
еЊвЊЃКЛљгкЖрыФКЭЕААзжЪЕФЫЎФ§НКЬхЯЕзїЮЊЩњЮяВФСЯЕФвЛИіживЊжжРр,ЪмЕНСЫЙуЗКЙизЂ,НќФъРДИќЕУЕНСЫ ГЄзуЕФНјВНЁЃЦфФ§НКЛЏЕФЛњжЦМШПЩвдЪЧВрЛљжЎМфЕФЮяРэзщзАКЭОлМЏ,вВПЩвдЪЧЗДгІадВаЛљжЎМфЕФПЩПиЛЏбЇЗД гІЁЃСНепаЭЌзїгУПЩНјвЛВНДйНјЦфЙІФмИФадЁЃЖрыФКЭЕААзжЪЖРгаЕФЩњЮяЛюадМАЦфНсЙЙКЭЙІФмЩЯЕФЖрбљадКЭПЩ ЕїадЮЊЩњЮяВФСЯЕФЗЂеЙЬсЙЉСЫжюЖрПЩФм,еЙЯжГіЙуЗКЕФгІгУЧАОАЁЃБОЮФЖдЛљгкЖрыФКЭЕААзжЪЕФЫЎФ§НКЕФЩшМЦЁЂжЦБИКЭгІгУЕШбаОПНјеЙНјааСЫМђвЊЕФНщЩмгызмНс,ВЂЖдИУСьгђЕФНјвЛВНЗЂеЙНјааСЫеЙЭћЁЃ
в§бдЃКВФСЯПЦбЇгыЩњУќПЦбЇЕФНЛВц,ЭЦЖЏСЫЩњЮяВФСЯЕФбИУЭЗЂеЙЁЃЫЎФ§НКЪЧвЛРрЗЧГЃживЊЕФЩњЮяВФСЯЁЃЫќ ЪЧгЩЧзЫЎадШ§ЮЌЭјзДНсЙЙгыДѓСПЫЎЙЙГЩЕФЖРЬиЗжЩЂЬхЯЕ,ФмЙЛдкЫЎжаШмеЭЖјВЛШмНтЁЃЖрЪ§ШэзщжЏОЭЪЧгЩ ЕААзжЪКЭОлЬЧЕШЩњЮяИпЗжзгзщГЩЕФЫЎФ§НКЁЃвђДЫ,ЫЎФ§НКдкЩњЮявНбЇСьгђ,ЬиБ№ЪЧвЉЮяЪЭЗХЁЂзщжЏЙЄГЬЁЂдйЩњвНбЇЁЂЩњЮяДЋИаЦїЁЂШсадДйЖЏЦївдМАШЫЙЄЯИАћЭтЛљжЪЕШЗНУцОпгаЙуЗКЕФгІгУЧАОА[1~3] ЁЃНќФъРД,Лљгк ЖрыФКЭЕААзжЪЕФЫЎФ§НК,вђЦфНсЙЙЩЯЕФОЋШЗадКЭЙІФмЩЯЕФЖрбљадЕУЕНЬиБ№ЕФЙизЂ,БОЮФНЋОЭИУЬхЯЕзіМђ вЊЕФНщЩмКЭзмНсЁЃ
1 БГОА
1.1 ЫЎФ§НК
ЙЙНЈЫЎФ§НКПЩвдЭЈЙ§ЮяРэНЛСЊКЭЛЏбЇНЛСЊЕФЗНЗЈ ЁЃЧАепЛљгкЗЧЙВМлМќЯрЛЅзїгУ , ШчОВЕчзїгУ ЁЂЪшЫЎ зїгУ ЁЂЧтМќЕШзїгУ;КѓепдђЪЧЛљгкЙВМлМќзїгУ ЁЃЦРМлЫЎФ§НКзїЮЊЩњЮяВФСЯЕФгІгУЧАОАЕФСНИіЗНУцЪЧЩњРэ ЛюадКЭЛњаЕадФм[4, 5] ЁЃЩњРэЛюаджївЊШЁОігкИпЗжзгЕФзщГЩ ЁЂЗжВММАЦфгыЯИАћжЎМфЕФЯрЛЅзїгУ;ЖјЛњаЕадФм жївЊШЁОігкНЛСЊЕуЕФЧПЖШКЭУмЖШ ЁЃСНепжБНггАЯьЯИАћЕФЩњГЄКЭЗжЛЏааЮЊ[3, 6] ЁЃНќФъРД , ЮЇШЦЫЎФ§НКЕФЙІ ФмЛЏКЭЛњаЕадФмИФадгПЯжСЫКмЖрГіЩЋЕФЙЄзї ЁЃвЛЗНУц , ШЫУЧЭЈЙ§МгШыФЩУзИДКЯЮяЛђДѓЗжзгОлМЏЬхВЮгыНЛ СЊ[7] , ИФБфНЛСЊЕуЕФаджЪ , НсКЯСНИіЩѕжСЖрИіаджЪЛЅВЙЕФЭјТч[8, 9] ЛђепРћгУВЛЭЌНЛСЊЛњРэаЭЌзїгУ[10, 11] ЕШЗНЗЈРДИФНјЫЎФ§НКЕФСІбЇадФм;СэвЛЗНУц , ЭЈЙ§в§ШыОпгаЩњРэЛюадЕФЛљдЊВЂПижЦЫќУЧдкЪБМфПеМфЩЯЕФ БэЯжРДИФНјЫЎФ§НКЕФЩњРэЛюад ЁЃзлКЯЕїПиетСНЗНУцЕФаджЪЪЧЫЎФ§НКбаОПЕФживЊЗНЯђ ЁЃ
1.2 ЖрыФКЭЕААзжЪ
ЖрыФКЭЕААзжЪОљЮЊвЛРргЩІС-АБЛљЫсвдыФМќЯрСЌЖјГЩЕФЛЏКЯЮяЁЃвЛАуАбКЌга100Иі(гаЪБЪЧ50Иі)вд ЩЯАБЛљЫсЕФЖрыФГЦЮЊЕААзжЪЁЃЫќУЧдкЬхФкЦ№ЕНСЫМЋЮЊживЊЕФЩњРэЙІФм,ПЩвдЫЕЪЧЙЙГЩЩњУќЕФЮяжЪЛљДЁЁЃ
ЫфШЛЦфзщГЩНіАќКЌ20жжЬьШЛАБЛљЫс,ЫќШДФмдкЩњУќЙ§ГЬжаЦ№ЕНИДдгЖрбљЕФЙиМќЙІФмЁЃетжївЊЙщвђгкЦфОЋШЗЕФађСаНсЙЙКЭЖрМЖЕФПеМфНсЙЙЁЃАБЛљЫсЩЯВЛЭЌаджЪЕФЙйФмЭХ(ЪшЫЎЁЂЧзЫЎЁЂЕчжаадЁЂЕчИКадЁЂМЋадЛљЭХ)ЕФЮЂУюЦНКтКЭЧЩУюНсКЯе§ЪЧЕААзжЪФмНјаагаађелЕўКЭЪЕЯжЖржжЙІФмЕФЙиМќЁЃ
ЖрыФПЩвдЭЈЙ§ЙЬЯрКЯГЩ,вВПЩвдЭЈЙ§ЕААзжЪНЕНтЕУЕНЁЃЖрыФЕФЙЬЯрКЯГЩЫфШЛжЛЪЪгУгаЯоГЄЖШЗжзгЕФКЯГЩ(ЭЈГЃаЁгк50ИіАБЛљЫс),ЕЋЪЧПЩвдЗНБуЕив§ШыИїжжЗЧЬьШЛАБЛљЫс[12] ЁЃЕААзжЪЕФЩњЮяКЯГЩдђЪЧгЩDNAЕФзЊТМгыЗвыЪЕЯжЕФвЛИіИпЖШОЋШЗПЩППЕФФЃАхОлКЯЗДгІЁЃЯжДњЛљвђЙЄГЬКЭжизщЕААзжЪЩњЮяКЯГЩЕФбИУЭЗЂеЙЮЊЕААзжЪЩњЮяВФСЯбаОПЬсЙЉСЫКмКУЕФЛљДЁ,ЖјЕААзжЪЙЄГЬИќНјвЛВНЭиеЙСЫЦфПЩаоЪЮадКЭЙІФмадЁЃР§Шч,schultzзщ[13,14]гыTirrellзщ[15,16]ГЩЙІНЋЗЧЬьШЛАБЛљЫсв§ШыЕААзжЪ,ЮЊЩшМЦгыВйзнЬиЪтЕААзжЪЬсЙЉСЫМЋДѓЕФБуРћЁЃЖЈЕуЭЛБфКЭЖЈЯђНјЛЏЕШММЪѕЕФГЩЪьИќШнаэШЫУЧЖдгкЭЌвЛжжЕААзжЪЕФаджЪНјааИФНјКЭЮЂЕї,ЦфНсЙЙКЭЙІФмПижЦЕФОЋШЗЫЎЦНЪЧКЯГЩИпЗжзгЬхЯЕРяЫљВЛФмБШФтЕФЁЃ
2 ЛљгкЖрыФ/ЕААзжЪ-КЯГЩИпЗжзгЕФЫЎФ§НКЬхЯЕ
КЯГЩИпЗжзгЫфШЛВЛОпгаЩњЮяДѓЗжзгЕФНсЙЙОЋШЗЖШ , ЩњРэЙІФмвВЯрЖдгаЯо , ЕЋЪЧЦфЕЅЬхНсЙЙИДдгЖрбљ , вВБШНЯШнвзаоЪЮВЂЪЕЯжСПВњЁЃЯжДњПЩПиОлКЯММЪѕЕФЗЂеЙИќЬсЙЉСЫПижЦКЯГЩИпЗжзгЗжзгСП ЁЂЗжЩЂЖШЕШживЊ НсЙЙВЮЪ§ЕФЖржжгааЇЗНЗЈЁЃвђЖј , КЯГЩИпЗжзггыЖрыФКЭЕААзжЪдкзїЮЊВФСЯЗНУцОпгаКмЖрЛЅВЙЕФгХЕу , Цф ИДКЯЫЎФ§НКЬхЯЕвВБИЪмЙизЂЁЃMichaelМгГЩ ЁЂЕуЛїЛЏбЇ[17] ЕШвЛЯЕСаПьЫйИпаЇЕФЛЏбЇаоЪЮЪжЖЮЮЊДДдьИДКЯ ЫЎФ§НКЬсЙЉСЫКЯЪЪЕФЪжЖЮ , МмЦ№СЫКЯГЩЬхЯЕКЭЩњЮяЬхЯЕжЎМфЕФЧХСК , ДйНјСЫетИіЗНЯђЕФХюВЊЗЂеЙЁЃ
2.1 ЖрыФPИпЗжзгЫЎФ§НК
ЖрыФвђЦфКЯГЩКЭаоЪЮЕФМђБуадБЛЙуЗКЕигІгУгкжЦБИИДКЯЫЎФ§НКЁЃЖрыФжЎМфЕФгаађОлМЏПЩвдаЮГЩЮяРэНЛСЊЁЃР§Шч,ОэЧњТна§ШчССАБЫсРСДЪЧгЩІС-Тна§вђЪшЫЎзїгУаЮГЩЕФЖрОлЬх(ШчЭМ1aЫљЪО)ЁЃеддЊЕмКЭСѕВЈЕШНЋЦфНгЕНОлввЖўДМСД(PEG)ЕФвЛЖЫ,СэвЛЖЫСЌЩЯБћЯЉЫсѕЅЁЃдкЙтКЭв§ЗЂМСзїгУЯТ,ИУДѓЗжзгЕЅЬхОлКЯаЮГЩНгжІИпЗжзгЫЂ,ВрСДЕФССАБЫсРСДОлМЏОЭаЮГЩСЫЮяРэФ§НК[18] ЁЃІТ-ЦЌВуНсЙЙЪЧвЛжжЛљгкЧтМќЕФОлМЏзїгУ (ШчЭМ1bЫљЪО)ЁЃKopecekзщЩшМЦСЫвЛРрЫЎНтОлТэРДЫсєћ(HPMA)-ЖрыФЕФНгжІЙВОлЮя[19] ЁЃЦфжаСНжжЖрыФПЩвдЛЅВЙаЮГЩІТ-ЦЌВуНсЙЙ,ДгЖјЪЙећИіЬхЯЕФ§НКЛЏ,ВЂФмзїЮЊЕїПиЙЧЯИАћЗжЛЏвдМАєЧЛљСзЛвЪЏПѓЛЏЕФЩњЮяВФСЯЁЃЭЌЪБ,ЖрыФЩЯЕФЙйФмЭХвВПЩвдБЛгУгкЪЕЯжЙВМлНЛСЊ,Р§ШчКЌгаЗДгІадЙйФмЭХЕФЖрыФФмКЭКЌгаЯргІЗДгІадЙйФмЭХЕФКЯГЩИпЗжзгНјааНЛСЊжЦБИЫЎФ§НКЁЃ
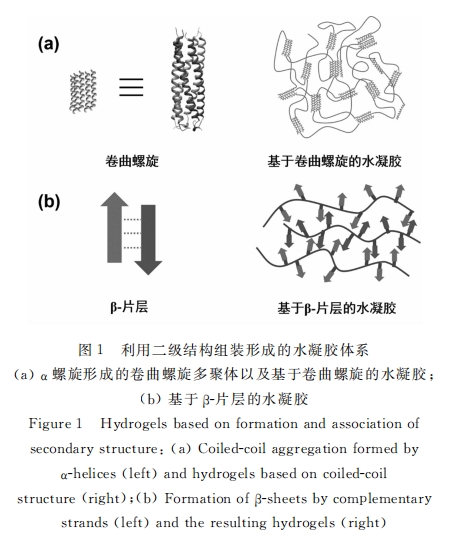
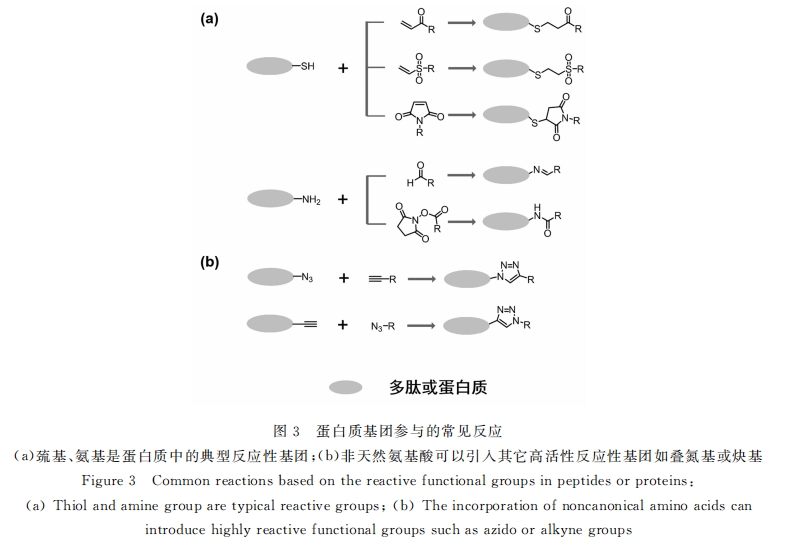
PEGЪЧвЛРр Оп га Км КУ Щњ Юя Яр Шн ад ЕФ ЫЎШмадКЯГЩИпЗжзг , БЛЙуЗКгІгУгкЩњЮяИпЗжзгЕФИФадКЭжЦБИЫЎФ§НКЁЃЛљгк PEG ЕФЫЎФ§НКвђЖјБЛШЯЮЊЪЧЫЎ Ф§НКЬхЯЕЕФЁАН№БъзМЁБЁЃвбгаКмЖрЙЄзїНЋ PEG КЭЖрыФЛђЕААзжЪНсКЯдквЛЦ№жЦБИЫЎФ§НКЁЃзюГЃгУЕФЗДгІЪЧ ЛљгкАыызАБЫсЕФлЯЛљЛђепРЕАБЫсЕФАБЛљЕФЗДгІЁЃР§Шч , HubbellзщЭЈЙ§ PEG ЕФввЯЉэПЖЫЛљгыКЌгаОЋАБ Ыс-ИЪАБЫс-ЬьЖЌАБЫсађСа(RGD)ЖрыФЦЌЖЮЩЯЕФлЯЛљжЎМфЕФ MichaelМгГЩЪЕЯжФ§НКЛЏ , ЗЂеЙжЇГжЯИАћ№ЄИН гыУИНЕНтЕФЯИАћЭтЛљжЪ[20~22] ЁЃИУЬхЯЕжаЛЙв§ШыСЫФмЙЛБЛЛљжЪН№ЪєЕААзУИ(MMP)ЛђбЊНЌУИЬивьадНЕНт ЕФЖрыФађСа ЁЃВЛЭЌЖрыФађСаЕФНЕНтЫйТЪВЛЭЌ , вдДЫОЭПЩвдЕїНкЬхЯЕЕФНЕНтЫйТЪЁЃИУЬхЯЕБЛГЩЙІгІгУдкЯИАћХрбјЙ§ГЬ ЁЃНсЙћБэУї , ЯИАћдкИќШнвзБЛНЕНтЕФЫЎФ§НКжаЪцеЙКЭЧЈвЦЕФЫйЖШИќПь ЁЃ
ЮЊСЫОЁСПМѕЩйФ§НКЛЏЙ§ГЬЖдгкЩњЮяЗжзгвдМАЯИАћЕФгАЯьВЂИќКУЕиПижЦФ§НКЛЏЙ§ГЬ,ЩњЮяе§НЛЕФЕуЛї ЛЏбЇБЛЙуЗКЕигІгУЕНЫЎФ§НКЕФжЦБИжаРДЁЃkopeekзщЖдPEGНјааЛЏбЇаоЪЮЕУЕНЖЫЛљЮЊЕўЕЊЕФЫФБл PEG,ВЂКЯГЩСЫЖЫЛљЮЊШВВЂКЌгаУИНЕНтЦЌЖЮЕФЖрыФGFLG1,РћгУШВЛљКЭЕўЕЊжЎМфЕФЕуЛїЗДгІ,ЪЕЯжСЫ PEGгыЖрыФЕФИДКЯЫЎФ§НК[23] ЁЃРрЫЦЕи,AnsethзщРћгУЕуЛїЩњЮяе§НЛЗДгІжЦБИСЫвЛЯЕСаPEGФ§НК,ВЂАб живЊЕФЩњЮяаХКХЙЬЖЈЕНЭјТчжаШЅ,вддіЧПвШЯйІТ-ЯИАћдкЫЎФ§НКжаЕФЗтзАГЩЛюТЪ[24] ЁЃЫћУЧИќНјвЛВНПЊДД адЕиНјааСЫНЋPEGЫЎФ§НКадФмдкШ§ЮЌПеМфжаЭМАИЛЏЕФбаОП[25,26] ЁЃБШШчЯѕЛљБНМзУббмЩњЮяЪЧвЛРрЙуЮЊШЫ жЊЕФЙтНЕНтЛљЭХ(ШчЭМ2ЫљЪО)ЁЃдкЖрыФНЛСЊЗжзгжав§ШыЯѕЛљБНМзУббмЩњЮяКѓ,ОЭПЩгЩЙтПЬММЪѕЕїПиЦфЙй ФмЛЏзДЬЌЁЃЕБдкИУНсЙЙФЉЖЫМгШыКЌRGDЦЌЖЮЪБ,ЙтееЛсЕМжТЕФЖрыФаХКХЪЭЗХ,етОЭПЩвдЗНБуЕиПижЦБЛЗт зАЯИАћЕФЧЈвЦКЭЗжЛЏ[26] ЁЃРћгУМгГЩЗДгІЕФЙйФмЛЏгыРћгУЯѕЛљБНМзУбЕФПЩПиНЕНтПЩвдЭЈЙ§ВЛЭЌВЈГЄЕФЙт МЄЗЂРДЗжБ№ЪЕЯжЁЃЫћУЧГЩЙІЕиНЋИУЗНЪНгІгУгкЫЎФ§НКадФмЕФШ§ЮЌЭМАИЛЏ,ЪЙГЩЯЫЮЌЯИАћФмЙЛбизХRGDЙйФмЛЏЕФПеМфНјааЩњГЄ[27,28] ЁЃетЮЊЕїПиЯИАћдкШ§ЮЌПеМфжаЕФЩњГЄЬсЙЉСЫЧПгаСІЕФЪжЖЮЁЃ
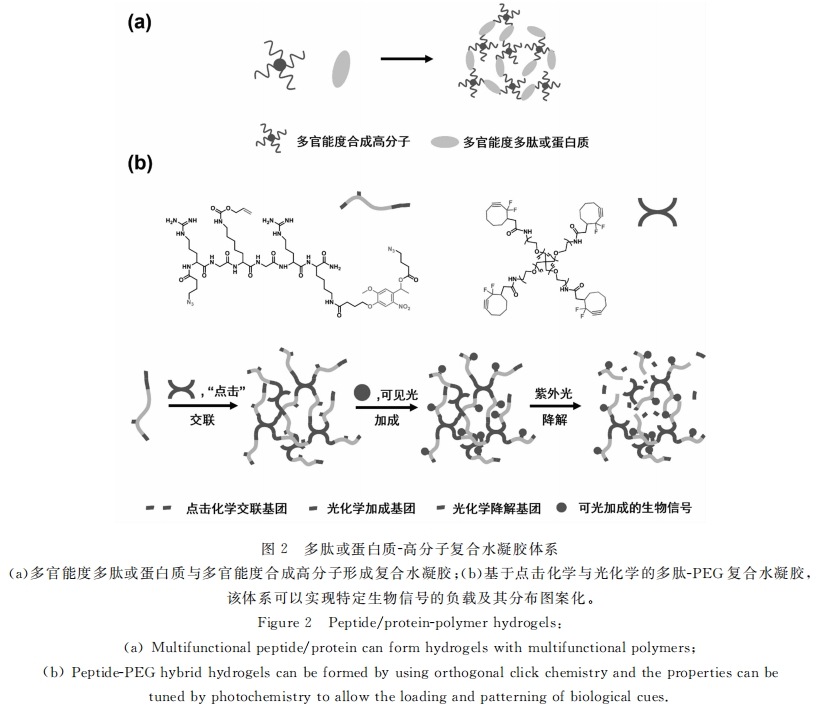
ЯрБШгкЕуЛїЛЏбЇ , УИДпЛЏЕФПЩПиХМСЊвВаэКЭЯИАћЙ§ГЬИќЮЊМцШн ЁЃБШШчЙШАБѕЃАЗзЊАЗУИПЩвдгааЇЕиДп ЛЏЙШАБѕЃАЗКЭРЕАБЫсжЎМфЕФвьыФМќаЮГЩ ЁЃEhrbarгы LutolfЁЂweberЕШжЦБИСЫСНжжАЫБл PEG, ЦфФЉЖЫЗжБ№ аЏДјСНРрЖрыФ:вЛРрКЌгаЙШѕЃАЗЪмЬхЕзЮя(Gln),вЛРрЪЧКЌгаЙЉЛљжЪН№ЪєЕААзУИНЕНтЕФЖрыФађСаЕФРЕАБЫс ЕзЮя(Lys)ЁЃСНжжДѓЗжзгдкЙШАБѕЃАЗзЊАЗУИЕФзїгУЯТЙВМлЯрСЌ , ОЭПЩвдаЮГЩЭјТчНсЙЙ ЁЃвђЮЊетИіЗНЗЈвВ ЪЪгУгкКЭЩњЮяДѓЗжзгЕФЙВМлХМСЊ , ЫљвдИУИДКЯЫЎФ§НКвВПЩБЛгУгкЙЬЖЈКЌгаЬиЖЈБъЧЉЕФШкКЯЕААз , БШШчВЖЛёШЫХпЩіЯИАћ(HEK-293T)ЗжУкГіЕФБъЧЉЩњГЄвђзг ЁЃМгдиСЫЩњГЄвђзгЕФЫЎФ§НКЬхЯЕПЩвдзїЮЊжИЕМЯИАћ ааЮЊЕФжЧФмШЫЙЄЯИАћЭтЛљжЪ[29] ЁЃ
2.2 ЕААзжЪPИпЗжзгЫЎФ§НК
ЛљгкЕААзжЪКЭКЯГЩИпЗжзгЕФЫЎФ§НККЭЖрыФ-ИпЗжзгЫЎФ§НКгаКмЖрЕФЯрЫЦад , ЦфаЮГЩЭЌбљМШПЩвдвРПП ЕААзжЪЛљдЊжЎМфЕФЮяРэОлМЏ , вВПЩвдРћгУЕААзжЪЕФЕфаЭЗДгІадЛљЭХ , БШШчРЕАБЫсЕФАБЛљ ЁЂАыызАБЫсЕФлЯЛљ ЕШ , ЩѕжСЪЧЗЧЬьШЛАБЛљЫсжаЕФИпЗДгІадЛљЭХ , ШчЕўЕЊЛљКЭШВЛљЕШ ЁЃлЯЛљРДдДгкЬьШЛАБЛљЫс , ЫќКЭЫЋМќжЎМф ЕФ MichaelМгГЩЗДгІдкЩњРэЬѕМўЯТОпгаНЯИпЕФбЁдёадКЭЗДгІад , ЖдЕААзжЪЩњРэЛюадЕФгАЯьНЯЩй , ЫљвдвВ зюЮЊГЃгУ(МћЭМ 3)ЁЃHubbellзщЩшМЦСЫФЉЖЫОпгаввЯЉЛљэПНсЙЙЕФ PEG, гыКЌгаЖрИіАыызАБЫсЕФжизщЯЫЮЌ ЕААздЕААзжЪзїЮЊНЛСЊЗжзг , РћгУ MichaelМгГЩЗДгІдЮЛаЮГЩЫЎФ§НКЗтзАЫЎФ§НК , ДгЖјНвЪОСЫВФСЯКъЙл ЕФСІбЇадФмЪЧБШ RGDађСаУмЖШИќЮЊживЊЕФгАЯьЯИАћЧЈвЦЫйТЪЕФвђЫи[30, 31] ЁЃyang зщЩшМЦВЂБэДяСЫОпга РрЗКЫиНсЙЙгђ(ULD)ЕФЕААзжЪ[32] ЁЃИУЕААзжЪФмЙЛаЮГЩЫФОлЬх , УПВПЗжЬсЙЉвЛИіПЩЙЉЗДгІЕФлЯЛљ , ПЩзїЮЊ ЫФЙйФмЖШЕФжизщЕААзжЪгыЖЫЛљЮЊТэРДѕЃбЧАЗЕФЫФБл PEGНјаа MichaelМгГЩЗДгІНЛСЊ ЁЃлЯЛљгыВЛЭЌРраЭ ЕФЫЋМќЗДгІЛюадВЛвЛ , ЫйТЪвВВЛОЁЯрЭЌ , ШнаэШЫУЧЖдЗДгІЕФЖЏСІбЇКЭФ§НКЪБМфгаНЯКУЕФПижЦ ЁЃЭЌЪБ , гЩгк НЛСЊЕФЮЛЕуШЗЖЈ , дкЭјТчжаЕФЗжВМКЭОрРыОљдШ , ЖдгкЛњаЕадФмвВгаКмКУЕФДйНјзїгУ ЁЃ
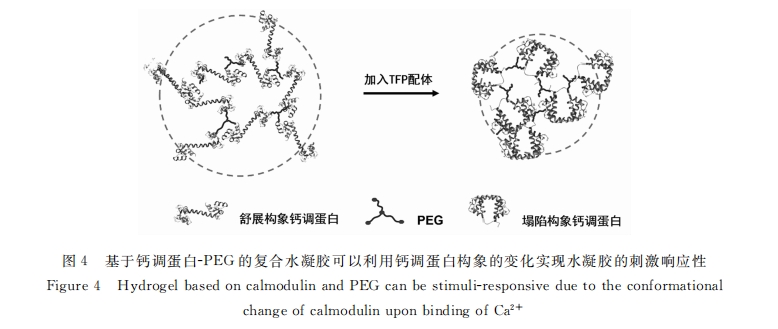
етжжПЩПиЕФФ§НКЛЏЙ§ГЬКЭЕААзжЪЕФЙЙЯѓБфЛЏКЭЙІФмзЊЛЛНсКЯдквЛЦ№ , ОЭгаПЩФмЪЕЯжДЬМЄЯьгІадЫЎФ§ НК ЁЃР§Шч , ИЦЕїЕААз(calmodulin, caM)ЪЧвЛжжНЯЮЊЪцеЙЕФ , ОпгабЦСхаЮзДЕФЕААзжЪ , дк ca2ЪЎ РызгДцдкЕФ ЧщПіЯТ , ЫќЕФЙЙЯѓЗЂЩњЬЎЫњ , вдБугкгывЛаЉаЁЗжзг(ШчШ§ЗњРрК TFP)НјааНсКЯ[33] ЁЃMurphy зщРћгУлЯЛљ гыБћЯЉЫсѕЅдкзЯЭтЙтЯТЕФ MichaelМгГЩЗДгІЙЙНЈГіСЫвЛжжИЦРызгХЈЖШЯьгІЕФИЦЕїЕААз-ИпЗжзгИДКЯЫЎФ§ НК[34] (МћЭМ 4)ЁЃдкИЦРызгКЭ TFP ЕФЛЗОГЯТ , Ф§НКбИЫйЪеЫѕ;ШчЙћНЋЦфжУгк EDTA ЛЗОГЯТ , дђЛсШмеЭВЂЪЭ ЗХГі TFPЁЃетжжгЩЕААзжЪЙЙЯѓБфЛЏЫљДјРДЕФКъЙладжЪзЊБфЙ§ГЬПЩвдбЛЗЗДИД , ЖрДЮНјаа , ГфЗжеЙЪОСЫЕА АзжЪдкЪЕЯжДЬМЄЯьгІадКЭжЧФмЛЏЫЎФ§НКЬхЯЕЗНУцЕФОоДѓгІгУЧАОА ЁЃ
КЭЖрыФЯрБШ , ЕААзжЪЕФЩњРэЛюадЖдЦфЙЙЯѓИќЮЊУєИа , дкРћгУЗДгІадВаЛљНјааЙйФмЛЏЙ§ГЬжаШнвзЪЇЛю ЁЃУИДпЛЏЕФИпаЇХМСЊПЩвдЙцБметИіШБЕу ЁЃРћгУФкКЌыФКЭзЊАБУИЖМПЩФмЪЕЯжетжжНЛСЊ ЁЃLutolfзщдђбЁгУСЫвЛИіЩдЮЂВЛЭЌЕФВпТд[35] (МћЭМ 5)ЁЃвбжЊввѕЃИЈУИ A(CoA)ЕФСзЫсЗКѕЃлЯЛљввАЗВПЗждкСзЫсЗКѕЃлЯЛљвв АЗЛљзЊвЦУИ(PPTase)ЕФДпЛЏзїгУЯТ , ФмгыЫПАБЫсЕФєЧЛљЙВМлЯрСЌ ЁЃЫљвд , ЫћУЧОЭЩшМЦСЫЭбѕЃЛљдиЬхЕААз ЖўОлЬх(ACP-ACP)вдМАФЉЖЫЮЊввѕЃИЈУИ A ЕФНгжІЖрБл PEGСД(CoA-PEG),СНепдк PPTaseДпЛЏЯТНЛСЊ аЮГЩЫЎФ§НК ЁЃетЪЧвЛИіИпЖШЩњЮяЯрШнадЕФЗДгІ , ЭЌЪБШнаэЙВМлМгдиКЌгаОЋАБЫс-ИЪАБЫс-ЬьЖЌАБЫс-ЫПАБЫс №ЄИНађСа(RGDS)ЕФЖрыФЦЌЖЮ , гааЇжЇГжБЛЗтзАЯИАћдкШ§ЮЌПеМфФкЕФЩњГЄ ЁЃ
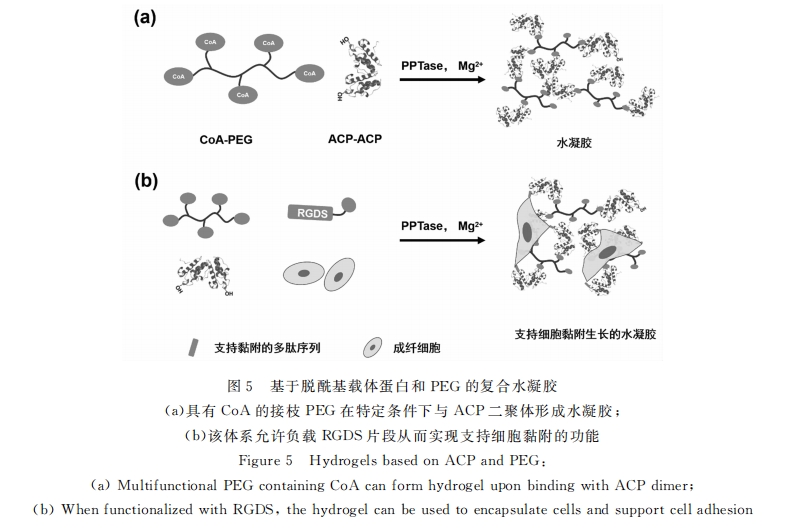
ЕААзжЪЯрЛЅзїгУвВЪЧЙЙНЈИДКЯЫЎФ§НКЕФгааЇЧ§ЖЏСІ ЁЃзщБЈЕРСЫРћгУОэЧњТна§ОлМЏНЛСЊЕФ ЕААзжЪ-ИпЗжзгИДКЯЫЎФ§НК[36] ЁЃИУЬхЯЕзщГЩЮЊ N-(2-єЧБћЛљ)-МзЛљБћЯЉѕЃАЗКЭ N-(NЁЏ, NЁЏ-ЖўєШМзЛљАББћ Лљ)-МзЛљБћЯЉѕЃАЗСНжжЕЅЬхЕФЙВОлЮя(HPMA-co-DAMA),вдМАОпгаОэЧњТна§(coiled-coil)НсЙЙЕФСНжжЕА АзжЪ CC1 гы CC2ЁЃЦфжа CC1 РДдДгкЧ§ЖЏЕААзЕФОЅВПЧјгђ(~150ИіАБЛљЫс), CC2дђЪЧЛљгкДгЭЗЩшМЦЕФ ЖрыФСД(~50ИіАБЛљЫс),ОпгаМЋИпЕФШШЮШЖЈад ЁЃЕААзжЪгыКЯГЩИпЗжзгжЎМфЪзЯШЭЈЙ§зщАБЫсБъЧЉКЭИпЗж згВрСДєШЛљжЎМфЕФ Ni2ЪЎ РызгХфЮЛТчКЯНгжІЯрСЌ , ЖјОэЧњТна§ЕФНјвЛВНОлМЏдђгааЇЕМжТСЫЮяРэФ§НКЕФаЮ ГЩ ЁЃетжжЗНЗЈОпгавЛЖЈЕФЦеЪЪад , вВЪЪгУгкБШШч zn2ЪЎ ЁЂCa2ЪЎ ЁЂMg2ЪЎ ЕШРызг , НЋгаПЩФмдЪаэЬхЯЕзїЮЊЩњЮяЛЗОГЯТЖдН№ЪєРызгЯьгІЕФЮЂаЭДЋИаЦї ЁЃ
Г§СЫОэЧњТна§жЎЭт , ЖўМлРызгФЭЪмЕААзжЪ(cutA)вВЪЧзюНќбаОПжаБШНЯШШУХЕФФмЙЛНјаагаађОлМЏЕФ ЕААзжЪ(МћЭМ 6a)ЁЃЫќРДдДгкГЌЪШШШЙХОњ pyrococcus horikoshii , ФмаЮГЩМЋЦфЮШЖЈЕФШ§ОлЬх[37] ЁЃIto зщЩш МЦСЫвЛжжгЩШ§ЙйФмЖШ cutA КЭЫФЙйФмЖШ PEG НЛСЊЖјГЩЕФЫЎФ§НКЬхЯЕ , ВЂдк cutA Ш§ОлЬхЕФ c ЖЫШкКЯСЫ TAXЯрЛЅзїгУЕААзжЪ I(Tip-1)ЁЃTip-1ОпгавЛЖЮХЬзДЭЌдДНсЙЙгђ(PDZ),ФмЙЛгыЯргІЕФЖрыФЖЮЪЖБ№ВЂНс КЯ ЁЃЫћУЧЭЌЪБКЯГЩСЫОпгаЪЖБ№ PDZНсЙЙгђЖрыФЖЫЛљЕФЫФБл PEG Зжзг ЁЃСНепздзщзАОЭаЮГЩОпгаЙцећЭј ТчЕФЫЎФ§НК ЁЃСїБфВтЪдЗЂЯж , ИУЬхЯЕГіЯжЕфаЭЕФМєЧаБфЯЁЯжЯѓ , ГЗШЅаЮБфжЎКѓЦфЛњаЕЧПЖШМДПЬЛиИД ЁЃет ПЩвдгЩ PDZНсЙЙгђгыЯргІЖрыФдкИпМєЧаСІЯТПЩФцЕФЗжРыКЭНсКЯРДНтЪЭ[38] ЁЃ
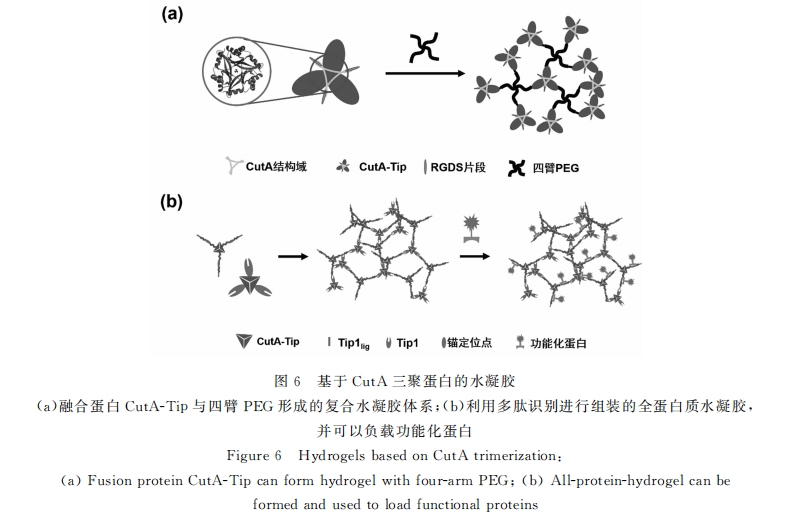
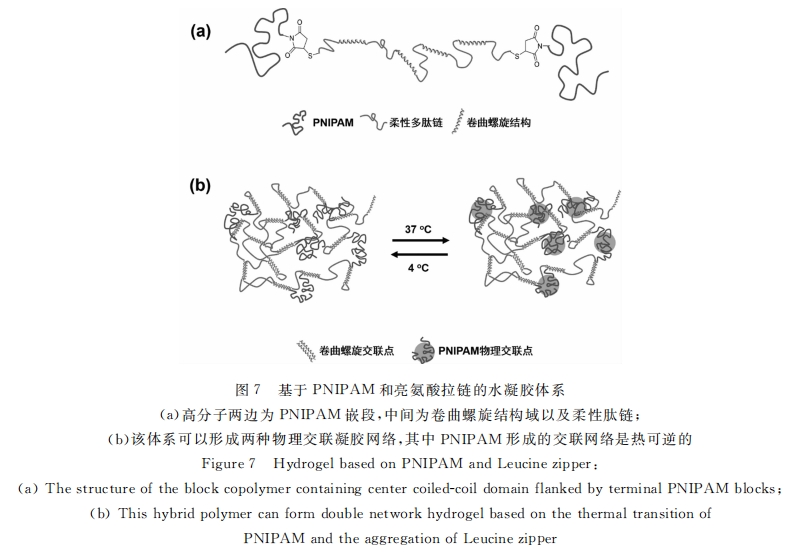
ЪЕМЪЩЯ , РћгУЕААзжЪЯрЛЅзїгУЪЕЯжЮяРэНЛСЊжЦБИЕФИДКЯЫЎФ§НКЕФСІбЇ адФмШдОпгаЯрЕБДѓЕФЬс Щ§ Пе Мф ЁЃНсКЯВЛЭЌЕФЮяРэНЛСЊЛђЛЏбЇНЛСЊЛњРэаЕїзїгУОЭЪЧвЛжжЗЧГЃКУЕФВпТд[39] ЁЃolsenзщНЋЕААзжЪ-ЕААз жЪЯрЛЅзїгУКЭИпЗжзгЕФЯрзЊБфНсКЯдквЛЦ№ , ЙЙНЈСЫОпгаСНжжНЛСЊЭјТчЕФЫЎФ§НК[40] (МћЭМ 7)ЁЃИУЧЖЖЮИДКЯ ЮяСНЖЫОпгаОл(N-вьБћЛљБћЯЉѕЃАЗ)(PNIPAM),Щ§ЮТКѓФмЙЛОлМЏаЮГЩЮяРэНЛСЊ , ЖјжаМфЕФЕААзжЪдђЭЈЙ§ ОэЧњТна§аЮГЩЕкЖўжжЮяРэНЛСЊ ЁЃетжжЫЋЭјТчНсЙЙЪЙЕУећИіЬхЯЕЕФСІбЇадФмгыПЙИЏЪДадФмЖМгаСЫМЋДѓЕФИФЩЦ ЁЃ
змЕФРДЫЕ , жЦБИЕААзжЪ-ИпЗжзгИДКЯЫЎФ§НК , жївЊЪЧЮЊСЫРћгУЕААзжЪЕФЩњЮяЛюадКЭКЯГЩЬхЯЕЕФвзжЦБИ КЭвзаоЪЮад , етРрЬхЯЕОпгавЛЖЈЕФДйНјЯИАћЩњГЄгыЗжЛЏЕФФмСІ ЁЃФПЧАЕФЬєеНжївЊдкгкНјвЛВНПЊЗЂКЭРћгУ гыЩњЮяЙ§ГЬЯрШнЕФЛЏбЇЗДгІ , дкзюДѓГЬЖШБЃжЄЕААзжЪЩњРэЛюадЕФЛљДЁЩЯЪЕЯжФ§НКЛЏ , ВЂНјвЛВНИФНјЛњаЕ адФм , ЪЕЯжЫЎФ§НКЙІФмЖрбљЛЏ ЁЂЭМАИЛЏКЭжЧФмЛЏ , ИќКУЕиФЃФтЬьШЛЯИАћЭтЛљжЪЕФЬиад , ПЊЭиЦфдкзщжЏЙЄГЬ жаЕФдЫгУ ЁЃ
3 ЭъШЋЛљгкЖрыФКЭЕААзжЪЕФЫЎФ§НКЬхЯЕ
ЖрыФЛђЕААзжЪгыКЯГЩИпЗжзгЕФИДКЯЫЎФ§НКЬхЯЕНсКЯСЫСНепЕФгХЕу , ЕЋЪЧдкВЛгАЯьЩњРэЛюадЕФЧАЬсЯТ ЪЕЯжПЩПиХМСЊЕФЗНЗЈЯрЕБгаЯо ЁЃЭЌЪБ , ЖдгкДѓЖрЪ§ЬхФкгІгУРДНВ , КЯГЩИпЗжзгЕФЩњЮяЯрШнадМАПЩНЕНтаджЪЕШвВашвЊЩїжиПМТЧ ЁЃвђДЫ , ЭъШЋЛљгкЖрыФЛђЕААзжЪЕФЫЎФ§НКЬхЯЕГЩЮЊвЛИіЗЧГЃЮќв§ШЫЕФЗНЯђ ЁЃЫќШЁЗЈздШЛ , ФмЙЛИќКУЕиЖдЩњЮяЬхЯЕНјааФЃФт ЁЃ
3.1 ЖрыФЫЎФ§НК
ЖрыФЕФздзщзАЪЧЗсИЛЖрВЪЕФЁЃЖрыФЕФзщГЩВЂВЛЯогкДЋЭГЕФ20жжЬьШЛАБЛљЫс,ЛЙАќРЈИїжжЗЧЬьШЛАБЛљЫсЁЃЭЌЪБ,МДЪЙНіКЌгаЪ§ИіВаЛљ,ЖрыФШдШЛПЩвдеЙЯжЧПСвЕФзщзАЯжЯѓ,аЮГЩгаађНсЙЙЁЃаьБјКЭбюжОФБЕШ[41]ЙЙНЈСЫDЙЙаЭЕФЫФыФ,етжжЫФыФОпганСЛљЖЫЛљADAБНБћАБЫсADAБНБћАБЫсADAРЕАБЫсADAРвАБЫсЕФНсЙЙ,ФмЙЛНсКЯзЯЩМДМЕШЩњЮяЛюадЗжзгЁЃСзЫсЛЏЕФЖрыФЧАЬхвЛОШЅСзЫсЛЏ,ОЭФмзщзАаЮГЩОпгаЯЫЮЌЙЧМмЕФФ§НКЁЃгЩгкDаЭАБЛљЫсгавЛЖЈЕФПЙНЕНтФмСІ,ИУЬхЯЕФмЙЛГЄЪБМфБЃГжЮШЖЈ,ВЂЛКТ§ЪЭЗХвЉЮяЁЃulijnЕШ[42]НіРћгУвЛжжФЉЖЫЪЧмЬМзбѕєЪЛљ(Fmoc)ЕФЖўыФ,ОЭЪЕЯжСЫЖрыФФ§НКЛЏЙ§ГЬ,ЦфжаІа-ІаЯрЛЅзїгУБЛШЯЮЊЪЧФ§НКЛЏЕФЙиМќЁЃЫцКѓGoughгыulijnЕШНЋмЬМзбѕєЪЛљ-ЖўБНБћАБЫсЖрыФ(FmocFF)гымЬМзбѕєЪЛљ-ОЋАБЫс-ИЪАБЫс-ЬьЖЌАБЫсЖрыФ(Fmoc-RGD)ЕФЫЎШмвКЛьКЯ,вВЕУЕНСЫОпгаФЩУзЯЫЮЌНсЙЙЕФЫЎФ§НК,ВЂПЩгУгкдіЧПЗтзАдкЦфжаЕФШЫРрецЦЄГЩЯЫЮЌЯИАћЕФдіжГгыЩњГЄ[43,44] ЁЃ
ЖрыФвВПЩвдЭЈЙ§ИќЮЊИДдгЕФЖўМЖНсЙЙЕФЯрЛЅзїгУЪЕЯжФ§НКЛЏЁЃЖрыФІТелЕўЕФгаађзщзАТЪЯШМЄЗЂСЫбаОПепЖдЖрыФФ§НКЕФбаОПЁЃР§Шч,Опга16ИіЙШАБЫс(E)-ИЪАБЫс(A)-РЕАБЫс(K)жиИДЕЅдЊЕФЖрыФ(EAK16),ПЩвдЭЈЙ§ЖрыФСДМфЧтМќзїгУаЮГЩІТелЕўЕФЦНУц,етаЉЦНУцдйЭЈЙ§ВаЛљМфЕФОВЕчзїгУНјааЖбЕў,ДгЖјаЮГЩЬиЖЈЕФШ§ЮЌНсЙЙ[45~46] ЁЃСэвЛжжIsKЕААзжаЕФзЊФЄНсЙЙгђОпгаИЛКЌРЕАБЫсЕФЖрыФЦЌЖЮ,вВФмЙЛЛљгкЩЯЪіЗНЪНаЮГЩФ§НК[47] ЁЃдкИУЙЄзїжа,AggeliЕШАбФмЙЛЗЂЩњФ§НКЛЏЕФЖрыФЬхЯЕЕФРэадЩшМЦзмНсЮЊШ§Иіддђ:(1)ВЛЭЌСДЕФВрЛљМфвЊОпгаЮќв§СІ(ШчЪшЫЎЯрЛЅзїгУЁЂОВЕчЯрЛЅзїгУЁЂЧтМќзїгУЁЂІа-ІаЖбЕўЕШ);(2)ыФСДЖЮВрЛљдкФГЗНЯђЩЯ(ЭЈГЃЮЊДЙжБгкІТелЕўЦНУцЕФЗНЯђ)ОпгавЛЖЈЕФЪЖБ№зїгУ,ПЩвдЯожЦвЛИіЗНЯђЩЯЕФздзщзА;(3)НсЙЙБэУцвЊЖдШмМСЗжзггаНЯЧПЕФЮќИНад[47] ЁЃ
етаЉЙцТЩжИЕМСЫздзщзАЖрыФЕФНјвЛВНРэадЩшМЦЁЃMessersmithЕШбЁдёСЫОпгаНЛЬцЪшЫЎЧзЫЎВаЛљЕФ ЖрыФЦЌЖЮ,ЦфађСаЮЊH2N-(FEFEFKFK)2-COOH,МђГЦЮЊFEK16ЁЃдкШШЛђЙтв§ЗЂЯТ,гЩжЌжЪЬхИКдиЕФТШ ЛЏИЦБЛЪЭЗХ,ОЭМЄЗЂСЫЖрыФЕФздзщзАаЮГЩФ§НК,етжжЗНЪНГЩЙІЪЕЯжСЫФ§НКЛЏЙ§ГЬЕФЙтПиЛђШШПи[48] ЁЃSchneiderЕШЩшМЦСЫОпга(VK)4VDPPTK(VK)3V-NH2 ађСаЕФЖрыФСД[49] ,ЫќдкЫсадЯТЪЧЮоЙцЯпЭХ,дкМю адЬѕМўЯТдђаЮГЩІТ-ЗЂМа,здзщзАГЩЮЊЮШЙЬЕФФ§НКЭјТчЁЃетжжЙЙЯѓЕФзЊБфЫцзХpHЕФЩ§ИпЖјЗЂЩњ,ЦфЫйТЪ гЩЖрыФХЈЖШКЭЛьКЯЫйТЪОіЖЈЁЃЛљгкІС-Тна§вдМАОэЧњТна§НсЙЙЕФФ§НКЬхЯЕвВЭЌбљЕиБэЯжГіpHУєИаадЁЃHartgerinkЕШ[50]ЩшМЦСЫФмаЮГЩІС-Тна§НсЙЙЕФЖрыФЦЌЖЮEIKQLESEISKLEQEIQSLEK,ИУТна§ЕФeКЭg ЮЛжУЮЊЙШАБЫс(E)ЁЃдкpHЩ§ИпЪБ,ЙШАБЫсШЅжЪзгЛЏ,дкОВЕчГтСІЕФзїгУЯТТна§НтОл,ЖрыФФ§НКОЭЛсШмНтЁЃ
ЖрыФЫЎФ§НКЕФНјвЛВНЙІФмЛЏвВЪЧбаОПепЧрэљЕФШШЕу ЁЃеХЪяЙтЕШ[51] ЗЂЯжОпгаФЩУзЯЫЮЌНсЙЙЕФЖрыФЫЎ Ф§НКЬхЯЕПЩвдАќВиВЂЛКЪЭШмОњУИ ЁЂУтвпЧђЕААз G ЕШВЛЭЌНсЙЙВЛЭЌЕчадЕФЕААзжЪ , ЦфЛКЪЭЫйЖШШЁОігкЕААз жЪДѓаЁЖјЗЧЕААзжЪЕчад , ВЂЧвЪЭЗХЕФЕААзжЪШдПЩвдБЃГждРДЕФИпМЖНсЙЙ[52] ЁЃетЫЕУїЗНЗЈОпгавЛЖЈЕФЦе ЪЪад , ЛЛбджЎ , ЭЈЙ§КЯРэЩшМЦЕїећЫЎФ§НКЕФЙЧМмГпДч , ОЭПЩвдОЋШЗЕїПиЦфЛКЪЭЙ§ГЬ[53] ЁЃетЮЊбаОПЯИАћЩњ РэКЭзщжЏЙЄГЬЬсЙЉСЫгаРћЕФЦНЬЈ ЁЃЕЋЪЧСэвЛЗНУц , ЯрБШЕААзжЪЖјбд , ЖрыФдкЦфЩњРэЙІФмЖрбљадКЭЩњЮяЙ§ ГЬЕФНЧЩЋЩЯЖМБШНЯгаЯо ЁЃвђДЫ , ШЋЕААзжЪЫЎФ§НКЕФбаОПвВе§ЗНаЫЮДАЌ ЁЃ
3.2 ШЋЕААзжЪЫЎФ§НК
ЛљвђжизщКЭЩњЮяКЯГЩЪжЖЮЕФГЩЪьЪЙЕУжизщЕААзжЪЕФжЦБИКЭаджЪЕїПиБфЕУГЃЙцЁЃдкЩњЮяЬхФк,ЕААзжЪЛљдЊжЎМфЕФНЛСЊЪЧаЮГЩЯИАћЭтЛљжЪКЭЦфЫќЕААзжЪВФСЯЕФживЊЙ§ГЬЁЃвђДЫ,ЭъШЋЛљгкЕААзжЪЕФЫЎФ§НКПЩвдзюДѓГЬЖШЕиФЃФтШЫЙЄЯИАћЭтЛљжЪКЭРћгУЬьШЛЕААзжЪЕФЩњЮяЛюадЁЃФПЧА,ЖрЪ§ШЋЕААзжЪЫЎФ§НКЖМЪЧЛљгкПЩПиШЗЖЈЕФЕААзжЪ-ЕААзжЪЯрЛЅзїгУЛђНЯВЛЙцећЕФЕААзжЪОлМЏЫљаЮГЩЕФЮяРэНЛСЊЁЃЧАепШчССАБЫсРСД(Leucinezipper)ЕФОлМЏзїгУ[54,55]ЁЂЖўМлРызгФЭЪмЕААз(cutA)ЕФШ§Ол[56,57] вдМАИЦ-ИЦЕїЕААз(calmodulin)ЕФНсКЯЕШ[58] ,КѓепШчЕЏадЕААзЕФШШОлМЏ[59] КЭИЛКЌИЌАБЫсЕФыФЖЮЕФОлМЏЕШ[60] ЁЃ
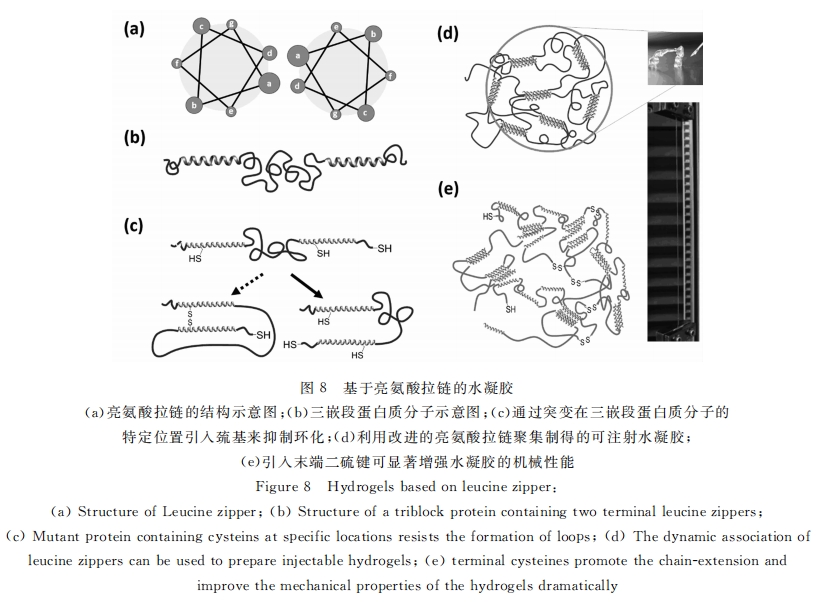
ССАБЫсРСД(Leucine zipper)ЪЧвЛжжФмаЮГЩОэЧњТна§НсЙЙЕФЕААзжЪ,гЩЖрИі71 Тна§ОлМЏаЮГЩ,ЦфЦп дЊжиИДЕЅдЊПЩвдБэЪОГЩabcdefg,ЦфжаСНзщТна§ЕФaгыdЮЊЪшЫЎЛљЭХ,ЭЈГЃЮЊССАБЫс,eгыgЮЊДјЕчВаЛљ (МћЭМ10a)ЁЃЫќУЧЭЈЙ§ССАБЫсЕФЪшЫЎзїгУаЮГЩОэЧњТна§ЁЃTirrellзщРћгУССАБЫсРСДЕФЪшЫЎЯрЛЅзїгУзї ЮЊНЛСЊЪжЖЮ,ЩшМЦСЫШ§ЧЖЖЮзщЕААзжЪЛљдЊAc10A[61] (МћЭМ10b),AДњБэССАБЫсРСД,c10ДњБэЧзЫЎШсадСДЁЃЫќПЩвдаЮГЩЫЎФ§НК,ВЂОпгаpHЯьгІадЁЃЕБpHЩ§ИпЪБ,ССАБЫсРСДAЩЯЙШАБЫсЕФРызгЛЏЪЙССАБЫсРСД НтОлЁЃКЫДХБэеїНјвЛВНЗЂЯж,ЬхЯЕжаЫЎШмадЕФШсадСДЖЮЪЧЖЏЬЌБфЛЏЕФ,ЖјССАБЫсРСДдђБэЯжЮЊБШНЯИеад ЕФНсЙЙ,етгыЩшЯыЕФНсЙЙЪЧЯрЮЧКЯЕФ[62] ЁЃ
KennedyЕШЗЂЯжССАБЫсРСДДцдкЖўОлЬх-ЫФОлЬхЕФЦНКт , етаЉОлМЏЬхЙВЭЌЙЙГЩСЫЫЎФ§НКЕФЮяРэНЛСЊ Еу , жаМфШсадыФСДЕФЫЎШмадПЩвдЗРжЙЖрОлЬхЕФЮоЙцОлМЏЮіГі[63] ЁЃШЛЖј , дкПЊЗХШмвКЛЗОГжа , ЕААзжЪЛсж№ НЅвдЛЗзДжаМфЬхЕФаЮЪНШмНтЕНШмвКжаШЅ ЁЃЮЊСЫЮШЖЈДЫРрЫЎФ§НК , ЫћУЧНЋССАБЫсРСД A жиИДЕЅдЊЩЯЕФ d ЮЛАБЛљЫсЭЛБфЮЊАыызАБЫс(МћЭМ 10c)ЁЃвђЮЊ AЧуЯђгкЗДЦНааОлМЏ , етжжЩшМЦЪЙжЎИќЧуЯђаЮГЩЗжзгМфЖўСђ МќЖјЗЧЗжзгФкЖўСђМќ , МѕЩйСЫГЩЛЗЕФПЩФмад , ЬсИпСЫЫЎФ§НКЕФЮШЖЈад[64] ЁЃ
НтОіССАБЫсРСДЫЎФ§НКВЛЮШЖЈадЕФСэвЛИіЗНЗЈЪЧВЩгУОпгаВЛЭЌОлМЏЬиадЕФССАБЫсРСДЁЃTirrellзщЩшМЦСЫСэвЛжжЛљгкДѓЪѓШэЙЧЙбОлЛљжЪЕААз(cOMP)ЕФPаЭССАБЫсРСДЁЃЩЯЪіЬхЯЕжаЕФССАБЫсРСДГЩЗжAЧуЯђгкаЮГЩЫФОлЬх,ЖјPЧуЯђгкаЮГЩЮхОлЬх,ВЂЧвPгыAФбвдаЮГЩвьОлЬхЁЃвђДЫ,ФЉЖЫГЩЗжВЛЭЌЕФШ§ЧЖЖЮНсЙЙPc10AОЭВЛШнвзаЮГЩЛЗзДЗжзгЖјШмНт,ЦфИЏЪДЫйТЪвЊЕЭгкAc10AЬхЯЕ2~3ИіЪ§СПМЖ[54,55, 65,66] ЁЃОпгаДЬМЄЯьгІадЕФЯрЫЦЬхЯЕдкekзщЕФЯЕЭГЙЄзїжаЕУЕНКмКУЕФЬхЯжЁЃЫћУЧЩшМЦСЫЫФжжШ§ЧЖЖЮЕААзжЪ[67] ,ЦфжаОэЧњТна§НсЙЙгђЕФЬиЖЈЮЛЕуБЛЬцЛЛГЩОпгае§ЕчКЩЕФРЕАБЫс,ЪЙЦфЮШЖЈадИїВЛЯрЭЌЁЃдВЖўЩЋЦзБэУї,ДЫРрЫЎФ§НКжаЕФЖўМЖНсЙЙдкМгШШЛђЛЏбЇБфадМСзїгУЯТЕФБфадЙ§ГЬЪЧПЩФцЕФЁЃетдкКъЙлЩЯБэЯжЮЊШмвККЭФ§НКжЎМфЕФзЊБфЁЃЛљгкССАБЫсРСДЕФЖЏЬЌаджЪ,OlsenЕШЛЙЗЂеЙСЫПЩзЂЩфЕФЫЎФ§НК[54] вдМАЖЯСбЩьГЄТЪИпДя3000%ЕФИпЕЏадЫЎФ§НК[10] (МћЭМ8d-e)ЁЃ
chenЕШвдcutAШ§ОлЬхзїЮЊШ§ЙйФмЖШНЛСЊЗжзг,ФкКЌыФ(Intein)ЕФМєНгзїгУзїЮЊХМСЊЪжЖЮ,жЦБИСЫвЛжжаТаЭЕФСНзщЗжШЋЕААзжЪЫЎФ§НК[68] ЁЃгЩгкФкКЌыФЕФМєНгЫйЖШПьЁЂаЇТЪИп[69] ,ВЂЧвЭъШЋКЭЯИАћЯрШн,вђДЫВЛрДЮЊвЛжжЙЙНЈПЩЛљвђБрТыЕФЫЎФ§НКЬхЯЕЕФКУЗНЗЈЁЃРрЫЦItoзщЕФcutA-Tip-PEGЬхЯЕ,chenЕШЙЙНЈСЫШ§ЙйФмЖШЕФcutA-Tip1ligКЭcutA-Tip1,ЦфжаTipligКЭTip1ФмЙЛЛЅЯрНсКЯЕМжТФ§НКаЮГЩЁЃжЕЕУвЛЬсЕФЪЧ,ИУЕААзжЪжаЛЙАќКЌзХвЛЖЮОпгаУЊЖЈзїгУЕФЖрыФађСаSH3lig ,дЪаэМгдиШЮвтОпгаSH3НсЙЙгђЕФШкКЯЕААз,ФЃЪНЛЏЕив§ШыЬиЖЈЙІФм[70] (МћЭМ6b)ЁЃДЫЭт,ЕЏадЕААзОпгаЖРЬиЕФЕЭСйНчШмНтЮТЖШ(LCST),дкЮТЖШЩ§ИпЕФЪБКђШмНтЖШНЕЕЭЁЃИљОнАБЛљЫсађСаЕФВЛЭЌ,ЫќУЧЕФЧзЫЎадКЭШШзЊБфЮТЖШОљгаВЛЭЌЁЃЫљвд,ЖдгквЛИіШ§ЧЖЖЮЕЏадЕААзРДЫЕ,ШчЙћСНЖЫОпгаНЯЕЭЕФШШзЊБфЮТЖШ,жаМфЖЮЪЧИпЖШЧзЫЎЕФ,ФЧУДдкШмвКжаЩ§ЮТЕФЪБКђ,дїЫЎЕФСНЖЫОЭЛсЗЂЩњОлМЏ,аЮГЩФ§НК[59] ЁЃdevriesЕШРћгУНЭФИЗЂНЭЛёЕУДѓСПИпЖШЧзЫЎЕФУїНКЕААзжЪ[71] ,НсКЯРрЫПЕААз,ЩшМЦЙЙНЈСЫвЛжжШ§ЧЖЖЮЕААзжЪ,ВЂНјвЛВНЭЈЙ§НЋРрЫПЕААзађСажаЕФЬиЖЈЮЛЕуЬцЛЛГЩФмЙЛРызгЛЏЕФзщАБЫс,ИГгшСЫФ§НКЛЏЙ§ГЬpHЯьгІад[72] ЁЃ
РћгУЬьШЛЕААзжЪЕФЗДгІадВаЛљвВПЩвдЪЕЯжНЛСЊЕФШЋЕААзжЪЭјТчЁЃЭЌЪБ,ЖдЕААзжЪЕФМђЕЅаоЪЮПЩвддЪ аэвдЖржжЖрбљЕФЗНЗЈРДЪЕЯжФ§НКЛЏЁЃЫфШЛетВЂВЛЭъШЋЪЧШЋЕААзжЪЛљЫЎФ§НК,ЕЋЫќДѓДѓЭиеЙСЫЕААзжЪЛљЫЎ Ф§НКЕФЗЖГыКЭЙІФм,ЬсЙЉСЫЦфИФадЕФаТЫМТЗЁЃР§Шч,здШЛНчжаЕФБДРрФмЙЛРЮРЮЙЬЖЈдкбвЪЏЩЯ,жївЊЙщЙІгк ЦфБДРр№ЄИНЕААз(MAP)жаЙиМќГЩЗж3,4-ЖўєЧЛљБНБћАБЫс(DOPA)ЕФДцдк,ЦфдРэБЛMessersmithЕШдкЛљ гкPEGЬхЯЕЕФКЯГЩИпЗжзгЫЎФ§НКжавбгаЬхЯж[73,74] ЁЃChaзщНјвЛВНдкЕААзжЪЬхЯЕРявВзіСЫЯъЯИЕФЙЄзїЁЃЫћУЧЪзЯШРћгУФЂЙНРвАБЫсУИЖджизщfp-1аЭMAPНјааКѓаоЪЮ,ЛёЕУКЌгаDOPAЕФMAPЕААзжЪ[75] ЁЃЫќ МШФмгыFe3+ аЮГЩЖрГнХфКЯЮя[76,77] ,гжФмдкбѕЛЏЬѕМў(ШчIO4- )ЯТаЮГЩЙВМлНЛСЊЁЃетЮЊШЋУцЬсЩ§ШЋЕААзжЪ ЫЎФ§НКЕФеГИНадКЭЛњаЕадФмЬсЙЉаТЕФвЛжжЫМТЗЁЃ
РћгУПЩПиЛЏбЇХМСЊЪЕЯжШЋЕААзжЪЫЎФ§НКЕФБЈЕРЯрЖдНЯЩйЁЃЬьШЛЕААзжЪ(БШШчУїНК)ЕФНЛСЊЭЈГЃПЩвдРћгУзЊАБУИРДЭъГЩ,ЕЋЦфНЛСЊЮЛЕуВЂВЛПЩПиЁЃзюНќ,Е§ЗДгІЕФГіЯжЮЊетИіФПБъЬсЙЉСЫПЩФмЁЃЕ§ЗДгІЪЧЗЂЩњдкЕ§ВЖЪж(ЕААзжЪЦЌЖЮ)КЭЕ§БъЧЉ(ЖрыФЦЌЖЮ)жЎМфЕФЗДгІ[78] ЁЃСНИіЦЌЖЮжЎМфЪзЯШЗЂЩњЗжзгЪЖБ№КЭНсКЯ,МЬЖјЗЂЩњздДпЛЏЗДгІ,дкЕ§ВЖЪжЕФРЕАБЫсКЭЕ§БъЧЉЕФЬьУХЖЌАБЫсжЎМф,ИпаЇаЮГЩвьыФМќ[79] ЁЃИУЗДгІОпгаИпЖШЕФЬивьад,КЭЯИАћЙ§ГЬМцШн,вбБЛГЩЙІЕигІгУгкЭиеЙЕААзжЪЕФЭиЦЫНсЙЙ(МћЭМ9)[79] ЁЃРћгУИУЗДгІжЦБИЕФЭјТчНсЙЙ(ЛђГЦЕ§ЭјТч)ЪЧвЛИіЭъШЋЩњЮяЯрШнЕФФ§НКЛЏЙ§ГЬ,ВЂПЩвдЭЈЙ§ЛљвђБрТыЕФЗНЪНв§ШыЩњЮяЛюадЛљдЊ(БШШчАзбЊВЁвжжЦвђзг),дЄМЦдкЛюЯИАћЕФЗтзАКЭааЮЊПижЦЗНУцНЋгаЙуЗКЕФгІгУЧАОАЁЃ
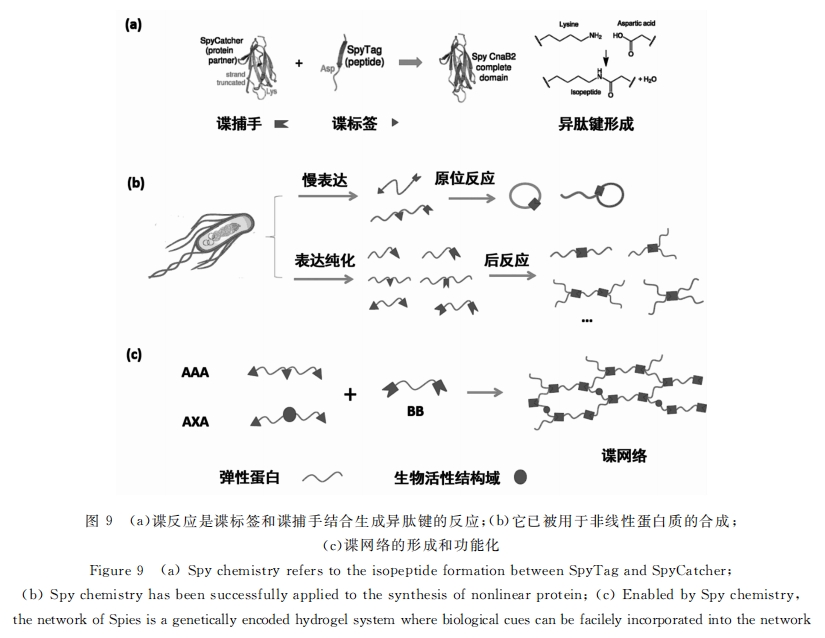
гЩДЫПЩМћ,ОпгаИпЖШЬивьадКЭЧПЧзКЭСІЕФЕААзжЪ-ЕААзжЪЯрЛЅзїгУдкШЋЕААзжЪЫЎФ§НКжаПЩвдЗЂЛгЦфЧПДѓЕФЙІФмЁЃЭЈЙ§ЛљвђЙЄГЬЕФЗНЗЈПЩвдБрМФ§НКЧАЬхЕФНсЙЙКЭЙІФм,ЖјЩњЮяКЯГЩдђБЃжЄЮвУЧФмЙЛМђБуЕиЛёЕУОпгаЯргІНсЙЙКЭЙІФмЕФФ§НКЧАЬхЕААзжЪЁЃСНжжЗНЪННсКЯ,ФмЙЛИќМгПьЫйМђБуЕиЛёЕУВЛЭЌЙІФмЕФФ§НКЧАЬхЕААзжЪ,ДгЖјЪЕЯжФЃПщЛЏЙЙНЈШЋЕААзжЪЫЎФ§НКЁЃР§Шч,дкЧАЬхЕААзжЪжаЧЖШывЛЖЮНсЙЙгђ,ЭЌЪБдкСэвЛЧАЬхЕААзжЪжав§ШыгыжЎНсКЯЕФЛЅВЙНсЙЙгђ,ОЭПЩвдНсКЯВЛЭЌЕФЙІФмЕААзЩшМЦжизщШкКЯЕААзжЪ,вджЦБИЙІФмадЫЎФ§НКЁЃНсЙЙгђжЎМфНсКЯЕФВЛЭЌЖЏСІбЇКЭВЛЭЌЮШЖЈадПЩвдБЛгУРДЮЂЕїЫЎФ§НКЕФКъЙладжЪЁЃ
4 змНсгыеЙЭћ
дкНќМИЪЎФъЕФПЦбЇЕРТЗЩЯ,ЕААзжЪЙЄГЬОРњСЫЗЩЫйЕФЗЂеЙ,ЖјВФСЯбаОПвВШеаТдТвьЁЃПЩвдЫЕ,ЖдКЯГЩИпЗжзгЫЎФ§НКЕФбаОПвбОЯрЕБГЩЪьЁЃФПЧА,ЩњЮяДѓЗжзг-КЯГЩИпЗжзгЕФИДКЯЫЎФ§НКЗНЯђШЁЕУСЫЯрЕБДѓЕФНјеЙ,ЭъШЋЛљгкЖрыФКЭЕААзжЪЕФЫЎФ§НКЬхЯЕвВе§ШеаТдТвьЁЃгаРэгЩЯраХ,РћгУЕААзжЪЕФгХЪЦЙЙНЈЫЎФ§НКВФСЯ,дкЮДРДШдОпгаМЋЦфЙуРЋЕФЧАОАвдМАгІгУЧБСІЁЃШЛЖјИУСьгђШдШЛДцдквЛЯЕСажиДѓЕФЬєеНЁЃЪзЯШ,ШчКЮевЕНвЛжжКЯЪЪЁЂМђБуПЩааЖјгжЩњЮяЯрШнЕФФ§НКЛЏЗНЗЈ,вдЪЪгІЙуЗКЕФЩњЮягУЭО? ЦфДЮ,ШчКЮРћгУЦфЬиЪтЕФЙІФмад,ЮЊЬхЯЕдіЬэЗсИЛЕФДЬМЄЯьгІадКЭжЧФмад,НјвЛВНЬсЩ§ЕААзжЪЫЎФ§НКЕФЩњЮяМлжЕ? ШчКЮеце§НЋВФСЯДгЖшадБфГЩЛюад,ДгБЛЖЏБфГЩжїЖЏ? здШЛНчОпгаИїЪНИїбљЁЂЙІФмЗсИЛЕФЕААзжЪ,вЊдкКЦШчбЬКЃЕФЕААзжЪПтжабАевКЯЪЪЕФЕААзжЪ,ВЂдЫгУЕНЫЎФ§НКЕФЙЙНЈКЭЙІФмЛЏжа,ашвЊНсЙЙЩњЮябЇКЭЩњЮяаХЯЂбЇЕФКЯзїЗЂеЙЁЃШЋЕААзжЪЫЎФ§НКЪЧвЛИіПЩвдвдЛљвђБрТыЕФЗНЪНШЅЕїПиВФСЯКъЙладФмЕФЖРЬиЬхЯЕ,ЮЊЮвУЧбаОПВФСЯНсЙЙЙІФмЙиЯЕЬсЙЉСЫЧПгаСІЕФЪжЖЮЁЃЯраХдкЮДРДМИФъФк,ШЋЕААзжЪЫЎФ§НКНЋЛсЭбгБЖјГі,ГЩЮЊЩњЮяВФСЯжаМЋОпЩњУќСІКЭЪЕМЪгІгУЧБСІЕФвЛИіСьгђЁЃ
Утд№ЩљУїЃКБОЮФЮЊаавЕНЛСїбЇЯАЃЌАцШЈЙщдзїепМАддгжОЫљгаЃЌШчгаЧжШЈЃЌПЩСЊЯЕЩОГ§ЁЃЮФеТБъзЂгазїепМАЮФеТГіДІЃЌШчашдФЖСдЮФМАВЮПМЮФЯзЃЌПЩдФЖСддгжОЁЃ
