
’Σ“ΣΘΚΝΫ«Ή–‘ΕύκΡΖ÷Ή”ΨΏ”–άύΥΤΧλ»ΜΝΉ÷§Ζ÷Ή”ΒΡΝΫ«ΉΧΊ–‘ΓΔ ΖαΗΜΒΡΖ÷Ή”ΫαΙΙΓΔ ΕάΧΊ–¬”±ΒΡΉιΉΑΧεΫαΙΙ“‘ΦΑΧΊ βΒΡ…ζΈο―ßΙΠΡή, «ΕύκΡΉ‘ΉιΉΑ―–ΨΩΒΡ»»ΒψΝλ”ρ. ±ΨΈΡΉήΫαΝΥΫϋΡξά¥ΙΊ”ΎΝΫ«Ή–‘ΕύκΡΉ‘ΉιΉΑ―–ΨΩΦΑ”Π”ΟΒΡΫχ’Ι, Ϋι…ήΝΥΦΗ÷÷≥ΘΦϊΒΡΝΫ«Ή–‘ΕύκΡ, ≤ΔΫχ“Μ≤Ϋ≤ϊ ωΤδΖ÷Ή”ΫαΙΙΧΊ’ςΓΔ ΉιΉΑ––ΈΣΚΆΜζάμΓΔ ΉιΉΑΧεΫαΙΙΚΆΙΠΡή“‘ΦΑ‘ΎΡ…ΟΉΦΦ θΚΆ…ζΈο“Ϋ―ßΝλ”ρ÷–ΒΡ”Π”Ο.
ΕύκΡΉ‘ΉιΉΑΙψΖΚ¥φ‘Ύ”Ύ…ζΟϋΧε÷–, «÷ΎΕύ…ζΟϋΜνΕ·ΚΆ…ζΈο―ßΙΠΡήΒΟ“‘ Βœ÷ΒΡΜυ¥Γ. Ή‘ΉιΉΑ «ΕύκΡΖ÷Ή””κ…ζΨψά¥ΒΡΡήΝΠ, ΨΏ”–ΧΊΕ®Α±ΜυΥα–ρΝ–ΒΡΕύκΡΩ…“‘ΉιΉΑ–Έ≥…ΨΏ”–ΧΊΕ®Ω’ΦδΫαΙΙΚΆ…ζΈο―ßΙΠΡήΒΡΒΑΑΉ÷ Ζ÷Ή”. ΕχΕύκΡΦΑΒΑΑΉΒΡ¥μΈσΉιΉΑ≤ΜΫωΜα‘λ≥……ζΈο―ßΙΠΡήΒΡ…Ξ ßΜρΗΡ±δ, Εχ«“Μα≤ζ…ζ“ΜœΒΝ–ΒΡ≤ΓάμΖ¥”Π. “ρ¥Υ, Ε‘ΕύκΡΖ÷Ή”ΒΡΉ‘ΉιΉΑ―–ΨΩ≤ΜΫω”–άϊ”Ύ…ν»κΝΥΫβ…ζΟϋœ÷œσ, Εχ«“Ε‘”Ύ–μΕύ÷Ί¥σΦ≤≤ΓΒΡ≤ΓάμΦΑ÷ΈΝΤΖΫΖ®ΒΡ―–ΨΩ“≤ΨΏ”– °Ζ÷÷Ί“ΣΒΡ“β“ε. ΗϋΈΣ÷Ί“ΣΒΡ «, ΕύκΡΉςΈΣΧλ»ΜΒΡ”≈ΝΦΉιΉΑΖ÷Ή”, ΨΏ”–ΧΊ βΒΡ…ζΈοΜν–‘ΚΆΝΦΚΟΒΡ…ζΈοœύ»ί–‘, ΡήΙΜΗ≥”η≤ΡΝœΕάΧΊΒΡ…ζΈο―ßΙΠΡή. “Ή”ΎΚœ≥…ΚΆΜ·―ß–ό Έ“≤ «ΕύκΡΖ÷Ή”ΒΡΆΜ≥ω”≈ Τ, Ά®Ιΐ20÷÷Χλ»ΜΑ±ΜυΥαΒΡ≈≈Ν–ΉιΚœ“‘ΦΑΆβ‘¥–‘ΙΠΡήΜυΆ≈ΒΡ“ΐ»κ, Ω…“‘ΧαΙ© ΐ“‘“ΎΦΤΒΡΕύκΡΉιΉΑΖ÷Ή”, ¥”Εχ–Έ≥…ΚΤ»γ―ΧΚΘΒΡΫαΙΙΦΑΙΠΡήΗς“λΒΡΕύκΡΉιΉΑΧε. “≤ « Βœ÷ΓΑΉ‘œ¬Εχ…œΓ±÷Τ±ΗΡ…ΟΉ/―«ΈΔΟΉΫαΙΙΒΡ–¬–Ά…ζΈοΙΠΡή≤ΡΝœΒΡ“ΜΧθ÷Ί“ΣΆΨΨΕ. œύ±»”ΎΤδΥϋΉ‘ΉιΉΑΧεœΒ, ΕύκΡΉ‘ΉιΉΑ‘Ύœ»ΫχΙΠΡή≤ΡΝœ÷Τ±ΗΓΔ Μυ“ρ÷ΈΝΤΓΔ …ζΈοΩσΜ·ΓΔ …ζΈο¥ΪΗ–“‘ΦΑΉι÷·ΙΛ≥ΧΒ»…ζΈο“Ϋ―ßΝλ”ρΨΏ”–ΙψάΪΒΡ”Π”Ο«ΑΨΑ, ήΒΫ―–ΨΩ»Υ‘±ΒΡΙψΖΚ÷Ί ”[1].
‘ΎΕύκΡΉ‘ΉιΉΑ―–ΨΩ÷–, ΉςΈΣΕ―ΤωΡΘΩιΒΡΕύκΡΖ÷Ή”ΫαΙΙΦΑ…ηΦΤ÷ΝΙΊ÷Ί“Σ, «ΨωΕ®ΕύκΡΉ‘ΉιΉΑ––ΈΣΦΑΉιΉΑΧε–ΈΟ≤ΚΆΙΠΡήΒΡ÷Ί“Σ“ρΥΊ. ΝΫ«Ή–‘ΕύκΡΖ÷Ή”ΫαΙΙΨΏ”–άύΥΤ”Ύ±μΟφΜν–‘ΦΝΒΡ ηΥ°Ν¥ΕΈΚΆ«ΉΥ°ΕΥΜυ, ‘Ύ ηΥ°–‘Ν¥ΕΈΦδΒΡ ηΥ°–‘Ής”ΟΝΠΚΆ«ΉΥ°–‘ΕύκΡΝ¥ΕΈΦδΒΡ«βΦϋ–≠Ά§«ΐΕ·œ¬, ΕύκΡΖ÷Ή”ΡήΙΜΉ‘ΉιΉΑ–Έ≥…Ιφ’ϊ”––ρΒΡΡ…ΟΉ/ΈΔΟΉΫαΙΙ. ‘ΎΉιΉΑΙΐ≥Χ÷–, ηΥ°–‘Ν¥ΕΈ”…”Ύ‘ςΥ°ΕχœύΜΞΨέΦ·, –Έ≥…ΉιΉΑΧεΒΡ ηΥ°–‘ΡΎΚΥ, Υυ≤ζ…ζΒΡ ηΥ°–‘Ής”ΟΝΠ ««ΐΕ·ΝΫ«Ή–‘Ζ÷Ή”Ή‘ΉιΉΑΚΆΈ§≥÷ΉιΉΑΧεΫαΙΙΈ»Ε®ΒΡ÷ς“Σ«ΐΕ·ΝΠ; ΕχκΡΝ¥ΦδΒΡ«βΦϋ‘ρ ΙΒΟœύΜΞΩΩΫϋΒΡ«ΉΥ°–‘ΕύκΡΝ¥ΕΈ“‘Ρ≥÷÷ΧΊΕ®ΒΡΕΰΦΕΫαΙΙΫτΟήΕ―Τω≈≈Ν–‘ΎΉιΉΑΧε±μΟφ, ”κΥ°Ϋ”¥Ξ, Ϋχ“Μ≤Ϋ«ΐΕ·ΉιΉΑΒΡΆξ≥…. ΝΫ«Ή–‘ΕύκΡΖ÷Ή”÷÷άύΖ±Εύ, Α¥ ηΥ°Ν¥ΕΈΒΡΙΙ≥…Ω…“‘Ζ÷ΈΣ ηΥ°Ν¥ΕΈ»Ϊ≤Ω”…Α±ΜυΥα≤–ΜυΙΙ≥…ΒΡ±μΟφΜν–‘κΡ(Surfactant-like peptide)ΚΆ ηΥ°Ν¥ΈΣ≥ΛΆιΜυΧΦΝ¥ΒΡΝΫ«Ή–‘ΕύκΡ(Peptide amphiphiles, PAs), ΕχΚσ’Ώ”÷Ω…“‘Α¥’’Ζ÷Ή”ΫαΙΙΒΡ≤ΜΆ§Ζ÷ΈΣ¥ΪΆ≥ΒΡΝΫ«Ή–‘ΕύκΡΚΆΫαΙΙΧΊ βΒΡΝΫ«Ή–‘ΕύκΡ, »γBola–ΆΝΫ«Ή–‘ΕύκΡΚΆGemini–ΆΝΫ«Ή–‘ΕύκΡΒ».
1 ”…Α±ΜυΥα≤–ΜυΙΙ≥… ηΥ°Ν¥ΕΈΒΡ±μΟφΜν–‘κΡΉ‘ΉιΉΑ
±μΟφΜν–‘κΡΒΡ ηΥ°Ν¥ΕΈ“ΜΑψ”…ΕύΗωΝ§–χΒΡ ηΥ°–‘Α±ΜυΥα≤–ΜυΉι≥…, «ΉΥ°Ν¥ΕΈΚ§”–1ΗωΜρ2Ηω¥χ”–1Μρ2Ηω’ΐΒγΚ…ΜρΗΚΒγΚ…ΒΡΑ±ΜυΥα≤–Μυ, ΫαΙΙΚΆ≥Ώ¥γ”κΧλ»ΜΝΉ÷§Ζ«≥ΘœύΥΤ[2]. ZhangΒ»[3~6]…ηΦΤΚœ≥…ΝΥ“ΜœΒΝ––ΓΖ÷Ή”±μΟφΜν–‘κΡ, »γ¥χ”–ΗΚΒγΚ…ΒΡDA6, DV6, D2V6, D2L6[3]ΚΆ D[4]2G4~10“‘ΦΑΒγ’ΐ–‘ΒΡKA6, H2V6, K2V6ΚΆK2L6[5]. ’β–©–ΓΖ÷Ή”±μΟφΜν–‘κΡ‘ΎΥ°÷–Ω…“‘ΉιΉΑ–Έ≥…”…Ρ…ΟΉΙήΙΙ≥…ΒΡΆχΉ¥ΫαΙΙ, Ρ…ΟΉΙή÷ήΈß¥φ‘Ύ”–Ρ…ΟΉΡ“≈ί; Εχ«ΉΥ°Ν¥ΕΈΒγΚ…ΒΡ’ΐΗΚ±δΜ·Μα”ΑœλΡ…ΟΉΙήΒΡ÷±ΨΕ. ΉιΉΑΜζάμ―–ΨΩ±μΟς, ΉιΉΑΧε÷–ΕύκΡΖ÷Ή”≤ΔΟΜ”–Α¥’’Ος»ΖΒΡΕΰΦΕΫαΙΙ≈≈Ν–, ΥΒΟς¥ΥάύΕύκΡΒΡΉιΉΑ÷ς“Σ“άΩΩ ηΥ°ΦδΉς”ΟΝΠΒΡ«ΐΕ·. ηΥ°Ν¥ΕΈΨέΦ·”ΎΉιΉΑΧεΡΎ≤Ω–Έ≥…ΥΪΖ÷Ή”≤ψ, Εχ«ΉΥ°Ν¥ΕΈΖΔ…ζΒγάκ”κΥ°Ζ÷Ή”–Έ≥…Υ°Μ·≤ψ, ΫϋΥΤ”ΎΧλ»ΜΝΉ÷§ΒΡΉιΉΑΖΫ Ϋ(ΆΦ1). ‘Ύ¥ΥΜυ¥Γ…œ, Ε‘ΕύκΡΫαΙΙΫχ––ΝΥ”≈Μ·, ΖΔœ÷“‘ΝΉΥαΜ·ΒΡSer≤–ΜυΈΣ«ΉΥ°Ν¥ΕΈ, A6ΜρV6ΈΣ ηΥ°Ν¥ΕΈ ±, ΕύκΡΒΡΉιΉΑ––ΈΣΗϋΫ”ΫϋΧλ»ΜΝΉ÷§[6]. ΕχΫΪ¥χ”–œύΖ¥ΒγΚ…ΒΡA6DΚΆA6KΙ≤Μλ ±, ΥφΉ≈A6D/A6KΜλΚœΡΠΕϊ±»¥”1ΓΟ1‘ωΦ”ΒΫ2ΓΟ1 ±, ΉιΉΑΧε¥”Ρ…ΟΉœΏΉΣ±δΈΣΡΛΫαΙΙ[7].
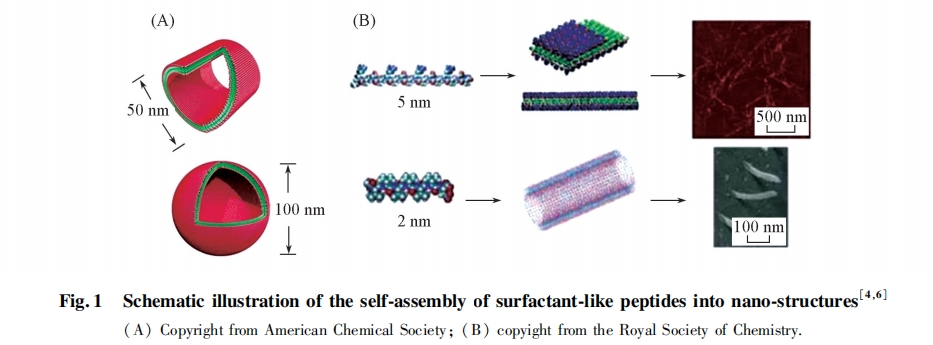
Ά®ΙΐΗΡ±δΑ±ΜυΥα≤–Μυ÷÷άύΚΆ ΐΡΩ, Ω…“‘ΒςΫΎΕύκΡΖ÷Ή”ΒΡΦΗΚΈΙΙ–Ά, ¥”Εχ”ΑœλΕύκΡΖ÷Ή”ΒΡΉιΉΑ––ΈΣΚΆΉν÷’ΉιΉΑΧε–ΈΟ≤. »γ“ΜΗωΖ«≥ΘΕΧΒΡI3KκΡΉιΉΑΒΡΥΪΖ÷Ή”≤ψ«ψœρ–Έ≥…Ψμ«ζΒΡ¬ί–ΐ¥χΉ¥ΫαΙΙ, ≤ΔΫχ“Μ≤Ϋ–Έ≥…÷±ΨΕ÷Μ”–~10 nmΒΡΙήΉ¥ΫαΙΙ[8]. XuΚΆLuΒ»[9]Ά®ΙΐΗΡ±δAmKΕύκΡ÷–Ala≤–ΜυΒΡ ΐΡΩ(m=3,6,9)ά¥ΗΡ±δΕύκΡΖ÷Ή”ΒΡΦΗΚΈΙΙ–Ά.
”…”Ύ¥Υάύ±μΟφΜν–‘κΡΒΡΫαΙΙ”κΧλ»ΜΝΉ÷§Ζ÷Ή”Ζ«≥ΘœύΥΤ, “ρ¥ΥΤδΉιΉΑ–Έ≥…ΒΡΥΪΖ÷Ή”≤ψΡΛΩ…“‘ΡΘΡβΧλ»ΜœΗΑϊΡΛΒΡΙΠΡή, ΦΪΨΏ”Π”Ο«±ΝΠ. »γ±μΟφΜν–‘κΡV6D, A6D, A6DΚΆV6KΩ…“‘ΦΪ¥σΒΊΧαΗΏΡΛΒΑΑΉΒΡΜν–‘ΚΆΈ»Ε®–‘[7], ”≈”Ύ¥ΪΆ≥ΒΡ±μΟφΜν–‘ΦΝN- °ΕΰΆιΜυ-D-¬σ―ΩΧ«ή’ΚΆN-–ΝΜυ-D-ΤœΧ«ή’[10~12]. ¥χ”–’ΐΒγΚ…ΒΡ±μΟφΜν–‘κΡΜΙΩ…“‘ΉςΈΣΩΙΨζ“©Έο Ι”Ο, ΥϋΩ…“‘Ά®Ιΐ”κœΗΨζœΗΑϊΡΛΝΉ÷§œύΜΞΉς”ΟΤΤΜΒœΗΨζΒΡœΗΑϊΡΛΫαΙΙ, ¥”Εχ…±ΥάœΗΨζ. »γ±μΟφΜν–‘κΡAmK(m=3, 6, 9)Ά®Ιΐ…χ»κΚΆΤΤΜΒœΗΨζœΗΑϊΡΛ±μœ÷≥ωΦΪΗΏΒΡΩΙΨζΜν–‘, «“ΩΙΨζΜν–‘ΥφΉ≈ ηΥ°Ν¥ΕΈΒΡΑ±ΜυΥα≤–ΜυAla ΐΡΩ‘ωΕύΕχ‘ω«Ω. Τδ÷–, A9KΕ‘ΗοάΦ œ“θ–‘ΨζΚΆΗοάΦ œ―τ–‘ΨζΕΦ”–…±ΟπΉς”Ο[13].
2 ηΥ°Ν¥ΕΈΈΣ≥ΛΆιΜυΝ¥ΒΡΝΫ«Ή–‘ΕύκΡΉ‘ΉιΉΑ
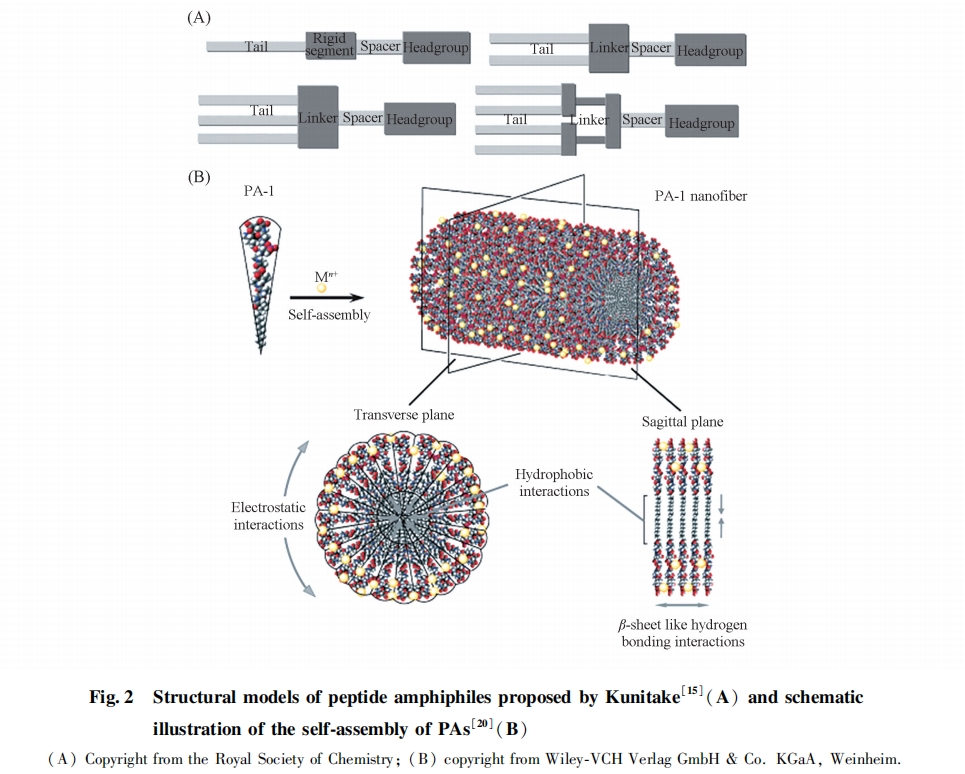
ηΥ°Ν¥ΕΈΈΣ÷§ΖΨΉε≥ΛΆιΜυΝ¥ΓΔ «ΉΥ°Ν¥ΕΈΈΣ«ΉΥ°–‘Α±ΜυΥα≤–ΜυΙΙ≥…ΒΡΝΫ«Ή–‘ΕύκΡΖ÷Ή”(PAs) «“ΜάύΉνΈΣ÷Ί“ΣΒΡΉιΉΑΕύκΡ[14], Ήν‘γ”…KunitakeΒ»[15]Χα≥ω, ΑϋΚ§”…1ΧθΜρΕύΧθ≥ΛΆιΜυΝ¥ΙΙ≥…ΒΡ ηΥ°Έ≤≤Ω“‘ΦΑ”…Α±ΜυΥα≤–ΜυΙΙ≥…ΒΡΝ§Ϋ”«χΓΔ Ζ÷Ητ«χΚΆ«ΉΥ°ΆΖ≤Ω[ΆΦ2(A)]. ≥ΛΆιΜυΝ¥÷°ΦδΒΡ ηΥ°Ής”ΟΝΠ“‘ΦΑΕύκΡΝ¥ΕΈΦδΒΡ«βΦϋΉς”Ο ««ΐΕ·¥ΥάύΕύκΡΖ÷Ή”ΉιΉΑΚΆΈ§≥÷ΉιΉΑΧεΫαΙΙΈ»Ε®ΒΡ÷Ί“Σ“ρΥΊ. StuppΒ»[16~23]…ηΦΤΝΥ“ΜœΒΝ–÷Ί“ΣΒΡPAsΖ÷Ή”, ≤…”Ο“ΜΕΈΠ¬-’έΒΰ–ρΝ–ΉςΈΣΝ§Ϋ”«χ, Εχ«ΉΥ°ΆΖ≤Ω‘ρ”…¥χΒγΒΡ«ΉΥ°–‘Α±ΜυΥα≤–ΜυΚΆΨΏ”–…ζΈοΜν–‘ΙΠΡήΒΡΕύκΡ–ρΝ–ΙΙ≥…[19]. PAsΖ÷Ή”‘ΎΥ°»ή“Κ÷–ΉιΉΑ ±, ηΥ°ΒΡ≥ΛΆιΜυΝ¥«ψœρ”Ύ‘ΎΉιΉΑΧεΡΎ≤ΩΨέΦ·–Έ≥… ηΥ°–‘ΡΎΚΥ, ΕχΑ±ΜυΥαΝ¥ΕΈ‘ρ“‘Π¬-’έΒΰΒΡΕΰΦΕΫαΙΙΕ―Τω≈≈Ν–‘ΎΉιΉΑΧε±μΟφ–Έ≥…«ΉΥ°–‘ΆβΩ«, ΫχΕχ–Έ≥…ΨΏ”–Ρ…ΟΉœΥΈ§ΫαΙΙΒΡΉιΉΑΧε. ”…”ΎΉιΉΑ–Έ≥…ΒΡœΥΈ§ΨΏ”–άύΥΤ”Ύ¥ΪΆ≥ΫΚ χΒΡΫαΙΙ, “ρ¥Υ“≤≥Τ÷°ΈΣœΥΈ§Ή¥ΫΚ χ[ΆΦ2(B)][20]. ”…”Ύ ηΥ°Ής”ΟΝΠ‘ΎPAsΉιΉΑΙΐ≥Χ÷–ΖΔΜ”Ή≈÷Ί“ΣΉς”Ο, “ρ¥ΥΆιΜυΝ¥≥ΛΕ»Ε‘ΤδΉιΉΑ”ΑœλΫœ¥σ. Β±ΆιΜυΝ¥Κ§ΧΦ ΐ≥§Ιΐ10 ±, Κ§”–C4G3SPRGDΕύκΡ–ρΝ–ΒΡPAsΖ÷Ή”‘ΎΥα–‘»ή“Κ÷–Ω…Ή‘ΉιΉΑ–Έ≥…”…Ρ…ΟΉœΥΈ§≤χ»ΤΕχ≥…ΒΡΥ°ΡΐΫΚ; ΕχΒ±ΆιΜυΝ¥Κ§ΧΦ ΐ–Γ”Ύ10 ±, ”…”ΎΆιΜυΝ¥ΦδΒΡ ηΥ°Ής”ΟΝΠΦθ»θ, PAsΖ÷Ή”≤ΜΡήΫχ––ΉιΉΑ[17]. ¥ΥΆβ, Ά®ΙΐΗΡ±δ«ΉΥ°ΕύκΡΝ¥ΕΈΦδΒΡ«βΦϋΉς”ΟΩ…“‘ΒςΩΊΉιΉΑΧεΫαΙΙΚΆΉιΉΑ––ΈΣ. ”…”ΎΦΪ–‘Α±ΜυΥα≤–ΜυΒΡ¥χΒγΉ¥Χ§ΡήΙΜ¥ΌΫχΜρΤΤΜΒΕύκΡΝ¥ΕΈΦδ«βΦϋΒΡ–Έ≥…, “ρ¥Υ, «ΉΥ°ΕύκΡΝ¥ΕΈ÷–Κ§”–¥χΒγΦΪ–‘Α±ΜυΥα≤–ΜυΒΡPAsΖ÷Ή”ΒΡΉιΉΑΕύΨΏ”–pHΟτΗ––‘[16,17,19]ΚΆΫπ τάκΉ”ΟτΗ––‘[20,21], «“≤ΜΆ§¥χΒγ–‘ΒΡPAsΖ÷Ή”Ε‘≤ΜΆ§Ϋπ τάκΉ”ΒΡΟτΗ––‘≤ΜΆ§. Ϋω”–ΗΚΒγ–‘Ϋœ»θΒΡPA7ΚΆ’ΐΒγ–‘ΒΡPA8Ε‘ΗΏ≈®Ε»ΒΡK+ΟτΗ–, ΡήΙΜ–Έ≥…ΡΐΫΚ; ΕχΨχ¥σΕύ ΐΒΡPAsΡΐΫΚΕ‘Mn2+, Ca2+, Cu2+, Gd3+Β»ΕΰΦέΜρ»ΐΦέΫπ τάκΉ”ΟτΗ–. ‘Ύ¥ΥΜυ¥Γ…œ, ΥϊΟ«Ϋχ“Μ≤Ϋάϊ”ΟΦΤΥψΡΘΡβ“‘ΦΑ Β―ιΙέ≤λΒΡΖΫΖ®Μφ÷ΤΝΥPAsΖ÷Ή”ΉιΉΑ±δΜ·ΒΡΓΑœύΆΦΓ±, Χ÷¬έΝΥPAsΖ÷Ή”‘ΎΥ°÷–Ή‘ΉιΉΑ––ΈΣΦΑΉιΉΑΧεΫαΙΙ±δΜ·”κΤδΖ÷Ή”ΫαΙΙΓΔ Ζ÷Ή”ΦδΉς”ΟΝΠΓΔ ―ΈάκΉ”≈®Ε»ΦΑ»ή“ΚpH÷Β÷°ΦδΒΡΙΊœΒ[18,22]. ΫϋΡξά¥, StuppΒ»[23]ΫΪ2÷÷¥χ”–œύΖ¥ΒγΚ…ΒΡPAsΖ÷Ή”‘ΎΥ°»ή“Κ÷–ΜλΚœ, άϊ”ΟΝΫ’ΏΦδΒΡΨ≤ΒγΈϋ“ΐΉς”ΟΉιΉΑ–Έ≥…Ρ…ΟΉœΥΈ§ΫαΙΙ, “ρ¥Υ’β÷÷ΉιΉΑΖΫΖ®”–Άϊ Βœ÷PAs‘Ύ…ζάμΧθΦΰœ¬ΒΡΉ‘ΉιΉΑ.
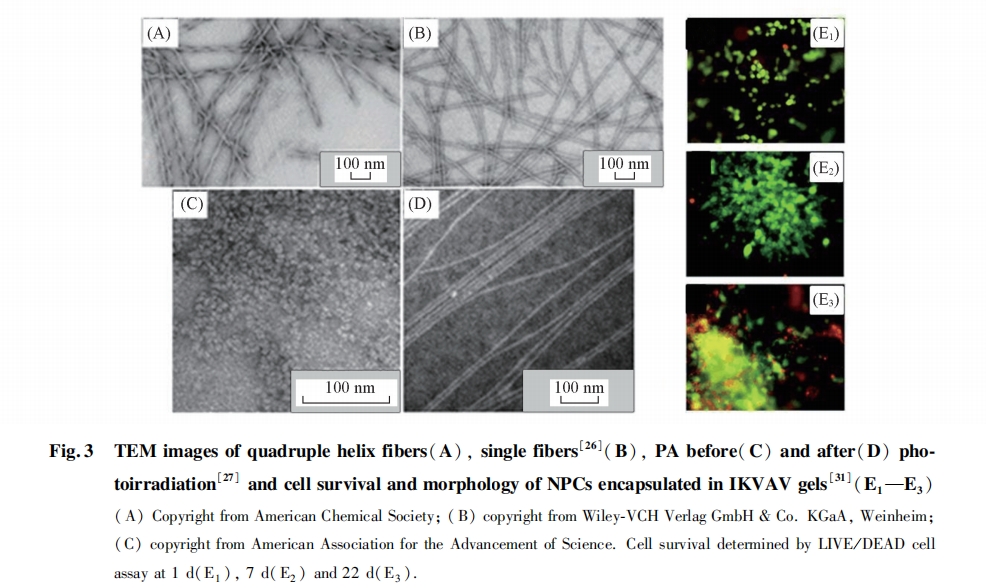
άϊ”Ο…œ ωPAsΕύκΡΖ÷Ή”ΕάΧΊΒΡΉ‘ΉιΉΑ–‘÷ , Ά®Ιΐ‘ΎΖ÷Ή”ΫαΙΙ÷–“ΐ»κΧΊ βΒΡΙΠΡήΜυΆ≈ΚΆΨΏ”–ΧΊΕ®…ζΈοΜν–‘ΒΡΙΠΡήΕύκΡ–ρΝ–, ¥”Εχ ΙPAsΒΡΉ‘ΉιΉΑ––ΈΣΓΔ ΉιΉΑΧεΫαΙΙΚΆΈοάμΜ·―ß–‘÷ ΖΔ…ζΗΡ±δ, Η≥”ηPAsΉιΉΑ≤ΡΝœΗϋΕύΒΡ”Π”ΟΦέ÷Β. StuppΒ»[24] ΫΪ4ΗωΝ§–χΒΡCys≤–Μυ“ΐ»κΒΫC16-C4G3SPRGDΕύκΡΖ÷Ή”ΫαΙΙ÷–, ‘ΎΥα–‘ΧθΦΰœ¬ΉιΉΑ–Έ≥…œΥΈ§Κσ, Cys≤–ΜυΚ§”–ΒΡέœΜυ‘ΎI2ΒΡΉς”Οœ¬ΖΔ…ζ―θΜ·ΫΜΝΣ. ’β÷÷‘ΎΉιΉΑΧεΡΎ≤Ω“ΐ»κΜ·―ßΫΜΝΣΒΡΖΫΖ®”––ßΧαΗΏΝΥœΥΈ§ΒΡ«ΩΕ», ΫΜΝΣΚσΒΡœΥΈ§‘ΎΦν–‘»ή“Κ÷–»‘ΡήΈ»Ε®¥φ‘Ύ, ΕχΈ¥Μ·―ßΫΜΝΣΒΡœΥΈ§‘ΎΦν–‘»ή“Κ÷–―ΗΥΌΖ÷Ϋβ. ”…Μ·―ßΫΜΝΣΒΡœΥΈ§–Έ≥…ΒΡΆχ¬γΫαΙΙΡήΙΜΡΘΡβœΗΑϊΆβΜυ÷ ΫαΙΙ, Ά®ΙΐΝΉΥαΜ·ΒΡSer≤–ΜυΕ‘Ca2+ΒΡΈϋΗΫ≤ζ…ζ…ζΈοΩσΜ·ΙΠΡή, –Έ≥…ΒΡΩσΜ·ΈοΖ«≥ΘΫ”ΫϋΧλ»Μτ«ΜυΝΉΜ“ ·ΫαΙΙ≥…Ζ÷. ΗΜΚ§ΉιΑ±Υα≤–Μυ–ρΝ–ΒΡΕύκΡΖ÷Ή”C16-A4G3H3ΉιΉΑ–Έ≥…ΒΡΡ…ΟΉœΥΈ§…œΨΏ”–ΡήΙΜ”κFe2+ΚΆFe3+άκΉ”ΫαΚœΒΡΜν–‘ΈΜΒψ, “‘¥ΥΈΣΡΘΑεΩ…“‘ΩσΜ·–Έ≥…¥≈–‘Ρ…ΟΉ≤ΡΝœ[25]. ΫΪΙβΟτ–‘ΒΡΝΎœθΜυ±ΫΜ·ΚœΈο“ΐ»κC16-GV3A3E3ΕύκΡΖ÷Ή”ΒΡ≤ύΝ¥…œ, ”…”ΎΝΎœθΜυ±Ϋ≤ύΝ¥ΜυΆ≈ΒΡΩ’ΦδΈΜΉη ΙΒΟPAsΖ÷Ή”ΡήΙΜΉιΉΑ–Έ≥…¬ί–ΐΉ¥œΥΈ§ΫαΙΙ, ≤ΔœύΜΞ≤χ»Τ–Έ≥…ΗϋΗΏΦΕΒΡΕΰ÷ΊΓΔ »ΐ÷ΊΚΆΥΡ÷Ί¬ί–ΐΫαΙΙ. ‘ΎΉœΆβΙβ’’…δΚσ, ≤ύΝ¥ΝΎœθΜυ±ΫΜυΆ≈ΖΔ…ζΙβΫβ, ¬ί–ΐΉ¥œΥΈ§ΫαΙΙΥφ÷°ΉΣ±δΈΣΤ’Ά®ΒΡΒΞΙ…Ρ…ΟΉœΥΈ§[ΆΦ3(A, B)][26]. ΫΪΝΎœθΜυ±ΫΜυΜ·―ßΝ§Ϋ”ΒΫC16-GV2E2RGDSΖ÷Ή”…œΫχ“Μ≤Ϋ÷Τ±Η≥ωΙβΩΊ¥ΞΖΔΒΡΥ°ΡΐΫΚ≤ΡΝœ. ¥χ”–ΝΎœθΜυ±ΫΒΡPAsΖ÷Ή”‘ΎΥ°÷–Ω…“‘ΉιΉΑ≥…«ρ–ΈΡ…ΟΉΫαΙΙ, Β±ΉœΆβΙβ’’…δΚσ, ΙβΫβΆ―¬δΝΎœθΜυ±ΫΜυΆ≈ΒΡPAsΖ÷Ή”ΉιΉΑ≥…ΨΏ”–Π¬-’έΒΰΫαΙΙΒΡΡ…ΟΉœΥΈ§, ΫχΕχœύΜΞ≤χ»Τ–Έ≥…Υ°ΡΐΫΚ. ”…”ΎPAsΖ÷Ή”Κ§”–œΗΑϊπΛΗΫΙΠΡήκΡΕΈRGDS, Ιβ’’Κσ–Έ≥…ΒΡΥ°ΡΐΫΚΡήΙΜ”––ߥΌΫχNIH3T3œΗΑϊΡΛΙ«ΦήΒΑΑΉΒΡmRNA±μ¥ο[ΆΦ3(C, D)][27]. ΤδΥϋΚ§”–RGDΙΠΡή–ρΝ–ΒΡPAsΖ÷Ή”ΉιΉΑ–Έ≥…ΒΡΥ°ΡΐΫΚ“≤Ά§―υΩ…“‘ΉςΈΣ”≈ΝΦΒΡœΗΑϊ≈ύ―χ÷ßΦή≤ΡΝœ[20,28,29], ≤ΔΩ…”Ο”Ύ÷Τ±ΗΧεΡΎΉΔ…δ≥…–ΆΥ°ΡΐΫΚ≤ΡΝœ, ”κœΥΈ§œΗΑϊ…ζ≥Λ“ρΉ”ΜλΚœΚσ”Ο”ΎΉι÷·–όΗ¥[30]. Κ§”–¥Ό…ώΨ≠ΆΜ…ζ≥ΛΒΡLeu-Lys-Val-Ala-ValΙΠΡή–ρΝ–ΒΡPAsΖ÷Ή”Ω…“‘‘ΎœΗΑϊ≈ύ―χ“Κ÷–‘ΎCa2+¥ΞΖΔœ¬ΉιΉΑ–Έ≥…ΨΏ”–Ρ…ΟΉœΥΈ§ΫαΙΙΒΡΥ°ΡΐΫΚ, άϊ”ΟΗΟΥ°ΡΐΫΚ≈ύ―χΚΆ”’ΒΦ…ώΨ≠‘ΣΗ…œΗΑϊΫχ––―Γ‘ώ–‘Ζ÷Μ·[ΆΦ3(E1~E3)][31]. ¥ΥΆβ, HartgerinkΒ»[32]ΫΪΕ‘œΗΑϊΜυ÷ Ϋπ τΒΑΑΉΟΗΟτΗ–ΒΡΕύκΡ–ρΝ–GTAGLIGQ“ΐ»κΒΫPAsΫαΙΙ÷–, ΉιΉΑ–Έ≥…ΒΡΥ°ΡΐΫΚΨΏ”–ΟΗΟτΗ––‘, ΡήΙΜ‘ΎœΗΑϊΖ÷ΟΎΒΡΔτ–ΆΫΚ‘≠ΟΗΒΡΉς”Οœ¬ΖΔ…ζΫΒΫβ, Ε‘”Ύ»ΥΧεΡΎΒΡ”Π”ΟΨΏ”–Ϋœ¥σ«±ΝΠ. ΉιΉΑΖΫΖ®ΒΡΗΡ±δΆ§―υΩ…“‘Ε‘PAsΒΡΉιΉΑΫχ––ΒςΩΊ, ≤ΔΦΪ¥σΒΊΆΊ’ΙΝΥΤδ…ζΈο―ß”Π”Ο. ΫΪΕύκΡC16-V3A3E3πΛ»ή“Κά≠≥…≥ΛΥΩΉ¥ΤΧ‘Ύ±μΟφΆΩ”–»»CaCl2»ή“ΚΒΡΤΫΑε…œ, ≤Δ‘Ύ80 Γφœ¬±ΘΈ¬30 minΚσΖΔœ÷, –Έ≥…ΒΡΧθΉ¥ΡΐΫΚΡΎ≤ΩΡ…ΟΉœΥΈ§≥ ΗΏΕ»”––ρΒΡΕ®œρ≈≈Ν–Ή¥Χ§, ≤ΜΆ§”ΎC16-V3A3E3‘ΎΥ°»ή“Κ÷––Έ≥…ΈόΙφ≤χ»ΤΒΡœΥΈ§. ΆΤ≤β‘ΎΗΏΈ¬œ¬±Γ≤ψ÷–ΒΡCa2+Ε‘±μΟφ¥χ”–ΗΚΒγΚ…ΒΡœΥΈ§≤ζ…ζΝΥΨ≤ΒγΉς”Ο, ”’ΒΦœΥΈ§≥ Ε®œρ”––ρ≈≈Ν–. Ϋχ“Μ≤Ϋ―–ΨΩΜΙΖΔœ÷, œΗΑϊΩ…“‘‘ΎΕ®œρ≈≈Ν–ΒΡœΥΈ§ χ…œΚήΚΟΒΊπΛΗΫ…ζ≥Λ, ’βΩ…Ρή «“ρΈΣΕ®œρ≈≈Ν–ΒΡœΥΈ§ χ–Έ≥…ΒΡΜΖΨ≥”κΧεΡΎΒΡœΗΑϊΆβΜυ÷ ΜΖΨ≥œύΥΤ, Ηϋ ”ΎœΗΑϊ…ζ≥Λ[33].
≥ΐΝΥ…œ ωΑϋΚ§≤ΜΆ§ΙΠΡήΝ¥ΕΈΒΡPAsΖ÷Ή”Άβ, ÷±Ϋ”ΫΪ≥ΛΆιΜυΝ¥”κΡ≥“ΜΕύκΡΝ¥ΕΈœύΝ§÷Τ±ΗΒΡΫαΙΙΗϋΈΣΦρΒΞΒΡPAsΖ÷Ή”“≤ΨΏ”–ΉιΉΑΡήΝΠ, ≤ΔΡήΙΜ–Έ≥…ΗϋΖαΗΜΒΡΉιΉΑΧε–ΈΟ≤. StuppΒ»[34]ΫΪC16ΆιΜυΝ¥”κΕΧκΡVEVEœύΝ§Ϋ”÷Τ±ΗΝΥ“Μ÷÷–¬–ΆΒΡPAsΖ÷Ή”(C16H31OVEVE), ”…”Ύ ηΥ°–‘Α±ΜυΥα≤–ΜυVal”κ«ΉΥ°–‘Α±ΜυΥα≤–ΜυGluΫΜΧφ≈≈Ν–, C16H31OVEVEΦΪ“Ή–Έ≥… ηΥ°–‘Val≤–ΜυΚΆ«ΉΥ°–‘Glu≤–ΜυΖ÷±π≈≈Ν–‘Ύ’έΒΰΟφΝΫ≤ύΠ¬-’έΒΰΒΡΕΰΦΕΫαΙΙ÷–. ‘ΎΥ°»ή“Κ÷–, ”…Val ≤–Μυ–Έ≥…ΒΡ ηΥ°ΟφΈΣΝΥ±ήΟβ”κΥ°Ϋ”¥ΞΕχœύΜΞΨέΦ·, ΙC16H31OVEVEΖ÷Ή”–Έ≥…ΝΥΕάΧΊΒΡΕΰΨέΧεΫαΙΙ, ¥”ΕχΉιΉΑ–Έ≥…ΩμΕ»¥ο150 nmΒΡΨό–ΆΡ…ΟΉ¥χΉ¥ΫαΙΙ(Giant nanobelts). Β±‘ω¥σ»ή“ΚpH÷Β ±, ΒγάκΒΡE≤–ΜυΦδΒΡΒγΚ…≈≈≥βΉς”Ο ΙΉιΉΑΧε≤ζ…ζΙφ‘ρΒΡΖλœΕ, ¥”Εχ–Έ≥…¥χ”–ΙΒ≤έΒΡΡ…ΟΉ¥χ(Grooved nanobelt). Ψό–ΆΡ…ΟΉ¥χΉ¥ΫαΙΙΒΡ–Έ≥…Ά§ ±“άάΒ”ΎΕύκΡΖ÷Ή”»ή“ΚΒΡ≈®Ε», ‘ΎΒΆ≈®Ε»(0.01%, ÷ ΝΩΖ÷ ΐ)œ¬ Ήœ»ΉιΉΑ–Έ≥…œΝ’≠ΓΔ ≈Λ«ζΒΡΡ…ΟΉ¥χ(Narrower and twisted nanoribbons); ΥφΉ≈»ή“Κ≈®Ε»ΒΡ…ΐΗΏ(0.05%), ΕύΧθ≈Λ«ζΒΡΡ…ΟΉ¥χΨέΦ·–Έ≥…Ηϋ¥σΒΡ¥χΉ¥ΫαΙΙ, ≤ΔΡήΙέ≤βΒΫΕΰ’ΏΙ≤¥φΒΡάύΥΤ…®÷ψΉ¥ΒΡ÷–ΦδΧ§ΫαΙΙ(Broom morphology); Β±»ή“Κ≈®Ε»¥οΒΫ0.1% ±, Ψό–ΆΡ…ΟΉ¥χΉ¥ΫαΙΙ–Έ≥…. ΕχΫΪC16H31OVEVEΒΡΖ÷Ή”ΫαΙΙΗϋΗΡΈΣC16H31OVVEE ±, ΈόΖ®–Έ≥…Val≤–Μυ ηΥ°ΟφœύΜΞΉς”ΟΒΡΕΰΨέΫαΙΙ, Ρ…ΟΉ¥χΉ¥ΫαΙΙΥφ÷°œϊ ß, ”÷÷Ί–¬ΉιΉΑ–Έ≥…Ρ…ΟΉœΥΈ§ΫαΙΙ. RosiΒ»[35]ΫΪ“ΜΕΈΨΏ”–ΗΏΕ»ΫπΚΆ“χ±μΟφ«ΉΚΆ–‘ΒΡΕύκΡΙΠΡήΝ¥ΕΈAYSSGAPPMPPE”κC12÷±Ϋ”ΦϋΝΣ‘Ύ“ΜΤπ–Έ≥…C12-PEPAUΝΫ«ΉκΡ, ‘Ύ”–HAuCl4¥φ‘ΎΒΡτ«““ΜυΏΏύΚ““ΝρΜ«Υα(HEPES)ΜΚ≥ε“Κ÷–, ΡήΙΜΉιΉΑΩσΜ·–Έ≥…ΗΏΕ»Ιφ’ϊΒΡΉσ ÷ΥΪ¬ί–ΐΕύκΡ-ΫπΡ…ΟΉΩ≈ΝΘ‘”Μ·ΫαΙΙ(ΆΦ4).

WangΒ»[36]ΫΪC12ΆιΜυΝ¥”κΠ¬-ΒμΖέ―υœΥΈ§κΡΒΡ“ΜΕΈΙΠΡή–ρΝ–EVHHQKL[AΠ¬(11~17)]œύΝ§÷Τ±Η≥ωC12-AΠ¬(11~17)ΝΫ«ΉκΡ. ‘ΎΒΆpH÷Β»ή“Κ÷–, C12-AΠ¬(11~17)ΡήΙΜΉιΉΑ≥…≥ΛΕ»≤ΜΒ»ΓΔ ÷±ΨΕ‘ΦΈΣ5 nmΒΡΑτΉ¥œΥΈ§; ΥφΉ≈pH÷ΒΒΡ…ΐΗΏ, ’β–©ΕΧΑτΉ¥œΥΈ§”÷ΡήΫχ“Μ≤ΫœύΜΞ≈≈Ν––Έ≥…≥ΛΒΡœΥΈ§ΫαΙΙ. Εχ»ή“ΚpH=10 ±, ΉιΉΑΧε‘ρΉΣ±δΈΣ≈Λ«ζΒΡ¥χΉ¥ΫαΙΙ, ΆΤ≤β≤ΜΆ§pH÷Β»ή“Κ÷–Ζ÷Ή”ΦδΉς”ΟΝΠΒΡ±δΜ· «ΒΦ÷¬ΉιΉΑΧε–ΈΟ≤ΖΔ…ζΗΡ±δΒΡ÷ς“Σ‘≠“ρ. CoreΒ»[37]œΒΆ≥ΒΊ―–ΨΩΝΥΆιΜυΝ¥ΒΡ≥ΛΕ»ΓΔ ΐΡΩ“‘ΦΑΈ¬Ε»Ε‘Κ§”–ΫΚ‘≠ΒΑΑΉΤ§ΕΈκΡGVKGDKGNP GWPGAP(IVH1)ΒΡPAsΖ÷Ή”ΉιΉΑ––ΈΣΒΡ”Αœλ. ΖΔœ÷Κ§”–ΒΞΧθΆιΜυΝ¥Μρ2ΧθC12~14ΆιΜυΝ¥ΒΡPAsΖ÷Ή”ΡήΙΜΉιΉΑ–Έ≥…«ρ–ΈΫΚ χ; ΕχΚ§”–2ΧθC16~18ΆιΜυΝ¥ΒΡPAsΖ÷Ή”‘ρΡήΉιΉΑ–Έ≥…≈ΧΉ¥ΫΚ χ, ≤Δ«“’β–©≈ΧΉ¥ΫΚ χ‘Ύ “Έ¬œ¬ΡήΫχ“Μ≤ΫœύΜΞΕ―Μΐ–Έ≥…Ηϋ¥σΒΡΑτΉ¥ΫαΙΙ. ΒΪΆ®Ιΐ…ΐΈ¬-ά以¥ΠάμΚσ, ΑτΉ¥ΫαΙΙ‘ρ»Ϊ≤ΩΉΣ±δΈΣ«ρ–ΈΫΚ χ. ¥ΥΆβ, ΫΪΆιΜυΝ¥”κΡ≥–©ΙΠΡήκΡ÷±Ϋ”œύΝ§, άϊ”ΟΤδΕάΧΊΒΡΉιΉΑ–‘ΡήΡήΙΜ”––ßΒΊΧαΗΏ‘≠ΙΠΡήκΡΒΡΉς”Ο–ßΙϊ. »γΫΪΨΏ”–…±ΨζΉς”ΟΒΡΙΠΡήκΡKLFKRHLKWKII(SC4)”κC12ΜρC18œύΝ§÷Τ±ΗΒΡ–¬–ΆΩΙΨζPAsΖ÷Ή”, Ε‘ΗοάΦ œ―τ–‘ΨζΒΡ…±Οπ–ßΙϊ±»‘≠SC4κΡΧαΗΏΝΥ30±Ε, …θ÷ΝΕ‘¥ΪΆ≥ΩΙΨζ“©ΈοΈό–ßΒΡΫπΜΤ…ΪΤœΧ―«ρΨζ“≤”–ΚήΚΟΒΡ…±ΟπΉς”Ο[38].
≥ΐΝΥ…œ ωΜ·―ßΦϋΝΣΆβ, Ά®ΙΐΨ≤ΒγΉς”ΟΦΑ÷ςΩΆΧεœύΜΞΉς”ΟΒ»Ζ«Ι≤ΦέΦϋΉς”ΟΝΠ“≤Ρή÷Τ±ΗΗ¥Κœ–ΆPAsΖ÷Ή”ΉιΉΑΡΘΩι. KrosΒ»[39]άϊ”ΟΜΖΚΐΨΪ”κΫπΗ’ΆιΒΡ÷ςΩΆΧεœύΜΞΉς”Ο, ΫΪΚ§”–ΫπΗ’ΆιΕΥΜυΒΡΕύκΡΖ÷Ή”–ό Έ‘ΎΜΖΚΐΨΪΡ“≈ί±μΟφ. Β±»ή“ΚpH=5.0 ±, Ρ“≈ί±μΟφΒΡΕύκΡΝ¥Ω…“‘Ά®Ιΐ«βΦϋΉς”Ο–Έ≥…Π¬-’έΒΰΒΡΕΰΦΕΫαΙΙ; ΕχΒ±»ή“ΚpH=7.4 ±, ΉιΉΑΧεΫαΙΙΩ…“‘¥”«ρ–ΈΡ“≈ίΫαΙΙΉΣ±δΈΣœΏ–‘œΥΈ§, ≤Δ«“Ρ“≈ί÷–ΑϋΙϋΒΡ“©ΈοΖ÷Ή”“≤Υφ÷° ΆΖ≈. SchermanΒ»[40]άϊ”Ο÷ςΩΆΧεœύΜΞΉς”ΟΫΪή≈κΡΓΔ Ήœ÷§“‘ΦΑΚυ¬ΪκεœύΝ§Ϋ”, ÷Τ±ΗΝΥ–¬–Ά»ΐ‘ΣΗ¥ΚœΖ÷Ή”(ή≈κΡ/Κυ¬Ϊκε/Ήœ÷§), ¥Υ»ΐ‘ΣΗ¥ΚœΖ÷Ή”ΡήΙΜΉ‘ΉιΉΑ–Έ≥…Ρ“≈ίΫαΙΙ. Β±ΧεœΒ÷–¥φ‘Ύ–¬ΒΡΩΆΧεΖ÷Ή”2,6-Εΰτ«ΜυίΝ ±, Υϋ”κΉœ÷§ΦδΗϋ«ΩΒΡΉς”ΟΜα Ιή≈κΡ”Έάκ≥ωά¥, ≤Δ–Έ≥…–¬ΒΡ»ΐ‘ΣΗ¥ΚœΖ÷Ή”(2,6-Εΰτ«ΜυίΝ/Κυ¬Ϊκε/Ήœ÷§), ΉιΉΑΧεΫαΙΙ“≤Υφ÷°”…Ρ“≈ίΉΣ±δΈΣΡ…ΟΉΝΘΉ”. Β±ΩΆΧεΖ÷Ή”±δΈΣΫπΗ’Άι ±, ‘ρΜα”κΚυ¬ΪκεΖ÷Ή”œύΜΞΉς”Ο, »ΐ‘ΣΗ¥ΚœΖ÷Ή”ΫβΧε, ΉιΉΑΧε“≤ΥφΉ≈œϊ ß. ΗϋΈΣΧΊ βΒΡ «, ”…”Ύή≈κΡΖ÷Ή”Υυ¥ΠΈΔΙέΜΖΨ≥ΒΡ≤ΜΕœ±δΜ·, ¥ΥΙΐ≥ΧΆ§ ±Αι”–”ΪΙβΒΡβßΟπΚΆΜ÷Η¥. ’β÷÷ΨΏ”–”ΪΙβΩΣΙΊΙΠΡήΒΡ–¬–ΆΡ…ΟΉ≤ΡΝœ‘Ύ…ζΈο≥…œώΚΆ“©Έο¥ΪΒίΒ»Νλ”ρœ‘ Ψ≥ωΝΦΚΟΒΡ”Π”Ο«±ΝΠ.
ΫϋΡξά¥, “Μ–©–¬ΒΡΙΊ”ΎPAsΉ‘ΉιΉΑ―–ΨΩ±ΜœύΦΧ±®Βά, DasΒ»[41]…ηΦΤΚœ≥…ΝΥ“Μ÷÷Κ§”–άΒΑ±Υα≤–ΜυΒΡ–¬–ΆPAsΖ÷Ή”, Τδ‘ΎΥ°»ή“Κ“‘ΦΑΥ°ΚΆ”–Μζ»ήΦΝΒΡΜλΚœ»ή“Κ÷–ΡήΙΜΉιΉΑ–Έ≥…ΨΏ”–¬ί–ΐΡ…ΟΉœΥΈ§ΫαΙΙΒΡΡΐΫΚΧεœΒ; Υυ–Έ≥…ΒΡ¬ί–ΐΡ…ΟΉœΥΈ§ΡήΙΜΉςΈΣΡΘΑε‘≠ΈΜΚœ≥…ΫαΙΙΨυ“ΜΒΡΫιΩΉΙηΒΞ±ΎΧΦΡ…ΟΉΙή. ΕχΫΪΨΏ”–‘≠Ώ≤Ώχ[(PPIX)Zn]¬γΚœΡήΝΠΒΡΉιΑ±Υα≤–Μυ“ΐ»κΒΫC16-AHL3K3ΕύκΡΖ÷Ή”÷–, Ά®ΙΐC16-AHL3K3ΉιΉΑ–Έ≥…ΒΡœΥΈ§’σΝ–ΫαΙΙΩ…“‘ΒςΩΊΫπ τΏ≤ΏχΒΡ”––ρ≈≈Ν–, ¥”Εχ÷Τ±Η≥ωΨΏ”–ΧΊ βΙβΈοάμ–‘ΡήΒΡ–¬–ΆΡ…ΟΉ≤ΡΝœ[42]. DaninoΒ»[43]…ηΦΤΚœ≥…ΝΥΝμ“Μ÷÷ΫαΙΙάύΥΤ”Ύ2ΗωPAsΖ÷Ή” ΉΈ≤œύΝ§Εχ≥…C12-Π¬12ΕύκΡΖ÷Ή”, ¥ΥάύΖ÷Ή”‘ΎΉιΉΑ ±ΡήΙΜΖΔ…ζΝ¥ΕΈΒΡΖ÷Ή”ΡΎ’έΒΰ¥”Εχ–Έ≥…≤ΜΆ§ΒΡΉιΉΑΧεΫαΙΙ.
―–ΨΩΖΔœ÷, ‘ΎΒΆΈ¬ΧθΦΰœ¬, C12-Π¬12 Ήœ»ΉιΉΑ–Έ≥…œΥœΗΒΡœΥΈ§, ΥφΉ≈≈ύ”ΐ ±ΦδΒΡ―”≥Λ, œΥΈ§ΫχΕχΉΣ±δΈΣ≈Λ«ζΒΡ¥χΉ¥ΫαΙΙ, ≤ΔΫχ“Μ≤ΫΨμ«ζ≥…Ρ…ΟΉΙήΉ¥ΫαΙΙ. …ΐΈ¬÷Ν45 ΓφΚσ, C12-Π¬12Ζ÷Ή”ΡΎ’έΒΰΖΫ ΫΖΔ…ζ±δΜ·, ΉιΉΑΧεΥφ÷°ΉΣ±δΈΣ«ρΉ¥ΫΚ χ.
StuppΒ»[44]‘ρΖΔ’ΙΝΥ“Μ÷÷≤ΌΩΊPAsΕύκΡΉιΉΑΒΡΖΫΖ®, ΥϊΟ«ΫΪΕύκΡ»ή“Κ”κ¥χ”–œύΖ¥ΒγΚ…ΒΡΨέΚœΈο»ή“ΚœύΜλΚœ, άϊ”ΟΒγΚ…ΒΡœύΜΞΉς”Ο ΙΕύκΡ‘Ύ2÷÷“ΚΧεΫγΟφΫχ––ΉιΉΑ, Ά®ΙΐΒςΫΎΆβΦ”Βγ≥ΓΒΡ«ΩΕ»ΚΆΖΫœρά¥ΗΡ±δHAΖ÷Ή”‘ΎΕύκΡ»ή“Κ÷–ΒΡ…χΆΗΥΌΕ»ΚΆΖΫœρ, ¥”ΕχΗΡ±δΡΛΫαΙΙ÷–ΒΡœΥΈ§ χ≈≈Ν–ΖΫœρ, ≤Δ ΙΡΛΚώΕ»ΦΑ”≤Ε»Β»ΈοάμΧΊ–‘ΖΔ…ζΗΡ±δ. SpoerkeΒ»[45]œΒΆ≥ΒΊ―–ΨΩΝΥΒγΚ…Ής”ΟΕ‘PAsΖ÷Ή”Ή‘ΉιΉΑΒΡ”Αœλ, ΖΔœ÷Ζ÷÷ΠΕΥΜυ¥χ”–ΒΡ≤ΜΆ§ΒγΚ…Ω…“‘ ΙPAs≥ …λ’ΙΜρΧ°œίΒΡΖ÷Ή”ΙΙ–Ά, ¥”Εχ–Έ≥…ΫΚ χΓΔ œΥΈ§ΚΆΥΪ≤ψΡΛΒ»≤ΜΆ§ΒΡΉιΉΑΫαΙΙ.
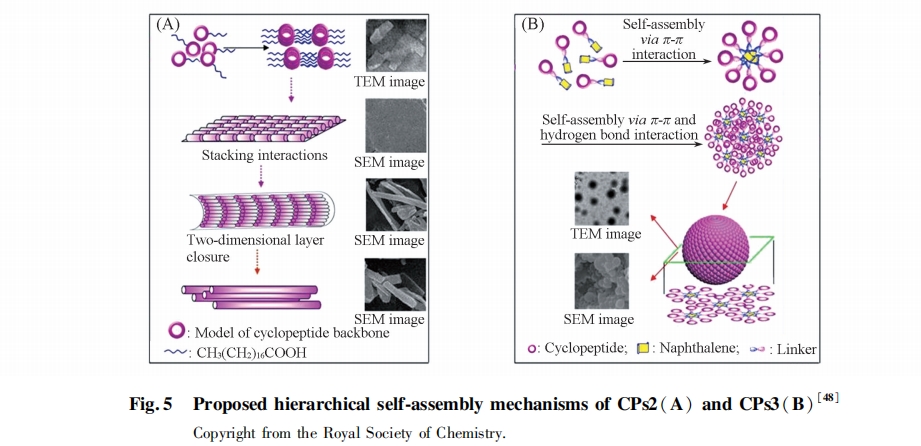
ΫϋΡξά¥Έ“Ο«‘ΎPAsΕύκΡΉ‘ΉιΉΑ―–ΨΩΖΫΟφ“≤Ϋχ––ΝΥ“ΜœΒΝ–œύΙΊΒΡΙΛΉς. ―–ΨΩΝΥ ηΥ°Ν¥ΫαΙΙΒΡ±δΜ·Ε‘PAsΕύκΡΉιΉΑΒΡ”Αœλ, ΖΔœ÷Έό¬έ «ΆιΜυΝ¥ΜΙ «ΖΦœψΉεΜυΆ≈”κRGD«ΉΥ°ΕΧκΡΙΙ≥…ΒΡPAsΖ÷Ή”ΕΦΡήΙΜΉιΉΑ–Έ≥…Ρ…ΟΉœΥΈ§ΚΆΫΚ χ, «“”…”Ύ ηΥ°Ν¥ΕΈΫαΙΙΒΡ≤ΜΆ§ΒΦ÷¬ΤδΉιΉΑΧε–ΈΟ≤‘Ύ≤ΜΆ§pH÷Β»ή“Κ÷–ΒΡ±δΜ·“≤≤ΜΆ§[46]. ΕχΆιΜυΝ¥ΒΡ≥ΛΕ»±δΜ·Ά§―υ“≤Ω…“‘œ‘÷χΒΊ”ΑœλΒΫPAsΖ÷Ή”ΒΡΉιΉΑΧΊ–‘ΚΆΉιΉΑΧε–ΈΟ≤, ΥφΉ≈»ή“ΚpH÷Β±δΜ·, Ϋœ≥ΛΒΡΆιΜυΝ¥(C16~C18)œύΜΞΉς”Ο–Έ≥…Ηϋ«ΩΒΡ ηΥ°Ής”ΟΩ…“‘±ΘΜΛΉιΉΑΧεΫαΙΙΟβ ήΨ≤Βγ≈≈≥βΉς”ΟΒΡ”Αœλ, ¥”Εχ ΙΒΟΥυ–Έ≥…ΒΡΡ…ΟΉœΥΈ§Έό¬έ‘ΎΥα–‘ΓΔ ÷––‘ΜΙ «Φν–‘ΜΖΨ≥÷–ΕΦΡήΈ»Ε®¥φ‘Ύ; ΕχΫœΕΧΒΡΆιΜυΝ¥(C12~C14)”…”ΎΤδ ηΥ°Ής”ΟΫœ»θ, œύ”ΠΒΡPAs‘ΎΥα–‘»ή“Κ÷–ΉιΉΑ–Έ≥…ΒΡΡ…ΟΉœΥΈ§ΥφΉ≈ΜΖΨ≥pH÷ΒΒΡ…ΐΗΏ‘ρ»Ϊ≤ΩΉΣ±δΈΣΡ…ΟΉΫΚ χΫαΙΙ[47]. Ϋχ“Μ≤ΫΫΪ«ΉΥ°–‘ΒΡΜΖκΡ”κ¥ΉΥαΓΔ ”≤÷§ΥαΚΆίΝΜΖœύΝ§Ϋ”ΜώΒΟΝΫ«Ή–‘ΕύκΡCPs1~3, ―–ΨΩΖΔœ÷CP1Ά®ΙΐΖ÷Ή”Φδ―Έ«≈ΚΆ«βΦϋΉς”ΟΡήΙΜΉιΉΑ–Έ≥…ΖΡ¥ΗΉ¥Ρ…ΟΉΫαΙΙ; CP2Ω…“‘Ά®ΙΐΆιΜυΝ¥ΦδΒΡ ηΥ°Ής”ΟΚΆκΡΝ¥ΦδΒΡ«βΦϋΡήΙΜΉιΉΑ–Έ≥…Ρ…ΟΉΙή; ΕχCP3‘ρΆ®ΙΐΖΦœψΜΖΦδΒΡΠ–-Π–œύΜΞΉς”ΟΚΆκΡΝ¥ΦδΒΡ«βΦϋΡήΙΜΉιΉΑ–Έ≥…Ρ…ΟΉ«ρΫαΙΙ(ΆΦ5)[48].
ΝμΆβ, Κ§”–2Χθ ηΥ°Ν¥ΒΡU–ΆPAsΡήΙΜ‘Ύ≤ΜΆ§pH÷Β»ή“Κ÷–ΖΔ…ζZ–ΆΚΆœΏ–ΆΙΙœσΒΡΉΣ±δ, ¥”Εχ“ΐΤπΉιΉΑΧεΫαΙΙΒΡ±δΜ·[49]. Έ“Ο«[50~52]ΫΪΩγΡΛκΡR8ΚΆΚΥΕ®ΈΜ–≈Κ≈κΡPKKKRKV”κC18ΆιΜυΝ¥œύΝ§÷Τ±ΗΒΡPAsΖ÷Ή”, ΤδΩ…“‘”κ“©ΈοΖ÷Ή”Μρ÷ΈΝΤΜυ“ρΙ≤ΉιΉΑ–Έ≥…Έ»Ε®ΒΡΡ…ΟΉ«ρΉ¥ΫαΙΙ, ΡήΙΜ”––ßΧαΗΏ“©Έο¥ΪΒίΚΆΜυ“ρΉΣ»Ψ–߬ [ΆΦ6(A)]. ΕχΫΪ ηΥ°–‘“©ΈοΖ÷Ή”ΑΔΟΙΥΊ(DOX)Ι≤ΦέΝ§Ϋ”ΒΫAc-GRGDSR8ΕύκΡΝ¥ΕΈ…œ, –Έ≥…«Α“©άύPAsΖ÷Ή”. ¥ΥάύPAsΖ÷Ή”ΡήΙΜΉιΉΑ–Έ≥…ΫΪDOXΑϋΙϋΤδ÷–ΒΡΡ…ΟΉΩ≈ΝΘ, “άάΒΙΠΡήκΡR8ΒΡ¥©ΡΛΉς”ΟΚΆRGDΒΡΑ–œρΉς”Ο, Βœ÷ΝΥΩΙΑ©“©ΈοΒΡΗΏ–ßΚΆ―Γ‘ώ–‘¥ΪΒί[ΆΦ6(B)][53].
3 ΨΏ”–ΧΊ βΖ÷Ή”ΫαΙΙΒΡΝΫ«Ή–‘ΕύκΡΉ‘ΉιΉΑ
ΫϋΡξά¥, “Μ–©ΨΏ”–ΧΊ βΫαΙΙΒΡΝΫ«Ή–‘ΕύκΡΖ÷Ή””…”ΎΤδΕάΧΊΒΡΉιΉΑ–‘Ρή»’“φ ήΒΫ―–ΨΩ’ΏΒΡΙΊΉΔ, Τδ÷–”–¥ζ±μ–‘ΒΡ «Bola–ΆΝΫ«Ή–‘ΕύκΡΚΆGemini–ΆΝΫ«Ή–‘ΕύκΡ.
3.1 Bola–ΆΝΫ«Ή–‘ΕύκΡΉ‘ΉιΉΑ
Bola–ΆΝΫ«Ή–‘ΕύκΡΖ÷Ή”ΫαΙΙ≥ ―ΤΝε–Έ, «ΫΪ2Ηω«ΉΥ°–‘ΕύκΡΝ¥ΕΈ”Ο“ΜΧθΜρΕύΧθ ηΥ°Ν¥ΕΈΝ§Ϋ”Τπά¥ΒΡΝΫΕΥ«ΉΥ°ΓΔ ÷–Φδ ηΥ°ΒΡΧΊ βΫαΙΙ, ”…”ΎΩαΥΤΡœΟάΆΝ÷χ»ΥΒΡ“Μ÷÷ΈδΤςBola(2–Γ«ρΖ÷±πœΒ”Ύ“ΜΗυœΗ…ΰΝΫΕΥ)ΕχΒΟΟϊ. Bola–ΆΝΫ«ΉκΡ”…”ΎΤδ±Ψ…μΫαΙΙΒΡΧΊΒψ”–Ή≈ΫœΈΣΕάΧΊΒΡΉιΉΑ––ΈΣ, Τδ÷–ΙήΉ¥ΫαΙΙ «Bola–ΆΝΫ«ΉκΡ÷–ΒΡ≥ΘΦϊΉιΉΑΫαΙΙ. MatsuiΒ»[54]…ηΦΤΚœ≥…ΝΥ“Μ÷÷Bola–ΆΝΫ«ΉκΡGG-C7-GG, Τδ‘ΎΦν–‘»ή“Κ(pH=8)÷–ΡήΙΜΉιΉΑ–Έ≥…Ωμ‘Φ2 ΠΧmΓΔ ≥Λ‘Φ10~40 ΠΧmΒΡ¬ί–ΐ¥χΉ¥ΫαΙΙ; ΕχΫΒΒΆ»ή“ΚpH÷ΒΒΫ5 ±, ‘ρΡήΙΜΉιΉΑ–Έ≥…ΤΫΨυ÷±ΨΕΈΣ500 nmΒΡ÷–Ω’Ρ…ΟΉΙή[ΆΦ7(A)]. ΉιΉΑΜζάμ―–ΨΩ±μΟς, ‘Ύ≤ΜΆ§pH÷Βœ¬, GG-C7-GGΖ÷Ή”Φδ«βΦϋ«ΩΕ»ΒΡ±δΜ· «ΨωΕ®ΉιΉΑΧε–ΈΟ≤ΒΡ÷ς“Σ“ρΥΊ[ΆΦ7(B)].
Β±œρΉιΉΑΧεœΒΦ”»κ“ΜΕ®≈®Ε»ΒΡNi2+άκΉ”Κσ, ΡήΙΜ‘ΎBolaκΡΡ…ΟΉΙήΦδ–Έ≥…Ϋπ τάκΉ”«≈(Ni ion bridge)ΚΆΙήΦδ«βΦϋΉς”Ο, ¥”Εχ ΙΖ÷…ΔΈό–ρΒΡBolaκΡΡ…ΟΉΙή÷Ί–¬Ε®œρ≈≈Ν––Έ≥…Ιφ’ϊΒΡΡ…ΟΉΙή χ. Τδ÷–Ρ…ΟΉΙή χΒΡ–Έ≥… ήΫπ τάκΉ”÷÷άύΓΔ ≈®Ε»ΓΔ Φ”»κ ±Φδ“‘ΦΑ «Ζώ¥φ‘Ύ““ΕΰΑΖΥΡ““Υα(EDTA)¬γΚœΦΝΒΡ”Αœλ[55]. ΥφΚσΥϊΟ«”÷Ε‘BolaκΡΡ…ΟΉΙήΒΡΙήΨΕΫχ––ΝΥΒςΩΊ, άϊ”ΟΕύΩΉΧΦΥαθΞΈΣΡΘΑε÷Τ±ΗΙήΨΕΨυ“ΜΒΡΕύκΡΡ…ΟΉΙή[56]. ¥ΥΆβ, Έ“Ο«[57]“≤Κœ≥…ΝΥ“Μ÷÷Bola–ΆΝΫ«Ή–‘ΕύκΡΖ÷Ή”, ΆιΜυΝ¥ΝΫΕΥΖ÷±πΝ§Ϋ”ΝΥD/LΙΙ–ΆœύΦδΒΡΜΖκΡ, ΗΟκΡΡήΙΜ‘ΎΥ°»ή“Κ÷–Ή‘ΉιΉΑ–Έ≥…ΒΞ±ΎΒΡΡ…ΟΉΙή. BolaκΡΡ…ΟΉΙήΨΏ”–“Ή–ό ΈΦΑΫαΙΙΈ»Ε®Β»”≈Βψ, ΨΏ”–ΙψΖΚΒΡ”Π”Ο«ΑΨΑ. Ά®ΙΐœρBolaκΡΡ…ΟΉΙή…œ“ΐ»κΗΜΚ§His≤–ΜυΒΡΙΠΡή–‘ΕΧκΡΖ÷Ή”, άϊ”ΟHis≤–ΜυΕ‘Ϋπ τάκΉ”ΒΡ¬γΚœΉς”Ο”’ΒΦΫπ τάκΉ”(Au, Cu, Ag, NiΒ»)‘ΎΡ…ΟΉΙή±Ύ…œΫαΨß…ζ≥Λ[58~60]. ΫΪΙβΟτ–‘ΒΡ≈ΦΒΣ±ΫΜυΆ≈“ΐ»κΒΫBolaκΡΡ…ΟΉΙή…œ, άϊ”Ο≈ΦΒΣ±Ϋ”κΠΝ-ΜΖΚΐΨΪΒΡ÷ςΩΆΧεœύΜΞΉς”Ο, Βœ÷ΝΥκΡΡ…ΟΉΙή‘Ύ–ό Έ”–ΠΝ-ΜΖΚΐΨΪΒΡΙη±μΟφΒΡΙβΩΊΈϋΗΫΚΆΫβάκ[61]. άϊ”Ο…ζΈοΩΙ‘≠-ΩΙΧεœύΜΞ Ε±πΚΆΉς”Ο, –ό Έ”–ΩΙΧεΒΡBolaκΡΡ…ΟΉΙήΡήΙΜ‘ΎΨΏ”–ΩΙ‘≠’σΝ–ΒΡΫπ±μΟφΫχ––”––ρΒΡΈϋΗΫΚΆ≈≈Ν–[62]. άϊ”Ο‘≠Ώ≤ΏχΔυ Zn(Δρ)”κBolaκΡΡ…ΟΉΙήΦδΒΡ«βΦϋΉς”Ο, Ω…ΫΪ‘≠Ώ≤ΏχΔυ Zn(Δρ)ΑϋΙϋ”ΎκΡΡ…ΟΉΙή…œ–Έ≥…Ϋπ τΏ≤ΏχΡ…ΟΉΙή, ”–Άϊ”Ο”ΎΡ…ΟΉΜ·―ߥΪΗ–ΤςΚΆΙβΒγ≤ΡΝœΒΡ÷Τ±Η[63]. ¥ΥΆβ, BolaκΡΡ…ΟΉΙήΜΙΩ…”Π”Ο”ΎΟβ“Ώ¥ΪΗ–Τς÷–, ”Ο”ΎΦλ≤βΒΞ¥Ωπε’ν≤ΓΕΨΒ»≤Γ‘≠Χε[64].
≥ΐΙήΉ¥ΫαΙΙΆβ, StuppΒ»[65]±®ΒάΝΥΝμ“Μ÷÷≤ΜΕ‘≥ΤΒΡBolaΝΫ«ΉκΡΖ÷Ή”, Τδ ηΥ°Ν¥ΝΫΕΥΖ÷±πΝ§Ϋ”ΝΥΕύκΡΝ¥ΕΈΚΆΒΆΨέ““Εΰ¥Φ. ‘ΎΥ°»ή“Κ÷–ΉιΉΑ ±, ηΥ°Ν¥ΕΈ…ν¬ώ”ΎΒΞΖ÷Ή”≤ψΡΎ≤Ω, –Έ≥…Ά§ ±ΨΏ”–«ΉΥ°–‘ΡΎΚΥΚΆ«ΉΥ°–‘±μΟφΒΡΥΪ«Ή–‘ΒΡΧΊ βœΥΈ§ΫαΙΙ. KogisoΒ»[66]…ηΦΤΚœ≥…ΝΥ“ΜœΒΝ–BolaΝΫ«ΉκΡVV-(CH2)n-VV, ≤ΔœΒΆ≥―–ΨΩΝΥ÷–Φδ ηΥ°Ν¥≥ΛΕ»Ε‘ΤδΉιΉΑ––ΈΣΒΡ”Αœλ. ΖΔœ÷Β±n=4Μρ6 ±, Ω…“‘‘ΎΥ°»ή“Κ÷–ΉιΉΑ–Έ≥…άύΥΤΫαΨßΙΧΧεΈοΒΡ≥ΝΒμ; Εχn=7~12 ±, Ω…“‘‘ΎΥ°»ή“Κ÷–“‘Π¬-’έΒΰ–Έ ΫΉιΉΑ–Έ≥…Ρ…ΟΉœΥΈ§ΫαΙΙ. ¥ΥάύBolaκΡœΥΈ§ΨΏ”–”κNi2+, Zn2+, Cu2+, Co2+Β»ΕΰΦέΫπ τάκΉ”¬γΚœΒΡΡήΝΠ, ΡήΙΜΫχ“Μ≤ΫΉιΉΑ–Έ≥…ΫαΨßΙΧΧε≥ΝΒμΈο, ΨΏ”–Ρ…ΟΉœΥΈ§ΫαΙΙΫΚΧεΖ÷…ΔΈο“‘ΦΑΥ°ΡΐΫΚΒ»[67].
3.2 Gemini–ΆΝΫ«Ή–‘ΕύκΡΉ‘ΉιΉΑ
Gemini–ΆΝΫ«Ή–‘ΕύκΡΖ÷Ή”ΒΡ…ηΦΤΝιΗ–‘¥”ΎGemini–Ά±μΟφΜν–‘ΦΝ. Gemini–Ά±μΟφΜν–‘ΦΝ «“Μ÷÷–¬–ΆΒΡ±μΟφΜν–‘ΦΝ, «”…2ΗωΒΞΝ¥±μΟφΜν–‘ΦΝΖ÷Ή”Ά®ΙΐΩΩΫϋΆΖ≤ΩΒΡΝΣΫαΜυΆ≈Μ·―ßΦϋΚœΕχ≥…ΒΡ. ¥”ΫαΙΙ…œΩ¥, άύΥΤ”Ύ2ΗωΒΞΝ¥Ζ÷Ή”ΒΡΕΰΨέΧε, Υυ“‘≥ΤΉςΥΪΉ”(Gemini)±μΟφΜν–‘ΦΝ. “ΜΑψ±μΟφΜν–‘ΦΝ‘ΎΥ°÷–ΉιΉΑ–Έ≥…ΨέΦ·Χε ±, ή2÷÷œύΖ¥Ής”ΟΝΠΒΡ”Αœλ: “Μ÷÷ « ηΥ°Ν¥ΕΈ÷°ΦδΒΡ ηΥ°œύΜΞΉς”Ο, «ΐΕ·±μΟφΜν–‘ΦΝΖ÷Ή”ΫτΟή≈≈Ν–; Νμ“Μ÷÷ ««ΉΥ°ΆΖ≤ΩΆ§÷÷ΒγΚ…ΒΡΨ≤Βγ≥βΝΠ, ΙΒΟ±μΟφΜν–‘ΦΝΖ÷Ή”ΦδΨύ‘ω¥σ, ΤΤΜΒΉιΉΑ. ΕχGemini–Ά±μΟφΜν–‘ΦΝΒΡΜ·―ßΦϋΝΣΫαΙΙ”––ßΒΊœϊ≥ΐΝΥ«ΉΥ°ΆΖΜυΦδΨ≤Βγ≈≈≥βΉς”Ο, ΙΒΟ ηΥ°Ν¥ΕΈΗϋΦ”ΫτΟήΒΊ≈≈Ν–, ‘ω«ΩΝΥΖ÷Ή”ΦδΒΡ ηΥ°œύΜΞΉς”Ο, ’β «Gemini–Ά±μΟφΜν–‘ΦΝ±»¥ΪΆ≥ΒΞΝ¥±μΟφΜν–‘ΦΝΨΏ”–ΗϋΗΏ±μΟφΜν–‘ΒΡΗυ±Ψ‘≠“ρ. ΗϋΈΣ÷Ί“ΣΒΡ «, GeminiΒΡΕάΧΊΫαΙΙΈΣΉ‘ΉιΉΑΙΐ≥ΧΧαΙ©ΝΥΗϋΦ”ΖαΗΜΒΡΫαΙΙΒςΩΊ“ρΥΊ, ¥”Εχ≤ζ…ζΝΥΗ¥‘”Εχ–¬”±ΒΡΉιΉΑΧΊ–‘[68~71]. LuisΒ»[72,73]±®ΒάΝΥ“ΜœΒΝ–Gemini–ΆΝΫ«ΉκΡ, ΖΔœ÷‘ΎΖ÷Ή”ΫαΙΙΓΔ »ήΦΝΦΪ–‘“‘ΦΑ»ή“ΚpHΒ»“ρΥΊΒΡΒςΩΊœ¬, ΤδΡήΙΜΉιΉΑ–Έ≥…ΦΪΈΣΖαΗΜΒΡΉιΉΑΧε–ΈΟ≤. WangΒ»[74]ΫΪ2ΧθΆιΜυΝ¥C12”κΠ¬-ΒμΖέ―υΕύκΡ(Amyloid-Π¬ peptide AΠ¬)ΙΠΡήΤ§ΕΈΒΡ≤ΜΆ§≤ΩΈΜœύΝ§÷Τ±ΗΝΥ2÷÷≤ΜΆ§ΒΡGemini–ΆΝΫ«ΉκΡ2C12-Lys-AΠ¬(12~17)ΚΆC12-AΠ¬(11~17)-C12. ―–ΨΩΖΔœ÷2C12-Lys-AΠ¬(12~17)‘Ύ≤ΜΆ§pHΖΕΈßΡΎΕΦΡήΉιΉΑ–Έ≥…Έ»Ε®ΒΡΡ…ΟΉœΥΈ§ΫαΙΙ; ΕχC12-AΠ¬(11~17)-C12‘ΎΫœΒΆpH÷Β»ή“Κ÷–Ω…“‘ΉιΉΑ–Έ≥…ΕΧΒΡ≈Λ«ζ¥χΉ¥ΫαΙΙ, ΥφΉ≈pH÷ΒΒΡ…ΐΗΏ, ΉΣ±δΈΣ±ΓΤ§Ή¥ΫαΙΙ[ΆΦ8(A)], ΈΣΆ®ΙΐΖ÷Ή”ΫαΙΙΒΡΦρΒΞ±δΜ· Βœ÷ΉιΉΑΧε–ΈΟ≤ΒΡΒςΩΊΧαΙ©ΝΥ“Μ÷÷–¬ΒΡΆΨΨΕ. ¥ΥΆβ, ¥χ”–’ΐΒγ–‘Α±ΜυΥα≤–ΜυΒΡ―τάκΉ”Gemini–ΆΝΫ«ΉκΡ”…”ΎΨΏ”–ΝΌΫγΫΚ χ≈®Ε»(cmc)ΒΆΓΔ Ε‘ΒγΫβ÷ ≤ΜΟτΗ–Β»”≈Βψ, ΡήΙΜ”κDNAΗ¥ΚœΉιΉΑ–Έ≥…ΝΘΨΕΗϋ–ΓΓΔ ΫαΙΙΗϋΦ”Έ»Ε®ΒΡΡ…ΟΉ«ρΉ¥ΫαΙΙ, ”–Άϊ≥…ΈΣ“Μ÷÷–¬–ΆΒΡΧεΡΎΉΔ…δΜυ“ρ‘ΊΧε≤ΡΝœ[ΆΦ8(B, C)][75,76].

ΫϋΡξά¥, Έ“Ο«[77]…ηΦΤΚœ≥…ΝΥ“ΜœΒΝ–ΨΏ”–≤ΜΆ§≥ΛΕ»ΆιΜυΝ¥ΒΡGAPsΖ÷Ή”, άϊ”ΟΤδΫχ––Ή‘ΉιΉΑ÷Τ±Η≥ωΖ÷÷ΠΉ¥œΥΈ§ΫαΙΙ. ‘ΎΨΏ”–ΝΦΚΟΤΫΚβΒΡΖ÷Ή”Φδ ηΥ°Ής”ΟΚΆ«βΦϋ–≠Ά§«ΐΕ·œ¬, (C10-C-O3)2ΚΆ(C12-C-O3)2Ζ÷Ή”ΡήΙΜΉιΉΑ–Έ≥…≥ΛΒΡΖ÷÷ΠΉ¥œΥΈ§; Εχ ηΥ°Ής”ΟΚΆ«βΦϋ–≠Ά§Ής”ΟΤΫΚβΒΡΤΤΜΒ, ΒΦ÷¬(C14-C-O3)2ΚΆ(C16-C-O3)2Ζ÷Ή”ΉιΉΑ–Έ≥…≤ΜΨΏ”–»ΈΚΈΖ÷÷ΠΫαΙΙΒΡΕΧœΥΈ§.
Ά®ΙΐΖ¬…ζ…ηΦΤΝΥΝμ“ΜάύΈ≤≤ΩΆιΜυΝ¥Κ§”–≤ΜΆ§ ΐΡΩΥΪΦϋΒΡGAPsΖ÷Ή”, άϊ”Ο’β–©ΦρΒΞΒΡΕύκΡΖ÷Ή”, ÷Τ±Η≥ω–ΈΟ≤ΚΆ≥Ώ¥γ≤ΜΆ§ΒΡΡ“≈ίΫαΙΙ, Τδ÷–Αϋά®”…Ρ…ΟΉΡ“≈ί÷πΦΕ»ΎΚœ–Έ≥…ΒΡΈΔΟΉΦΕœΗΑϊΉ¥ΒΡΨό–ΆΡ“≈ί. Μζάμ―–ΨΩΖΔœ÷, Ρ“≈ί–ΈΟ≤ΚΆ≥Ώ¥γΒΡ±δΜ·Ω…Ρή «”κΥ≥ ΫΥΪΦϋ“ΐΤπΒΡGAPsΖ÷Ή”‘ΎΡ“≈ίΥΪΖ÷Ή”≤ψΡΛ÷–ΒΡ≈≈Ν–ΫτΟή≥ΧΕ»ΒΡ±δΜ·”–ΙΊ[78]. GAPsΖ÷Ή”Ε‘œύ”ΠΒΡΆ§‘¥–‘ΒΞΝ¥PAsΖ÷Ή”ΒΡΉ‘ΉιΉΑΨΏ”–ΚήΚΟΒΡΒςΩΊΉς”Ο, ΡήΙΜ ΙΉιΉΑΧε”–œΥœΗΒΡ≥ΛœΥΈ§ΉΣ±δΈΣΨυ“ΜΒΡΓΔ ΫαΙΙΗϋΦ”Έ»Ε®ΒΡΕΧΑτΉ¥œΥΈ§. ΕΧΑτΉ¥œΥΈ§‘Ύ”…GAPsΧΊ βΥΪΝ¥ΫαΙΙ≤ζ…ζΒΡœΥΈ§Φδ ηΥ°Ής”ΟΒΡ«ΐΕ·œ¬, Ϋχ“Μ≤ΫΉ‘ΖΔΒΊ÷Ί–¬Ε®œρ≈≈Ν–, ≤Δ÷πΫΞ―”≥Λ≥ΛΕ», Ήν÷’”…―”…λΖΫœρ≤ΜΆ§ΒΡΤΫ––≈≈Ν–ΒΡœΥΈ§œύΜΞΫΜ÷·ΫχΕχ–Έ≥…ΝΥΗ¥‘”ΒΡΓΔ ΗΏΕ»”––ρΒΡΓΑ±ύ÷·–ΆΓ±œΥΈ§’σΝ–. Ά§ ±, ¥ΥάύΕύκΡΜΙΨΏ”–≥ω…ΪΒΡΫγΟφΉιΉΑΡήΝΠ, ‘Ύ≤ΜΆ§«Ή ηΥ°–‘ΫγΟφ…œΩ…“‘ΖΫ±ψ―ΗΥΌΒΊ–Έ≥…ΨΏ”–Ω’ΦδΝΔΧεΫαΙΙΒΡΕύκΡΡ…ΟΉ’σΝ–[79].
4 ’Ι Άϊ
Ϋϋ–©ΡξΕύκΡΉ‘ΉιΉΑ―–ΨΩΦΑΤδ‘ΎΡ…ΟΉΩΤΦΦΓΔ …ζΈο“Ϋ”ΟΝλ”ρΒΡ”Π”Ο ήΒΫΝΥΦΪ¥σΙΊΉΔ≤Δ»ΓΒΟΝΥœ‘÷χΒΡΫχ’Ι, Ά®Ιΐ‘Ύ÷νΕύΝλ”ρΒΡΫΜ≤φ…χΆΗ, ‘Ύ–¬≤ΡΝœ÷Τ±ΗΖΫΟφœ‘ Ψ≥ωΨό¥σΒΡ”Π”Ο«±ΝΠ. Τδ÷–, –¬–ΆΒΡΝΫ«Ή–‘ΕύκΡΉιΉΑΖ÷Ή”ΒΡ…ηΦΤΚΆ÷Τ±ΗΓΔ ≤ΌΩΊΕύκΡΉιΉΑ–¬ΖΫΖ®ΒΡΫ®ΝΔΓΔ ΨΏ”–ΧΊ β–‘÷ ΚΆΫαΙΙΒΡΉιΉΑΧεΒΡΙΙΫ®ΓΔ ΕύκΡΉιΉΑΜζάμΒΡ…ν»κΫ“ Ψ“‘ΦΑ“‘ΉιΉΑΧεΈΣΙΙ÷ΰΜυ‘ΣΒΡΗΏ≤ψ¥ΈΉιΉΑΒ»―–ΨΩ±ΊΫΪ≥…ΈΣΝΫ«Ή–‘ΕύκΡΉ‘ΉιΉΑ―–ΨΩΝλ”ρ–¬ΒΡ»»ΒψΚΆΖΔ’ΙΖΫœρ, Ω…“‘‘ΛΤΎ, ΥφΉ≈ΕύκΡΉ‘ΉιΉΑΦΦ θΚΆΜυ¥Γάμ¬έ―–ΨΩΒΡ≤ΜΕœΖΔ’Ι, ΕύκΡΉ‘ΉιΉΑ±ΊΫΪΦΪ¥σΒΊΆΤΕ·…ζΟϋΩΤ―ßΓΔ …ζΈο“Ϋ―ß“‘ΦΑ≤ΡΝœ―ßΒΡ≥ΛΉψΫχ≤Ϋ.
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷ΨΓΘ
