐ»γΡ…ΟΉ«ρΜρΕΧΡ…ΟΉΑτΘ®ΆΦ 3a).»ΜΕχΘ§ΥφΉ≈ ηΥ°±»ΒΡΫΒΒΆΘ§Ω…“‘Ά®ΙΐA–Έ≥…ΗΏΉίΚα±»ΒΡœΥΈ§ΫαΙΙ6ΦΊΘ®ΆΦ 3b) ±μΟςΘ§Β± ηΥ°±»‘ωΦ” ±Θ§Ή‘ΉιΉΑκΡ«ψœρ”ΎΨέΦ·–Έ≥…«ρ–ΈΫαΙΙΓΘœύ±»÷°œ¬Θ§Meng Β»»Υ[30]. ΖΔœ÷ ηΥ°œύΜΞΉς”ΟΒΡ‘ωΦ”ΜαΗΡ±δΉιΉΑΫαΙΙΒΡ–ΈΧ§Θ§¥”≤ΜΙφ‘ρΒΡΨέΦ·Χε±δΈΣ«ρ–ΈΡ“≈ίΘ§ΉνΚσ±δΈΣΡ…ΟΉΙήΫαΙΙΘ®ΆΦ 3c), ’β“ΜΩ¥ΥΤΟ§ΕήΒΡΫα¬έΩ…Ρή «”…”ΎΆ§÷÷ΒγΚ…Φδ¥φ‘ΎΫœ«ΩΒΡ≈≈≥βΝΠΘ§ΕχΉςΈΣ«ΉΥ°ΉιΖ÷ΒΡKΘ§Τδ ΐΝΩ‘ΫΕύΘ§≤ζ…ζΒΡ≈≈≥βΉηΝΠ‘Ϋ¥σΘ§“ρ¥ΥΘ§κΡΒΡΉ‘ΉιΉΑ–ΈΟ≤ «”… ηΥ°œύΜΞΉς”ΟΚΆΨ≤Βγ≈≈≥β÷°ΦδΒΡΤΫΚβΨωΕ®ΒΡΓΘZhao et al[31]. Ιέ≤λΒΫ““κφ (ACN) ΤΤΜΒ ηΥ°œύΜΞΉς”ΟΘ§≤Δ±®ΒάΝΥΤδΕ‘Ή‘ΉιΉΑΒΡ”ΑœλΓΘ»γΆΦΥυ ΨΆΦ 3d, KI ΒΡ‘≠œΥΈ§4K”…ΥΡΗωΠ¬-’έΒΰΉι≥…Θ§Τδ÷–I4Π¬ ’έΒΰΤ§Τπ ηΥ°ΚΥ–ΡΒΡΉς”ΟΓΘ‘ΎΥ°÷–Θ§Ιέ≤λΒΫκΡΖ÷Ή”ΫτΟήΕ―Μΐ–Έ≥…‘≠œΥΈ§Θ§œΥΈ§ΡΎ≤Ω≥δΒ± ηΥ°ΚΥ–ΡΘ§Υ°Ζ÷Ή”«ψœρ”Ύ¥”ΝΫΗω Π¬ ’έΒΰΤ§÷°Φδ“ί≥ωΓΘ»ΜΕχΘ§‘Ύ““κφ/Υ°ΜλΚœΈοΘ®1:1 ΡΠΕϊ±»Θ©ΚΆ““κφ÷–Θ§κΡΖ÷Ή”Ε―Μΐ≤ΜΧΪΫτΟήΘ§»ήΦΝΖ÷Ή”“≤Ζ÷≤Φ‘Ύ ηΥ°ΚΥ–Ρ÷–ΓΘΖ÷Ή”Ε·ΝΠ―ßΡΘΡβΫχ“Μ≤Ϋ±μΟςΘ§““κφΖ÷Ή”Ω…“‘Ϋχ»κκΡΒΡ ηΥ°≤ΩΖ÷≤Δœς»θ ηΥ°œύΜΞΉς”ΟΘ§¥”ΕχΤΤΜΒœύΝΎ’έΒΰΤ§÷°ΦδΒΡΝ§–χ–‘[32]’β–©ΕάΧΊΒΡΫαΙΙ≤ΜΫωΩ…“‘ΉςΈΣ÷ßΦήΙΙΫ®Ης÷÷Ρ…ΟΉΤςΦΰΘ§ΜΙΩ…“‘”Ο”ΎΉΑ‘Ί“©ΈοΘ§ Βœ÷“©ΈοΒΡΩΊ÷Τ ΆΖ≈ΓΘ33Θ§34]ΓΘ
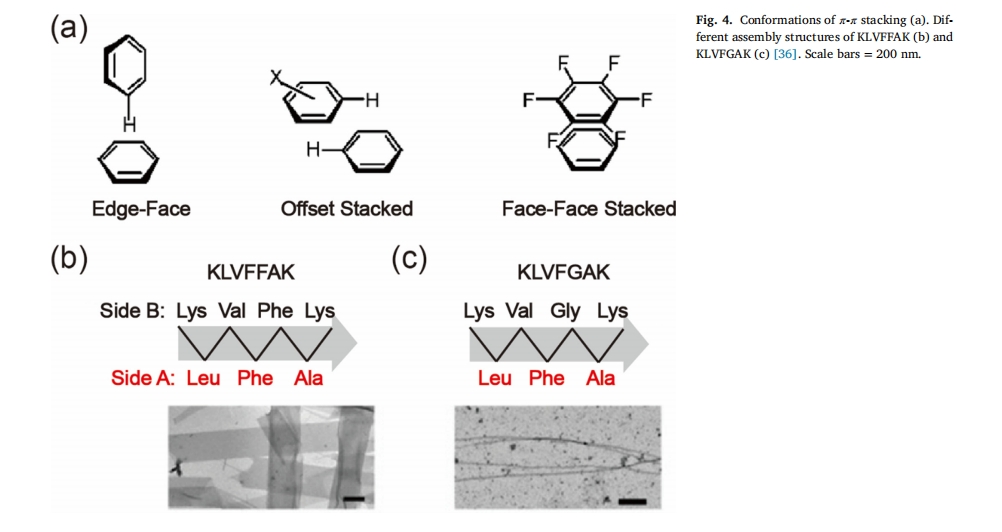
Π–-Π–Ε―ΜΐœύΜΞΉς”Ο”÷≥ΤΖΦœψœύΜΞΉς”ΟΘ§÷ς“ΣΖΔ…ζ‘ΎΠ–-Π–Ι≤ινΧεœΒ÷–Θ§ΤδΝΠΡήΈΣ8®C12 kJ/mol[35],”κ“ΜΑψΒΡ ηΥ°œύΜΞΉς”Ο≤ΜΆ§Θ§Π–-Π–ΜΐάέœύΜΞΉς”ΟΨΏ”–ΖΫœρ–‘Θ§…φΦΑΨΏ”–ΧΊΕ®Ω’Φδ≈≈Ν–ΒΡ ηΥ°œύΜΞΉς”ΟΓΘΆ®≥Θ”–»ΐ÷÷Μΐάέ–Έ ΫΘ§Αϋά®±ΏΕ‘ΟφœύΜΞΉς”ΟΓΔΤΪ“ΤΕ―ΜΐΙΙœσΚΆΟφΕ‘ΟφΕ―ΜΐΦΗΚΈΫαΙΙΘ®ΆΦ 4aΘ©[36],Π–-Π–Ε―Μΐ÷ς“Σ «”…ΖΦœψΉεΑ±ΜυΥα≤–ΜυΘ®»γ±Ϋ±ϊΑ±Υα[37Θ§38], …ΪΑ±Υα[39]ΚΆά“Α±Υα[40]) ‘ΎκΡ–ρΝ–÷–ΓΘΤδΥϊΖΦœψΜυΆ≈ΒΡ“ΐ»κ[41]Θ§άΐ»γ Fmoc ΜυΆ≈[42]ΚΆΕΰΟ·Χζ[43]Θ§“≤Ρή≤ζ…ζΠ–-Π–Ε―ΜΐΓΘ‘ΎΚ§”–ΖΦœψΜΖΒΡκΡΫαΙΙ÷–Θ§’β÷÷ΜΖΒΡ”––ρΕ―Μΐ“ΐΒΦκΡΉιΉΑΧεΒΡΕ®œρ…ζ≥ΛΘ§Ήν÷’–Έ≥…ΗϋΦ”ΫτΟή”––ρΒΡΡ…ΟΉΫαΙΙ[44Θ§45],Π–-Π–ΒΡΜΐάέ≤ΜΫωΧαΗΏΝΥκΡΖ÷Ή”ΒΡΉ‘ΉιΉΑΡήΝΠ[46Θ§47] Εχ«“‘Ύ“Μ–©≤Γάμ–‘ΒμΖέ―υΒΑΑΉΨέΦ·Ιΐ≥Χ÷–ΤπΉ≈ΨωΕ®–‘Ής”Ο[48Θ§49],±»»γΘ§ΒμΖέ―υΑΏΩιΒΡΕ―Μΐ «ΝΌ¥≤≈–ΕœΑΔΕϊ¥ΡΚΘΡ§≤ΓΘ®ADΘ©ΒΡΫπ±ξΉΦ[50],¥Ό ΙΒμΖέ―υΒΑΑΉΉ‘ΨέΦ·ΒΡ÷ς“ΣΝΠΝΩ «ΒΑΑΉ÷ ÷–ΖΦœψΉεΑ±ΜυΥα≤–Μυ÷°ΦδΠ–-Π–ΦϋΒΡΜΐάέΘ§“ρ¥ΥΘ§ΤΤΜΒΒμΖέ―υΒΑΑΉΖ÷Ή”ΦδΒΡΠ–-Π–œύΜΞΉς”Ο≥Θ±Μ”Ο”Ύ“÷÷ΤΒμΖέ―υΑΏΩιΒΡ–Έ≥…Θ§“‘ΧΫΥςADΒΡ÷ΈΝΤ≤Ώ¬‘ΓΘKLVFFAK «KLVFFAE AΠ¬Θ®16®C22Θ©ΒΡ“ΜΗωΆΜ±δΧεΘ§Β±KLVFFAKκΡΕΈ÷–ΒΡΒΎΕΰΗω±Ϋ±ϊΑ±Υα±ΜΗ Α±Υα»Γ¥ζΘ®KLVFGAKΘ©ΚσΘ§Π–-Π–Ε―ΜΐΉς”Ο±»KLVFFAK»θΒΟΕύΘ§“ρ¥ΥΚαœρ…ζ≥ΛΒΡ«ΐΕ·ΝΠΟςœ‘Φθ»θΘ§Ή‘ΉιΉΑ”…ΫœΩμΒΡ¥χΉ¥ΫαΙΙ±δΈΣΫœ’≠ΒΡΡ…ΟΉœΥΈ§ΫαΙΙΘ®ΆΦ 4b, c)[51],¥ΥΆβΘ§Π–-Π–Ε―ΜΐΩ…“‘ΫΒΒΆΉ‘ΉιΉΑκΡΒΡΝΌΫγΨέΦ·≈®Ε»Θ§ΧαΗΏΉιΉΑΫαΙΙΒΡΈ»Ε®–‘ΚΆΤδΥϊΝΠ―ß–‘Ρή[52Θ§53]ΓΘ
ΈόΖΫœρ–‘ΒΡΨ≤ΒγΝΠΑϋά®Ά§ΒγΚ…Έϋ“ΐΚΆ“λΒγΚ…≈≈≥βΓΘΨ≤ΒγΝΠά¥‘¥”ΎκΡ–ρΝ–÷–ΒΡΥα–‘ΜρΦν–‘Α±ΜυΥαΓΘΨ≤ΒγΉς”ΟΥδ»Μ±»Ϋœ»θΘ§ΡήΝΩΈΣ0.5ΓΪ4kJ/molΘ§ΒΪΕ‘κΡΉ‘ΉιΉΑΒΡ”ΑœλΫœ¥σΓΘ‘Ύ“‘Ψ≤ΒγΝΠΈΣ÷ςΒΦΒΡκΡΫαΙΙ÷–Θ§Ά§ΒγΚ…≤ζ…ζΒΡΨ≤Βγ≈≈≥βΝΠΆυΆυ±μœ÷ΈΣκΡΉ‘ΉιΉΑΒΡΉηΝΠΜρΈ»Ε®ΝΠΓΘ¥χΒγΤ§ΕΈ“ΜΑψΈΣ«ΉΥ°Ν¥Θ§”ωΥ°≈ρ’ΆΘ§ ηΥ°Τ§ΕΈ”ωΥ°Χ°Υζ ’ΥθΨέΦ·–Έ≥…ΡΎΚΥΓΘΥυ–Έ≥…ΒΡΫΚ χΒΡ≥Ώ¥γ»ΓΨω”Ύ ηΥ°ΚΥΒΡ±μΟφ’≈ΝΠ”κ«ΉΥ°Ω«÷–œύΝΎΤ§ΕΈΒΡΨ≤Βγ≈≈≥β÷°ΦδΒΡΤΫΚβΓΘ”κΆ§÷÷ΒγΚ…≈≈≥βœύΖ¥Θ§’ΐΒγΚ…ΚΆΗΚΒγΚ…÷°ΦδΒΡΨ≤Βγ“ΐΝΠΩ…“‘ΩΊ÷ΤκΡΖ÷Ή”ΒΡΨέΦ·––ΈΣΘ§≤Δ ΙΥϋΟ«“‘’ΐΒγΚ…ΚΆΗΚΒγΚ…≤–Μυ±Υ¥ΥΫ”ΫϋΒΡΖΫ ΫΉ‘ΉιΉΑΓΘRamachandran ΖΔœ÷Θ§‘ΎΨ≤Βγ“ΐΝΠΒΡΉς”Οœ¬Θ§¥χœύΖ¥ΒγΚ…ΒΡκΡΩ…“‘Ή‘ΖΔΨέΦ·≤ΔΉιΉΑ‘Ύ“ΜΤπΘ§–Έ≥…ΡήΙΜΗΚ‘Ί“©ΈοΚΆΙΠΡήΒΑΑΉΒΡκΡ‘ΊΧε’ΐΗΚΒγΚ…ΦδΒΡΨ≤Βγ“ΐΝΠΩ…“‘ΩΊ÷ΤκΡΖ÷Ή”ΒΡΨέΦ·––ΈΣΘ§ ΙΤδΉ‘ΉιΉΑΘ§ Ι¥χ’ΐΒγΚΆ¥χΗΚΒγΒΡ≤–ΜυœύΜΞΩΩΫϋΓΘRamachandranΖΔœ÷Θ§‘ΎΨ≤Βγ“ΐΝΠΒΡΉς”Οœ¬Θ§¥χœύΖ¥ΒγΚ…ΒΡκΡΩ…“‘Ή‘ΖΔΨέΦ·ΉιΉΑ‘Ύ“ΜΤπΘ§–Έ≥…ΡήΙΜΗΚ‘Ί“©ΈοΚΆΙΠΡήΒΑΑΉΒΡκΡ‘ΊΧε’ΐΗΚΒγΚ…ΦδΒΡΨ≤Βγ“ΐΝΠΩ…“‘ΩΊ÷ΤκΡΖ÷Ή”ΒΡΨέΦ·––ΈΣΘ§ ΙΤδΉ‘ΉιΉΑΘ§ Ι¥χ’ΐΒγΚΆ¥χΗΚΒγΒΡ≤–ΜυœύΜΞΩΩΫϋΓΘRamachandranΖΔœ÷Θ§‘ΎΨ≤Βγ“ΐΝΠΒΡΉς”Οœ¬Θ§¥χœύΖ¥ΒγΚ…ΒΡκΡΩ…“‘Ή‘ΖΔΨέΦ·ΉιΉΑ‘Ύ“ΜΤπΘ§–Έ≥…ΡήΙΜΗΚ‘Ί“©ΈοΚΆΙΠΡήΒΑΑΉΒΡκΡ‘ΊΧε[54]Β±¥χœύΖ¥ΒγΚ…ΒΡΑ±ΜυΥα≤–ΜυΖ÷≤Φ‘ΎκΡ–ρΝ–ΒΡΝΫΕΥ ±Θ§κΡ‘ΎΉ‘ΉιΉΑΙΐ≥Χ÷–«ψœρ”Ύ≤…”ΟΖ¥œρΤΫ––ΒΡΖ÷Ή”≈≈Ν–[55Θ§56],StuppΒ»―–ΨΩ÷Λ ΒΘ§¥χœύΖ¥ΒγΚ…ΒΡΑ±ΜυΥα≤–ΜυΦδΨ≤Βγ“ΐΝΠΉς”Οœ¬–Έ≥…ΒΡΡ…ΟΉœΥΈ§ΨΏ”–Ηϋ«ΩΒΡ»»Έ»Ε®–‘[57]. άϊ”ΟΟ…ΧΊΩ®¬όΡΘΡβ―–ΨΩΝΥκΡΉ‘ΉιΉΑΙΐ≥Χ÷–Ψ≤ΒγœύΜΞΉς”ΟΕ‘Έ§≥÷ΉιΉΑ–ΈΟ≤ΒΡ÷Ί“Σ–‘ΓΘΫαΙϊ±μΟςΘ§Ρ…ΟΉœΥΈ§«ΉΥ°±μΟφ¥χΒγΑ±ΜυΥα≤–Μυ÷°ΦδΒΡΨ≤ΒγœύΜΞΉς”ΟΩ…“‘Έ»Ε®œΥΈ§ΫαΙΙ[58]Ά®ΙΐΧΫΨΩΖ÷Ή”ΨΜΒγΚ…Ε‘Ή‘ΉιΉΑΒΡ”ΑœλΘ§ΖΔœ÷’ϊΧεΨΜΒγΚ…ΈΣ1Ήν”–άϊ”ΎΉ‘ΉιΉΑœΥΈ§ΒΡ–Έ≥…[59]ΓΘ
Ή‘ΉιΉΑΧεœΒΒΡpH÷Β «”ΑœλΨ≤Βγ–ß”ΠΒΡ÷Ί“Σ“ρΥΊΘ§ΒςΫΎpH÷ΒΜαΒΦ÷¬ΕύκΡΒγΚ…ΒΡ±δΜ·Θ§ΫχΕχΗΡ±δΕύκΡΒΡ≈≈Ν–ΖΫ ΫΘ§“ΐΖΔΉ‘ΉιΉΑ¥”œΥΈ§ΒΫΡ…ΟΉΙήΒΡΉΣ±δΓΘΜυ”Ύ¥ΥΙφ¬…Θ§…ηΦΤ≥ωΉνΕΧΉ‘ΉιΉΑάκΉ”ΜΞ≤ΙκΡΖ÷Ή”KFFE[60]. Honggang Cui Β»»Υ÷ΛΟςΝΥκΡ EF4E ΚΆ KF4¥χ“Μ÷÷ΒγΚ…ΒΡ K ÷ΜΡή‘ΎΆ§÷÷ΒγΚ…≈≈≥βΧθΦΰœ¬–Έ≥…’≠ΒΡ≈Λ«ζ¥χΚΆœΗœΥΈ§ΓΘ”–»ΛΒΡ «Θ§Β±¥χœύΖ¥ΒγΚ…ΒΡ E ΚΆ K ≤–Μυ±Μ“ΐ»κΒΫΆ§“ΜΗω EF ÷– ±4KκΡΘ§’ΐΗΚΒγΚ…œύΜΞ÷–ΚΆΘ§ΤΝ±ΈΝΥΖ÷Ή”ΦδΒΡΨ≤Βγ≈≈≥βΘ§ΉιΉΑΧεΚαœρ…ζ≥ΛΘ§Ήν÷’–Έ≥…“ΜΧθΩμ¥χΓΘ±ΨΈΡ«ΑΟφΧαΒΫΒΡ≤ΜΆ§Ζ÷Ή”œύ”ΠΉιΉΑΧεΒΡ–Έ≥…Μζάμ»γΆΦΥυ ΨΆΦ 5a[61]ΓΘ
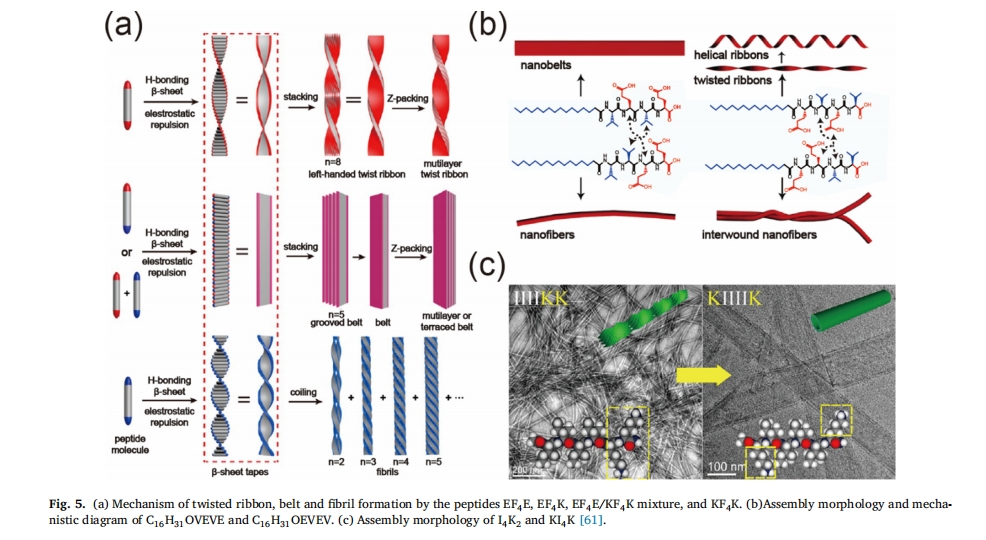
κΡΒΡΑ±ΜυΥα–ρΝ–”κΤδΉ‘ΉιΉΑΫαΙΙΟή«–œύΙΊ[32Θ§62],Ά®ΙΐΗΡ±δκΡΑ±ΜυΥα≤–ΜυΒΡ–ρΝ–Θ§Ω…“‘ΒΟΒΫΆξ»Ϊ≤ΜΆ§ΒΡ“ΜΈ§Ρ…ΟΉΫαΙΙ[62]. »γΆΦΥυ ΨΆΦ 5bΘ§κΡC16H31 OVEVEΚΆΨΏ”–ΫΜΧφ«ΉΥ°ΚΆ ηΥ°Α±ΜυΥα≤–ΜυΒΡC16H31 OEVEVΩ…“‘–Έ≥…ΗϋΩμΒΡ¥χΓΘœύ±»÷°œ¬Θ§κΡC16H31 OVVEE ΚΆ≤ΜΫΜΧφ≈≈Ν–ΒΡC16H31 OEEVV÷ΜΡή–Έ≥…ΫœœΗΒΡœΥΈ§ΓΘ’β «“ρΈΣVEVEΚΆEVEVΒΡΫΜΧφ≈≈Ν–Ω…“‘ΚήΚΟΒΊΖ÷…ΔΒγΚ…Θ§ΫΒΒΆEE÷°ΦδΒΡ≈≈≥βΝΠΓΘ‘ΎEEVVΚΆVVEEΒΡΝ§–χ≈≈Ν–÷–Θ§ΝΫΗω¥χΒγΑ±ΜυΥαEE±Υ¥ΥΩΩΫϋΘ§Ψ≤Βγ≈≈≥βΝΠΫœ«ΩΓΘΆ§―υΘ§―–ΨΩΜΙΖΔœ÷Θ§ΨΓΙήI4K2ΚΆ KI4K ΨΏ”–œύΆ§ ΐΝΩΚΆ÷÷άύΒΡΑ±ΜυΥα, ΤδΉιΉΑ–ΈΧ§¥φ‘ΎΟςœ‘≤ν“λ (ΆΦ 5c). Ε‘”ΎI4K2Θ§Ζ÷Ή”“ΜΕΥ¥χ’ΐΒγΒΡK≤–ΜυΒΦ÷¬ΧεœΒ≤ζ…ζ«ΩΝ“ΒΡΨ≤Βγ≈≈≥βΘ§¥”Εχœς»θΠ¬-Τ§≤ψ÷°ΦδΒΡ ηΥ°Έϋ“ΐΝΠΘ§¥”Εχ–Έ≥…œΗΡ…ΟΉœΥΈ§ΓΘœύΖ¥Θ§KI¥χΒγ≤–Μυ4KΈΜ”ΎΖ÷Ή”ΝΫΕΥΘ§Ε‘≥ΤΒΡΫαΙΙΒΦ÷¬ΝΫΕΥΒΡK“ΐΤπΒΡΨ≤Βγ≥βΝΠ¥σ¥σΦθ»θΘ§Εχ«“ ηΥ°œύΜΞΉς”Ο ήΨ≤Βγ–ß”ΠΒΡ”ΑœλΫœ–ΓΓΘ
«βΦϋ «“Μ÷÷ΧΊ βΒΡΖΕΒ¬ΜΣΝΠΘ§ΨΏ”–ΖΫœρ–‘ΚΆ±ΞΚΆ–‘ΓΘΆ®≥Θ«ιΩωœ¬Θ§«βΦϋ«ΩΕ»¥σ”ΎΖΕΒ¬ΜΣΝΠΘ§–Γ”ΎΙ≤ΦέΦϋΓΘκΡ÷ςΝ¥÷°ΦδΒΡ«βΦϋΡή‘ΦΈΣ4®C12 kJ/mol[23]”…”ΎκΡΫαΙΙ÷–Κ§”–ΫœΕύΒΡC=OΚΆNHΜυΆ≈Θ§Ω…“‘–Έ≥…ΫœΕύΒΡ«βΦϋΘ®N-HΓ≠OΘ©ΓΘ«βΦϋ «”ΑœλκΡΕΰΦΕΫαΙΙΒΡ÷Ί“Σ“ρΥΊΘ§Ζ÷ΈΣΖ÷Ή”ΡΎ«βΦϋΚΆΖ÷Ή”Φδ«βΦϋΓΘ”κ¥ΪΆ≥±μΟφΜν–‘ΦΝΒΡΫαΙΙœύ±»Θ§ΝΫ«Ή–‘κΡΉν¥σΒΡ«χ±π‘Ύ”ΎΖ÷Ή”Ι«Φή÷–¥φ‘Ύ¥σΝΩ«βΦϋΘ§¥Ό ΙΉιΉΑΧε―Ί≥Λ÷αΖΫœρ…ζ≥ΛΘ§’β“≤ «ΈΣ ≤Ο¥±μΟφΜν–‘ΦΝ“ΜΑψ«ψœρ”Ύ–Έ≥…«ρ–ΈΜρΑτΉ¥ΫαΙΙΘ§ΕχκΡ«ψœρ”Ύ–Έ≥…œΥΈ§Ή¥ΫαΙΙΒΡ‘≠“ρΓΘZhu et al[63]÷ΛΟς«βΦϋΡή ΙΨΏ”–ΫΜΧφ«ΉΥ°–‘ΚΆ ηΥ°–‘Α±ΜυΥα≤–ΜυΒΡκΡ–Έ≥…Π¬-’έΒΰΫαΙΙΘ§ΗΟΫαΙΙ¥Ι÷±”ΎœΥΈ§ΒΡ≥Λ÷α≈≈Ν–Θ§¥”Εχ±ήΟβœΥΈ§ΒΡ≈Λ«ζΜρΆδ«ζΓΘ“ρ¥ΥΘ§”…«βΦϋ«ΐΕ·ΒΡΠ¬-’έΒΰΒΡ–Έ≥…ΒΦ÷¬κΡΖ÷Ή”Ή‘ΉιΉΑΚΆΨέΦ·Θ§–Έ≥…ΨΏ”–ΗΏ≥ΛΨΕ±»ΒΡ‘≤÷υ–ΈΡ…ΟΉœΥΈ§ΓΔΡ…ΟΉ¥χΚΆΡ…ΟΉΙή[63®C67],Pahrump Β»»ΥΖΔœ÷Ρ…ΟΉœΥΈ§ΚΥ–Ρ÷–ΥΡΗωΑ±ΜυΥα≤–Μυ÷°ΦδΒΡΖ÷Ή”Φδ«βΦϋΩ…“‘ ΙκΡ«ψœρ”Ύ–Έ≥…¥Ι÷±”ΎΡ…ΟΉœΥΈ§≥Λ÷αΒΡ Π¬ Τ§≤ψΫαΙΙΓΘ”…¥ΥΫ“ ΨΝΥΗΜΚ§«βΦϋΒΡΖ÷Ή”«ψœρ”Ύ–Έ≥…ΗΏΉίΚα±»ΒΡœΥΈ§ΫαΙΙ[68],œύ±»÷°œ¬Θ§‘ΕάκΚΥ–ΡΒΡΑ±ΜυΥα≤–Μυ‘ΎΉ‘ΉιΉΑΚΆΈ§≥÷œΥΈ§Έ»Ε®–‘ΚΆΫαΙΙΗ’–‘ΖΫΟφΒΡΉς”ΟΫœ»θΓΘ«βΦϋΒΡΤΤΜΒΜρΗΡ±δΒΦ÷¬‘≤÷υ–ΈΡ…ΟΉœΥΈ§ΒΡΫαΙΙΖΔ…ζ±δΜ·Θ§άΐ»γ≥ΛœΥΈ§ΒΡΦΗΚΈ–ΈΉ¥ΖΔ…ζΗΡ±δΘ§ΒΦ÷¬«ρ–ΈΫΚ χΘ§…θ÷ΝΉ‘ΉιΉΑκΡ–ΈΧ§œϊ ß[69]ΓΘ
κΡΒΡΉ‘ΉιΉΑ «”…Εύ÷÷Ζ«Ι≤ΦέΦϋΝΠ«ΐΕ·ΒΡΘ§’β–©ΝΠΨωΕ®ΝΥκΡΖ÷Ή”œρ≤ΜΆ§ΖΫœρ…ζ≥ΛΘ§¥”Εχ–Έ≥…≤ΜΆ§–ΈΟ≤ΒΡΉιΉΑΧεΓΘ÷Μ”–‘Ύ≥δΖ÷άμΫβ…œ ωΖ«Ι≤ΦέΦϋΝΠΒΡΜυ¥Γ…œΘ§≤≈ΡήΆ®ΙΐΒςΫΎΗς÷÷Ζ«Ι≤ΦέΦϋΝΠΒΡœύΕ‘¥σ–Γά¥”––ßΩΊ÷ΤΉιΉΑΧεΒΡ–ΈΟ≤ΚΆ≥Ώ¥γΓΘ
2.2 ≤…”ΟPDCsΒΡΩ…ΉΣΜ·Ρ…ΟΉ“©Έο
ΫΪ“©Έο”κΉ‘ΉιΉΑκΡΫαΚœΩ…–Έ≥…Ης÷÷Ρ…ΟΉΒίΥΆœΒΆ≥Θ§Ω… ”ΠΑ–œρ–‘ΓΔ≥ΛΤΎ÷ΆΝτ–‘ΚΆΧαΗΏ“©ΈοΈ»Ε®–‘ΓΘ¥ΥΆβΘ§ΝΘΨΕΚΆ–ΈΧ§Ε‘Ρ…ΟΉΒίΥΆœΒΆ≥ΒΡ«ε≥ΐ¬ ΓΔ¥©ΆΗ–߬ ΓΔΑ–œρ÷ΆΝτΚΆœΗΑϊΤςΖ÷≤Φ”–œύΒ±¥σΒΡ”Αœλ[70]άΐ»γΘ§‘Φ100Ρ…ΟΉΒΡΡ…ΟΉΝΘΉ””–άϊ”Ύ‘ω«Ω…χΆΗ–‘ΚΆ±ΘΝτ–ß”ΠΘ®EPRΘ©ΒΡ―ΣΙήΆβ…χ[71],ΒΪ≤Μάϊ”Ύ…ν»κ÷ΉΝωΉι÷·ΓΘ–ΓΡ…ΟΉΝΘΉ”Θ®<20Ρ…ΟΉΘ©ΨΏ”–…ν»κ÷ΉΝωΉι÷·ΓΔΚΥΡΛΆ®ΆΗ–‘ΚΟΒΡ”≈ ΤΘ§ΒΪΝΘΨΕΧΪ–Γ≤Μ“Ή±ΜΜζΧε«ε≥ΐΘ§‘Ύ÷ΉΝωΡΎ–νΜΐ≤ΜΉψ[72]Θ§”κ«ρ–ΈΩ≈ΝΘœύ±»Θ§ΨΏ”–ΗΏ≥ΛΨΕ±»ΒΡΡ…ΟΉœΥΈ§ΚΆΑτΉ¥–ΈΉ¥‘Ύ÷ΉΝω≤ΩΈΜ±ΘΝτΚΆœΗΑϊ…ψ»ΓΖΫΟφΨΏ”–Οςœ‘”≈ Τ[73]œύ±»÷°œ¬Θ§Ϋœ¥σΒΡœΥΈ§Ή¥ΚΆΑτΉ¥Ρ…ΟΉΫαΙΙΗϋ»ί“Ή±ΜΆχΉ¥ΡΎΤΛœΒΆ≥≤ΕΜώΘ§Ψ≤¬ωΗχ“©Κσ¥φ‘ΎΥ®»ϊΖγœ’[74]Θ§“ρ¥ΥΘ§―Α’““Μ÷÷Φ·≥…Εύ÷÷–ΈΧ§”≈ ΤΒΡ÷«Ρή–ΈΧ§ΉΣΜ·ΒίΥΆœΒΆ≥“ΐΤπΝΥΙψΖΚΒΡ―–ΨΩ–Υ»ΛΓΘΕ‘”ΎΉ‘ΉιΉΑΒΡPDCsΘ§Ω…“‘Ά®ΙΐΒςΩΊΉ‘ΉιΉΑΝΠ Βœ÷–ΈΧ§ΉΣΜ·Θ§Η≥”ηPDCs”≈‘ΫΒΡ÷ΈΝΤ–ßΙϊΓΘ
ΉνΫϋΘ§‘Ϋά¥‘ΫΕύΒΡ―–ΨΩ–Υ»ΛΉΣœρΒςΩΊPDCΉ‘ΉιΉΑΝΠΘ§ΗυΨίΧΊΕ®Φ≤≤Γ÷ΈΝΤΒΡ–η“ΣΕ®÷ΤΉ‘ΉιΉΑκΡΒΡΡ…ΟΉΫαΙΙΘ§…θ÷Ν»ΟΉ‘ΉιΉΑΡ…ΟΉΫαΙΙΗυΨίΦ≤≤ΓΒΡΧΊ’ςœλ”Π–‘ΒΊΖΔ…ζ–ΈΧ§±δΜ·Θ§≤Δ±Μ‘ΛΤΎΚΆ÷Λ ΒΩ…¥οΒΫΗϋΚΟΒΡ–ßΙϊΓΘΜυ”Ύ–ΈΧ§Ε‘“©ΈοADMEΒΡ”ΑœλΘ§―–ΨΩ»Υ‘±÷¬ΝΠ”ΎΩΣΖΔΗς÷÷Ω…±δ–ΈΡ…ΟΉΝΘΉ”Θ®NPsΘ©ΓΘΗς÷÷œλ”Π≤Ώ¬‘“―±Μάϊ”Οά¥¥ΞΖΔPDCsΒΡ≥Ώ¥γΚΆ–ΈΉ¥±δΜ·Θ§άΐ»γΧεΡΎΫΜΝΣΜρΩ«Ζ÷άκ[75]ΓΘ
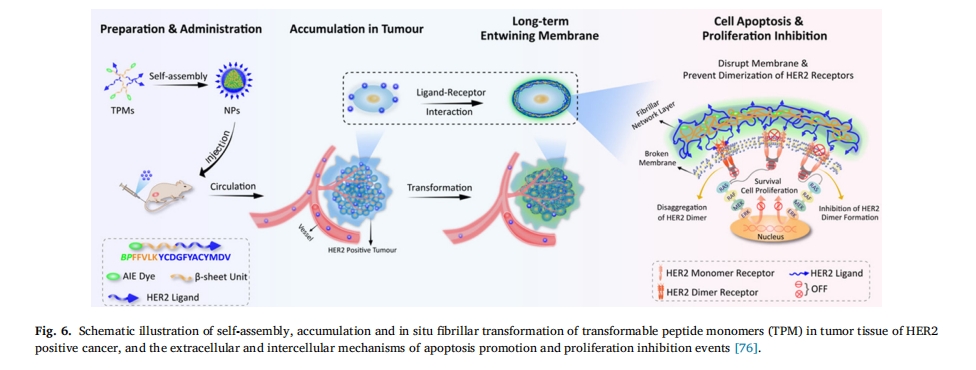
άΐ»γΘ§ZhangΒ»»Υ…ηΦΤΒΡ“Μ÷÷ΈόΕΨΩ…ΉΣΜ·κΡ[76]ΓΘ’β–©κΡΡήΙΜ‘ΎΥ°»ή“Κ÷–Ή‘ΉιΉΑ≥…Ω…ΉΔ…δΒΡ«ρ–ΈΫΚ χΘ§ΉΔ…δΚσ“ΜΒ©Α–œρΫαΚœΑ©œΗΑϊ…œΒΡ HER2Θ§ΥϋΟ«ΨΆΜαΫΜΝΣ–Έ≥…Ρ…ΟΉœΥΈ§Θ§‘ΎΑ©œΗΑϊ±μΟφ–Έ≥…œλ”ΠΓΘΫΜΝΣœΥΈ§ΒΡΗ≤Η«”––ßΒΊΤΤΜΒΝΥ HER2 ΕΰΨέΜ·≤ΔΒΦ÷¬Α©œΗΑϊΒρΆωΘ®ΆΦ 6). Chen Β»±®ΒάΝΥ”…ΕύΑΖ“ΐΖΔΒΡ–ΈΧ§Ω…ΉΣ±δΒΡ≥§Ζ÷Ή”κΡΡ…ΟΉ“©ΈοΘ§Τδ”…≥§Ζ÷Ή”ΒΞ‘ΣΘ®Κυ¬Ϊ[7]κε (CB[7])Θ©ΓΔΠ¬-Τ§≤ψ–Έ≥…ΒΞ‘ΣΘ®κΡ FFVLKΘ©ΚΆ“©ΈοΒΞ‘ΣΘ®œ≤ ςΦν (CPT)Θ©Ήι≥…ΓΘFFVLK-CPT Ι≤ινΈοΆ®Ιΐ CB[7]-Phe-÷ςΩΆΧεœύΜΞΉς”Ο”κ CB[7] Ζ«Ι≤ΦέΗ¥ΚœΘ§ΥφΚσΉ‘ΉιΉΑ≥… NPΘ§Β± CB[7] Ά®ΙΐΑ©œΗΑϊ÷–Ιΐ±μ¥οΒΡœΗΑϊΡΎΨΪΑΖΫβάκ ±Θ§NP Ω…“‘ΉΣΜ·ΈΣΈΔœΥΈ§Θ§œ‘÷χ‘ω«Ω CPT ‘ΎΑ©œΗΑϊ÷–ΒΡΜΐάέΚΆ±ΘΝτΘ§≤ΔΫΒΒΆΧεΡΎœΒΆ≥ΕΨ–‘Θ®ΆΦ 7Θ©[77]. Ι®Β»»Υ[78]ΫΪ“ΏΟγ”κκΡΫαΚœΘ§…ηΦΤ≥ω“Μ÷÷ΨΏ”–Ω…ΗΡ±δ–ΈΧ§ΒΡΗΏ–ßΩΙ÷ΉΝω“ΏΟγΓΘ÷ΉΝωΒΡΥα–‘ΈΔΜΖΨ≥Ω…”’ΒΦΡ…ΟΉ“ΏΟγΖΔ…ζœ‘÷χΒΡ–ΈΧ§±δΜ·Θ§¥”Ρ…ΟΉ«ρΘ®÷±ΨΕ‘Φ 100 Ρ…ΟΉΘ©ΒΫΡ…ΟΉΤ§Θ®≥ΛΕ»ΜρΩμΕ»ΈΣΦΗΈΔΟΉΘ©ΓΘ≥… λΒΡΉιΉΑΧεΩ…“‘Μζ–ΒΒΊΤΤΜΒΡΎΧεΡΛΘ§ΩΙ‘≠κΡ±Μ÷±Ϋ”‘ΥΥΆΒΫœΗΑϊ÷ ÷–ΓΘ÷Ί–¬ΉιΉΑΒΡΡ…ΟΉΤ§ΜΙΆ®ΙΐΦΛΜνΧΊΕ®ΒΡ―Ή÷ΔΆΨΨΕά¥‘ω«Ω÷ΉΝωΟβ“ΏΝΠΓΘ
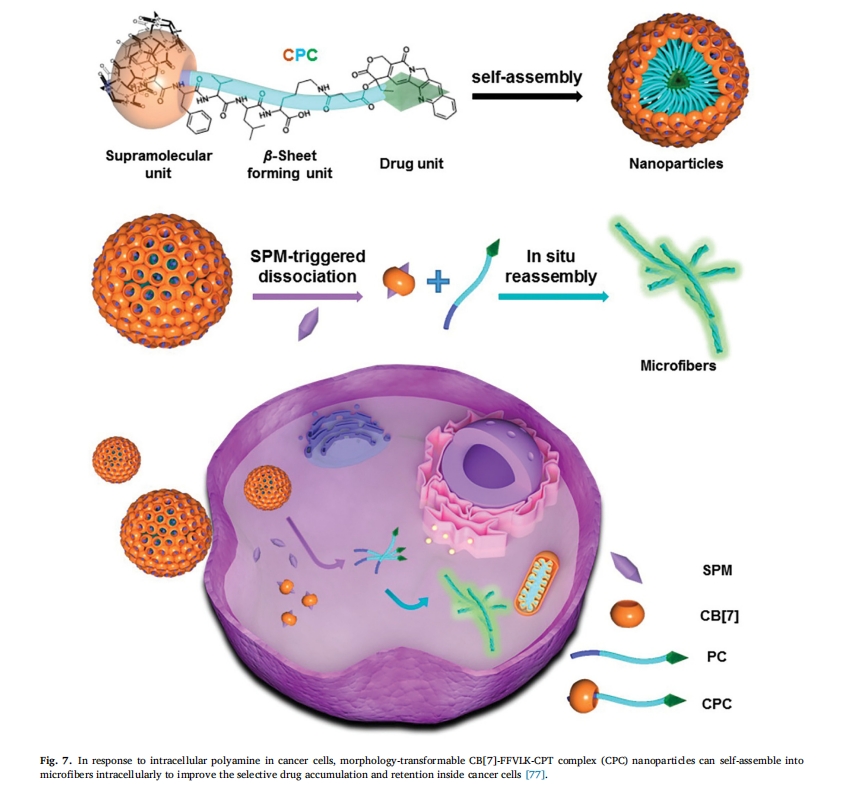
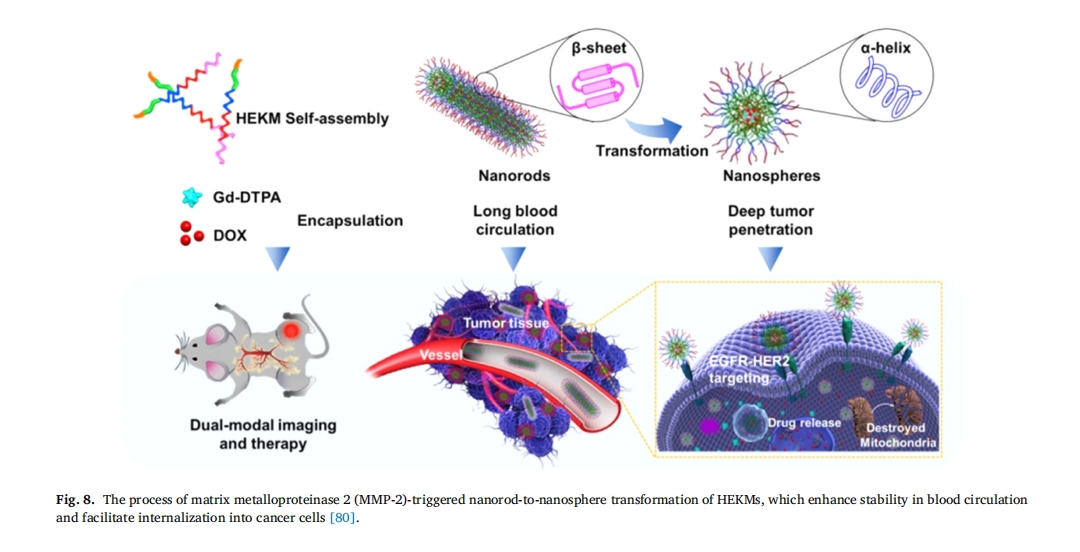
¥σΝΩ―–ΨΩ±μΟςΘ§ΑτΉ¥Ρ…ΟΉΝΘΉ”―≠ΜΖ ±ΦδΫœ≥ΛΘ§”–άϊ”Ύ“©Έο‘Ύ≤Γ‘νΡΎ≥÷–χ–νΜΐΘ§Εχ«ρ–ΈΡ…ΟΉΝΘΉ”ΨΏ”–…νΕ»œΗΑϊ¥©ΆΗΡήΝΠ[79]ΓΘ“ρ¥ΥΘ§Wang Β»»Υ[80]. ΩΣΖΔΝΥ“Μ÷÷Μυ”Ύ÷ΉΝωΒςΩΊΒΡ÷«ΡήΫΚ χœΒΆ≥Θ§≥ΤΈΣHEKMΘ§ΗΟœΒΆ≥”…ΑτΉ¥Ω≈ΝΘΉι≥…Θ§’β–©Ω≈ΝΘ ή÷ΉΝωΈΔΜΖΨ≥ΒςΫΎΕχΉΣ±δΈΣΗϋ–ΓΒΡΫΚ χΓΘΡ…ΟΉ«ρ–ΈΫΚ χΉψΙΜ–ΓΘ§Ω…“‘‘Ύ÷ΉΝω÷–ΨέΦ·Θ§ Βœ÷…ν≤ψ…χΆΗΓΘ’β÷÷Ατ«ρΉΣΜ·ΒΡΡ…ΟΉΒίΥΆœΒΆ≥‘ω«ΩΝΥΑΔΟΙΥΊΘ®DOXΘ©ΒΡœΗΑϊΑ–œρ–‘ΓΔΡΎΜ·–‘ΚΆ÷ΉΝω÷ΈΝΤΡήΝΠΘ§ΧαΗΏΝΥ÷ΈΝΤ–ßΙϊΘ®ΆΦ 8Θ©ΓΘ
2.3 Α–œρκΡ
PDCs ΒΡ”Π”ΟΡΩ±ξ «Ά®Ιΐ―”≥ΛΫœΕΧΒΡ―≠ΜΖΑκΥΞΤΎΚΆΧαΗΏΑ–œρ–߬ ά¥‘ω«ΩΜ·ΝΤ“©ΈοΒΡΝΤ–ßΓΘ≥Θ”ΟΒΡΑ–œρ≤Ώ¬‘Αϋά®Ά®Ιΐ EPR –ß”Π±ΜΕ·Α–œρ÷ΉΝωΉι÷·Θ§“‘ΦΑάϊ”ΟΑ–ΈΜΗΏ±μ¥οΜρΧΊ“λ–‘±μ¥οΒΡ ήΧε Βœ÷Α–œρ“©ΈοΒίΥΆΓΘ”κ¥ΪΆ≥ΩΙΧεΒ»ΗΏΖ÷Ή”ΝΩΚΆΗ¥‘”ΫαΙΙΒΡΑ–œρ≈δΧεœύ±»Θ§–ΓΖ÷Ή”Α–œρκΡ ήΒΫ‘Ϋά¥‘ΫΕύΒΡ―–ΨΩΙΊΉΔΓΘΜυ”Ύ‘Ύ÷ΉΝω―ΣΙή…œΙψΖΚΙΐ±μ¥οΒΡ’ϊΚœΥΊΦ“Ήε≥…‘±Θ§ΉνΒδ–ΆΒΡ÷ΉΝωΙι≥≤κΡΜυ–ρ « RGD[81]. ΩΤ“ΝΈδΡΰ[82]Β»±®ΒάΠΝ5Π¬1’ϊΚœΥΊΫαΚœΦΝΨυΚ§”–RGDΘ§ΜΖΉ¥κΡ±μœ÷≥ω±»œΏ–‘ΫαΚœΗϋ«ΩΒΡΫαΚœΝΠΓΘΥφΚσΘ§ΖΔœ÷ΝΥ–μΕύΨΏ”–÷ΉΝω¥©ΆΗΡήΝΠΒΡRGD–ρΝ–Θ®Φ«ΈΣiRGDΘ§ΓΑiΓ±¥ζ±μΡΎΜ·Θ©[83]Θ§‘ΎΙΐ»ΞΒΡΦΗ °ΡξάοΘ§iRGD ‘Ύ–μΕύ÷ΉΝωΡΘ–Ά÷–‘ω«ΩΝΥ“©ΈοΒΡΜΐάέΚΆΝΤ–ß[84Θ§85].2018ΡξΘ§iRGDΉςΈΣΦΣΈςΥϊ±θΚΆabraxaneΒΡ“»œΌΑ©ΧμΦ”ΦΝΫχ»κIΤΎΝΌ¥≤ ‘―ιΘ®ΝΌ¥≤ ‘―ιΚ≈ΘΚNCT03517176Θ©ΓΘ
Νμ“Μ÷÷÷ΉΝωΑ–œρκΡ «œΗΑϊΆβΜυ÷ (ECM) Α–œρκΡΘ§“ρΈΣ ECM ΒΡ≥…Ζ÷‘ΎΦΗΚθΥυ”– ΒΧε÷ΉΝω÷–ΕΦΙΐΕ»±μ¥οΓΘΙΊ”Ύ ECM Α–œρκΡΒΡ±®ΗφΑϋά®œΥΝ§ΒΑΑΉ”ρΆβ B (EDB-FN) ΚΆΦΓκλΒΑΑΉ-C ΫαΚœκΡ [86Θ§87], ≤ψ’≥Ν§ΒΑΑΉ-≥≤ΒΑΑΉΗ¥ΚœΈοΫαΚœκΡ[88]ΓΔΜυ÷ Ϋπ τΒΑΑΉΟΗΑ–œρκΡΓΔΫαΒόΉι÷·…ζ≥Λ“ρΉ” (CTGF/CCN2) ΫαΚœκΡ[89]“‘ΦΑ‘¥Ή‘ΧΞ≈Χ…ζ≥Λ“ρΉ” (PlGF-2) ΒΡ ECM ΫαΚœκΡ[90]Hubbell ΦΑΤδΆ§ ¬Ά®ΙΐΡΘΖ¬Χλ»Μ≈δΧεΩΣΖΔΝΥ ECM ΫαΚœκΡΘ§’β–©κΡ‘ΎΑ©÷Δ÷ΈΝΤΖΫΟφΨΏ”–Νν»Υ–ΥΖήΒΡ”Π”ΟΓΘΝμ“ΜΗωΨΏ”–Ψό¥σ«±ΝΠΒΡάΐΉ” « Bicycle Therapeutics ΩΣΖΔΒΡκΡ-“©Έο≈ΦΝΣΈο (BT1718)[91]BT1718”…ΥΪΜΖ≈δΧεΉι≥…Θ§Ά®ΙΐΕΰΝρΦϋΩ…Ν―ΫβΒΡΝ§Ϋ”Χε”κΟάΒ«ΡΨΥΊ≈ΦΝΣΘ§Ω…Α–œρECM÷–ΒΡΜυ÷ Ϋπ τΒΑΑΉΟΗ14Θ®MMP-14Θ©ΓΘBT1718ΒΡI/IIaΤΎ ‘―ι’ΐ‘ΎΜυ”ΎΆμΤΎ ΒΧεΝωΜΦ’ΏΘ®»ΐ“θ–‘»ιœΌΑ©ΚΆΖ«–ΓœΗΑϊΖΈΑ©ΘΜΝΌ¥≤ ‘―ι±ύΚ≈ΘΚNCT03486730Θ©Ϋχ––ΓΘ
¥ΥΆβΘ§≤Γ±δ≤ΩΈΜΧΊ“λ–‘±μ¥οΒΡ≤ΜΆ§ ήΧε“≤ «Α–œρκΡΒΡ÷Ί“ΣΑ–ΒψΓΘάΐ»γΘ§»ιœΌΑ©ΓΔ¬―≥≤Α©ΓΔΉ”Ι§ΡΎΡΛΑ©ΚΆ«ΑΝ–œΌΑ©Β»Α©÷Δ÷–Ιΐ±μ¥οΒΡLHRH ήΧεΘ§ΈΣΑ–œρ“©Έο δΥΆΧαΙ©ΝΥΦΪΚΟΒΡ≤Ώ¬‘ΓΘΫΪDOXΚΆ2-ΏΝΩ©Άι-DOX”κLHRHΑ–œρκΡΫαΚœΘ§Ω…“‘‘ωΦ”Α©œΗΑϊΒΡΑ–œρ–‘Θ§ΫΒΒΆ“©ΈοΕΨ–‘ΓΘLindgrenΒ»[92]. Ν¥Ϋ” YTA2ΚΆ YTA4κΡ”κΦΉΑ±ΒϊΏ ΫαΚœ–Έ≥…PDCΓΘ”κ”ΈάκΦΉΑ±ΒϊΏ œύ±»Θ§ΫαΚœΗ¥ΚœΈο ΙΑ–ΟΗΕΰ«β“ΕΥαΜΙ‘≠ΟΗΒΡΉς”Ο‘ωΦ”ΝΥ15-20±ΕΓΘ‘¥Ή‘œΗΑϊ±μΟφΒΑΑΉΒΡœΗΑϊΦδ’≥ΗΫΖ÷Ή”-1Θ®ICAM-1Θ©κΡ–ρΝ–Ω…“‘±ΜΑΉ―Σ≤ΓTœΗΑϊΧΊ“λ–‘ΒΊΡΎΆΧΓΘICAM-1κΡΩ…“‘”κ÷ΈΝΤ“©ΈοΝ§Ϋ”Θ§¥”ΕχΩ…“‘ΧαΗΏ“©Έο‘ΎΑ©÷ΔΚΆΉ‘…μΟβ“Ώ÷ΈΝΤ÷–ΒΡ”––ß–‘ΓΘ’β÷÷Φ≤≤ΓΒΡ÷ΈΝΤ–ßΙϊΦθ…ΌΝΥΜ·ΝΤΒΡΗ±Ής”ΟΓΘJeffreyΒ»»ΥΖΔœ÷10ΚΆ20ΨέΧεΒΡΧλΕ§Α±ΥαΚΆΙ»Α±Υα‘ΎΑ–œρΙ«’έΖΫΟφΖ«≥Θ”––ßΓΘΥϊΟ«ΒΡ Β―ι±μΟςΘ§Υα–‘Α±ΜυΥαΒΡΕΧΙ―κΡΩ…“‘ΫΪκΡ”––ß‘ΊΚ…Ε®ΈΜΒΫΙ«’έ¥Π91ΓΘ±»Ε‘’’Ζ«Α–œρκΡ”––ß‘ΊΚ…”––ß 9 ±Ε[93]. ΗϋΕύΒΡΥα–‘κΡΫαΚœΈο’ΐ‘Ύ―–ΨΩ÷–”Ο”Ύ―Γ‘ώ–‘ΒΊΫΪΗς÷÷“©Έο δΥΆΒΫΙ«ςάΘ§Αϋά®ΦΛΥΊΓΔΩΙ…ζΥΊΓΔ–ΓΗ…»≈ RNA (siRNA) ΚΆΜυ“ρ÷ΈΝΤ[94®C98]±μ 2ΓΘ
ΉήΫαΝΥ”Ο”ΎΙΙΫ®Α–œρΒίΥΆœΒΆ≥ΒΡκΡΒΡΉν–¬―–ΨΩΓΘ
2.4 œΗΑϊ¥©ΆΗκΡ
œΗΑϊΡΛ «“ΜΒά÷Ί“ΣΒΡ…ζάμΤΝ’œΘ§œό÷ΤΝΥ–μΕύΒΑΑΉ÷ ΓΔΚΥΥαΒ»…ζΈο¥σΖ÷Ή”ΚΆΡ…ΟΉ‘ΊΧεΘ®÷§÷ ΧεΓΔΫΚ χΓΔΡ…ΟΉΩ≈ΝΘΘ©Ωγ÷ ΡΛ‘Υ δΘ§“ρ¥ΥΧαΗΏ“©ΈοΆΗΙΐœΗΑϊΡΛΒΡ–ß¬ ”»ΈΣ÷Ί“ΣΓΘœΗΑϊ¥©ΆΗκΡΘ®CPPsΘ©ά¥‘¥”ΎΉΣ‘ΥΒΑΑΉΒΡΉΣΒΦΫαΙΙ”ρΘ§ «“ΜάύΡή¥©ΆΗœΗΑϊΡΛΒΡ–ΓΖ÷Ή”ΕΧκΡΘ§“ΜΑψ”…5~30ΗωΑ±ΜυΥα≤–ΜυΉι≥…ΓΘ¥©ΆΗκΡΗΜΚ§ΨΪΑ±ΥαΓΔάΒΑ±ΥαΒ»Φν–‘Α±ΜυΥαΘ§ΨΏ”–…ζάμΧθΦΰœ¬¥χ’ΐΒγΚ…ΓΔΝΫ«Ή–‘ΓΔ¥©ΆΗœΗΑϊΡΛΡήΝΠ«ΩΒ»ΧΊΒψΓΘΥϋΩ…“‘‘ΎΧεΡΎΚΆΧεΆβΩλΥΌΒΦ÷¬“©ΈοΦΑΤδ‘ΊΧεΩγ‘ΫœΗΑϊΡΛΤΝ’œΘ§ «Ήν”–«ΑΆΨΒΡ“©Έο δΥΆ‘ΊΧε÷°“Μ[138]Ή‘1994ΡξFawellΒ»±®ΒάΒΎ“ΜΗωά¥‘¥”ΎHIV-1ΒΑΑΉΘ®RKKKRRQRRRΘ©ΒΡœΗΑϊ¥©ΆΗκΡTat“‘ά¥Θ§“―ΖΔœ÷Εύ÷÷œΗΑϊ¥©ΆΗκΡΘ§Αϋά®TATΓΔMAPΓΔtransportanΚΆΗΜΚ§Arg/Lys≤–ΜυΒΡ–≈Κ≈ΉΣΒΦκΡVP22ΓΘCPPΡΎΜ·ΒΡΕύ÷÷ΡΘ–Ά“―±ΜΧα≥ωΘ§”κCPPΩγΡΛœύΙΊΒΡ±ΜΙψΖΚ»œΩ…ΒΡΜζ÷Τ÷ς“Σ «ΡήΝΩΕάΝΔΒΡ÷±Ϋ”ΩγΡΛΜζ÷ΤΘ®Αϋά®ΒΙΫΚ χΡΘ–Ά[139]Θ§ΩΉœΕ–Έ≥…[140]“‘ΦΑΓΑΒΊΧΚΓ±ΡΘ–Ά[141Θ§142]Θ©ΘΜΝμ“Μάύ «ΡήΝΩ“άάΒ–‘ΡΎΆΧΉς”ΟΘ®Αϋά®–ΓΈ―ΚΆΆχΗώΒΑΑΉΖ«“άάΒΜρ“άάΒ–‘ΆΨΨΕ[143®C145]ΚΆΨόΑϊ“ϊΉς”Ο[146]Θ©CPP ΡΎΜ·ΡΘ–ΆΒΡ’Σ“Σ»γœ¬ΘΚΆΦ 9aΓΘ
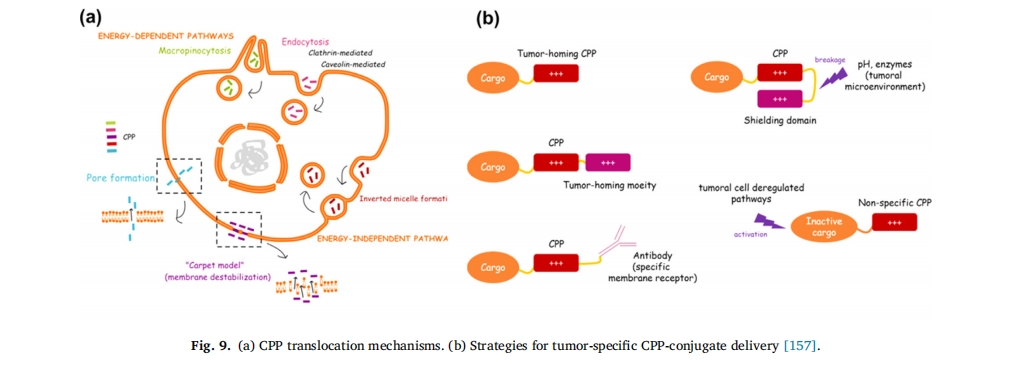
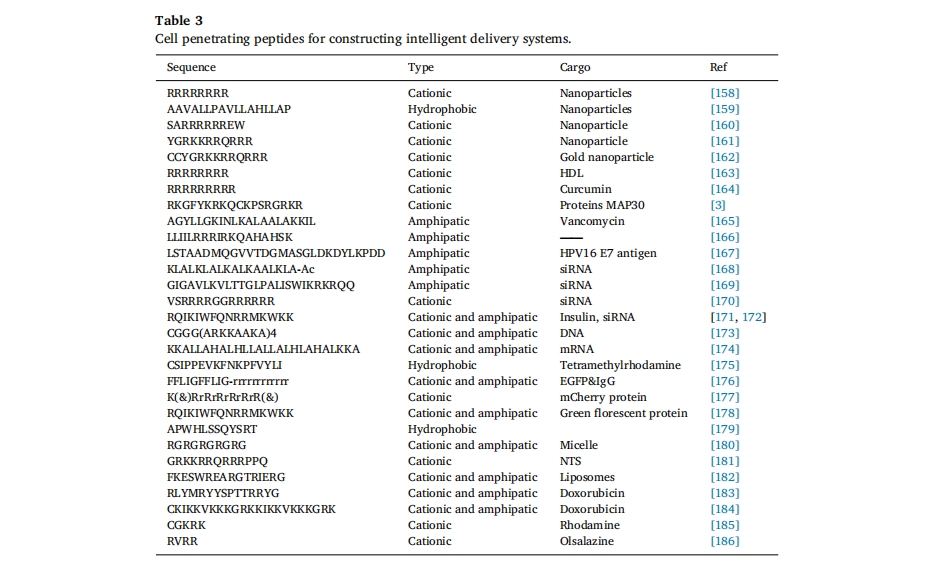
ΥφΉ≈CPPsΉς”ΟΜζάμΒΡ÷πΫΞ≤ϊΟςΘ§¥σΝΩΗϋΗΏ–ßΒΡΙΠΡή–‘…χΆΗΦΝ–ρΝ–±ΜΚœ≥…[147]Θ§÷ΉΝω…χΆΗ»‘»Μ « CPP ―–ΨΩΒΡ÷ΊΒψΘ§÷ΉΝωΧΊ“λ–‘ CPP ΫαΚœΈοΒίΥΆ≤Ώ¬‘»γœ¬ΆΦ 9b. ¥ΥΆβΘ§Ωγ‘Ϋ―ΣΡ‘ΤΝ’œ « CPP ΩΣΖΔΒΡΝμ“ΜΗω”–œΘΆϊ«“ΨΏ”–Χτ’Ϋ–‘ΒΡΝλ”ρΓΘ“―ΖΔœ÷ΗϋΕύ CPP ΡήΙΜΫΪ…ώΨ≠÷ΈΝΤ“©Έο‘ΥΥΆΒΫΡ‘ Β÷ ÷–ΓΘ’β–© CPP Έό–η ήΧεΦ¥Ω…¥©ΆΗ―ΣΡ‘ΤΝ’œΘ§”…ΕΧΒΡΝΫ«Ή–‘ΚΆ/Μρ―τάκΉ”–ρΝ–Ήι≥…Θ§ΨΏ”–Ά®ΙΐΈϋΗΫΫιΒΦΒΡΉΣΑϊΆΧΜζ÷ΤΩγ‘ΫœΗΑϊΡΛΒΡΡήΝΠ[148]¥ΥΆβΘ§Μυ”Ύ CPP ΒΡΤδΥϊΙΠΡή“≤ΒΟΒΫΝΥΖΔ’ΙΘ§“‘‘ω«ΩΩγ BBB ΡήΝΠΘ§Αϋά®Ε‘ΧΊΕ® ήΧεΒΡ«Ω«ΉΚΆΝΠΚΆΫιΒΦΉΣΑϊΆΧΉς”ΟΒΡΡήΝΠ[149]Θ®ΆΦ 10)ΓΘKwonΒ»»Υ…ηΦΤΝΥ“Μ÷÷Μυ”Ύ¥©ΆΗκΡTAT-ΧλΕ§θΘΑΖΟΗΗ¥ΚœΈοΒΡ’κΕ‘ΝήΑΆœΗΑϊΑΉ―Σ≤ΓΒΡ”––ßΒίΥΆœΒΆ≥[150]. ≥ΐΝΥ‘ω«Ω÷ΉΝω…χΆΗ–‘÷°ΆβΘ§CPP ΜΙ±Μ”Ο”ΎΩΎΖΰ“»ΒΚΥΊΒΡ≥Δ ‘ΓΘΨί±®ΒάΘ§R8 κΡ±Μ”Ο”Ύ–ό Έ“»ΒΚΥΊ NPs “‘ΩΥΖΰ’≥“ΚΚΆ…œΤΛΤΝ’œΘ§ Ι“»ΒΚΥΊ…ζΈοάϊ”ΟΕ»ΧαΗΏ 1.9 ±ΕΘ§“ρ¥Υœ‘ Ψ≥ωΩΎΖΰκΡ/ΒΑΑΉ÷ ΒίΥΆΒΡΨό¥σ«±ΝΠ[151]¥”÷°«ΑΒΡΩΣ¥¥–‘ΙΛΉςά¥Ω¥Θ§CPPsite 2.0 ΐΨίΩβΑϋΚ§¥σ‘Φ 1850 Ηω CPP –ρΝ–Θ§‘ΛΦΤ’β“Μ ΐΉ÷ΫΪΦΧ–χ‘ωΦ”[152]. ≥§Ιΐ 25 ÷÷ CPP ΫαΚœ“©Έο’ΐ‘ΎΝΌ¥≤ΩΣΖΔ÷–Θ§”Ο”Ύ÷ΈΝΤ―Ή÷ΔΓΔΧέΆ¥ΓΔΑ©÷ΔΓΔ–Ρ‘ύ≤ΓΚΆΥΞάœΒ»Εύ÷÷Φ≤≤Γ[153®C156]±μ 3ΓΘ
ΉήΫαΝΥΫϋΦΗ °Ρξά¥±®ΒάΒΡ¥σΝΩœΗΑϊ¥©ΆΗκΡΒΡ―–ΨΩΘ§’β–©κΡ“―≥…ΙΠ”Π”Ο”Ύ‘ω«ΩΒΑΑΉ÷ ΓΔΚΥΥαΚΆ–ΓΖ÷Ή”“©ΈοΒΡΙΠ–ßΓΘ
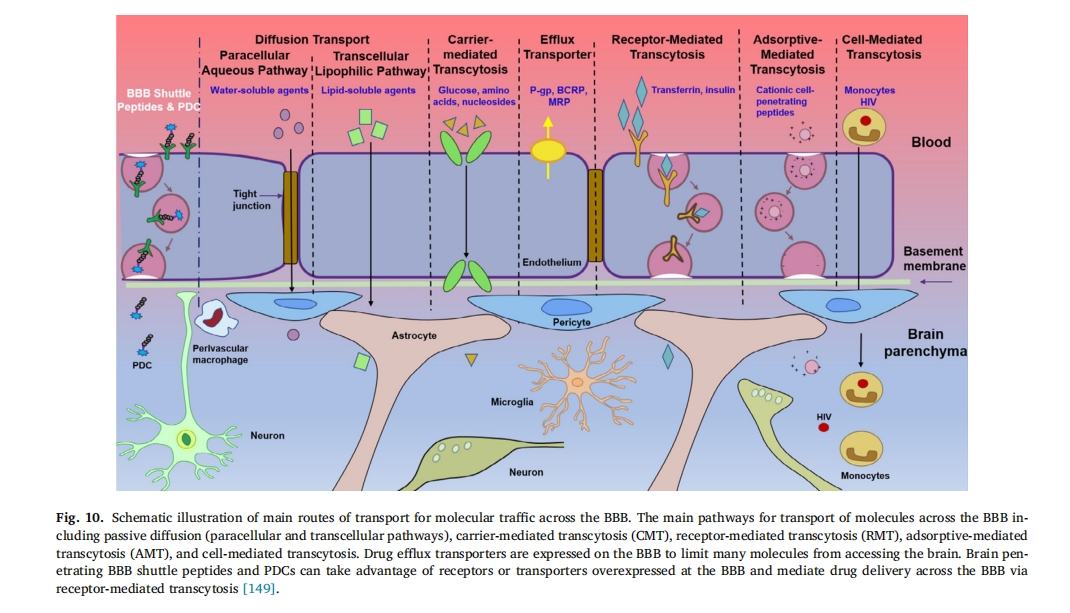
2.5 Ζ¥”ΠκΡ
œλ”Π–‘κΡΫΪ ήΒΫœΒΆ≥/Άβ≤ΩΜΖΨ≥±δΜ·ΒΡ”ΑœλΘ§“‘¥ΧΦΛκΡΉ‘ΉιΉΑΜρ“©Έο ΆΖ≈Θ§Αϋά®Έ¬Ε»ΓΔpHΓΔΟΗΓΔGSH ΚΆάκΉ”≈®Ε»Β»Θ§ΈΣκΡ“©Έο δΥΆœΒΆ≥ΒΡΙΙΫ®ΧαΙ©Ω…ΩΊ–‘ΓΘΈ¬Ε»œλ”Π–‘κΡΆ®≥Θ”Ο”ΎΙΙΫ®Ω…ΉΔ…δΒΡ»»œλ”ΠΥ°ΡΐΫΚΘ§”Ο”Ύ“©Έο δΥΆΚΆœΗΑϊΖβΉΑΓΘάΐ»γΘ§ά¥Ή‘Β·–‘ΒΑΑΉ (ELP) ΒΡΈεκΡ–ρΝ– VPGXG ΒΡΙ―Ψέ÷ΊΗ¥–ρΝ–‘ΎΒΆ”Ύ 35ΓψC ΒΡΈ¬Ε»œ¬»ήΫβΘ§‘ΎΧεΡΎΉΔ…δΚσ‘ΎΧεΈ¬ (37ΓψC) œ¬–Έ≥…Υ°ΡΐΫΚΓΘ“ρ¥ΥΘ§ELP ―υκΡΩ… ΙœΗΑϊΚΆΤδΥϊ÷ΈΝΤΦΝ‘Ύ “Έ¬œ¬”κ»ή“Κ÷–ΒΡΉ‘…μΫαΚœΘ§≤Δ‘Ύ…ζάμΈ¬Ε»œ¬–Έ≥…ΡΐΫΚΉ¥Μυ÷ Θ§”Ο”ΎΉι÷·ΙΛ≥ΧΚΆΤδΥϊ“ΫΝΤ”Π”ΟΓΘ”–ΦΗ÷÷Φ≤≤ΓΜαΒΦ÷¬ pH ΤΫΚβΗΡ±δΘ§άΐ»γ―Ή÷ΔΚΆΑ©÷ΔΓΘκΡ“≤Ω…“‘Κœάμ…ηΦΤ“‘œλ”ΠΥα–‘Θ§»ΜΚσ ΆΖ≈±Μ≤ΕΜώΒΡΖ÷Ή”Θ§άΐ»γ“©ΈοΓΔΒΑΑΉ÷ ΚΆΚΥΥαΓΘΑ©œΗΑϊΒΡΨ÷≤ΩœΗΑϊΆβ pH ÷ΒΩ…Ρή±»ΫΓΩΒœΗΑϊΒΆ 0.5-1 Ηω pH ΒΞΈΜΘ§’β‘ –μ Ι”Ο pH œλ”ΠκΡΕ‘Υα–‘ΜΦ≤ΓΉι÷·Εχ≤Μ «ΫΓΩΒΉι÷·ΨΏ”–ΗϋΗΏΒΡΑ–œρΧΊ“λ–‘κΡ“≤Ω…“‘Κœάμ…ηΦΤΘ§ ΙΤδΕ‘Υα–‘Ής≥ωΖ¥”ΠΘ§»ΜΚσ ΆΖ≈±Μ≤ΕΜώΒΡΖ÷Ή”Θ§»γ“©ΈοΓΔΒΑΑΉ÷ ΚΆΚΥΥαΓΘΑ©œΗΑϊΒΡΨ÷≤ΩœΗΑϊΆβ pH ÷ΒΩ…Ρή±»ΫΓΩΒœΗΑϊΒΆ 0.5-1 Ηω pH ΒΞΈΜΘ§“ρ¥ΥΘ§Ά®Ιΐ Ι”Ο pH œλ”ΠκΡΘ§Ω…“‘Ε‘Υα–‘ΜΦ≤ΓΉι÷·Θ®Εχ≤Μ «ΫΓΩΒΉι÷·Θ© Βœ÷ΗϋΗΏΒΡΑ–œρΧΊ“λ–‘κΡ“≤Ω…“‘Κœάμ…ηΦΤΘ§ ΙΤδΕ‘Υα–‘Ής≥ωΖ¥”ΠΘ§»ΜΚσ ΆΖ≈±Μ≤ΕΜώΒΡΖ÷Ή”Θ§»γ“©ΈοΓΔΒΑΑΉ÷ ΚΆΚΥΥαΓΘΑ©œΗΑϊΒΡΨ÷≤ΩœΗΑϊΆβ pH ÷ΒΩ…Ρή±»ΫΓΩΒœΗΑϊΒΆ 0.5-1 Ηω pH ΒΞΈΜΘ§“ρ¥ΥΘ§Ά®Ιΐ Ι”Ο pH œλ”ΠκΡΘ§Ω…“‘Ε‘Υα–‘ΜΦ≤ΓΉι÷·Θ®Εχ≤Μ «ΫΓΩΒΉι÷·Θ© Βœ÷ΗϋΗΏΒΡΑ–œρΧΊ“λ–‘[187]. ΝΚΒ»»Υ[188]≥…ΙΠ÷Τ±ΗΝΥ“Μ÷÷–¬–Ά pH œλ”ΠκΡΝΫ«ΉΡ…ΟΉΉιΉΑΧεΘΜ‘Ύ’βάοΘ§ΥϋΒΡΕΰΦΕΫαΙΙΩ…“‘¥” pH 7.4 ±ΒΡ Π¬ Τ§≤ψ±δΈΣ pH 5.0 ±ΒΡ ΠΝ ¬ί–ΐΘ§¥”ΕχΒΦ÷¬–ΈΧ§¥”Ρ…ΟΉ«ρ―ί±δΈΣΡ…ΟΉœΥΈ§ΓΘMatthew Β»»Υ±®ΒάΝΥ“Μ÷÷ pH œλ”ΠκΡΜυΡ…ΟΉœΥΈ§ΉιΉΑΧεΘ§ΗΟΉιΉΑΧεάϊ”ΟΤœΧ―Χ«―θΜ·ΟΗ (GOx) Ά®ΙΐœϊΚΡ–Έ≥…ΒΡ…ζάμΤœΧ―Χ«ΓΑ»ΦΝœΓ±ά¥ΫΒΒΆΨ÷≤Ω pH ÷ΒΘ§¥”Εχ«ΐΕ·Ζ«ΤΫΚβκΡΥ°ΡΐΫΚΆχ¬γΓΘ‘ΎΒΆ―ΣΧ«ΖΔΉςΤΎΦδΘ§―ΣΧ«œ¬ΫΒΒΦ÷¬ GOx ≤ζ…ζΒΡΤœΧ―Χ«ΥαΦθ…ΌΘ§…ζάμΜΖΨ≥÷–ΒΡ pH ÷Β…ΐΗΏΓΘpH œλ”ΠκΡΥ°ΡΐΫΚΥφΉ≈ pH ÷ΒΒΡ…ΐΗΏΕχ»ήΫβ≤Δ ΆΖ≈“»ΗΏ―ΣΧ«ΥΊΘ§ΧαΙ©ΤœΧ―Χ«-pH œλ”ΠΡ…ΟΉœΥΈ§Υ°ΡΐΫΚΖΫΖ®ά¥ δΥΆ“»ΗΏ―ΣΧ«ΥΊά¥Ε‘ΩΙΒΆ―ΣΧ«[189]ΓΘ
“Μ–©Ζ¥”Π–‘κΡΩ…ΉςΈΣΟΗΒΉΈοΘ§Τδ“©ΈοΒίΥΆΧΊ–‘Μα ήΒΫΧεΡΎΟΗΒΡ”ΑœλΓΘΨί±®ΒάΘ§Εύ÷÷ΟΗ‘Ύ÷ΉΝω≤ΩΈΜΚΆ»±―Σ«χ”ρΨέΦ·ΒΡΤΒ¬ ΗϋΗΏΘ§άΐ»γ MMPΓΔΉι÷·ΒΑΑΉΟΗΚΆΦν–‘ΝΉΥαΟΗΓΘJin Zhang Β»[190]. ΩΣΖΔΝΥMMP-2œλ”Π«ΕΚœκΡΘ§ΗΟκΡΩ…“‘‘Ύ…ζάμΧθΦΰœ¬Ή‘ΉιΉΑ≥…«ρ–ΈNPΘ§Β±ΟΗMMP-2‘Ύ÷ΉΝω≤ΩΈΜΙΐ±μ¥ο ±Θ§Ά®ΙΐΫΒΒΆCAC÷ΒΉΣΜ·ΈΣ≥ΛΡ…ΟΉœΥΈ§ΓΘ«ρ–ΈΒΫœΥΈ§ΒΡΉΣ±δΩ…“‘‘ωΦ”÷ΉΝω÷–ΒΡΜθΈοGd 3+≈®Ε»Θ§¥”ΕχΗΡ…ΤMRI–≈Κ≈ΓΘ¥ΥΆβΘ§Jinseong KimΒ»»Υ÷Τ±ΗΒΡ«Α“©NP[191].”…Ή‘ΉιΉΑΒΡΒΑΑΉΟΗBΧΊ“λ–‘Ω…Ν―ΫβκΡΘ®FRRGΘ©ΚΆDOXΫαΚœΈοΉι≥…Θ§Ά®ΙΐΑ©œΗΑϊ÷–ΙΐΕ»±μ¥οΒΡΒΑΑΉΟΗBΫιΒΦΒΡΟΗ¥ΌΫΒΫβΧΊ“λ–‘ ΆΖ≈DOXΓΘ
3 Ν§Ϋ”Τ§ΕΈ
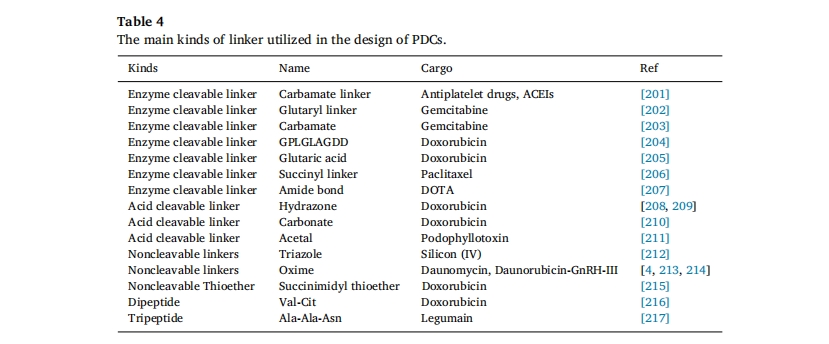
Ν§Ϋ”Μυ «―”≥ΛPDC―≠ΜΖ ±ΦδΚΆΩΊ÷ΤΜθΈοœλ”Π–‘ ΆΖ≈ΒΡΙΊΦϋΘ§“ΜΑψ―Γ‘ώΕύκΡΡ©ΕΥΘ®Αϋά®ΕύκΡΒΡNΕΥΜρCΕΥΘ©ΚΆΑκκΉΑ±ΥαΕΥΜυΉςΈΣΝ§Ϋ”ΈΜΒψΘ§”…”ΎΡ©ΕΥΝ§Ϋ”Ε‘ΕύκΡΙΠΡή”Αœλ≤Μ¥σΘ§¬μά¥θΘ―«ΑΖ-ΑκκΉΑ±ΥαMichaelΦ”≥…Ζ¥”ΠΨΏ”–Ζ¥”ΠΕ·ΝΠ―ßΩλΓΔ―Γ‘ώ–‘ΗΏΓΔΖ¥”ΠΧθΦΰΈ¬ΚΆΒ»ΧΊΒψΘ§±ΜΙψΖΚ”Ο”ΎPDCΚœ≥…Ζ¥”ΠΈΜΒψ[192]ΓΘ¥ΥΆβΘ§”…”Ύ–μΕύκΡ‘Ύ–ρΝ–÷–ΫωΚ§”–“ΜΗωΑκκΉΑ±Υα≤–ΜυΉςΈΣΖ¥”ΠΈΜΒψΘ§“ρ¥ΥΩ…“‘ΧαΗΏ≈ΦΝΣ–ß¬ ΓΘΝ§Ϋ”Ή”±Ί–κ‘Ύ―≠ΜΖ÷–±μœ÷≥ωΈ»Ε®–‘Θ§“‘Ζά÷Ι“©ΈοΙΐ‘γΚΆΖ«ΧΊ“λ–‘ ΆΖ≈Θ§’βΕ‘PDCs‘ΎΧεΡΎ‘Υ δΙΐ≥Χ÷–ΒΡΈ»Ε®–‘ΤπΉ≈÷ΝΙΊ÷Ί“ΣΒΡΉς”ΟΓΘΝ§Ϋ”Ή”ΒΡ…ηΦΤ»ΓΨω”Ύ…φΦΑPDCsΒΡΜζ÷ΤΚΆΦ≤≤Γ÷ΈΝΤΒΡ–η“ΣΓΘΝ§Ϋ”Ή”ΒΡ±δΜ·Η≥”ηΒίΥΆœΒΆ≥Εύ÷÷ΙΠΡήΘ§’βΫΪΕ‘’ϊΗωœΒΆ≥ΒΡ–‘ΡήΚΆ“©ΈοΗΚ‘Ί≤ζ…ζ÷Ί¥σ”ΑœλΓΘ”Ο”ΎPDCsΒΡΝ§Ϋ”Φϋάύ–ΆΕύ÷÷Εύ―υΘ§Ω…Ζ÷ΈΣΩ…Ν―ΫβΚΆ≤ΜΩ…Ν―Ϋβ–Έ Ϋ[193]Ω…Ν―ΫβΒΡΝ§Ϋ”ΦϋΑϋά®ΟΗΜρΜ·―ßΫιΒΦΒΡΝ―Ϋβ“‘ΦΑ…φΦΑ pH ΚΆ»ΐκΡΙ»κΉΗ κΡ (GSH) ΒΡΈΔΜΖΨ≥ΈΔΖ¥”ΠΝ―Ϋβ±μ4ΓΘ
ΉήΫαΝΥPDCs…ηΦΤ÷–÷ς“Σ Ι”ΟΒΡΝ§Ϋ”Χεάύ–ΆΘ§Ν§Ϋ”Χε‘ΎΧεΡΎΒΡΈ»Ε®–‘“―ΒΟΒΫ÷Λ ΒΘ§¥”¥σΒΫ–ΓΒΡΥ≥–ρΈΣΘΚθΘΑΖ>Α±ΜυΦΉΥαθΞ>θΞ>ΧΦΥαθΞ[194]ΓΘ
ΫΓΩΒΉι÷·œΗΑϊΒΡpH÷ΒΈΣ7.4Θ§Εχ ΒΧε÷ΉΝωΉι÷·ΡΎΈΔΜΖΨ≥ΒΡpH÷ΒΒΆ”Ύ’ΐ≥ΘΉι÷·ΓΘ195Θ§196]ΓΘάϊ”Ο÷ΉΝωΈΔΜΖΨ≥ΒΡΈ¬ΚΆΥα–‘Θ§–μΕύpHΟτΗ–ΒΡΜ·―ßΦϋ±Μ”Ο”ΎΩΊ÷Τ÷ΈΝΤ“©Έο¥”Α–ΈΜPDCs÷–ΒΡ ΆΖ≈ΓΘ’β–©Φϋ‘ΎΧεΡΎ―≠ΜΖpHœ¬ «Έ»Ε®ΒΡΘ§ΒΪΩ…“‘‘ΎΥα–‘pHœ¬Υ°ΫβΘ§Αϋά®Υθ»©ΓΔ―«ΑΖΓΔκ¬ΚΆΗς÷÷Υαœλ”Π–‘κΡΤ§ΕΈΓΘGSHΩ…“‘Ν―Ϋβ“ΜœΒΝ–Ω…≤τ»κΫ”ΆΖΒΡΜ·―ßΦϋΘ§Αϋά®ΕΰΝρΦϋΓΔΝρθΞ/ΈχΜ·Έο/μΎΓΔΕΰΈχΜ·Έο/ΕΰμΎΜ·ΈοΓΔΕΰΟ·ΧζΚΆΫπ τΝρ¥ΦΫ”ΆΖΓΘ÷ΉΝω÷–ΒΡGSHΥ°ΤΫΚήΗΏΓΘ‘Ύ“©ΈοΒίΥΆΖΫΟφΘ§Ψ÷≤Ω‘ωΦ”ΒΡGSH≈®Ε»”–άϊ”ΎΩΊ÷Τ”––ß‘ΊΚ…ΒΡ ΆΖ≈ΓΘ…ΐΗΏΒΡGSH≈®Ε»“‘¥ΌΫχΫ”ΆΖΒΡΝ―Ϋβ±Μ”Ο”ΎΕύ÷÷÷ΉΝωΑ–œρ≤Ώ¬‘ΓΘΟΗ«–Ν§Ϋ”Έο «÷ΈΝΤ PDC ΒΡΝμ“Μ÷÷≥ΘΦϊ≤Ώ¬‘ΓΘΕΰκΡ Val-Ala Μρ Val-Cit «ΙψΖΚ Ι”ΟΒΡΟΗ«–Ν§Ϋ”ΈοΓΘ’βΝΫ÷÷ΕΰκΡ‘Ύ»ΥΧε―≠ΜΖ÷–ΕΦ±μœ÷≥ωΝΦΚΟΒΡΈ»Ε®–‘Θ§≤Δ«“Ϋω‘ΎΉι÷·ΒΑΑΉΟΗΜρτ»ΥαθΞΟΗ 1 ¥φ‘Ύœ¬≤≈Μα±Μ«–Ην[193]MMP ΙψΖΚ¥φ‘Ύ”ΎΧεΡΎΘ§“―≥…ΈΣ÷ΎΕύ―–ΨΩΒΡ÷ςΧβΓΘάΐ»γΘ§Tang Β»»Υ[197]Β»άϊ”ΟMMP-2ΟτΗ–κΡ(PVGLIG)ΉςΈΣ“©ΈοΉœ…Φ¥Φ”κκΡ≈ΦΝΣΈοΒΡΝ§Ϋ”–ρΝ–Θ§’β÷÷–¬ΒΡΗχ“©œΒΆ≥Ω…“‘ΩΊ÷ΤΉœ…Φ¥ΦΒΡ ΆΖ≈Θ§¥”Εχ‘ΎΫΚ÷ ΝωΜΦ’Ώ÷–ΜώΒΟΗϋΚΟΒΡ“÷÷Τ–ßΙϊΓΘ÷λΒ»―–ΨΩΝΥΕ‘MMP-2ΟτΗ–ΒΡΑΥκΡ(GPLGIAGQ)Θ§ΥϋΡή‘Ύ÷ΉΝω≤ΩΈΜ ΆΖ≈ΫΚ χΓΔsiRNAΒ»Θ§Φθ…Ό“©Έο‘ΎΖ«Α–Ήι÷·ΒΡ ΆΖ≈[198Θ§199¥ΥΆβΘ§ΜΙΩ…“‘Ά®ΙΐΫ”ΆΖΒΡ…ηΦΤά¥‘ωΦ”“©ΈοΗΚ‘Ί[200]ΓΘ
4 ¥Π”ΎΩΣΖΔΚΆΝΌ¥≤ ‘―ιΫΉΕΈΒΡ PDC
4.1 ΜθΈο
ΤάΦέΡΡ–©Κρ―Γ“©ΈοΩ…“‘ΉΣΜ·ΈΣPDCsΒΡ±ξΉΦ÷ς“ΣΑϋά®‘ω«Ω―≠ΜΖΈ»Ε®–‘ΚΆΧαΗΏ–߬ ΒΡ÷ΛΨίΓΘΩ…Ν―ΫβΝ§Ϋ”Ή” ΆΖ≈ΒΡΜθΈο±Ί–κ”–“ΜΗωΩ…––ΒΡΝ§Ϋ”ΒψΓΘ“ΜΑψά¥ΥΒΘ§Κρ―Γ“©ΈοΒΡIC50ΫœΒΆ50»ήΫβ–‘ΦΪ≤ν[218].PDC ÷– Ι”ΟΒΡΜθΈο ΨάΐΑϋά® DOX[219]ΓΔΉœ…Φ¥ΦΘ®PTXΘ©[220®C222], »αΚλΟΙΥΊ[223]ΚΆΦΣΈςΥϊ±θ[224]…φΦΑ’β–©÷ΉΝω÷ΈΝΤΦΝΒΡ÷ς“Σ’œΑ≠”κΥ°»ή“Κ÷–ΒΡ»ήΫβ–‘≤νΓΔΕ‘÷ΉΝωΉι÷·ΒΡ…χΆΗ–‘≤ν“‘ΦΑΕ‘ΫΓΩΒœΗΑϊΒΡ≤ΜΝΦ”Αœλ”–ΙΊΓΘΈΣΝΥΫβΨω’β–©Έ ΧβΘ§¥ΪΆ≥…œΜα Ι”ΟΗ≥–ΈΦΝά¥»ήΫβ≤Μ»ή–‘“©ΈοΘ§ΒΪ’β÷÷≤Ώ¬‘Μα¥χά¥Ϋχ“Μ≤ΫΒΡΈ ΧβΘ§άΐ»γΧμΦ”»ήΦΝΒΡΕΨ–‘Ής”ΟΓΘάΐ»γΘ§≥ΐΤδΥϊΗ±Ής”ΟΆβΘ§Kolliphor EL “―”Ο”ΎΓΑ»ήΫβΓ±Ήœ…Φ¥Φ (Taxol) ΉςΈΣ»ιΦΝΘ§≤Δ«“”κΙΐΟτΖ¥”ΠΚΆ÷ήΈß…ώΨ≠≤Γ±δ”–ΙΊ[225]ΫΪΩΙΑ©“©Έο”κ«ΉΥ°–‘κΡΫαΚœ≤ΜΫωΩ…“‘ΜΚΫβ’β÷÷»ήΫβΕ»Έ ΧβΘ§ΜΙΩ…“‘ΧαΙ©“ΐ»κΕνΆβΙΠΡήΒΡΜζΜα[226]ΓΘ
ΈΣΝΥΩΥΖΰΜ·ΝΤ“©ΈοDOX―≠ΜΖ ±Φδ”–œόΚΆΆ―Α–ΕΨ–‘ΒΡΨ÷œό–‘Θ§ChilkotiΒ»»Υ[209]. ΩΣΖΔΝΥ“Μ÷÷ΑΉΒΑΑΉΫαΚœκΡΉΚΚœΈοABD-DOXΓΘPDCsΩ… Ι÷ΉΝω≤ΩΈΜDOXΒΡΗΜΦ·≈®Ε»‘ωΦ”4±ΕΘ§≤ΔΩ…Ϋχ“Μ≤ΫΫΪDOXΒΡ“©–ß≥÷–χ ±Φδ―”≥Λ120±ΕΓΘΥ°»ή–‘≤νΚΆΕΨ–‘¥σœό÷ΤΝΥΙμΨ ΕΨΥΊΘ®PPTΘ©‘ΎΝΌ¥≤Α©÷Δ÷ΈΝΤ÷–ΒΡ”Π”ΟΓΘ÷ήΒ»[227]…ηΦΤΝΥ“Μ÷÷Μυ”ΎκΡΒΡPPTΉΚΚœΈοΘ§ΧαΗΏΝΥ“©ΈοΒΡ»ήΫβΕ»Θ§¥σ¥σΫΒΒΆΝΥ”ΈάκPPTΒΡ»ή―ΣΜν–‘Θ§œ‘÷χΧαΗΏΝΥΩΙΑ©ΝΤ–ßΓΘΦΣΈςΥϊ±θΒΡΩλΥΌ―ΣΫ§ΫΒΫβΓΔ»Ϊ…μΕΨ–‘ΚΆΡΆ“©–‘¥σ¥σœς»θΝΥΤδΉς”Ο≤Δœ‘÷χΫΒΒΆΝΥΤδΝΤ–ßΓΘΉνΫϋΩΣΖΔΒΡΗΡΫχ≥ΘΙφΜ·ΝΤΒΡΖΫΖ®÷°“Μ «Μυ”ΎPDCsΫΪΜ·ΝΤ“©Έο÷±Ϋ”Α–œρΑ©œΗΑϊΘ§¥σ¥σΧαΗΏΝΥΦΣΈςΥϊ±θΒΡ÷ΈΝΤ÷Η ΐΓΘ¥ΥΆβΘ§Ε‘”Ύ–μΕύ…ζΈο¥σΖ÷Ή”»γΒΑΑΉ÷ ΚΆΚΥΥα“‘ΦΑΡ…ΟΉ‘ΊΧεΘ®÷§÷ ΧεΓΔΫΚ χΚΆΡ…ΟΉΩ≈ΝΘΘ©ά¥ΥΒΘ§œΗΑϊΡΛ «÷Τ‘ΦΥϋΟ«ΩγΡΛ‘Υ δΒΡ÷Ί“Σ…ζάμΤΝ’œΓΘ“ρ¥ΥΘ§ΧαΗΏ“©ΈοΒΡœΗΑϊΡΛ¥©ΆΗ–߬ ÷ΝΙΊ÷Ί“ΣΓΘΫΪ ηΥ°–‘“©Έο”κ«ΉΥ°–‘κΡΉιΚœΩ…“‘ΗΡ±δΖ÷Ή”ΦΪ–‘Θ§”––ßΧαΗΏΡΛ¥©ΆΗΡήΝΠΓΘ–μΕύœΗΑϊ¥©ΆΗκΡ“―±Μ≥…ΙΠ”Ο”Ύ‘ω«ΩΒΑΑΉ÷ ΓΔΚΥΥαΚΆ–ΓΖ÷Ή”“©ΈοΒΡΙΠ–ßΓΘάΐ»γΘ§¥χ’ΐΒγΚ…ΒΡœΗΑϊ¥©ΆΗκΡ“―±Μ”Ο”ΎΝ§Ϋ”¥χΗΚΒγΚ…ΒΡΚΥΥα¥σΖ÷Ή”Θ§“‘ΙΙΫ®Ρ…ΟΉΒίΥΆœΒΆ≥ά¥÷ΈΝΤΗς÷÷“≈¥ΪΦ≤≤ΓΓΘ¥ΥάύΖΫΖ®Ω…“‘ΧαΗΏΚΥΥα¥σΖ÷Ή”‘ΎΧεΡΎΒΡΈ»Ε®–‘ΚΆΩγΡΛΡήΝΠΓΘ–μΕύ¥©ΡΛκΡ“―±Μ≥…ΙΠ”Ο”Ύ‘ω«ΩΒΑΑΉ÷ ΓΔΚΥΥαΚΆ–ΓΖ÷Ή”“©ΈοΒΡ“©–ßΓΘάΐ»γΘ§¥χ’ΐΒγΚ…ΒΡ¥©ΡΛκΡ“―±Μ”Ο”ΎΝ§Ϋ”¥χΗΚΒγΚ…ΒΡΚΥΥα¥σΖ÷Ή”Θ§ΙΙΫ®Ρ…ΟΉΒίΥΆœΒΆ≥ά¥÷ΈΝΤΗς÷÷“≈¥ΪΦ≤≤ΓΓΘ¥ΥάύΖΫΖ®Ω…“‘ΧαΗΏΚΥΥα¥σΖ÷Ή”‘ΎΧεΡΎΒΡΈ»Ε®–‘ΚΆΩγΡΛΡήΝΠΓΘ–μΕύ¥©ΡΛκΡ“―±Μ≥…ΙΠ”Ο”Ύ‘ω«ΩΒΑΑΉ÷ ΓΔΚΥΥαΚΆ–ΓΖ÷Ή”“©ΈοΒΡ“©–ßΓΘάΐ»γΘ§¥χ’ΐΒγΚ…ΒΡ¥©ΡΛκΡ“―±Μ”Ο”ΎΝ§Ϋ”¥χΗΚΒγΚ…ΒΡΚΥΥα¥σΖ÷Ή”Θ§ΙΙΫ®Ρ…ΟΉΒίΥΆœΒΆ≥ά¥÷ΈΝΤΗς÷÷“≈¥ΪΦ≤≤ΓΓΘ¥ΥάύΖΫΖ®Ω…“‘ΧαΗΏΚΥΥα¥σΖ÷Ή”‘ΎΧεΡΎΒΡΈ»Ε®–‘ΚΆΩγΡΛΡήΝΠΓΘ228Θ§229ΕΈ œΒ»…ηΦΤΝΥ“Μ÷÷ΨΏ”–«ΩΡΛ¥©ΆΗ–‘ΚΆΩΙ÷ΉΝωΜν–‘ΒΡ¥©ΆΗκΡ-PTX≈ΦΝΣΈοΘ§‘ω«ΩΝΥPTXΒΡΩΙ÷ΉΝωΜν–‘Θ§Ά§ ±ΩΥΖΰΝΥPTX»ήΫβ–‘≤νΚΆ÷ΉΝωΡΆ“©–‘ΒΡΈ Χβ[230]ΓΘ
4.2 ΝΌ¥≤ ‘―ι÷–ΒΡ PDC
ΨΓΙή PDC ΒΡΫαΙΙ”κΩΙΧε-“©Έο≈ΦΝΣΈο (ADC) œύΥΤΘ§ΒΪ’β–©ΙΠΡή–‘κΡ≈ΦΝΣΈο±» ADC ΨΏ”–Εύ÷÷”≈ ΤΘ§Αϋά®―Γ‘ώ–‘ΓΔΗϋ…νΒΡ÷ΉΝω…χΆΗΓΔΩλΥΌΆβ…χΚΆΗϋ¬ΐΒΡ…ω‘ύ«ε≥ΐΓΘPDC “©Έο Lu-dotatate177 “―ΜώΒΟΟάΙζ ≥ΤΖ“©ΤΖΦύΕΫΙήάμΨ÷ (FDA) ≈ζΉΦ”Ο”Ύ÷ΈΝΤΈΗ≥ΠΒά“»œΌ…ώΨ≠ΡΎΖ÷ΟΎ÷ΉΝω[231]ΓΘ»ΜΕχΘ§ΜΙ”–ΗϋΕύΒΡPDCs¥Π”ΎΩΣΖΔΫΉΕΈΘ§άΐ»γTH1902Θ§“Μ÷÷ΉΑ‘ΊΕύΈςΥϊ»ϋΒΡPDCΘ§”Ο”Ύ÷ΈΝΤ»ΐ“θ–‘»ιœΌΑ©ΚΆ¬―≥≤Α©ΓΘTH1904 «“Μ÷÷ΉΑ‘ΊDOXΒΡPDCΘ§Ω…”Ο”Ύ÷ΈΝΤ¬―≥≤Α©ΓΘ’βΝΫ÷÷PDCΕΦΑ–œρΑ©÷Δ÷–sortilin 1 (SORT1) ήΧεΒΡΙΐΕ»±μ¥οΘ§Αϋά®»ΐ“θ–‘»ιœΌΑ©ΓΔ¬―≥≤Α©ΓΔΖΈΑ©ΓΔΫα÷±≥ΠΑ©ΓΔΤΛΖτΑ©ΚΆ“»œΌΑ©[232]Νμ“Μ÷÷Μυ”Ύ≥ΤΈΣΉ‘––≥ΒκΡΒΡΉιΚœΈοΒΡ PDC “©Έο“≤Ζ«≥Θ ήΜΕ”≠ΓΘΉ‘––≥ΒκΡΆ®≥Θ≥ΛΕ»ΈΣ 9-20 ΗωΑ±ΜυΥαΘ§–ρΝ–÷–”–»ΐΗωΑκκΉΑ±Υα≤–ΜυΓΘ’β–©ΑκκΉΑ±Υα≤–Μυ”κ–ΓΖ÷Ή”Ϋ”ΆΖΖ¥”ΠΘ§ΫΪκΡœό÷ΤΈΣΗ’–‘ΙΙœσΓΘΉ‘––≥ΒΕΨΥΊΫαΚœΈοΩ…”ΟΉς“©ΈοΖ÷Ή”ΒΡΉΣ‘ΥΒΑΑΉΓΘ“©ΈοΗΫΉ≈‘ΎΉ‘––≥ΒκΡ…œΘ§“‘»Ζ±ΘΙΙœσ≤Μ ήΉηΑ≠[233]. Εύ÷÷Ή‘––≥ΒΝΤΖ®“―Ϋχ»κ’κΕ‘ΧΊΕ®÷ΉΝωΒΡΝΌ¥≤ ‘―ι[234]±μ 5ΓΘ
ΉήΫαΝΥΟάΙζFDAΆχ’Ψ…œΖΔ±μΒΡ…φΦΑPDCsΒΡΝΌ¥≤―–ΨΩΫχ’Ι±®ΗφΘ®https://www.fda.gov/Θ©ΓΘ
5 Ϋα¬έΚΆΈ¥ά¥’ΙΆϊ
±ΨΉέ ω«ΩΒςΝΥPDCΉςΈΣΙΙΫ®ΗΡΫχΒΡ÷«Ρή“©Έο δΥΆœΒΆ≥ΒΡΕύ―υΜ·ΙΛΨΏΒΡ«±ΝΠΘ§Αϋά®Τδ“Ή”ΎΚœ≥…ΓΔΙΠΡήΕύ±δΓΔ“©ΈοΗΚ‘ΊΙψΖΚΓΔ–ΈΧ§Εύ―υ–‘ΚΆΩ…Βς–‘ΓΘ”κADCœύ±»Θ§”…”ΎκΡ«χΚΆΝ§Ϋ”Τ§ΕΈΒΡ≥ω…ΪΕύΙΠΡή…ηΦΤΘ§PDCΨΏ”–ΗϋΕύΒΡΜζΜαΚΆΩ…Ρή–‘ΓΘΨΓΙήΨΏ”–’β–©”≈ ΤΘ§PDC‘ΎΫχ»κΝΌ¥≤ ±»‘ΟφΝΌ–μΕύΧτ’ΫΓΘ÷ς“ΣΧτ’Ϋ÷°“Μ «ΤδΈ»Ε®–‘≤νΚΆ…ω‘ύ«ε≥ΐΥΌΕ»ΩλΓΘPDC±Ί–κ‘Ύ―≠ΜΖ÷–±Θ≥÷Έ»Ε®Θ§“‘Ζά÷ΙœΗΑϊΕΨ–‘”––ß‘ΊΚ…ΒΡ‘Λœ» ΆΖ≈ΚΆΥφ÷°Εχά¥Βѻڅ챩¬ΕΓΘΈΣΝΥ÷ΈΝΤ ΒΧεΝωΘ§Ά®≥Θ–η“ΣΗϋ≥ΛΒΡPDC―≠ΜΖ ±ΦδΘ§“‘±ψ”–ΉψΙΜΒΡ ±ΦδΫΪPDCsΒίΥΆ÷ΝΕώ–‘Ήι÷·≤Δ¥©ΆΗΓΘNPΖβΉΑΒΡPDCsΚΆPEGΜ·¥ζ±μΝΥ―”≥ΛPDCs‘ΎΧεΡΎΑκΥΞΤΎΒΡ”–Έϋ“ΐΝΠΒΡ≤Ώ¬‘ΓΘ≥ΐΝΥ¥ΪΆ≥ΒΡΦΝ–ΆΚΆΗχ“©ΆΨΨΕΘ®»γΨ≤¬ωΜρΦΓ»βΉΔ…δΘ©ΆβΘ§ΜΙ”ΠΧΫΥς–¬ΒΡΗχ“©≤Ώ¬‘Θ§»γ±««ΜΚΆ…ύœ¬PDCΗχ“©Θ§“‘±ήΟβPDCΫΒΫβΓΘ‘ΎΩΙΨζΝλ”ρΘ§Υδ»Μ‘Ύ…ηΦΤΚΆ”≈Μ·Ηϋ”––ßΒΡΩΙΨζκΡΜρ“©ΈοΖΫΟφ»ΓΒΟΝΥ‘Ϋά¥‘ΫΕύΒΡ≥…ΨΆΘ§ΒΪ…ηΦΤPDCs“‘―Γ‘ώ–‘ΒΊΑ–œρΧΊΕ®÷÷άύΒΡœΗΨζΕχ≤Μ «ΙψΤΉœΗΨζ»‘»Μ «“ΜΗωΙΊΦϋΧτ’ΫΓΘœό÷ΤPDC―–ΨΩΒΡΝμ“ΜœνΙΊΦϋΦΦ θ «»±ΖΠ”Ο”ΎPDC≤β ‘ΒΡΉ®”–“©¥ζΕ·ΝΠ―ßΖΫΖ®ΓΘΩΣΖΔΗΏ–ßΒΡPDCs–η“Σ…ηΦΤ≥ωΨΏ”–Εύ÷÷ΙΠΡήΒΡΙΠΡήκΡΘ§άϊ”Ο»ΥΙΛ÷«ΡήœΒΆ≥ΒΡΚΘΝΩ ΐΨίΖ÷ΈωΡήΝΠΘ§ΧΫΥςΉιΉΑ–ΈΟ≤”κΑ±ΜυΥαάύ–ΆΚΆ–ρΝ–÷°ΦδΒΡΙΊœΒΘ§ Βœ÷PDCsΉ‘ΉιΉΑΫαΙΙΚΆ“©Έο ΆΖ≈ΒΡΨΪΉΦΒςΩΊΓΘ¥ΌΫχPDCsΒΡΩΤ―ßΉΣΜ·Θ§ΜΙ–η“Σ‘ΎΚœ≥…ΦΦ θ…œΉω≥ω≈§ΝΠΘ§Αϋά®ΩΊ÷Τ‘”÷ ΓΔ±Θ÷Λ“©ΈοΒΡΈ»Ε®–‘Β»ΓΘ Βœ÷PDCΉ‘ΉιΉΑΫαΙΙΚΆ“©Έο ΆΖ≈ΒΡΨΪΉΦΒςΩΊΓΘ¥ΌΫχPDCsΒΡΩΤ―ßΉΣΜ·Θ§ΜΙ–η“Σ‘ΎΚœ≥…ΦΦ θΖΫΟφœ¬ΙΠΖρΘ§Αϋά®ΩΊ÷Τ‘”÷ ΓΔ±Θ÷Λ“©ΈοΒΡΈ»Ε®–‘Β»ΓΘ Βœ÷PDCΉ‘ΉιΉΑΫαΙΙΚΆ“©Έο ΆΖ≈ΒΡΨΪΉΦΒςΩΊΓΘ¥ΌΫχPDCsΒΡΩΤ―ßΉΣΜ·Θ§ΜΙ–η“Σ‘ΎΚœ≥…ΦΦ θΖΫΟφœ¬ΙΠΖρΘ§Αϋά®ΩΊ÷Τ‘”÷ ΓΔ±Θ÷Λ“©ΈοΒΡΈ»Ε®–‘Β»ΓΘ
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷ΨΓΘ