
еЊвЊЃКЗжСбФкКЌыФЭЈЙ§МєНгЗДгІЪЕЯжЖрыФЦЌЖЮЕФСЌНгЃЌ ОпгаИпаЇЧвЮоКлЕФЬиЕуЃЌ ЪмЕНСЫЙуЗКЙизЂ. БОЮФЛљгкЗжСбФкКЌыФЕФНсЙЙЬиеїгыМєНгЗДгІЙ§ГЬЃЌ НсКЯНќФъРДЙигкЗжСбФкКЌыФадФмгХЛЏКЭгІгУбаОПНјеЙНјааСЫзлКЯЦРЪіЃЌ НвЪОЦфзїЮЊвЛжжШеНЅГЩЪьЕФЕААзжЪЙЄГЬЛЏММЪѕдкЕААзжЪЛЏбЇКЯГЩСьгђЕФЧАОАЃЌ ВЂМђвЊЗжЮіСЫФПЧАЗжСбФкКЌыФЙЄОпУцСйЕФЮЪЬтгыЬєеНЃЌ ВЂЖдПЩФмЕФНтОіЗНАИНјааСЫеЙЭћ.
ЗвыКѓаоЪЮМЋДѓЕиЗсИЛСЫЕААзжЪЕФНсЙЙгыЙІФмЕФЖрбљадЃл1Ѓн. етаЉаоЪЮЕААздквЉЮяПЊЗЂЃл2ЃнЁЂ ЕААзжЪзщбЇЃл3ЃнвдМАЕААзжЪНсЙЙ-ЙІФмЙиЯЕНтЮіЃл4~8ЃнЕШЗНУцОпгаживЊЕФгІгУ. ФПЧАЃЌ ЗвыКѓаоЪЮЕААзПЩвдЭЈЙ§ЬьШЛЬсШЁКЭЩњЮяБэДяЕШЗНЗЈЛёШЁ. ЕЋетаЉЗНЗЈШдДцдквЛЯЕСаЮЪЬтЃЌ Р§ШчЃЌ ЬьШЛЬсШЁФбвдЛёЕУОљвЛаоЪЮЕФЕААзбљЦЗЃЌ ЖјЩњЮяБэДяЛёШЁКЌЖржжЗвыКѓаоЪЮЕФЕААзбљЦЗвРШЛДцдкРЇФбЕШ. ЕААзжЪЛЏбЇКЯГЩММЪѕЪЧЛёЕУЗвыКѓаоЪЮЕААзЕФвЛжжаТаЫММЪѕЃЌ ИУММЪѕЛљгкЙЬЯрЖрыФКЯГЩММЪѕКЭЖрыФЦЌЖЮСЌНгММЪѕ. ЙЬЯрЖрыФКЯГЩММЪѕЃл9ЃнПЩвджБНгНЋИїжжЗЧЬьШЛАБЛљЫсМАДјгааоЪЮЛљЭХЕФАБЛљЫсЗНБуЕив§ШыЖрыФСДжаЬиЖЈЕФЮЛЕу. ОЁЙмЙЬЯрЖрыФКЯГЩММЪѕЕФКЯГЩФмСІЩЯЯовЛжБдкВЛЖЯЬсИпЃл10ЃнЃЌ ЕЋДѓВПЗжЕААзШдШЛФбвдЭЈЙ§ЙЬЯрКЯГЩвЛДЮадЛёШЁ. вђДЫЃЌ ЪЙгУЖрыФЦЌЖЮСЌНгММЪѕФмЙЛНЋКЌЬиЖЈаоЪЮЕФЖрыФЦЌЖЮНјааСЌНгЃЌ МЋДѓЕиРЉеЙСЫЕААзбљЦЗЕФЛёШЁЗЖЮЇЃЈЭМ1ЃЉ. ЭЈЙ§ЖрыФЦЌЖЮСЌНгЃЌ баОПепУЧвбОКЯГЩСЫДјгагЋЙтБъМЧЃл11ЃнЃЌ C13КЭN15ЕШЭЌЮЛЫиБъМЧЃл12ЃнЃЌ СзЫсЛЏЁЂ МзЛљЛЏМАЗКЫиЛЏЕШЗвыКѓаоЪЮЃл13ЃнЃЌ вдМАКЌЗЧЬьШЛАБЛљЫсЃл14ЃнЕШЙІФмЕФЕААзжЪ.
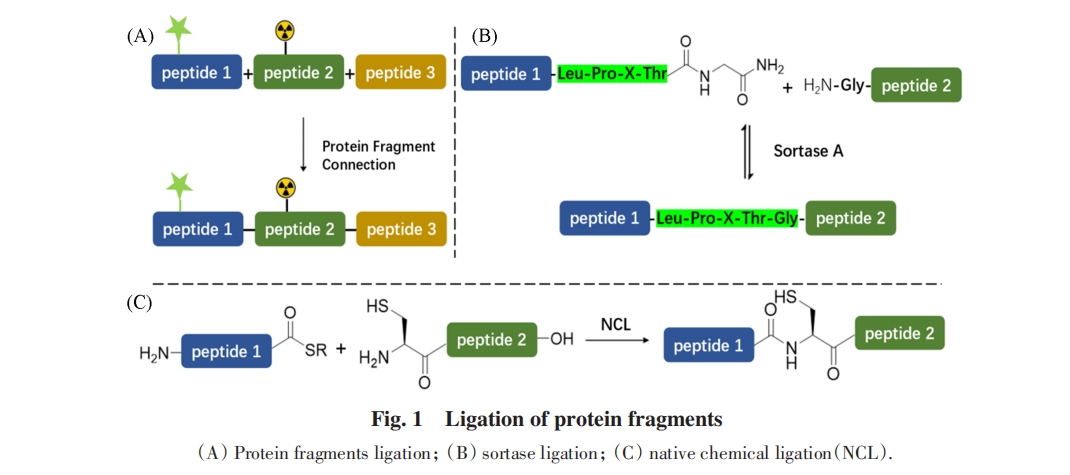
ФПЧАЃЌ гІгУзюЙуЗКЕФЛЏбЇСЌНгММЪѕЪЧздШЛЛЏбЇСЌНгЃЈNCLЃЉ. ИУММЪѕгЩжЅМгИчДѓбЇЕФKentЕШЃл15Ѓнгк1994ФъЬсГіЃЌ ЭЈЙ§вЛИіЖрыФЦЌЖЮЕФC-ЖЫСђѕЅгыСэвЛИіN-ЖЫCysЖрыФЦЌЖЮЕФбЁдёадЛЏбЇЗДгІЃЌ дкСЌНгЮЛЕуДІаЮГЩЬьШЛѕЃАЗМќ. дчЦкЃЌ C-ЖЫСђѕЅЕФКЯГЩЪмЕНЮШЖЈадВюЕФРЇШХЃЛНќФъРДЭЈЙ§ѕЃыТбѕЛЏЕФC-ЖЫСђѕЅдЮЛКЯГЩМАСЌНгВпТдМЋДѓЕиДйНјСЫетвЛЗНЗЈЕФгІгУЃл16~20Ѓн. ШЛЖјЃЌ гЩгкNCLЗДгІвРППЫцЛњХізВЃЌ ЦфСЌНгаЇТЪЪмЕзЮяХЈЖШЕФжЦдМЃЌ NCLЭЈГЃашвЊmmol/LМЖБ№ЕФЕзЮяХЈЖШ. ЕБЕзЮяХЈЖШЕЭгк1 mmol/LЪБСЌНгаЇТЪЕЭЃЌ ЪЙЕУNCLдкжЌЕААзЁЂ ФЄЕААзЕШФбШмЕААзЦЌЖЮСЌНгЪБВњЩњРЇФб. ЮЊдіМгNCLдкЕЭХЈЖШЬѕМўЯТЕФЗДгІаЇТЪЃЌ баОПепУЧЬсГіСЫЖржжНтОіЗНАИЃЌ жївЊгаЃКЯђЗДгІЬхЯЕМгШыгаЛњШмМСЛђепШЅЙИМСвддіМгЦЌЖЮШмНтЖШЃл21ЃнЃЛдкжїСДЩЯв§ШыПЩвЦГ§ЕФдіШмаоЪЮвдДђЦЦЧтМќЭјТчЁЂ ЯїШѕОлМЏЧуЯђЃл22ЃЌ23ЃнЃЛЭЈЙ§DNA-DNAЁЂ ЕААз-ЕААз/аЁЗжзгЕШЧПЯрЛЅзїгУФЃАхИЈжњЦЌЖЮСЌНгЃЌ ЪЙЦЌЖЮСЌНгдкНќЫЦЗжзгФкЗДгІЕФЬѕМўЯТНјааЃл24ЃнЕШ. ДЫЭтЃЌ ЪЙгУЛюадИќИпЕФЮјѕЅДњЬцСђѕЅзїЮЊѕЃЛљЙЉЬхЕФЗНАИвВФмЪЙЕУЗДгІдкИќЕЭХЈЖШЯТНјааЃл25Ѓн. ЫфШЛетаЉВпТдгажњгкКЯГЩОпгаЬєеНадЕФФПБъЃЌ ЕЋвВДцдкдіМгАВзАКЭвЦГ§БъЧЉЛђФЃАхЕФЖюЭтВйзїЁЂ БъЧЉзюМбАВзАЮЛжУФбвддЄВтМАЮјѕЅЕФЮШЖЈадНЯЕЭЕШЙЬгаЮЪЬтЃЌ ЕМжТетаЉЗНЗЈдкЮШНЁадКЭВйзїБуНнадЗНУцДцдкВЛзу.
СэвЛЗНУцЃЌ ЛЏбЇУИЗЈвђЦфФмЙЛдкЕЭХЈЖШЯТИпаЇЗДгІЃЌ дкЕААзжЪКЯГЩжавВБЛЙуЗКгІгУЃл26~28Ѓн. ЕЋетРрСЌНгЗДгІЭЈГЃашвЊдкЕзЮяжав§ШыУИЬивьадЪЖБ№ађСаЃЌ етаЉЪЖБ№ађСадкЗДгІКѓЭљЭљЛсБЃСєдкВњЮяЕААзЕФађСажаЃЌ СєЯТађСаЁААЬКлЁБЃЌ ШчSortase AУИЕФВаСєађСаLPXTGЃл29Ѓн. ЫфШЛПЩвдЭЈЙ§бЁдёФПБъЕААзжЪЙЬгаДцдкЕФУИЕФЪЖБ№ађСазїЮЊСЌНгЮЛЕуЃЌ ЕЋетаЉЧщПіЪЧМЋЦфХМШЛЕФЃЛЭЈЙ§ИпЭЈСПЩИбЁММЪѕЖдSortase AНјааађСаНјЛЏЃл30~33ЃнЃЌ вдМАЪЙгУдЄЯШжЦБИГЩСђѕЅЕФЕААзЦЌЖЮзїЮЊСЌНгЕзЮяЃл34ЃнЕШВпТдПЩвдМѕЩйSortase AУИЕФађСавРРЕадЃЌ ВЂМѕЩйЗДгІЕФПЩФцадЃл35ЃнЃЌ ЕЋвЊЪЕЯжЭъШЋЮоКлЕФЦЌЖЮСЌНгШдШЛРЇФб.
вђДЫЃЌ бАеввЛжжФмЙЛИпаЇСЌНгЕААзЦЌЖЮЧвСЌНгКѓЮоКлЕФЕААзСЌНгЙЄОпЃЌ ГЩЮЊбаОПепЙизЂЕФЗНЯђ. NCLЗДгІКЭSortase AУИСЌНгЙ§ГЬжаЖМОРњСЫСђѕЅКЭѕЃЛљS-NЧЈвЦЙ§ГЬЃЛдкЩњУќНјЛЏЙ§ГЬжаЃЌ ЗКЫиЛЏУИМЖСЊЗДгІЕШЙ§ГЬжавВЙлВьЕНСђѕЅжаМфЬхКЭѕЃЛљS-NЧЈвЦЙ§ГЬЃл36ЃнЃЌ гЩДЫПЩМћетЪЧЩњГЩаТЕФыФМќЕФвЛжжгааЇЭООЖ. ЪТЪЕЩЯЃЌ ШЫУЧвбОЗЂЯжСЫвЛРрФмЙЛЭЈЙ§СђѕЅжаМфЬхКЭѕЃЛљЧЈвЦТЗОЖЪЕЯжЖдЕААзжЪЦЌЖЮЕФИпаЇЮоКлСЌНгЕФЕААзЃКФкКЌыФЃЈInteinЃЉ. ФкКЌыФПЩвдЗжЮЊЫГЪНФкКЌыФЃЈCis-inteinЃЉгыЗжСбФкКЌыФЃЈSplit inteinЃЉСНДѓРр. ЦфжаЃЌ ЗжСбФкКЌыФПЩвдгыашвЊНјааСЌНгЕФЕААзЦЌЖЮЗжБ№ЯрСЌЃЌ вбОдкДѓСПЕААзжЪСЌНггыКЯГЩжаеЙЯжГігІгУМлжЕЃЌ ЪмЕНСЫбаОПепЕФЙуЗКЙизЂ. БОЮФжївЊНщЩмЗжСбФкКЌыФзїЮЊвЛжжЕААзжЪКЯГЩЙЄОпЃЌ НсКЯЗжСбФкКЌыФдкЕААзжЪКЯГЩаоЪЮСьгђЕФгІгУАИР§ЃЌ ЗжЮіСЫЗжСбФкКЌыФЯрБШЦфЫќЗНЗЈЕФгХЪЦЃЌ змНсЗжСбФкКЌыФЕФЗЂеЙЯжзДЃЌ ВЂеЙЭћСЫЦфЮДРДЕФЗЂеЙЧАОА.
1 ЗжСбФкКЌыФЕФЗЂЯж
1.1ЁЁФкКЌыФЕФЗЂЯж
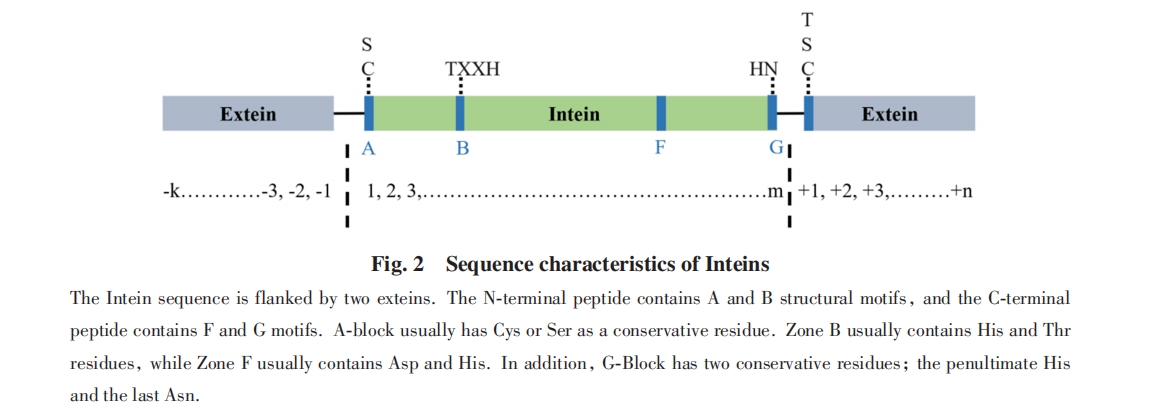
1990ФъЃЌ StevensЕШЗЂЯжФ№ОЦНЭФИЃЈSaccharomy cescerevisiaeЃЉTFP1ЛљвђЕФЗвыВњЮяЛсдкЗвыКѓБЛЧаИюГЩ2ИіЕААзЃЌ жабыЕФађСаМєЧаВњЩњЗжзгСПЮЊ50000ЕФЕААзЃЌ СНЖЫЕФЦЌЖЮЦДНгаЮГЩЗжзгСПЮЊ69000ЕФЕААз. ДЫКѓбаОПепдкИќЖрЮяжжжаЗЂЯжСЫРрЫЦЕФЯжЯѓЃл37ЃЌ38Ѓн. КѓајбаОПЗЂЯжЃЌ етжжВхШыЫожїЕААзађСаЕФЕААзПЩвдДпЛЏздЩэДгЫожїЕААзжаЖЯСбЃЌ ЭЌЪБЪЙСНВрЕФЫожїЕААзЦЌЖЮЭЈЙ§ыФМќСЌНгЦ№РДаЮГЩГЩЪьЕААзЃл39Ѓн. етжжОпгаМєНгСНВрЕААзЦЌЖЮФмСІЕФЕААзБЛУќУћЮЊФкКЌыФЃЈInteinЃЉЃЌ ЖјСНЖЫБЛМєНгЕФЕААзЦЌЖЮБЛУќУћЮЊЭтЯдыФЃЈExteinЃЉЃЈЭМ2ЃЉ.
1.2ЁЁЗжСбФкКЌыФЕФЗЂЯж
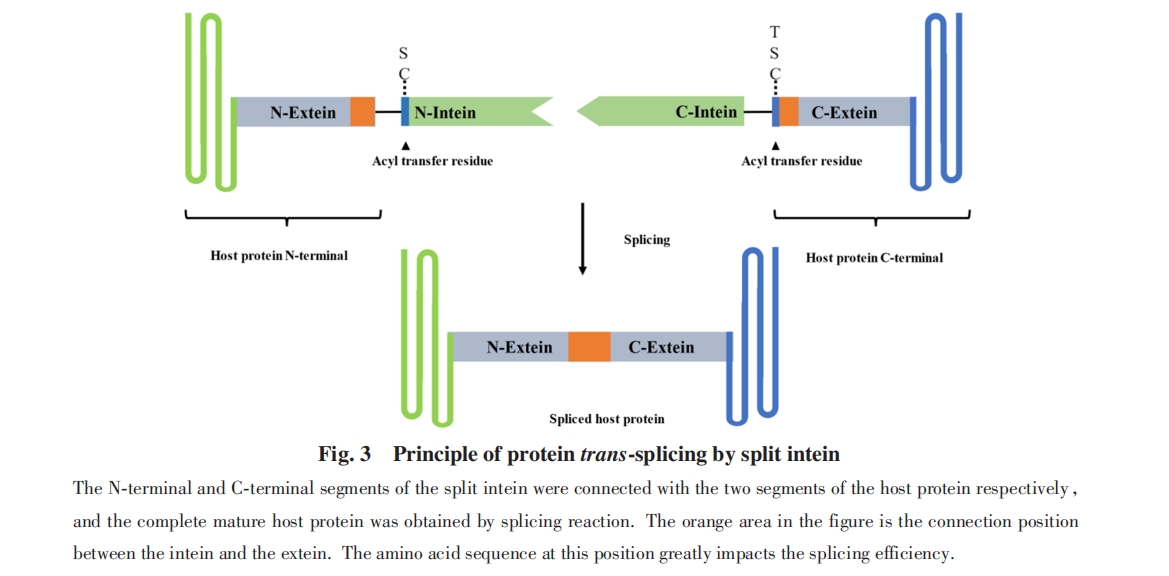
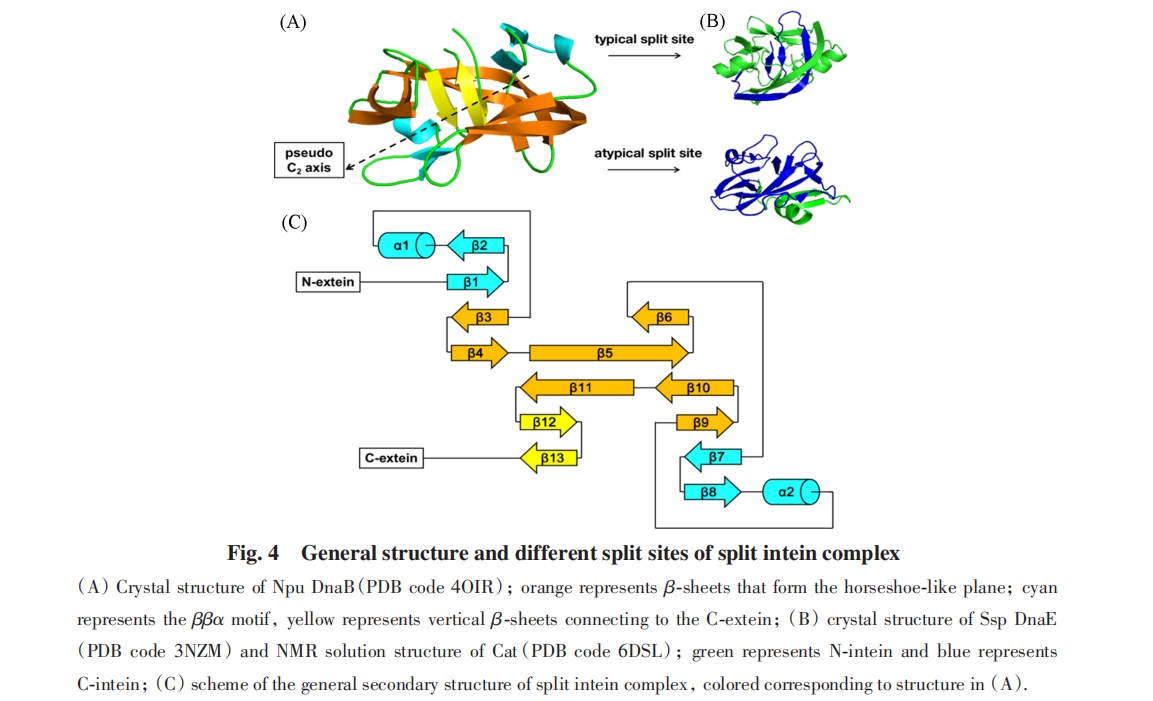
ЗжСбФкКЌыФЪЧЗжСбЮЊ2ИіЖРСЂДцдкЕФЧАЬхЦЌЖЮЕФФкКЌыФЃЌ УПИіЦЌЖЮЗжБ№гы1ИіЭтЯдыФШкКЯЃЈЭМ3ЃЉ. 2ИіЧАЬхЦЌЖЮЗЧЙВМлНсКЯВЂзщзАГЩЕфаЭЕФФкКЌыФНсЙЙЃЌ ДгЖјЛжИДЗжСбФкКЌыФЕФМєНгЛюадЃЌ ДпЛЏСНВрЕФЭтЯдыФЗЂЩњЕААзжЪЗДЪНМєНгЃЈPTSЃЉ. ЦфПЩвдРДдДгкЬьШЛСЌајФкКЌыФЕФШЫЙЄЗжСбЃЌ вВдкздШЛНјЛЏжаДцдк.
1969ФъЃЌ AnfinsenЕШЃл40ЃнЗЂЯждкЬиЖЈЕФЬѕМўЯТЃЌ ЗжСбГЩ2ИіЦЌЖЮЕФУИФмЙЛЗЧЙВМлНсКЯВЂЛжИДЭъећУИЕФДпЛЏЙІФм. 1998ФъЃЌ PerlerЕШЃл41ЃнНЋетвЛЫМЯыдЫгУЕНФкКЌыФЕФЩшМЦИФдьжа. ЫћУЧНЋPsp Pol-1ФкКЌыФжаЕФвЛИіыФМќЧаЖЯЃЌ НЋЦфЗжСбЮЊN-ЖЫЦЌЖЮЃЈIntNЃЉКЭC-ЖЫЦЌЖЮЃЈIntCЃЉЃЌ НЋ2ИіЦЌЖЮЗжБ№БэДяДПЛЏКѓдкЬхЭтЙВИДадЃЌ НсЙћБэУї2ИіЦЌЖЮЭЈЙ§ЗЧЙВМлзїгУаЮГЩИДКЯЮяВЂЛжИДМєНгЛюад.
ЭЌФъЃЌ PaulusЕШЃл42ЃнНЋMtu RecAФкКЌыФЗжСбЮЊN-ЖЫКЭC-ЖЫ2ИіЦЌЖЮЃЌ ВЂИїздШкКЯЕНЯргІЕФЭтЯдыФЩЯЃЌ ЗжБ№БэДяДПЛЏКѓЙВИДадЃЌ 2ИіЦЌЖЮНсКЯаЮГЩСЫОпгаМєНгЙІФмЕФИДКЯЮя. ЬНОПСЫМєНгЗДгІЙ§ГЬЖдбѕЛЏЛЙдЛЗОГКЭpHжЕЕФвРРЕадЃЌ ВЂжИГіетжжМєНгЯЕЭГЪЧвЛжжгаЧБСІЕФЕААзжЪСЌНгЙЄОп. ДЫКѓЃЌ баОПепЖдЬьШЛСЌајФкКЌыФНјааСЫДѓСПЕФШЫЙЄЖЯСбГЂЪдЃЌ ЗЂеЙГіЖржжШЫЙЄЗжСбФкКЌыФЃЈБэ1ЃЉ.
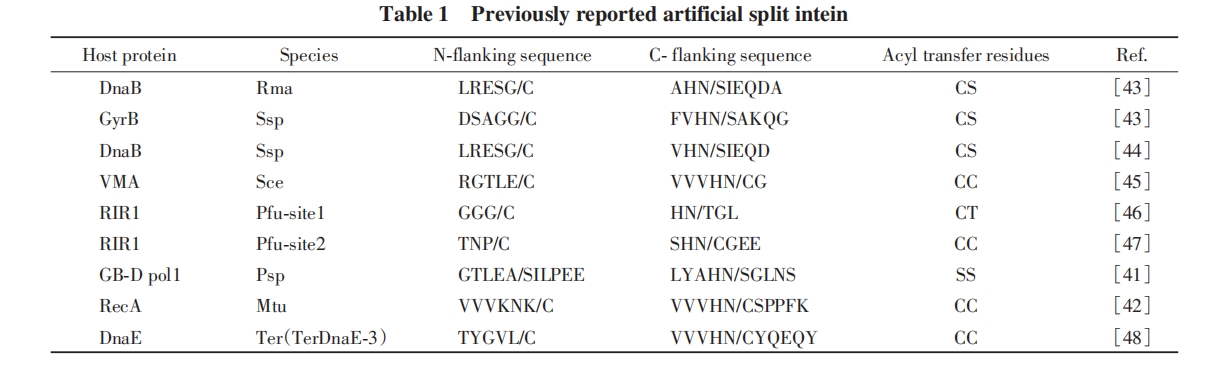
1998ФъЃЌ LiuЕШЃл49ЃндкSsp DnaEЃЈРЖдхЕФDNAОлКЯУИЂѓЕФДпЛЏбЧЛљAЃЉЕААзжаМјЖЈГіЪзИіЬьШЛЕФЗжСбФкКЌыФ. DnaEЕФN-ЖЫКЭC-ЖЫВПЗжгЩ2ИіЕЅЖРЕФЛљвђDnaE-NКЭDnaE-CЗжБ№БрТы. дкДѓГІИЫОњжаВтЪдЪБЃЌ 2ИіЗжСбФкКЌыФЦЌЖЮЭЈЙ§ЗЧЙВМлзїгУНсКЯаЮГЩФкКЌыФИДКЯЮя. ИУИДКЯЮяВЛНіОпгаРрЫЦФкКЌыФЕФађСаЬиеїЃЌ ЖјЧвБэЯжГіЕААзжЪЗДЪНМєНгЛюадЃЌ МєНгКѓЕУЕНЭъећЕФDNAОлКЯУИЂѓбЧЛљA. етвЛЗЂЯжБэУїЃЌ ЗжСбФкКЌыФЪЧЬьШЛДцдкЕФ. гыШЫЙЄЗжСбФкКЌыФЯрБШЃЌ ЬьШЛЗжСбФкКЌыФОпгаПЩдкЗЧБфадЬѕМўЯТзщзАМАМєНгаЇТЪИќИпЕШгХЪЦ.
2009ФъЃЌ MootzЕШЃл50ЃнЗЂЯжСЫСэвЛжжаТЕФЬьШЛЗжСбФкКЌыФNpu DnaE. Npu DnaEОпгаМЋИпЕФМєНгЗДгІЛюадЃЌ дк37 ЁцЯТЗДЪНМєНгЗДгІЕФАыЫЅЦкЃЈМДЗДгІЮяЕФХЈЖШЯћКФвЛАыЪБЫљашЕФЪБМфt1/2ЃЌ sЃЉНіЮЊ63 sЃЌ МєНгЗДгІЫйТЪГЃЪ§kЃЈs-1ЃЉ>3.5ЁС10-3 s-1. ДЫКѓЃЌ баОПепУЧЭЈЙ§ЛљвђЪ§ОнБШЖдКЭЪЕбщбщжЄЕШЪжЖЮЃЌ ЗЂЯжСЫИќЖрРДздВЛЭЌЮяжжЕФЬьШЛЗжСбФкКЌыФЃЈБэ2ЃЉЃЌ НјвЛВНРЉДѓСЫЗжСбФкКЌыФзїЮЊЕААзжЪСЌНгЙЄОпЕФПЩааадгыгІгУЧАОА.
1.3ЁЁЗжСбФкКЌыФЕФЗжРр
ЗжСбФкКЌыФПЩвдИљОнЦфМФжїЛљвђЙщРрЮЊОпгавХДЋЭЌдДЙиЯЕЕФМвзхРр. ДЫЭтЃЌ ВЛЭЌЕФЗжСбФкКЌыФдкЗДгІЙ§ГЬжажаМфЬхДцдкВювьЃЌ вВПЩвдОнДЫЖдЗжСбФкКЌыФНјааРрБ№ЛЎЗж. дкНсЙЙЩЯЃЌ ФкКЌыФвЛАуГЪЯжЖрИіІТЦЌВуЕФНсЙЙЃЌ ЗжСбЕФЮЛЕувЛАудкІТЦЌВужЎМфЕФLoopЧјЃЌ ДгВЛЭЌЧјгђЖЯСбЕФФкКЌыФОпгаВЛЭЌЕФЬиадЃЌ ФмЙЛЗсИЛЗжСбФкКЌыФЕФгІгУГЁОАЃЌ вђДЫбаОПепвВИљОнЗжСбФкКЌыФЕФЗжСбЮЛЕуЖдЦфНјааЙщФЩ.
дМ70%ЕФФкКЌыФЕФЫожїЕААзгыDNAИДжЦКЭDNAаоИДгаЙиЃл60ЃнЃЌ ВЂЧвдкНјЛЏЗЂеЙЙ§ГЬжаБэЯжГіЛљвђЫЎЦНЧЈвЦЕФЬиеїЃл61Ѓн. вђДЫЗжСбФкКЌыФПЩвдАДееЦфРДдДЕФЫожїЛљвђНјааЗжРрЃЌ РДдДгкЭЌРрЛљвђЕФЗжСбФкКЌыФЕФађСаЭЌдДадНЯИпЃЌ ВЂЧвОпгаРрЫЦЕФЬиеї. РДдДгкВЛЭЌЮяжжЕФDna EЗжСбФкКЌыФЖМФмдкЯрЭЌЕФЭтЯдыФађСаНјааЧаИюЃЌ ВЂЧвбаОПвВЗЂЯжЦфМфДцдкНЛВцЗДгІадЃЌ ШчNpu DnaEЕФIntNЦЌЖЮКЭSsp DnaEЕФIntCЦЌЖЮвВПЩвдЗЧЙВМлНсКЯВЂаЮГЩОпгаМєНгЗДгІЛюадЕФИДКЯЬхЃл62Ѓн. баОПепЖдгкдДгкDnaEЃЌ GyrBКЭDnaBЕШМвзхЕФЗжСбФкКЌыФвбОНјааСЫЯЕЭГЕФбаОПЃЌ ВЂЧвПЊЗЂСЫЖржжгІгУ.
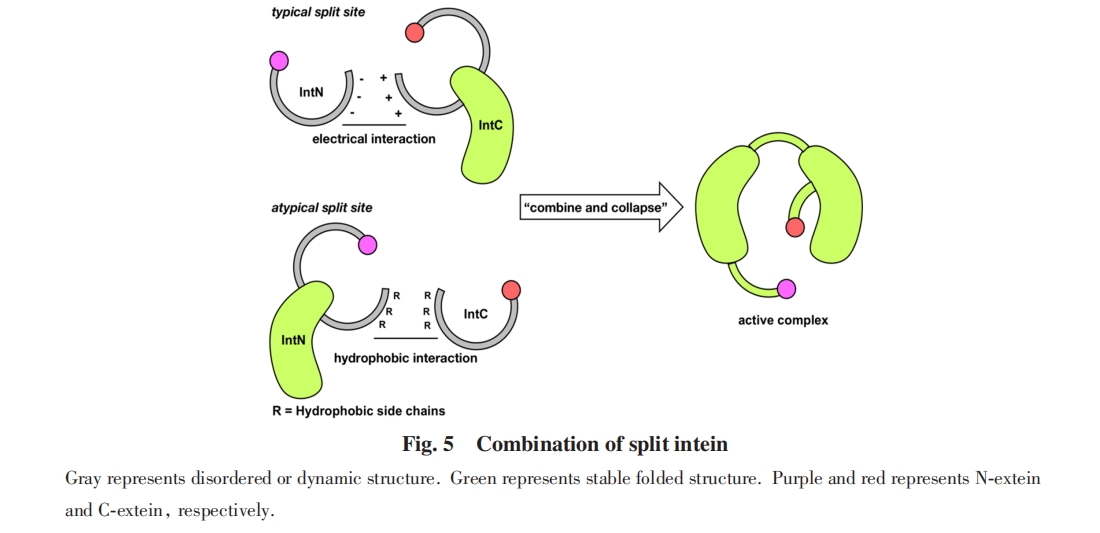
ЗжСбФкКЌыФДпЛЏЕФМєНгЗДгІЙ§ГЬЩцМАЖрВНЙиМќЕФѕЃЛљзЊвЦЗДгІЃЌ ЙиМќАБЛљЫсВаЛљдкетаЉѕЃЛљзЊвЦЙ§ГЬжааЮГЩСђѕЅЛђепбѕѕЅжаМфЬхЃЌ ЖдЗДгІаджЪВњЩњЯджјгАЯь. вЛАуЖјбдЃЌ СђѕЅжаМфЬхЮШЖЈадНЯВюЃЌ дкЬхЭтгІгУжаОГЃЙлВьЕНжаМфЬхСђѕЅМќЫЎНтЕМжТЕФИБВњЮяЃЌ ЖјВЩгУбѕѕЅжаМфЬхЕФЗжСбФкКЌыФдђЗДгІЫйТЪНЯТ§. вђДЫбаОПепИљОнЗжСбФкКЌыФ1ЮЛКЭCЖЫЭтЯдыФ+1ЮЛЕФАБЛљЫсЃЌ НЋЦфЗжЮЊCCКЭCSЃЈTЃЉСНРр. CCРрЗжСбФкКЌыФЃЈШчSsp DnaEМАSce VMAЕШЃЉ1ЮЛКЭCЖЫЭтЯдыФ+1ЮЛЕФАБЛљЫсОљЮЊАыызАБЫсЃЌ ЗДгІОРњ2ИіСђѕЅжаМфЬхЙ§ГЬ. CSЃЈTЃЉРрЗжСбФкКЌыФЃЈШчSsp DnaBЃЌ gp41ЕШЃЉ1ЮЛЮЊАыызАБЫсЃЌ CЖЫЭтЯдыФ+1ЮЛЮЊЫПАБЫсЛђЫеАБЫсЃЌ ЗДгІжаОРњ1ИіСђѕЅжаМфЬхКЭ1ИібѕѕЅжаМфЬхЙ§ГЬ. Г§ДЫСНДѓРрЭтЃЌ ЛЙгаЩйЪ§ЗжСбФкКЌыФдкЗДгІжаОРњ2ИібѕѕЅжаМфЬхЙ§ГЬЃЌ ШчNeq DNA polymeraseЗжСбФкКЌыФЃл55Ѓн.
ЗжСбФкКЌыФвЛАуЗжЮЊ2ИіЦЌЖЮЃЌ 2ИіЦЌЖЮЗЧЙВМлНсКЯКѓдкНсЙЙКЭЙІФмЩЯНгНќгкСЌајЕФФкКЌыФ. ЮоТлЪЧШЫЙЄЗжСбЕФФкКЌыФЛЙЪЧЬьШЛРДдДЕФЗжСбФкКЌыФЃЌ ЦфЗжСбЮЛЕуОљЮЛгкНсЙЙЩЯЕФЛЗЧјЃЈLoopЃЉЃЌ ДгЖјБмУтгАЯьНсКЯКЭМєНгЗДгІЛюад. ИљОнЗжСбЮЛЕуВЛЭЌЃЌ ФкКЌыФПЩвдБЛЗжЮЊS0ЃЌ S1КЭS11ФкКЌыФЃл63Ѓн. S0ФкКЌыФДгдЪМФкКЌыФжаЙщГВКЫЫсФкЧаЧјгђЮЛЕуЧаИюЃЌ ШчSsp DnaEКЭNpu DnaEЕШ. S1ФкКЌыФДгНгНќNЖЫЕФЮЛжУЧаИюЃЌ ШчSsp DnaBКЭM86 DnaBЕШ. S11ФкКЌыФДгНгНќCЖЫЕФЮЛжУЧаИюЃЌ ШчSsp GyrB S11КЭSsp DnaB S11ЕШ. ДЫЭтЃЌ LiuЕШЃл63ЃнНЋDnaBДгS1КЭS0ЮЛЕуЗжЮЊ3ЖЮЃЌ вРШЛБЃСєНсКЯМАМєНгЙІФм.
2 ЗжСбФкКЌыФЕФЗДгІдРэ
2.1ЁЁЗжСбФкКЌыФЕФНсЙЙ
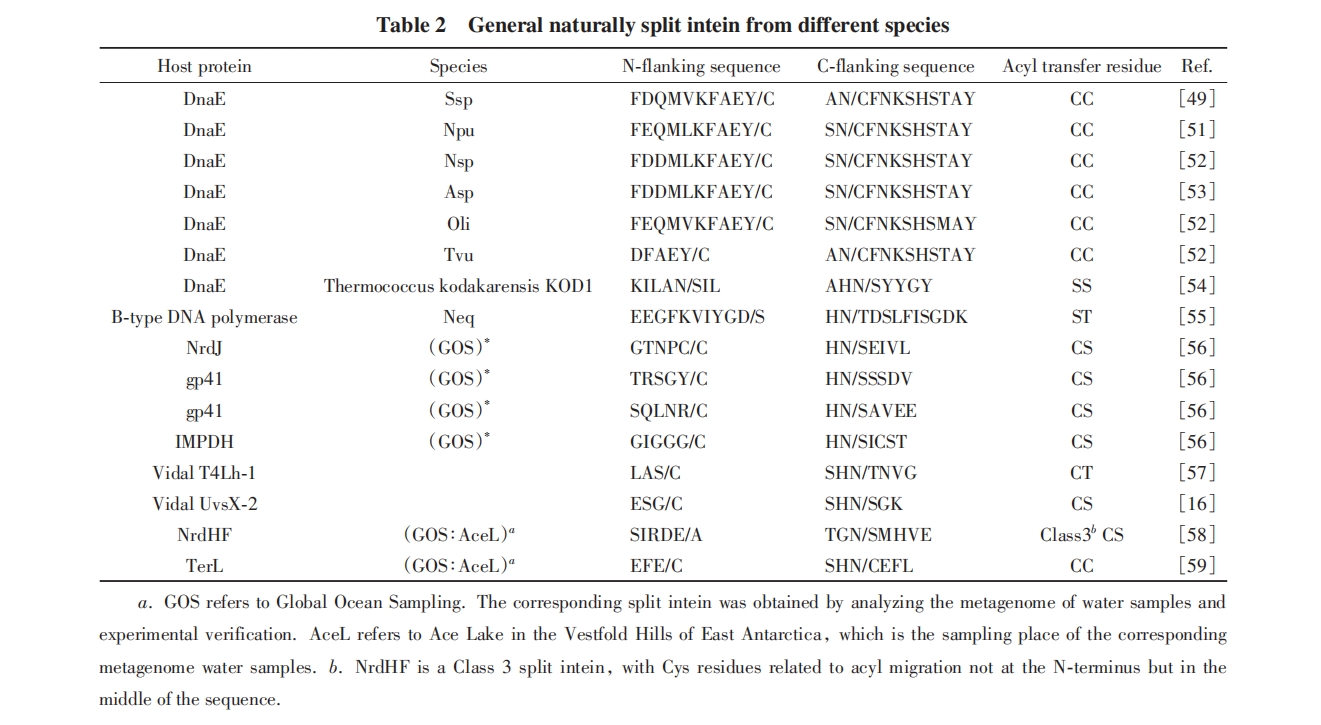
ЗжСбФкКЌыФПЩвдПДзїЪЧЭъећФкКЌыФЕФЗжСбаЮЪНЃЌ ЦфИДКЯЮяЕФНсЙЙгыЭъећФкКЌыФЯрЫЦЃл64ЃнЃЌ ЪЧЮБC2ЖдГЦадЕФТэЬуаЮЕААзЃЌ жївЊгЩІТелЕўзщГЩ. СНзщЗДЦНааЕФІТелЕўГЩТэЬуЕФЦНУцЃЌ ЦНУцЩЯЯТИїгавЛзщ2ИіІТелЕўКЭвЛЬѕІСТна§аЮГЩЕФНсЙЙгђЃЈІТІТІС motifЃЉЃЌ ЦфжавЛВрЕФІТелЕўбгЩьВЂДЙжБДЉЙ§ТэЬужабыЕФloopЃЌ гыNЖЫЭтЯдыФЯрСЌ. ТэЬужабыСэвЛЖдДЙжБгкЦНУцЕФІТелЕўгыCЖЫЭтЯдыФЯрСЌ. СНЖЮЭтЯдыФЮЛгкЦНУцЕФЭЌвЛВрЃлЭМ4ЃЈAЃЉКЭЃЈCЃЉЃнЃл65Ѓн.
ЗжСбФкКЌыФгаЖржжПЩФмЕФЗжСбЗНЪНЃЌ ЖрЪ§ЗжСбФкКЌыФЗжСбГЩвЛДѓвЛаЁ2ИіЦЌЖЮЃЌ ЗжСбЮЛЕуПЩФмдкППНќCЖЫЕФвЛВрЃЌ вВПЩФмдкППНќNЖЫЕФвЛВр. ЦфжаЃЌ ППНќCЖЫЕФЗжСбБЛГЦЮЊОЕфЃЈTypicalЃЉЗжСбЃЌ ЦфаЁЦЌЖЮвЛАуАќРЈжабыДЙжБЕФвЛзщІТелЕўКЭШЦЦНУцІТелЕўЕФвЛВПЗж. ППНќNЖЫЕФЗжСбБЛГЦЮЊЗЧОЕфЃЈAtypicalЃЉЗжСбЃЌ ЦфаЁЦЌЖЮвЛАуАќРЈжабыДЙжБЕФloopКЭЦНУцвЛВрІТІТІС motifЕФвЛВПЗж. ДЫЭтЃЌ LiuЕШЃл63ЃнЭЈЙ§ШЫЙЄЩИбЁSsp DnaBПЩФмЕФЗжСбЮЛЕуЃЌ ЪЕЯжСЫ3ЖЮЗжСбФкКЌыФЕФМєЧаЗДгІ. вбОдкNpu DnaEЩЯЪЕЯжРрЫЦЕФ3ЦЌЖЮЗжСбФкКЌыФМєЧаЗДгІЃЌ ЫЕУїЭЈЙ§ИФдьЕУЕНЖргк2ЦЌЖЮЕФЗжСбФкКЌыФЪЧПЩФмЕФЃлЭМ4ЃЈBЃЉКЭЃЈCЃЉЃн.
ЗжСбФкКЌыФашвЊИїзщЗжНсКЯКѓВХФмелЕўГЩе§ШЗЕФЙЙЯѓ. 2013ФъЃЌ MuirЕШЃл66ЃнЭЈЙ§КЫДХЙВеёВЈЦзЃЈNMRЃЉЙлВтСЫШмвКжаNpu DnaEЗжСбФкКЌыФНсКЯЧАКѓЕФЙЙЯѓЃЌ ЗЂЯжЗжСбФкКЌыФЦЌЖЮНсКЯЧАЃЌ аЁЦЌЖЮДІгкЮоађзДЬЌЃЌ ДѓЦЌЖЮДІгкЙЙЯѓЖЏЬЌБфЛЏЕФзДЬЌ. СНЦЌЖЮЕФЯрЛЅзїгУгеЕМаЁЦЌЖЮДгЮоађНсЙЙелЕўЮЊЮШЖЈЙЙЯѓЃЌ ВЂгеЕМДѓЦЌЖЮаЮГЩИќМгЮШЖЈЕФНсЙЙ. РрЫЦЕФNMRаХКХЬиеївВдкSsp DnaEКЭCatСНжжЗжСбФкКЌыФЩЯБЛбщжЄЃл67ЃЌ68Ѓн. 2018ФъЃЌ RibЈЎЕШЃл64ЃнНтЮіСЫNeq B-type DNA polymerase1ЃЈNeq Pol1ЃЉ NЖЫДѓЦЌЖЮКЭСНЦЌЖЮИДКЯЮяЕФОЇЬхНсЙЙЃЌ ЗЂЯжIntCЕЅЖРДцдкЕФНсЙЙгыаЮГЩИДКЯЮяКѓЕФНсЙЙгаУїЯдЧјБ№. етаЉНсЙћЫЕУїЗжСбФкКЌыФИїзщЗжЮоЗЈЖРСЂЮШЖЈелЕўГЩе§ШЗЕФЙЙЯѓ.
2.2ЁЁЗжСбФкКЌыФЕФНсКЯФЃЪН
ОВЕчЯрЛЅзїгУБЛШЯЮЊЪЧЗжСбФкКЌыФСНЦЌЖЮЪЖБ№КЭНсКЯЕФживЊЖЏСІ. 2007ФъЃЌ MuirЕШЃл69ЃнЭЈЙ§ађСаБШЖдЗЂЯжВЛЭЌЮяжжDnaEЗжСбФкКЌыФЯдЪОГіБЃЪиЕФЕчКЩЗжРыЬиеїЃКCЖЫаЁЦЌЖЮКЌгаМюадВаЛљМЏжаЦЌЖЮЃЌ NЖЫДѓЦЌЖЮКЌгаЫсадВаЛљМЏжаЦЌЖЮ. 2ИіЕчадЯрЗДЕФЦЌЖЮНщЕМЕФОВЕчЯрЛЅзїгУФЃЪНдкSsp DnaEМА Neq Pol1ЕФОЇЬхНсЙЙжаЖМЕУЕНСЫбщжЄЃл52ЃЌ64Ѓн.
ЗЧОЕфЗжСбФкКЌыФСНЦЌЖЮЕФНсКЯУцгыОЕфЗжСбФкКЌыФВЛЭЌ. 2018ФъЃЌ MuirЕШЃл67ЃнЭЈЙ§NMRНтЮіСЫвЛжжЗЧОЕфЗжСбФкКЌыФCatЕФНсЙЙЃЌ ЗЂЯж2ИіФкКЌыФЦЌЖЮЕФЯрЛЅзїгУНчУцжївЊЪЧЪшЫЎВаЛљЃЌ ЖрЪ§ДјЕчВаЛљБЉТЖдкШмМСжа. ЩњЛЏЪЕбщНсЙћвВБэУїЃЌ CatСНИіЦЌЖЮЕФНсКЯЧПЖШЪмШмвКРызгЧПЖШЕФгАЯьКмаЁЃЌ БэУїЪшЫЎЯрЛЅзїгУдквЛаЉЗЧОЕфЗжСбФкКЌыФЕФНсКЯжаЦ№ИќжївЊЕФзїгУЃЈЭМ5ЃЉ
2.3ЁЁЗжСбФкКЌыФЕФМєЧаЛњРэ
ЗжСбФкКЌыФЕФМєНгЪЧздЗЂЙ§ГЬЃЌ ВЛашвЊЦфЫќЕФЛюЛЏФмРДдД. ећИіЗДгІАќРЈN-SѕЃЛљЧЈвЦЁЂ СђѕЅНЛЛЛЁЂ ЬьЖЌѕЃАЗЛЗЛЏМАS-NѕЃЛљЧЈвЦЕШ4ИіЙ§ГЬЃЈЭМ6ЃЉ.
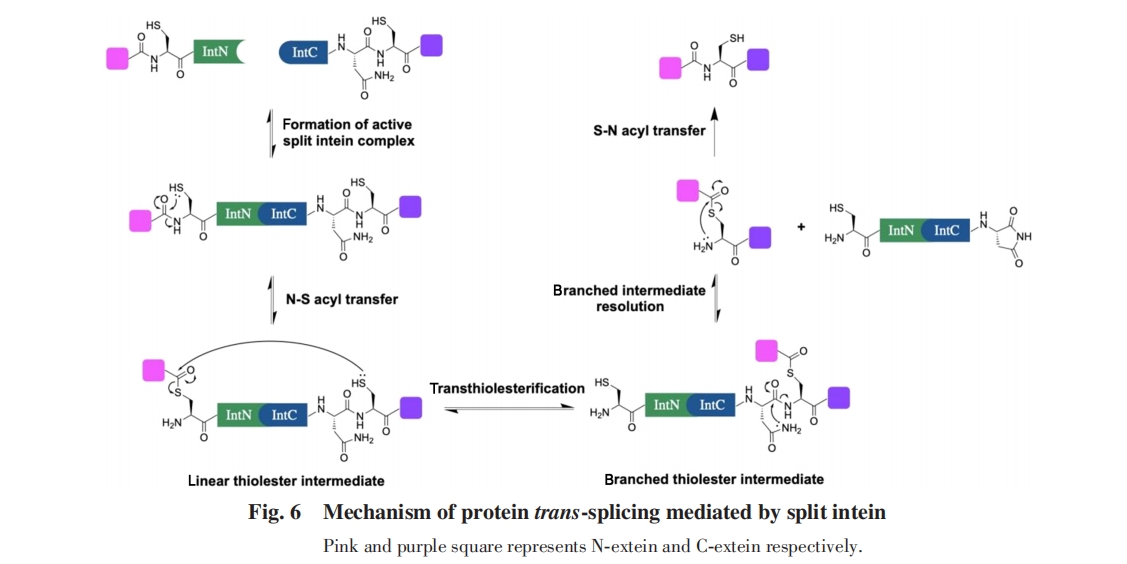
ЕквЛВНЃЌ ФкКЌыФN-ЖЫЕФCysВрСДлЯЛљНјЙЅжїСДѕЃАЗМќЃЌ ЗЂЩњN-SѕЃЛљЧЈвЦЃЌ ЪЙN-ЖЫЭтЯдыФЭЈЙ§СђѕЅМќСЌНгдкФкКЌыФЩЯ. дкДЫЙ§ГЬжаЃЌ ЗжСбФкКЌыФжажСЩйга3ИіАБЛљЫсЦ№ЕНДпЛЏзїгУЃЌ АќРЈгылЯЛљЯрЛЅзїгУЕФF-Block AspвдМАгыѕЃАЗМќЯрЛЅзїгУЕФB-Block HisКЭB-Block Thr. F-Block AspЦ№ЕНМюДпЛЏзїгУЃЌ ДйНјCysВрСДлЯЛљЕФШЅжЪзгЛЏЃЌ діЧПСЫлЯЛљЧзКЫадЃЌ ЕуЭЛБфЪЕбщБэУїИУАБЛљЫсЖдN-SѕЃЛљЧЈвЦЪЧБивЊЕФЃл70Ѓн. ЩЯЪіЛњРэгыNMRЙлВтЕНЕФлЯЛљpKaБфЛЏвдМАРэТлМЦЫуЕФНсЙћЯрЮЧКЯЃл71ЃЌ72Ѓн. жЪзгЛЏЕФB-Block HisЦ№ЕНЫсДпЛЏзїгУЃЌ дкжЪзгЛЏКѓГЩЮЊЧтМќЙЉЬхЃЌ гыѕЃАЗМќєЪЛљбѕЯрЛЅзїгУЃЌ діЧПСЫєЪЛљЕФЧзЕчад. етвЛЯрЛЅзїгУЕУЕНСЫРэТлМЦЫуЕФжЇГжЃл72ЃнЃЌ ВЂНјвЛВНдкSsp DnaB M86ФкКЌыФЕФОЇЬхНсЙЙжаЕУЕНСЫШЗШЯЃл73Ѓн. B-Block ThrЦ№ЕНМюДпЛЏзїгУЃЌ гыѕЃАЗМќЕФАБЛљЧтЯрЛЅзїгУЃЌ ЪЙѕЃАЗМќЦЋРыРэЯыМќВЮЪ§ЃЌ ЭЈЙ§ЬсЩ§ЕзЮяФмСПНЕЕЭСЫЛюЛЏФм. етвЛЛњРэЕУЕНСЫSsp DnaEФкКЌыФОЇЬхНсЙЙЕФжЇГжЃл74ЃнЃЌ ЕуЭЛБфЪЕбщБэУїИУАБЛљЫсЖдN-SѕЃЛљЧЈвЦЪЧБивЊЕФЃлЭМ7ЃЈAЃЉЃнЃл75Ѓн.
ЕкЖўВНЃЌ C-ЖЫЭтЯдыФЕФCysВрСДлЯЛљНјЙЅN-ЖЫЭтЯдыФгыN-ЖЫФкКЌыФжЎМфЕФСђѕЅМќЃЌ ЭЈЙ§СђѕЅНЛЛЛЪЙСНЖЮЭтЯдыФЭЈЙ§СђѕЅМќЯрСЌЃЌ ДгЯпадСђѕЅжаМфЬхзЊЛЏЮЊжЇСДСђѕЅжаМфЬх. QM/MMМЦЫуНсЙћБэУїЃЌ F-Block AspдкДЫЙ§ГЬжаЭЌбљЦ№ЕНМюДпЛЏЕФзїгУЃЌ ДйНјСЫC-ЖЫЭтЯдыФЕФCysЕФШЅжЪзгЛЏ. ЕуЭЛБфЪЕбщБэУїЃЌ НЋетИіAspЭЛБфЮЊGluЃЌ AsnЃЌ AlaЛђGlyЖМЛсЯджјвжжЦжЇСДжаМфЬхЕФаЮГЩЃлЭМ7ЃЈBЃЉЃнЃл76Ѓн.
ЕкШ§ВНЃЌ C-ЖЫФкКЌыФЕФC-ЖЫAsnВрСДѕЃАЗНјЙЅC-ЖЫФкКЌыФгыC-ЖЫЭтЯдыФжЎМфСЌНгЕФѕЃАЗМќЃЌ аЮГЩЖўѕЃбЧАЗЮхдЊЛЗЃЌ ФкКЌыФгыЭтЯдыФЭбРы. F-BlockКЭG-BlockжаЕФ2ИіHisЦ№ЕНСЫДпЛЏзїгУ. ЦфжаЃЌ F-Block HisДйНјСЫC-ЖЫAsnВрСДѕЃАЗЕФШЅжЪзгЛЏЃЌ Ц№ЕНМюДпЛЏзїгУ. жЪзгЛЏКѓЕФG-Block HisгыѕЃАЗМќєЪЛљбѕЯрЛЅзїгУЃЌ Ц№ЕНСЫдіЧПєЪЛљЧзЕчадЕФЫсДпЛЏзїгУ. ЩЯЪіЯрЛЅзїгУдкSsp DnaBЕФОЇЬхНсЙЙжаЕУЕНСЫЙлВтЃл77ЃнЃЌ вВЕУЕНСЫМЦЫуНсЙћЕФжЇГжЃл78Ѓн. ДЫЭтЃЌ MuirЕШЃл79ЃнЗЂЯжЖдгкNpu DnaEФкКЌыФЃЌ дкЮоЗЈаЮГЩжЇСДжаМфЬхЕФC1AЭЛБфЬхжаЃЌ AsnЛЗЛЏЫйТЪЛсНЕЕЭжСЭЛБфЧАЕФ1/200ЃЌ ЫЕУїжЇСДжаМфЬхЕФаЮГЩДйНјСЫAsnЕФЛЗЛЏЃлЭМ7ЃЈCЃЉЃн.
зюКѓЃЌ дкФкКЌыФДгЬхЯЕжаЭбРыКѓЃЌ СНЖЮЭтЯдыФМфЕФСђѕЅМќЭЈЙ§S-NѕЃЛљЧЈвЦаЮГЩЬьШЛыФМќЃЌ НјЖјЕУЕНМєЧаВњЮя. ИУЙ§ГЬВЛашвЊЦфЫќИЈжњЃЌ дкжаадЬѕМўЯТФмЙЛздЗЂНјааЃл80Ѓн.
дквЛаЉЗжСбФкКЌыФжаЃЌ ЩЯЪіЛњРэжаЕФCysБЛSerКЭThrЬцДњЃЌ ЗДгІЙ§ГЬжааЮГЩЯргІЕФбѕѕЅжаМфЬх ЃЈБэ1КЭБэ2ЃЉ. ДЫЭтЃЌ ВПЗжФкКЌыФЕквЛВНМєЧаЕФЧзКЫАБЛљЫсЮЛгкФкКЌыФФкВПЖјВЛЪЧNЖЫЃЌ MootzЕШЃл58ЃнБЈЕРСЫЕквЛИізёбетжжЛњРэЕФЬьШЛЗжСбФкКЌыФNrdHFЃЌ NrdHFВЮгыЕквЛВНN-SѕЃЛљЧЈвЦЕФCysЮЛгкIntCађСаЕФжаМфВПЮЛ.
Г§СЫФкКЌыФађСаФкВПАБЛљЫсЕФДпЛЏзїгУЃЌ ЗжСбФкКЌыФЛЙБэЯжГіЖдЭтЯдыФађСаЕФУєИаадЃКППНќМєНгЮЛЕуЕФЭтЯдАБЛљЫсЭЛБфЖдЗДгІаЇТЪКЭВњТЪгаУїЯдгАЯьЃЌ АЕЪОЭтЯдађСаПЩФмгыЙиМќАБЛљЫсДцдкЯрЛЅзїгУ. MuirЕШЃл81ЃнНЋNpu*ФкКЌыФЃЈSsp DnaE-NКЭNpu DnaE-CЕФзщКЯЃЉC-ЖЫЭтЯдыФДгЬьШЛЕФCFNИФЮЊSGVЃЌ ЗЂЯжCЖЫМєНгаЇТЪУїЯдЯТНЕ. ЭЈЙ§ЖЈЯђНјЛЏЃЌ ЕУЕНСЫПЩвдИќгааЇМєНгSGVзїЮЊC-ЖЫЭтЯдыФЕФmNpu*ЃЌ ЦфжаАќРЈF-Block D124YЭЛБф. ОЇЬхНсЙЙбаОПЯдЪОЃЌ CЖЫЭтЯдыФCFNжаЕФPheгыДйНјAsnЛЗЛЏЕФF-Block His125ДцдкІа-ІаЖбЕўзїгУЃЌ ЪЙHis125ДІгкгаРћгкДпЛЏЕФШЁЯђЃЌ ЖјD124YЭЛБфЪЙTyrДњЬцСЫPheЕФзїгУЃЌ ДгЖјМѕЧсСЫЗжСбФкКЌыФЖдC-ЖЫЭтЯдыФЕФУєИаад. MootzЕШЃл82ЃнЗЂЯжНЋSsp DnaB N-ЖЫЭтЯдыФЕФGlyИФБфЮЊL-AlaЛсЕМжТМєНгаЇТЪУїЯдЯТНЕЃЌ ЖјИФБфЮЊD-AlaЕФаЇТЪЯТНЕЗљЖШНЯаЁ. ЭЈЙ§ЙлВьОЇЬхНсЙЙЃЌ ШЯЮЊN-ЭтЯдыФЕФАБЛљЫсВрСДПЩФмгыB-Block HisВрСДВњЩњЮЛзшГхЭЛЃЌ ИЩШХHisЕФДпЛЏЙІФмЃЌ ЖјЦфЫќГЏЯђЕФВрСДЃЈШчD-AlaЃЉВњЩњЕФЮЛзшГхЭЛНЯаЁ. етаЉЪЕбщНсЙћЫЕУїЭтЯдыФгыФкКЌыФжЎМфДцдкЛЅзїЃЌ ПЩФмЪЧЭтЯдађСаУєИаадЕФживЊдвђ.
3 ЗжСбФкКЌыФЕФЙЄГЬЛЏИФдьгыадФмгХЛЏ
ЗжСбФкКЌыФвбОГЩЮЊвЛжжЙуЗКгІгУЕФЕААзжЪСЌНгЙЄОпЃЌ ЕЋдкЦфгІгУЙ§ГЬжавРШЛДцдквЛаЉЬєеНЃЌ вђДЫбаОПепЯЃЭћФмЛёЕУОпгаИќгХадФмЕФФкКЌыФЃЌ АќРЈМєЧаЛюадКЭЮШЖЈадИпЁЂ ЖдЭтЯдыФМєНгЮЛЕуИННќАБЛљЫсађСаЕФФЭЪмадЧПЃЌ ВЛЭЌЗжСбФкКЌыФМфЕФЗДгІе§НЛЃЌ ЗжСбФкКЌыФађСаЖЬЃЌ C-ЖЫЛђN-ЖЫГЄЖШдкЙЬЯрКЯГЩЗЖЮЇФкЕШ. ЮЊЛёЕУОпгаЩЯЪіЬиадЕФЗжСбФкКЌыФЃЌ баОПепУЧНјааСЫЙуЗКЕФЬНОПгыГЂЪд.
3.1ЁЁЗжСбФкКЌыФЕФМєЧаЛюадКЭЮШЖЈад
2016ФъЃЌ MuirЕШЃл83ЃнЪЙгУРэадЩшМЦЗНЗЈЛёЕУСЫОпгаЧПЮШЖЈадКЭИпМєЧаЛюадЕФЗжСбФкКЌыФ. ЭЈЙ§ЗжЮіDnaEМвзхЕФNpuЃЈПьЃЉКЭSspЃЈТ§ЃЉЗжСбФкКЌыФжЎМфМєНгаЇТЪЕФВювьЗЂЯжЃЌ ДѓЖрЪ§ЙиМќВаЛљдкПеМфНсЙЙЩЯгыЛюадЮЛЕужБНгСкНќ. вдетаЉЙиМќАБЛљЫсВаЛљзїЮЊЖдЦыЮЛЕуЃЌ Жд73ИіDnaEМвзхЕФФкКЌыФНјааБШЖдЃЌ зюжеЕУЕНСЫвЛИіаТаЭЗжСбФкКЌыФCfa. CfaЯдЪОГіПьЫйЕФЕААзжЪМєНгЛюадвдМАИпЕФШШЮШЖЈадКЭБфадЮШЖЈад. дк30 ЁцЯТЃЌ CfaЕФМєЧаЫйТЪБШДЫЧАБЈЕРЕФDnaEМвзхжазюПьЕФNpu DnaEПь2.5БЖЃЌ Цфt1/2НіЮЊ20 s. ЭЌЪБЃЌ CfaЖдБфадЬѕМўОпгаКмЧПЕФФЭЪмадЃЌ дк3~4 mol/L GnЁЄHClЛђ8 mol/LФђЫиЕШЧПБфадЬхЯЕжаЖМБЃГжСМКУЕФМєНгЛюадЃЌ МєНгЗДгІЦНКтГЃЪ§K>1ЁС10-2 s-1.
3.2ЁЁЗжСбФкКЌыФЖдМєНгЮЛЕуЕФМцШнад
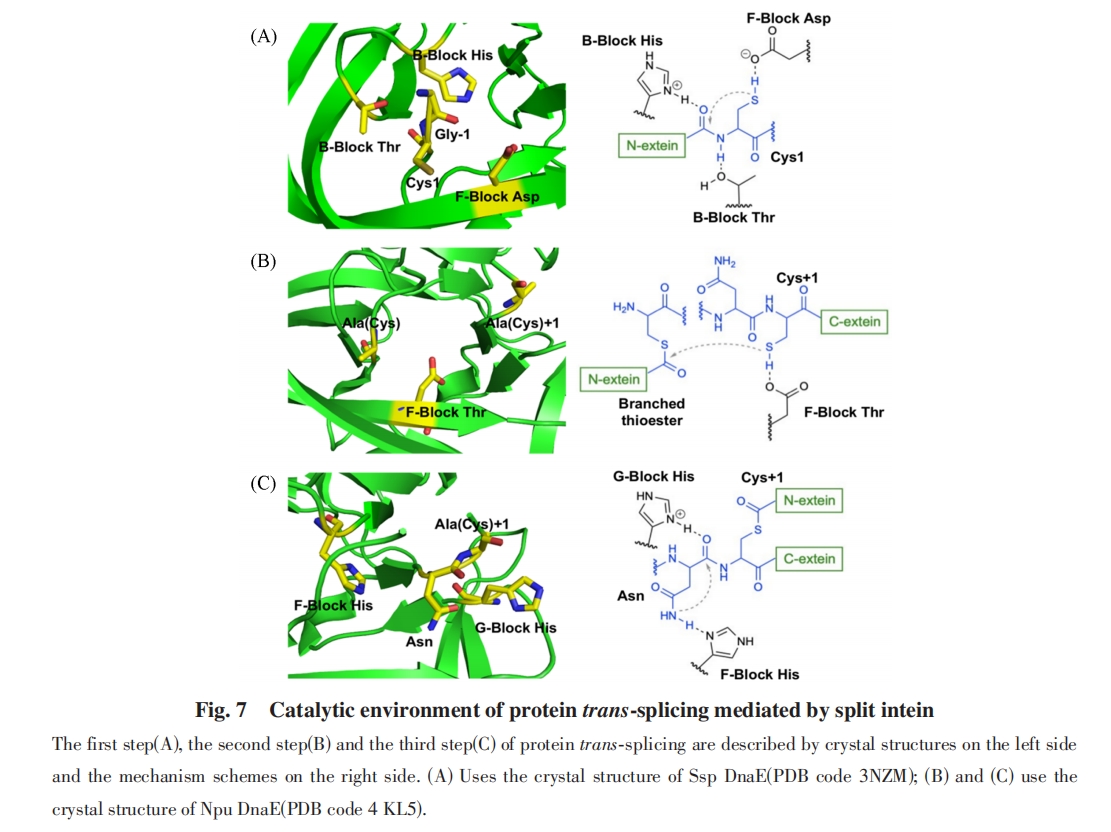
2011ФъЃЌ LiuЕШЃл84ЃнЭЈЙ§ЖЈЯђНјЛЏММЪѕЃЌ ЕУЕНСЫОпгаИпЭтЯдыФМєНгЮЛЕуМцШнадЕФФкКЌыФ. НЋSsp DnaBФкКЌыФЛљвђВхШыЕНПЈФЧУЙЫиПЙадЕААзжЪЃЈKanRЃЉЕФЛљвђжаЃЌ ЪЙЕУКЌФкКЌыФЕФKanRЕААзЧАЬхУЛгаПЈФЧУЙЫиПЙадЃЌ жЛгаЗЂЩњСЫЕААзМєНгЗДгІЕУЕНЭъећKanRЕААзЃЌ ПЈФЧУЙЫиПЙадЛжИДЕФОњжъВХФмдкКЌПЈФЧУЙЫиЕФХрбјЛљжаДцЛю. ЖдДцЛюОњжъЕФжЪСЃНјааВтађЃЌ МДПЩЛёЕУФмЙЛдкИУВхШыЮЛЕуЗЂЩњМєНгЗДгІЕФФкКЌыФЛљвђађСаЃЈЭМ8ЃЉ. ОЙ§ЖрИіЩИбЁбЛЗЃЌ НјЛЏЕУЕНСЫЖрИіаТаЭФкКЌыФЃЌ ФмЙЛдкЖрИіВЛЭЌЕФЭтЯдыФМєНгЮЛЕуКЭВЛЭЌЕФЕААзжЪжаЖМЯдЪОГіИпМєНгЛюад. ЦфжаЃЌ M86ЭЛБфЬхФкКЌыФОпгазюКУЕФМєНгЗДгІадФм. НЋM86ЭЈЙ§ШЫЙЄЗжСбЮЊNЖЫгЩ11ИіАБЛљЫсзщГЩЕФЗжСбФкКЌыФ. гыЭЛБфЧАЕФЗжСбDnaBФкКЌыФЯрБШЃЌ M86ДпЛЏЕААзжЪЗДЪНМєНгЗДгІЕФЫйТЪдіМгСЫ60БЖЃЌ ЗДгІГЃЪ§kM86=2.5ЁС10-3 s-1ЃЌ ВЂЧвЗжСбФкКЌыФЦЌЖЮЕФKdжЕЬсИпСЫвЛИіЪ§СПМЖ.
2017ФъЃЌ MuirЕШЃл85ЃнБЈЕРСЫЭтЯдыФађСаФЭЪмадУїЯдЬсИпЕФздШЛЗжСбФкКЌыФ. His125ЪЧЕААзжЪМєНгзюКѓвЛВНжаAsn137ЛЗЛЏЕФЙиМќДпЛЏВаЛљЃЌ ЭЈЙ§ЩшМЦHis125ЫљдкЕФЛЗЃЈВаЛљ122~124ЃЉЃЌ ПЩвдгааЇЕиИФБфHis125ЙЙЯѓЖЏСІбЇЃЌ ДгЖјЯджјдіМгМєЧаЗДгІЖд+2гы+3ЮЛжУЕФЭтЯдыФађСаФЭЪмадЃЌ ВЂЬсИпСЫМєЧаЫйТЪ. ЛљгкЖдЕААзжЪМєНгЙиМќЕФДпЛЏЮЛЕуЕФРэНтЃЌ MuirЕШЖдЗжСбФкКЌыФHis125ЫљдкЕФЛЗНјааСЫБЅКЭЭЛБфЃЌ ШЛКѓЪЙгУЛљгкЯИАћЕФбЁдёЯЕЭГРДМјЖЈФмЙЛдкВЛЭЌЭтЯдыФМєНгЮЛЕуЯТДпЛЏгааЇМєНгЕФЭЛБфЬхЃЌ ЗЂЯжДѓЖрЪ§ОњТфдкВаЛљ122~124ДІКЌгаGXPЛљађЃЈGRPЃЌ GEPЛђGPPЃЌ ДњЬцЬьШЛERDађЃЉЃЌ ЦфжаGEPађСаОпгазюИпЕФМєНгЛюадЃЈЭМ9ЃЉ.
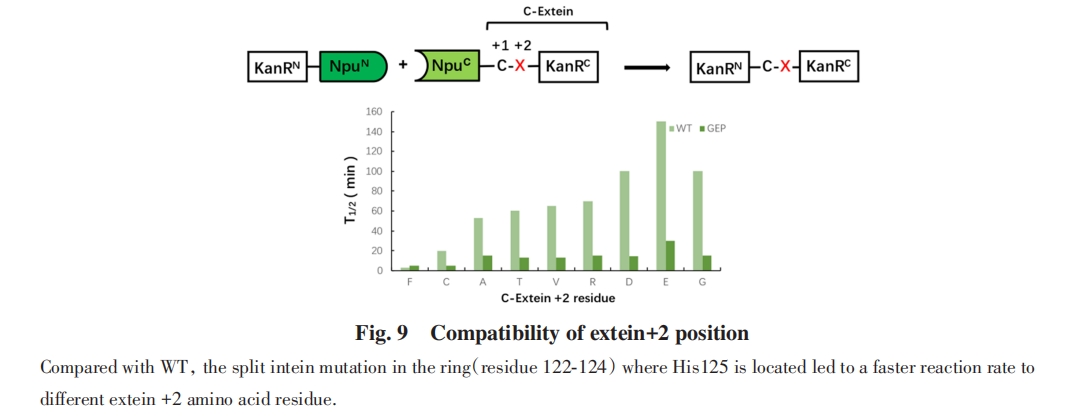
баОПБэУїЃЌ Npu DnaEФкКЌыФЕФЭтЯдыФађСаЦЋКУжївЊОжЯогкCЖЫЭтЯдыФ+1ЮЛЕФДпЛЏАыызАБЫсКЭ+2ЮЛЕФДѓЮЛзшЪшЫЎВаЛљЃЌ НЋЬьШЛ+2ЮЛPheЭтЯдыФађСаВаЛљЭЛБфЮЊЬхЛ§НЯаЁЕФВаЛљЃЈР§ШчAlaЃЉЛсЕМжТМєНгТЪЯджјНЕЕЭЃл79Ѓн. MuirЕШЃл85ЃнвдGFPЕААззїЮЊФЃаЭЃЌ БэеїСЫЖдгкВЛЭЌЭтЯдыФЮЛЕуЕФМєНгЛюад. ОЙ§GEPађСаИФдьЕФNpu DnaEФкКЌыФгыЮДИФдьЕФNpu DnaEФкКЌыФЯрБШЃЌ ЖдВЛЭЌЕФ+2КЭ+3ЮЛЭтЯдыФађСаЕФМєНгВњТЪОљгаЯджјЬсИпЃЈЭМ10ЃЉ.

3.3ЁЁЗжСбФкКЌыФЕФЗДгІе§НЛад
ЗжСбФкКЌыФЕФNЖЫКЭCЖЫЦЌЖЮжЎМфОпгаЧПСвЕФЕчКЩ-ЕчКЩЯрЛЅзїгУ. 2019ФъЃЌ IwaïЕШЃл86ЃнЛљгкЖдЗжСбФкКЌыФ2ИіЦЌЖЮНсКЯЙ§ГЬЁАВЖЛёКЭелЕўЛњжЦЁБжаЕФОВЕчЯрЛЅзїгУЃЌ НЋ2ИіЗжСбФкКЌыФЦЌЖЮНсКЯЙиМќЮЛжУЕФАБЛљЫсЕчКЩЭјТчНјааИФдьЃЌ ЪЕЯжСЫ2жжDnaBМвзхЗжСбФкКЌыФНсКЯЕФе§НЛад. етвЛВпТдвВЪЪгУгкДгЦфЫќЫГЪНМєНгФкКЌыФжаВњЩњаТЕФЗжСбФкКЌыФ.
2020ФъЃЌ WangЕШЃл87ЃнЩИбЁМьВтСЫЬьШЛДцдкЕФЗжСбФкКЌыФЕФМєНгЗДгІЃЌ НЈСЂСЫАќКЌ15жжЗжСбФкКЌыФЕФе§НЛЗжСбФкКЌыФПт. баОПНсЙћБэУїЃЌ е§НЛЗжСбФкКЌыФПЩвдгыЖрИіЗжСбзЊТМвђзгНсКЯЃЌ вддкЛюЩњЮяЬхжаЪЕЯжИДдгЕФТпМБэДяЙ§ГЬЃЌ ЛЙПЩвдгУгкЖрЦЌЖЮЕААзЕФЬхЭтСЌНг. зїепвдSasGЕААзжиИДЦЌЖЮНјааеЙЪОЃЌ ЪЙгУе§НЛЗжСбФкКЌыФзщКЯЃЌ вЛЙјЭъГЩСЫ6ИіЦЌЖЮЕФЦДНгЃЈЭМ11ЃЉЃЌ жЄУїСЫе§НЛЗжСбФкКЌыФРЉеЙПтЕФЖрЙІФмадКЭОоДѓЧБСІ.

ОЁЙмЗжСбФкКЌыФНщЕМЕФPTSдкЕААзжЪАыКЯГЩЩЯШЁЕУСЫвЛЖЈЕФГЩЙІЃЌ ЕЋвВДцдквЛаЉОжЯоЃЌ ШчаоЪЮЛљЭХЕФбЁдёадв§ШыЮЛЕуЭЈГЃжЛФмЮЛгкФПБъЕААзЕФNЖЫЛђCЖЫ. ЫцзХдНРДдНЖрЕФЗжСбФкКЌыФБЛМјЖЈЛђЩшМЦГіРДЃЌ ЗжСбФкКЌыФЙЄОпЯфж№НЅЗсИЛЃЌ дкЭЌвЛФПБъЕААзЕФАыКЯГЩжаЭЌЪБРћгУ2жжЯрЛЅе§НЛЕФЗжСбФкКЌыФНјааЖрДЮPTSЕФВпТдГЩЮЊПЩФм.
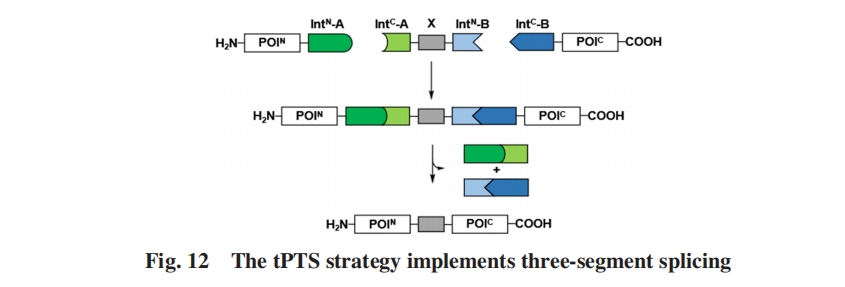
MuirЕШЃл88ЃнЪзДЮПЊЗЂСЫЭЈЙ§2жже§НЛЗжСбФкКЌыФЪЕЯжвЛЙјШ§ЦЌЖЮЗДЪНМєНгЕФВпТдЃЌ ИУЗНЗЈБЛГЦЮЊДЎСЊЕААзЗДЪНМєНгЃЈtPTSЃЉ. ФПБъЕААзЗжЮЊ3ИіЦЌЖЮЃЌ МДPOINКЭPOICвдМАвЛИіЭЈЙ§БэДяЛђЛЏбЇКЯГЩЛёШЁЕФДјгаЫљашаоЪЮЛљЭХЕФНЯЖЬжааФЖрыФЦЌЖЮXЃЈЭМ12ЃЉ. ЫљЪЙгУЕФ2Иіе§НЛЗжСбФкКЌыФЃЈAКЭBЃЉгыPOINКЭ POICШкКЯБэДяЃЈPOIN+IntN-AМАIntC-B+POICЃЉЃЌ ЖјXгыAКЭBЕФСэЭт2ИіЦЌЖЮШкКЯЃЈIntC-A+X+IntN-BЃЉ. етжжаТгБЕФЯЕЭГПЩвдЪЕЯж3ИіЖрыФЦЌЖЮЕФвЛЙјЛЏзщзАЃЌ КЯГЩСЫ304ИіВаЛљДѓаЁЕФc-CrkIIЕААзЃЌ ЦфгЩвЛИіSH2НсЙЙгђКЭ2ИіSH3НсЙЙгђзщГЩЃЌ УПИіНсЙЙгђЗжБ№гыЯргІЗжСбФкКЌыФЙЙГЩtPTSЕФЧАЬхЦЌЖЮ. ЗДгІдк22 ЁцЯТНјааСЫ48 hЃЌ вддМ48%ЕФЪеТЪЭъГЩСЫШ§ЦЌЖЮМєНг. ИУЗНЗЈЮЊНЯДѓЕФЖрНсЙЙгђЕААзЕФАыКЯГЩМАЕААзађСажабыЬиЖЈЮЛЕув§ШыЛЏбЇаоЪЮЬсЙЉСЫвЛжжЗНБуЕФЗНЗЈ.
3.4ЁЁЗжСбФкКЌыФЕФЪЙгУБуНнад
ЬьШЛЖЯСбЕФgp41-1ФкКЌыФЪЧФПЧАБЈЕРЕФзюаЁЕФЗжСбФкКЌыФжЎвЛЃЌ ОпгаЗЧГЃИпЕФЗДЪНМєНгЛюадЃл56ЃнЃЌ gp41-1ФкКЌыФгЩ88ИіВаЛљЕФIntNКЭ37ИіВаЛљЕФIntCзщГЩ. ЮЊСЫЪЙФкКЌыФЖдЭтЯдыФаджЪЕФИЩШХИќаЁЃЌ ШЫУЧЭЈГЃЯЃЭћЪЙФкКЌыФОЁПЩФмаЁ. IwaïЕШЃл86ЃнГЂЪдДгgp41-1ФкКЌыФЕФздШЛЗжСбЮЛЕуДІНиЖЬ2ИіВаЛљЃЌ ДгЖјЛёЕУИќаЁЕФЗжСбФкКЌыФ. ШЛЖјЃЌ етжжШБЪЇЪЙФкКЌыФЕФЕААзжЪМєНгаЇТЪНЕЕЭСЫдМ40%ЃЌ зїепКѓајЪдЭМгХЛЏФкКЌыФЕФАБЛљЫсађСавдЛжИДНиЖЬЕФgp41-1ФкКЌыФЕФМєНгЛюадЕНдгаЫЎЦНЃЌ ЕЋУЛгаГЩЙІ. етБэУїgp41-1ФкКЌыФЕФДѓаЁПЩФмвбДяЕНЪЕЯжЙІФмЕФзюаЁжЕ.
NeqЃЌ NpuКЭSsp DnaEЕШДѓВПЗжЬьШЛЗжСбФкКЌыФЖМдкГЃЙцЕФФкКЌыФЗжСбЮЛЕуS0ДІЖЯСбЃЌ ИУЮЛЕувВГфЕБЙщГВКЫЫсФкЧаУИВхШыЕФБъзМЮЛжУ. ИУЮЛЕуППНќФкКЌыФCЖЫЃЌ ЖЯСбВњЩњдМ100ИіАБЛљЫсВаЛљЕФIntNКЭ35ИіАБЛљЫсВаЛљЕФIntC. IntCНЯЖЬЃЌ БугкгыCЖЫЭтЯдыФЦЌЖЮЙВЭЌЙЬЯрКЯГЩЃЌ вђДЫетбљЕФЗжСбФкКЌыФЮЊЕААзжЪCЖЫАыКЯГЩвдМАБъМЧЬсЙЉСЫЗНБу. ШЛЖјЃЌ гЩгкЕААзжЪЙЬЯрКЯГЩФмСІЕФЯожЦЃЌ ЭЈЙ§ЙЬЯрКЯГЩгыN-ФкКЌыФЯрСЌЕФЕААзЪЧЯрЖдРЇФбЕФЃЌ етвВЯожЦСЫЗжСбФкКЌыФдкNЖЫАыКЯГЩЕААзжЪЛђепЕААзжЪNЖЫаоЪЮЕШГЁОАЕФгІгУ. вђДЫЃЌ баОПепУЧЯЃЭћбАевЛђПЊЗЂОпгаИќЖЬNЖЫЦЌЖЮЕФЗжСбФкКЌыФ.
2004ФъЃЌ LiuЕШЃл63ЃнНЋSsp DnaBУдФуФкКЌыФДгЗЧГЃЙцЕФЗжСбЮЛЕуЧаИюЃЌ БЈЕРСЫ3ИіаТЕФЗжСбЮЛЕуS1ЃЌ S8КЭS11ЃЌ Дгет3жжЮЛЕуЧаЖЯЃЌ ЖМВњЩњСЫФмЙЛНјааЕААзжЪЗДЪНМєНгЕФСНЖЮЪНЗжСбФкКЌыФЃЈЭМ13ЃЉ. ЦфжаS1ЗжСбФкКЌыФЕФNЖЫНіга11ИіАБЛљЫс. гЩгкВЛЭЌЕФФкКЌыФОпгаЯрЫЦЕФНсЙЙЃЌ етаЉаТЕФЗжСбЮЛЕуПЩвдЭЦЙуЕНЦфЫќФкКЌыФ. LiuЕШЃл63ЃнЛЙЪзДЮжЄУїСЫЗжСбЮЊ3ЖЮЕФЗжСбЕФФкКЌыФвРШЛПЩвдзщГЩИДКЯЮяВЂЛжИДЕААзжЪМєНгЛюад.

2014ФъЃЌ MootzЕШЃл59ЃнДгКъЛљвђзщЪ§ОнжаМјЖЈВЂБэеїСЫаТЕФЗжСбФкКЌыФAceL-TerL. етЪЧЪзДЮЗЂЯжДцдкЬьШЛЗЧЕфаЭЗжСбЕФФкКЌыФ. жЛга25ИіАБЛљЫсЕФNЖЫЦЌЖЮЪЧЦљНёЮЊжЙзюЖЬЕФЬьШЛIntNЦЌЖЮЃЌ взгкЭЈЙ§ЙЬЯрКЯГЩЛёШЁ. дк8 ЁцЕФЕЭЮТЯТЃЌ ИУФкКЌыФОпгазюИпДя90%ЕФЕААзжЪЗДЪНМєНгаЇТЪ. зїепЭЈЙ§ЖЈЯђЕААзжЪНјЛЏбЁдёСЫНјвЛВНИФНјЕФЭЛБфЬх. ЙЄГЬЛЏЕФФкКЌыФЭЛБфЬхдк37 ЁцЯТОпгаИќИпЕФМєНгаЇТЪЃЌ ЗДгІЫйТЪГЃЪ§k>1.8ЁС10-3 s-1ЃЌ ВЂдкЛЏбЇБъМЧЖржжЕААзжЪгІгУжаеЙЯжГіЬьШЛађСазюИп50БЖЕФаЇТЪ.
4 ЗжСбФкКЌыФдкЕААзжЪРыЬхгыдкЬхКЯГЩжаЕФгІгУ
4.1ЁЁРыЬхЕААзжЪАыКЯГЩ
ЗжСбФкКЌыФНщЕМЕФPTSЮЊЕААзжЪАыКЯГЩЬсЙЉСЫвЛжжаТЭООЖ. ВЛЭЌгкNCLЃл15ЃнКЭБэДяЕААзСЌНг ЃЈEPLЃЉЃл89~92ЃнЕШБъзМЕААзжЪЛЏбЇСЌНгЗДгІашвЊНЯИпХЈЖШЃЈвђЦфвРРЕгкЖрыФЦЌЖЮЕФЫцЛњХізВЃЉЃЌ PTSФмЙЛдкІЬmol/LХЈЖШЬѕМўЯТЗДгІЃЌ ЯдЪОГіНЯЕЭХЈЖШвРРЕадЃЈвђЦфгЩ2ИіЦЌЖЮжЎМфНєУмЕФЕААзжЪ-ЕААзжЪЯрЛЅзїгУДйГЩЃЉЃл69ЃнЃЌ етЪЙЕУЗжСбФкКЌыФГЩЮЊЕААзжЪАыКЯГЩЕФгаЮќв§СІЕФЙЄОп.
ЭЈЙ§PTSЭъГЩАыКЯГЩЕФФПБъЕААзЃЈPOIЃЉНЋБЛВ№ЗжЮЊNЖЫЃЈPOINЃЉгыCЖЫЃЈPOICЃЉ2ИіЧАЬхЦЌЖЮЃЌ ЦфзїЮЊЭтЯдыФЗжБ№гыЯргІЕФЗжСбФкКЌыФЦЌЖЮЯрСЌНгЃЌ гЩДЫзщГЩЕФШкКЯЬхЦЌЖЮЗжБ№ЖМПЩвддкPTSЗДгІжЎЧАЕЅЖРжЦБИВЂНЋЫљашЕФаоЪЮв§ШыЯргІЕФЦЌЖЮжа. гЩгкаэЖрОЙ§гХЛЏЕФЗжСбФкКЌыФЦЌЖЮНЯаЁЃЌ ЙЪКЌгаНЯаЁЗжСбФкКЌыФЦЌЖЮЕФФПБъЕААзЧАЬхЦЌЖЮЃЈPOIN⁃IntNЛђ IntC⁃POICЃЉПЩЭЈЙ§ЙЬЯрЖрыФКЯГЩЛёЕУЃЌ ДгЖјПЩвдШнвзЕиНЋаоЪЮЛљЭХбЁдёадв§ШыФПБъЕААзЕФN-ЖЫЛђC-ЖЫ.
MootzЕШЃл11ЃнНЋSsp DnaBЗжСбФкКЌыФгУгкЕААзжЪАыКЯГЩвдЪЕЯжNЖЫаоЪЮ. ИУIntNЦЌЖЮНіКЌ11ИіАБЛљЫсЃЌ БугкЭЈЙ§ЙЬЯрЖрыФКЯГЩЭЌЪБжЦБИIntNгыКЌаоЪЮЛљЭХЕФФПБъЕААзЦЌЖЮЃлЭМ14ЃЈAЃЉЃн. ЮЊжЄУїPTSдкN-ЖЫаоЪЮЕААзжЪАыКЯГЩжаЕФаЇЙћЃЌ Жд2жжЕААзжЪЕзЮяНјааСЫN-ЖЫгЋЙтЫиБъМЧ. КЯГЩыФ5-Fl-SEFSG-IntNгыШкКЯБэДяЕФЕААзЦЌЖЮIntC-Trx-His6вдЕШФІЖћХЈЖШЃЈ62 ІЬmol/LЃЉдк25 ЁцЯТЗДгІ22 hЃЌ ОДПЛЏКѓЕФАыКЯГЩВњЮяFl-Trx-His6змЪеТЪдМЮЊ30%. ЕкЖўИіР§згЪЧЗжзгСП30700ЕФІТ-ФкѕЃАЗУИЃЈІТLacЃЉ. ШкКЯБэДяЕААзЦЌЖЮIntC-ІТLac-His6гы5-Fl-SEFSG-IntNдк25 ЁцЃЌ 12 ІЬmol/LХЈЖШЯТЙВЗѕг§42 hЃЌ вддМ30%ЕФЪеТЪЕУЕНМєНгВњЮя Fl-ІТLac-His6.
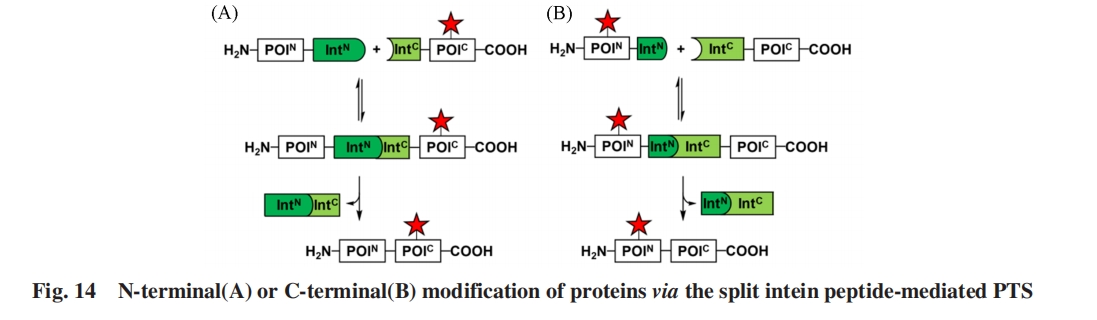
LiuЕШЃл93ЃнБЈЕРСЫвЛжжаТаЭЙЄГЬЛЏS11 Ssp GyrBЗжСбФкКЌыФ. ЦфЗжСбЮЛЕудкC-ЖЫИННќЃЌ ВњЩњГЄЖШдМ150ИіАБЛљЫсВаЛљЕФIntNЦЌЖЮКЭГЄЖШНіЮЊ6ИіАБЛљЫсВаЛљЕФIntCЦЌЖЮЃЌ ПЩгУгкНЋКЯГЩыФСЌНгЕНБэДяЕААзЕФCЖЫЃлЭМ14ЃЈBЃЉЃнЃл94Ѓн. етаЉаТгБЕФАќКЌЗЧГЃЖЬЕФNЖЫЛђCЖЫЦЌЖЮЕФЗжСбФкКЌыФЯджјРЉеЙСЫЦфдкЕААзжЪАыКЯГЩжаЕФгІгУЃЌ ЪЙЕУНЋаоЪЮЛљЭХЬэМгЕНФПБъЕААзЕФNЖЫЛђCЖЫИќМгЗНБу.
РћгУЗжСбФкКЌыФЛЙПЩвдЪЕЯжЕААзжЪЕФЮЛЕубЁдёадаоЪЮ. ЮЊСЫПЫЗўЖдКЌгаЖрИіАыызАБЫсЕФФПБъЕААзФбвдЪЕЯжбЁдёадаоЪЮЕФЯожЦЃЌ МДдкЦфжавЛИіЬиЖЈАыызАБЫсЮЛЕуЕФВрСДлЯЛљв§ШыаоЪЮЖјВЛгАЯьЦфгрАыызАБЫсВаЛљЃЌ MootzЕШЃл13ЃнНЋАыызАБЫсаоЪЮгыPTSНсКЯЪЙгУ. ОпЬхРДЫЕЃЌ ШЫЙЄЗжСбЕФSsp DnaBКЭMxe GyrAЕФIntCЦЌЖЮЕФЭтЯдыФ+1ЮЛЕуЗжБ№гЩЫПАБЫсЛђЫеАБЫсЖјЗЧАыызАБЫсРДНщЕМPTS. НЋетаЉЗжСбФкКЌыФЦЌЖЮгыОпгаЕЅИіАыызАБЫсЕФЖЬыФађСаЃЈГЦЮЊCysTagЃЉШкКЯБэДяЃЌ дкИУЦЌЖЮжав§ШыгЋЙтЭХЁЂ ЩњЮяЫиМАОлввЖўДМЃЈPEGЃЉЕШаоЪЮКѓЃЌ ОPTSЛёЕУШЋГЄЕФФПБъЕААз. ИУЗНЗЈЪЕЯжСЫдкКЌгаЖрИіАыызАБЫсЕФФПБъЕААзЃЌ ШчСђбѕЛЙЕААзЃл13ЃнЁЂ ІТ-ФкѕЃАЗУИЃл13ЃнЁЂ ШЫЩњГЄМЄЫиКЭЗЧКЫЬЧЬхыФКЯГЩУИTycAЃл95ЃнЕШЕААзжЪжаЕФЕЅвЛАыызАБЫсЮЛЕуЕФбЁдёадаоЪЮ.
НќЦкЃЌ PlessЕШЃл96ЃнНЋИУВпТдгІгУгкЬхФкЕААзжЪаоЪЮЃЌ дЪаэНЋЖржжаоЪЮЃЈАќРЈСзЫсЛЏЁЂ ввѕЃЛЏФЃФтЮяКЭЗЧЬьШЛАБЛљЫсЕШЃЉЭЌЪБв§ШыЛюЕФецКЫЯИАћжа. ЭЈЙ§ЬхФкtPTSВпТдЃЌ дкNaV1.5РызгЭЈЕРЕФАћФкгђКЭP2X2ЪмЬхЕФАћЭтгђжав§ШыСЫаоЪЮЃЌ баОПСЫЗвыКѓаоЪЮдкРызгЭЈЕРЙІФмвдМАЪмЬхгыХфЬхНсКЯжаЕФзїгУ. е§НЛЗжСбФкКЌыФНщЕМЕФДЎСЊЕААзЗДЪНМєНгдкЬхЭтКЭЬхФкЕФЕААзЖрЦЌЖЮМєНгЖМеЙЯжСЫСМКУЕФаЇЙћЃЌ гаЭћдкЕААзжЪАыКЯГЩжаЕУЕНИќЖрЕФгІгУ.
4.2ЁЁЦЌЖЮЭЌЮЛЫиБъМЧ
дкФПБъЕААзжЪжав§ШыОпгаNMRаЇгІЕФ15NМА13CЕШжидзгКЫЕФЭЌЮЛЫиБъМЧПЩПДзїЪЧЩЯЪіЭЈЙ§PTSНЋЛЏбЇаоЪЮв§ШыЕААзжЪЕФвЛИіОпЬхР§згЃЌ етвЛММЪѕзюживЊЕФгІгУЪЧгУгкКЫДХЙВеёВЈЦзбаОПРДБэеїЕААзжЪЕФНсЙЙКЭЖЏСІбЇ. ШЛЖјЖдгкНЯДѓЕААзжЪРДЫЕЃЌ ШЋГЄЭЌЮЛЫиБъМЧЕФбљЦЗжаЯджјЕФЗхжиЕўЪЙЕУКЫДХЙВеёЦзЕФНтЮіБфЕУРЇФб.
ЪЙгУЗжСбФкКЌыФНщЕМЕФPTSПЩвдНЋОпгажиЭЌЮЛЫиБъМЧЕФЕААзжЪЦЌЖЮгыЮоБъМЧЕФЦЌЖЮСЌНгЦ№РДЃЌ ЩњГЩВПЗжЦЌЖЮБъМЧЕФШЋГЄЕААзжЪбљЦЗЃЌ ЯджјМђЛЏКЫДХЙВеёВЈЦз. NakamuraЕШЃл46Ѓнгк1998ФъЪЙгУСЫРДздЙХОњPyrococcus furiosusЕФФкКЌыФЃЈPI-PfuIЃЉЃЌ ВЂНЋЦфШЫЮЊЗжЮЊ2ИіЦЌЖЮЃЌ ЗжБ№гызїЮЊКЫДХЙВеёБэеїЕФАаЕААзRNAОлКЯУИІСбЧЛљЕФCЖЫНсЙЙгђЃЈІСCЃЉЕФ2ИіЦЌЖЮШкКЯЃЈЭМ15ЃЉ. ОЭЌЮЛЫиБъМЧЕФЦЌЖЮгыСэвЛЮДБъМЧЦЌЖЮдкЬхЭтЛьКЯВЂЗЂЩњЗДЪНМєНгЗДгІЃЌ ЪзДЮЛёЕУСЫгУгкКЫДХЙВеёБэеїЕФВПЗжЦЌЖЮЭЌЮЛЫиБъМЧЕФЕААзжЪбљЦЗ.
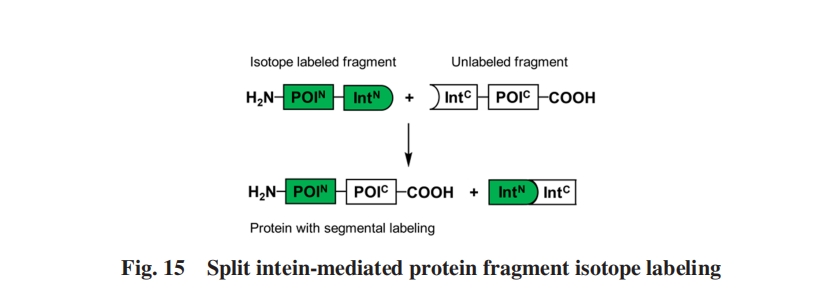
ОЁЙмзюГѕБъМЧЕФФПБъЕААзжЪНігадМ9000 DaЃЌ ЕЋИУЯЕЭГКѓРДГЩЙІЕигУгкжЦБИЗжЖЮЭЌЮЛЫиБъМЧЕФТѓбПЬЧНсКЯЕААзЃЈ42000 DaЃЉЃл97ЃнМАF0F1ATPУИІТбЧЛљЃЈ52000 DaЃЉЃл98ЃнЃЌ жЄУїСЫЭЈЙ§ЗжСбФкКЌыФЪЕЯжЕФЗжЖЮЭЌЮЛЫиБъМЧПЩгУгкИќДѓФПБъЕААзжЪЕФNMRНсЙЙЗжЮі.
IwaïЕШЃл12ЃнПЊЗЂСЫвЛжжЭЈЙ§Ш§ЦЌЖЮЕААзжЪМєНгРДЪЕЯжЦЌЖЮЭЌЮЛЫиБъМЧЕФаТВпТд. РћгУОпгаЯрЭЌађСаЕЋЗжСбЮЛЕуВЛЭЌЕФNpuDnaEЙЄГЬЛЏЗжСбФкКЌыФЃЌ ЖдРДздLyngbya majusculaЕФCuracin AЃЈCurAЃЉЕААзЭЈЙ§Ш§ЦЌЖЮЕААзМєНгРДЪЕЯжВПЗжЦЌЖЮЭЌЮЛЫиБъМЧ. ИУЕААзАќКЌ3ИіЫГађЕФѕЃЛљдиЬхЕААзЃЈACPЃЉНсЙЙгђ. ет 3ИіНсЙЙгђжЎМфИпЖШЕФађСаЭЌвЛадЃЈ93%~100%ЃЉЕМжТЦфЫљга3ИіНсЙЙгђОљБЛЭЌЮЛЫиБъМЧЕФбљЦЗЕФNMRаХКХбЯжижиЕў. вђДЫЃЌ бЁдёадЕиЖдИУШ§НсЙЙгђЕААзжЪЕФжааФНсЙЙгђНјааСЫЭЌЮЛЫиБъМЧЃЌ ЭЈЙ§ЗжСбФкКЌыФНщЕМЕФPTSЃЌ вддМ40%ЕФЪеТЪЛёШЁСЫгУгкNMRЗжЮіЕФВПЗжЦЌЖЮЭЌЮЛЫиБъМЧЕФШЋГЄCurAЕААз. ИУЗНЗЈНЋВПЗжЦЌЖЮЭЌЮЛЫиБъМЧВпТддкКЫДХЙВеёбаОПжаЕФгІгУЭиеЙЕНСЫЖрНсЙЙгђЕААзжаЕФНсЙЙгђ-НсЙЙгђЯрЛЅзїгУбаОП.
IwaïЕШЃл99ЃнЛЙПЊЗЂСЫНЋЭЌЮЛЫиБъМЧЕФЦЌЖЮгыЮДБъМЧЕФЦЌЖЮдкЭЌвЛЯИОњХрбјЮяжавЛДЮЩњГЩЃЌ ЫцКѓжБНгЗЂЩњМєНгЗДгІРДЬхФкЛёШЁЗжЖЮЭЌЮЛЫиБъМЧЕФШЋГЄЕААзЕФВпТд. ЦфЛљБОдРэЪЧдкЕЅИіЯИАћХрбјЙ§ГЬжаЕФВЛЭЌЪБМфБэДяКЌЗжСбФкКЌыФЕФЕААзЦЌЖЮЧАЬхЃЌ ВЂдкИїИіБэДяВНжшжЎМфИќЛЛЮДБъМЧЛђЭЌЮЛЫиБъМЧЬѕМўЕФХрбјЛљ. етбљЃЌ жЛгавЛжжЧАЬхЕААзЦЌЖЮЪЧБЛЭЌЮЛЫиБъМЧЕФЃЌ МєНгЗДгІКѓЕУЕНЕФМДЪЧжЛгавЛИіЦЌЖЮБЛЭЌЮЛЫиБъМЧЕФФПБъЕААзЃЌ ЪЕЯжСЫЗжЖЮЭЌЮЛЫиБъМЧЕААзжЪЕФжБНгЬхФкЛёШЁЃл98Ѓн.
4.3ЁЁЖрыФКЭЕААзжЪЕФЛЗЛЏ
ЛљгкЯИАћЕФЛЗыФЩИбЁЗНЗЈЯрЖдгкЪЩОњЬхеЙЪОЕШЬхЭтЩИбЁЗНЗЈЃЌ ПЩвдИќЮЊЮШНЁЕиЕУЕНОпгаЬхФкЛюадЕФЛЗыФЃЌ ВЂМцОпНЯЕЭЕФЯИАћЖОадЁЂ НЯКУЕФШмНтЖШЕШаджЪ. ЗжСбФкКЌыФНщЕМЕФPTSвђЦфЦЌЖЮПЩвджБНгЭЈЙ§БэДяЛёШЁВЂЧвПЩвдНЋСНВрЕФЖрыФЦЌЖЮСЌНгЦ№РДЕФЬиадЃЌ ПЩБЛгУРДдкЯИАћФкЩњГЩЛЗыФ. МДЭЈЙ§2ИіЗжСбФкКЌыФЦЌЖЮЕФНсКЯЃЌ ЦШЪЙЦфСНВрСЌНгЕФЯпадЖрыФЕФNЖЫКЭCЖЫЭЈЙ§ыФМќЭЗ-ЮВСЌНгЃЌ ЭъГЩЛЗЛЏЃЈЭМ16ЃЉ.
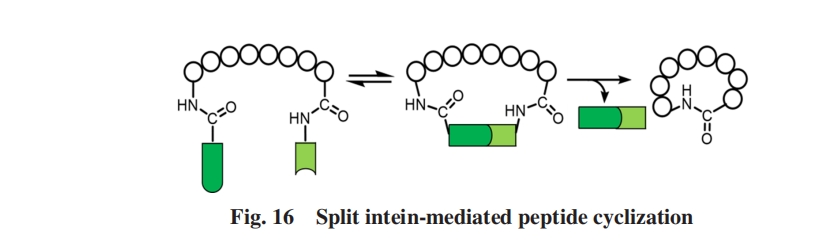
ЮЊДЫЃЌ BenkovicЕШЃл2ЃнПЊЗЂСЫЗжСбФкКЌыФНщЕМЕФЖрыФКЭЕААзжЪЛЗзДСЌНгЃЈSICLOPPSЃЉММЪѕ. етЯюММЪѕПЩвдЪЕЯжЬхФкЕФЛЗыФЮФПтЙЙНЈЃЌ ВЂНсКЯЙІФмЩИбЁЃЌ ЕУЕНСЫаэЖрОпгаЩњЮяЛюадЕФЛЗыФЃЌ ШчРвАБЫсУИвжжЦМСpseudostellarin FЃл2ЃнМАМзЛљзЊвЦУИвжжЦМСЕШЃл100Ѓн.
ЮЊСЫдіМгЩњГЩЛЗыФЕФЙІФмЛљЭХЖрбљадЃЌ SchultzЕШЃл101ЃнЭЈЙ§гывХДЋУмТыРЉеЙММЪѕНЋЗЧЬьШЛАБЛљЫсВхШыЕНЯЕЭГЩњГЩЕФЛЗыФжаЃЌ ЗсИЛСЫПЩЩњГЩЛЗыФЮФПтЕФЖрбљадЃЌ ЧвЩИбЁГіСЫHIVЕААзУИЛЗыФвжжЦМС. LindquistЕШЃл102ЃнЙЙНЈСЫЕквЛИіНЭФИМцШнЕФЛЗыФЮФПтЃЌ ВЂЩИбЁГіСЫПЩдкНЭФИМАЖЏЮяФЃаЭжаЬивьадНЕЕЭ ІС-synucleinЖОадЕФЛЗыФЗжзг.
ЮЊСЫПЊЗЂПЩвдЕїПиЕААзжЪ-ЕААзжЪЯрЛЅзїгУЕФЛЗыФЗжзгЃЌ BenkovicЕШЃл103ЃнећКЯСЫSICLOPPSгыЗДЯђЫЋдгНЛЯЕЭГЃЈRTHSЃЉ2жжММЪѕ. дкЫожїЯИАћжаОSICLOPPSММЪѕВњЩњЕФЛЗыФЮФПтЭЈЙ§RTHSЯЕЭГжБНгНјааЩИбЁЃЌ МјЖЈГіСЫЕїНкFKBP12-РзХСУЙЫи-FRAPЯрЛЅзїгУКЭHIV-1ЕААзУИКЭКЫЬЧКЫмеЫсЛЙдУИЯрЛЅзїгУЕФЛЗыФЗжзгЃЌ еЙЪОСЫSICLOPPSММЪѕЙЙНЈЛЗыФЮФПтВЂгУгкЛёШЁПЩЕїПиЕААзжЪ-ЕААзжЪЯрЛЅзїгУЕФЛЗыФЕФЧБСІ.
СэЭтЃЌ баОПепУЧЛЙПЊЗЂСЫНЋЗжСбФкКЌыФгыСЌајФкКЌыФСЊгУРДЙЙНЈЛЗыФЕФЗНЗЈЃл104ЃнЃЌ вдМАЭЈЙ§PTSаЮГЩыФМќЭъГЩыФЛЗЛЏКѓЃЌ дйРћгУАыызАБЫсВаЛљаЮГЩДѓЛЗыФФкВПЕФЖўСђМќРДЙЙНЈЫЋЛЗыФВњЮяЕФВпТдЃл105ЃнЃЌ етЬхЯжСЫЗжСбФкКЌыФЯрЙиЗНЗЈгУгкЙЙНЈЛЗыФРрЗжзгЕФСМКУаЇЙћМАЙуЗКгІгУ.
ЛљгкЗжСбФкКЌыФЕФЖрыФЛЗЛЏЛЙПЩгыЦфЫћаТаЫЩИбЁММЪѕНсКЯЃЌ ParkЕШЃл106ЃнПЊЗЂСЫвЛжжГЦЮЊCWCPSЃЈCustom-designed warhead-armed cyclic peptide screeningЃЉЕФВпТдЃЌ НЋПЩгыАаЕААзЛюадЮЛЕуНсКЯЕФЕЏЭЗЛЏбЇЛљЭХв§ШыгЩЗжСбФкКЌыФНщЕМЕФPTSЩњГЩЕФЛЗыФвжжЦМСЃЌ дйЭЈЙ§НЭФИЫЋдгНЛНјааЯИАћФкЩИбЁ. ЙЙЯѓЮШЖЈЕФЛЗыФжаЕФЕЏЭЗЛЏбЇЛљЭХгыЛюадЮЛЕуНсКЯКѓЕФЕФНтРыИќТ§ЃЌ ЧзКЭСІЯджјдіЧП. ЭЈЙ§ИУЗНЗЈЗЂЯжСЫвЛжжеыЖдАЉжЂАаЕуHDAC8гааЇЕФвжжЦМСCY5-6QЃЌ ЦфБэЯжГіКмЧПЕФНсКЯЧзКЭСІЃЈKD=15.1 nmol/LЃЉКЭвжжЦзїгУЃЈIC50=0.61 ІЬmol/LЃЉ. етБэУїЛљгкЗжСбФкКЌыФЕФЗНЗЈЪЧЯИАћФкЙЙНЈЛЗыФЕФвЛИіЯрЖдЮШНЁЕФВпТдЃЌ дкгаЧАЭОЕФЛЗыФвЉЮяЧАЬхЕФЗЂЯжжаЗЂЛгзїгУ.
4.4ЁЁЬѕМўадЕААзМєНг
ЬѕМўадЕААзМєНгЃЈCPSЃЉМДЭЈЙ§вЛИіЭтдДЕїНкЦїРДМЄЛюЛђвжжЦЕААзМєНг.ЛљгкЗжСбФкКЌыФЕФCPSЯЕЭГЕФЛљБОдРэЪЧНЋШЫЙЄЩшМЦЕФВЛФмздЗЂНсКЯЕФЗжСбФкКЌыФШкКЯЕНФПБъЕААзЕФ2ИіЦЌЖЮжаЃЌ ЭЈЙ§в§ШыЬиЖЈЭтдДЗжзгЛђЙтееЕШЗНЪНжиНЈЦфМєНгЛюадЃЛЛђЪЧдкЛюадЮЛЕуВаЛљДІЛђИННќНјааЛЏбЇаоЪЮЃЌ ЫцКѓЭЈЙ§ЙтгеЕМЁЂ УИЧаЕШЗНЪНШЅГ§аоЪЮЃЌ ДгЖјЪЙЦфЛжИДМєНгЛюад. вђДЫЃЌ ПЩвдЭЈЙ§ПижЦФкКЌыФЕФЛюадЃЌ РДЕУЕНвЛжжЫцвтЁАМЄЛюЁБШЮКЮЕААзжЪЕФЗНЗЈЃЌ ЩѕжСдкЬхФквВПЩвдЪЪгУ.
ШЫЙЄЗжСбЕФSce VMA ФкКЌыФдкIntNКЭ IntCЦЌЖЮжЎМфНіБэЯжГіЗЧГЃЕЭЕФЧзКЭСІЃЌ дкЕААзжЪМєНгжаМИКѕЮоЛюад. MuirЕШЃл107ЃнНЋРзХСУЙЫиНсКЯНсЙЙгђFKBP12КЭFRBШкКЯЕН2ИіЗжСбФкКЌыФЦЌЖЮКѓЃЌ МгШыРзХСУЙЫиЃЌ ПЩгеЕМ2ИіЦЌЖЮЯрЛЅППНќВЂНсКЯЃЌ НјЖјЕМжТЗжСбФкКЌыФЦЌЖЮЯрЛЅППНќВЂелЕўГЩОпгаPTSЛюадЕФИДКЯЮяЃлЭМ17ЃЈAЃЉЃн. ЭЈЙ§ЪЕЯжФЃаЭЕААзMBPКЭHis-tagЕФCPSЃЌ ЪзДЮЭъГЩСЫЕААзжЪЗДЪНМєНгЗДгІЕФаЁЗжзгМЄЛюЕїПи. MootzЕШЃл108ЃнРћгУРрЫЦЕФЫМТЗЃЌ НЋЗжСбФкКЌыФЦЌЖЮЗжБ№гыFKBP12ЕФF36MЭЛБфЬхШкКЯЃЌ ИУЭЛБфЬхЮоашЭтМгаЁЗжзгМДПЩЗЂЩњЭЌдДЖўОлЛЏЃЌ НјЖјНщЕМЗжСбФкКЌыФЗЂЩњPTS. ЖјДЫЪБдйМгШыРзХСУЙЫиЕШаЁЗжзгЃЌ ЛсЭЈЙ§гыИУНсЙЙгђНсКЯЖјзшЖЯЦфЖўОлЛЏЃЌ НјЖјЪЕЯжPTSЗДгІЕФЬѕМўадвжжЦЃЌ ЬсЙЉСЫвЛжжЧБдкЕФНЯЮЊЭЈгУЕФЪжЖЮгУгкЗвыКѓЕФЕААзжЪЙІФмЕїПи. Г§аЁЗжзгЯьгІЕФCPSЯЕЭГЭтЃЌ MuirЕШЃл109ЃнЛЙПЊЗЂСЫЙтЯьгІадCPSЬхЯЕЃЌ НЋдДгкФтФЯНцЕФЛсдкЙтееЯТЗЂЩњЖўОлЕФЙтУєЩЋЫиBЃЈPhyBЃЉгыЙтУєЩЋЫиЯрЛЅзїгУвђзг3ЃЈPIF3ЃЉгыSce VMAЗжСбФкКЌыФСНЖЫЦЌЖЮЗжБ№ШкКЯЃлЭМ17ЃЈBЃЉЃн. РрЫЦгкFKBP12гыFRBЖдРзХСУЙЫиЕФЯьгІЃЌ PhyBгыPIF3дкЙтееЯТЗЂЩњЖўОлЃЌ ЪЙЕУVMAЗжСбФкКЌыФвВЯрЛЅППНќВЂЛжИДМєНгЛюад. РћгУетвЛЬхЯЕЃЌ вдMBP-Flag tagзїЮЊФЃаЭМєНгВњЮядкНЭФИжаЪЕЯжСЫЙтЯьгІадCPS. ДЫЭтЃЌ SilverЕШЃл110ЃнЛЙНЋШЫЙЄЩшМЦЕФЛЅЯржЎМфОпгаНЯЧПЧзКЭСІЕФОэЧњТна§ЃЈCoiled coilЃЌ CCЃЉв§ШыVMAЗжСбФкКЌыФЃЌ ЪЕЯжСЫгЋЙтЫиУИЕФCPS.
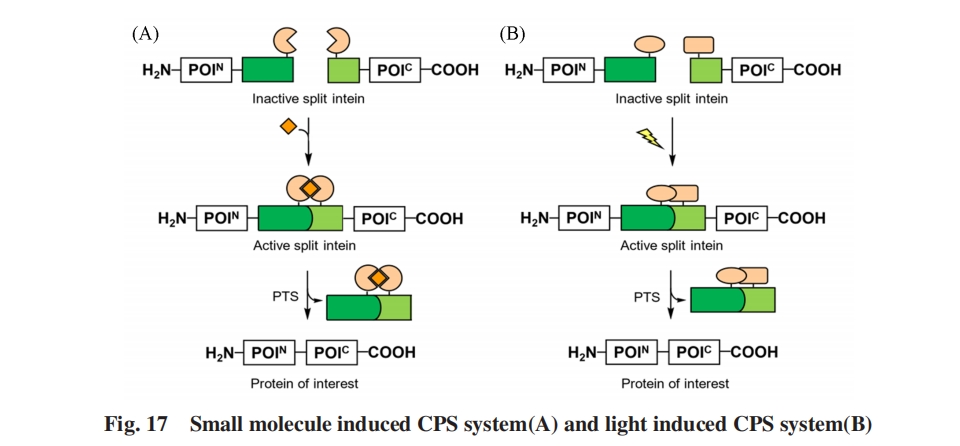
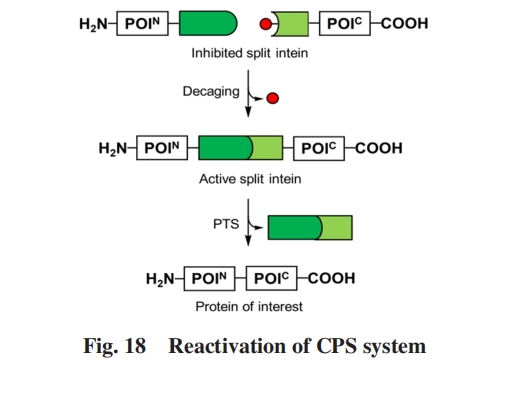
ЩЯЪіCPSЯЕЭГЖМЪЧЭЈЙ§НЋОпгадквЛЖЈЬѕМўЯТОпгаНЯИпНсКЯСІЕФЭтМгНсЙЙгђгыБОЩэНсКЯСІНЯЕЭЕФVMAЗжСбФкКЌыФШкКЯЪЕЯжЕФ. ЖјMuirЕШЃл111ЃнБЈЕРСЫЭЈЙ§в§ШыЦфЫќЛљЭХЃЌ вжжЦБОЩэОЭОпгаНЯИпНсКЯСІЕФЗжСбФкКЌыФЕФЛюадЃЌ дйдквЛЖЈЬѕМўЯТвЦГ§вжжЦЛљЭХЭъГЩЭбС§ЃЌ ДгЖјЪЕЯжCPSЕФаТВпТдЃЈЭМ18ЃЉ. КЯГЩСЫдкPTSЕФЙиМќЫПАБЫсВаЛљЩЯЭЈЙ§ІТ-єЧЛљаЮГЩЕФѕЅМќЖјЗЧГЃЙцЕФЭЈЙ§ІС-АБЛљЕФѕЃАЗМќСЌНгКѓајЦЌЖЮЕФSsp DnaEФкКЌыФЦЌЖЮРрЫЦЮяЃЌ ЧвЦфІС-АБЛљЛљЭХБЛЭтМгЕФБЃЛЄЛљЭХаоЪЮ. етзшЖЯСЫИУЦЌЖЮЕФO-NѕЃЛљзЊвЦЙ§ГЬЃЌ ЪЙЕУМєНгЗДгІЮоЗЈЗЂЩњ. БЃЛЄЛљЭХПЩвдЭЈЙ§зЯЭтЯпееЩфЛђЕААзУИМєЧаШЅГ§ЃЌ ДгЖјЛжИДИУЦЌЖЮЕФМєНгЗДгІЛюад. ЭЈЙ§жЦБИПЙОњыФmagaininЕФРрЫЦЮяеЙЪОСЫИУCPSЯЕЭГЕФЧБдкгІгУ. MootzЕШЃл112ЃнНЋЦљНёвбжЊМєНгЗДгІЫйТЪзюПьЕФgp41-1ЗжСбФкКЌыФNЖЫЦЌЖЮжаБЃЪиЕФБНБћАБЫсЭЈЙ§вХДЋУмТыРЉеЙММЪѕЭЛБфЮЊПЩЙтЛюЛЏЕФСкЯѕЛљмаЛљРвАБЫсЃлO-ЃЈ2-NitrobenzylЃЉ-L-tyrosineЃЌ ONBYЃн. дк365 nmЕФзЯЭтЯпееЩфЯТЃЌ СкЯѕЛљмаЛљБЃЛЄЛљБЛЭбГ§ЃЌ МєНгЗДгІвдгывАЩњаЭgp41-1ЗжСбФкКЌыФМИКѕЯрЭЌЕФЫйТЪЗЂЩњ. дНРДдНЖрЕФCPSВпТдЕФГіЯжЃЌ ЪЙЕУИУЗНЗЈгаЭћГЩЮЊПЩвддкЪБПеВуУцЩЯЕїПиЬхФкЕААзжЪЙІФмЕФЪжЖЮ.
5 змНсгыеЙЭћ
ОЙ§баОПепЕФГжајЬНЫїЃЌ ЗжСбФкКЌыФНщЕМЕФЕААзжЪЗДЪНМєНгвбОгЩвЛжжгаШЄЕФЕААзжЪЗвыКѓЯжЯѓЗЂеЙГЩЮЊвЛжжгІгУЙуЗКЕФИпаЇСЌНгЗНЗЈ. ЖдЕААзжЪЗДЪНМєНгЕФЛЏбЇЙ§ГЬЕФНтЮіЃЌ ЪЙШЫУЧИќКУЕиРэНтЗжСбФкКЌыФНсЙЙгыЙІФмжЎМфЕФЙиЯЕ. ПьЫйЗДЪНМєНгЗжСбФкКЌыФЕФЗЂЯжЁЂ ФкКЌыФЕФађСаНјЛЏЁЂ ФкКЌыФЮоКлМєНгВпТдЕФПЊЗЂЁЂ е§НЛФкКЌыФПтЕФНЈСЂЕШЃЌ ЖМДѓДѓИФНјСЫЛљгкФкКЌыФЕФСЌНгММЪѕЃЌ ЪЙЦфГЩЮЊШеНЅГЩЪьЕФЕААзжЪСЌНгЙЄОпЃЌ ЗжСбФкКЌыФНщЕМЕФЕААзжЪЗДЪНМєНгММЪѕЕФгІгУвбОХюВЊЗЂеЙ. ЗжСбФкКЌыФЯЕЭГДйНјСЫНсЙЙЩњЮябЇЕФЗЂеЙЃЌ ЬиБ№ЪЧКЫДХЙВеёВЈЦзНсЙЙНтЮіжаЕФгІгУЃЛЗжСбФкКЌыФЯЕЭГвВДйНјСЫКЯГЩЛђАыКЯГЩИќДѓЁЂ ИќИДдгЁЂ ДјгаЖржжаоЪЮЕФЕААзжЪЃЛДЫЭтЃЌ ЛљгкЗжСбФкКЌыФММЪѕЕФЛЗзДыФПтвВвбГЩЮЊвЉЮяЗЂЯжЕФгааЇВпТд.
ШЛЖјЃЌ ШЫУЧвВБиаыШЯЪЖЕНФПЧАЖдЗжСбФкКЌыФЕФРэНтКЭбаОПЩаВЛЭъЩЦ. ЫфШЛЖдгкЗжСбФкКЌыФНсКЯвдМАКѓајЗДгІЙ§ГЬвбОНјааСЫбаОПЃЌ ЕЋЪЧЖдгкЗДгІЙ§ГЬжаИќЯИжТЕФНсЙЙБфЙЙгыДпЛЏЙІФмБфЛЏЕФЗжЮіЛЙБШНЯДжВк. ИќКУЕиРэНтСђѕЅНЛЛЛЙ§ГЬжа2ИілЯЛљШчКЮППНќЃЌ жЇСДСђѕЅШчКЮЖдЬьЖЌѕЃАЗЛЗЛЏЙ§ГЬВњЩњДйНјЃЌ ЖМгажњгкИќКУЕиаЕїМєНгЗДгІЕФИїИіВНжшЕФЖЏСІбЇЃЌ ДгЖјЬсИпећЬхЕФСЌНгаЇТЪЃЌ БмУтC-ЖЫЛђN-ЖЫЫЎНтИБВњЮяЕФГіЯж. ДЫЭтЃЌ ФПЧАЖдЗжСбФкКЌыФдкЧПБфадЁЂ ИпЮТЁЂ бѕЛЏадЕШЬѕМўЯТЕФЮШНЁадбаОПЩаВЛГфзуЃЌ ПЩФмдкгыЦфЫќЛЏбЇЗДгІСЊгУЪБДјРДВЛШЗЖЈад.
ЫцзХИќЖрЕФЮЂЩњЮяЛљвђзщБЛВтађЃЌ ЗжСбФкКЌыФЕФЪ§СПНЋбИЫйдіГЄЃЌ ЮДРДгаЭћЗЂЯжОпгаЖРЬиаджЪЃЈР§ШчФмЙЛФЭЪмИпЮТЁЂ ИпбЮЕШМЋЖЫЬѕМўЃЉЕФаТаЭЗжСбФкКЌыФЃЌ ЗсИЛШЫРрЖдЗжСбФкКЌыФЕФШЯЪЖЃЌ НвЪОЯжНёЩаЮДШЯЪЖЕНЕФЧБСІЃЌ ЮЊЗжСбФкКЌыФММЪѕЕФЗЂеЙДјРДаТЕФЛњгіЃЌ ВЂзюжеЛёЕУЭЈгУаЭФкКЌыФЃЌ ФмЙЛЖдШЮКЮЭтЯдыФађСаОљОпгаМєПьЫйНгЖЏСІбЇЃЌ ВњТЪИпЃЌ ЗжСбЦЌЖЮвзгкЭЈЙ§ЛЏбЇКЯГЩММЪѕЛёШЁЃЌ ЧвСЌНгЛюадвзгкЭЈЙ§ЭтдДДЬМЄПижЦЕШ.
Утд№ЩљУїЃКБОЮФЮЊаавЕНЛСїбЇЯАЃЌАцШЈЙщдзїепМАддгжОЫљгаЃЌШчгаЧжШЈЃЌПЩСЊЯЕЩОГ§ЁЃЮФеТБъзЂгазїепМАЮФеТГіДІЃЌШчашдФЖСдЮФМАВЮПМЮФЯзЃЌПЩдФЖСддгжОЁЃ
