
’Σ“ΣΘΚ…ηΦΤΚœ≥…ΝΥΕύΗωΨΏ”– 2 ΗωΜν–‘–ρΝ–ΒΡœΏ–‘ΚΆΜΖΉ¥ΕύκΡΦΑΨΏ”–ΒΞΗωΜν–‘–ρΝ–ΒΡΕΧΝ¥ΕύκΡ, ―–ΨΩΝΥΥϋΟ«ΒΡ…±ΨζΜν–‘, ΖΔœ÷Τδ…±ΨζΜν–‘Υ≥–ρΈΣ≥ΛΝ¥κΡ>ΜΖΉ¥κΡ>ΕΧΝ¥κΡ, ΧΊ±π «œΏ–‘ΒΡ Linear-KT ΚΆ Linear-KS Ε‘Εύ÷÷ΗοάΦ œ“θ–‘ΨζΚΆ―τ–‘ΨζΨυΨΏ”–ΫœΗΏΒΡ…±ΨζΜν–‘. ≤…”Ο MTT Ζ®ΩΦ≤λΝΥ Linear-KT ΚΆ Linear-KS Ε‘’ΐ≥ΘœΗΑϊΒΡΕΨ–‘, Τδ÷– Linear-KS ±μœ÷≥ωΫœΒΆΒΡœΗΑϊΕΨ–‘, ”≈”Ύ―τ–‘Ε‘’’Εύ’≥ΨζΥΊ B. άϊ”ΟΦΤΥψΡΘΡβΒΡΖΫΖ®ΦΤΥψΝΥΕύκΡ”κœΗΨζœΗΑϊΡΛ÷–ΝΉ÷§θΘΗ ”Ά(DMPG)ΒΡœύΜΞΉς”Ο. ΫαΙϊ±μΟς, ΕύκΡΚΆ DMPG ΒΡΫαΚœΡή“≤±μœ÷≥ω≥ΛΝ¥κΡ>ΜΖΉ¥κΡ>ΕΧΝ¥κΡΒΡΙφ¬…, ΧΊ±π « Linear-KT ΚΆ Linear-KS ΨΏ”–ΫœΗΏΒΡΫαΚœΡή. ≥ΛΝ¥κΡΚ§”– 2 ΗωΜν–‘–ρΝ–, Ω…ΧαΙ©ΕύΗωΚ…’ΐΒγΒΡΑ±ΜυΥα”κΚ…ΗΚΒγΒΡΝΉ÷§ΫαΚœ, ΫαΚœΡήΫœ¥σ, …±ΨζΜν–‘Ϋœ«Ω. Ά§ ±, »α–‘ΒΡΫαΙΙΦΑ Linear-KT ΚΆLinear-KS ÷–ΥΩΑ±ΥαΚΆΥ’Α±ΥαΒΡBΧΦ…œΒΡτ«ΜυΩ…”κΝΉ÷§…œΒΡτ Μυ–Έ≥…ΕύΗω«βΦϋ, Ϋχ“Μ≤Ϋ‘ω¥σΝΥΫαΚœΡή. ΦΤΥψΡΘΡβΒΡΖΫΖ®ΈΣΩΙΨζκΡΒΡ…±ΨζΜν–‘¥”άμ¬έ…œΧαΙ©ΝΥ“ΜΕ®ΒΡ“άΨί.
ΩΙΨζκΡ(Antimicrobial peptides, AMPs) «Υό÷ςΖά”υΧεœΒ÷–Χλ»ΜΟβ“ΏœΒΆ≥Υυ≤ζ…ζΒΡΨΏ”–“ΜΕ®…±Ψζ–ßΙϊΒΡΕΧκΡάύΈο÷ , ΙψΖΚΖ÷≤Φ”ΎΉ‘»ΜΫγ÷–, ΨΏ”–ΙψΤΉΩΙΨζ–‘. ΚήΕύΩΙΨζκΡ≤ΜΫωΕ‘ΗοάΦ œ“θ–‘ΨζΦΑΗοάΦ œ―τ–‘ΨζΨΏ”–…±ΨζΉς”Ο, Εχ«“Ε‘Ρ≥–©’φΨζΓΔ ‘≠…ζ…ζΈοΓΔ …θ÷Ν÷ΉΝωœΗΑϊΚΆ≤ΓΕΨ“≤ΨΏ”–“ΜΕ®ΒΡ“÷÷ΤΉς”Ο[1 ~ 4].―–ΨΩ±μΟς, ΩΙΨζκΡΒΡ÷ς“ΣΉς”ΟΜζάμ «’κΕ‘…ζΈοΡΛ, Ά®ΙΐΤΤΜΒ…ζΈοΡΛΒΡΆξ’ϊ–‘, ΙΤδ≤ζ…ζΩΉΕ¥Μρ’Ώ…ζΈοΡΛΆΏΫβΕχ ΙΡΎ≤ΩΈο÷ ΆβΝς, Μρ’Ώ¥©ΆΗ…ζΈοΡΛΫχ»κœΗΑϊΡΎ≤Ω, Η…‘Λ“Μ–©÷Ί“ΣΒΡœΗΑϊΙΐ≥Χ, ¥”ΕχΤπΒΫ…±ΨζΒΡΉς”Ο[5]. ”…”ΎΩΙΨζκΡΉς”Ο÷ς“Σ’κΕ‘”ΎœΗΑϊΡΛΫαΙΙ, ΤδΤ’±ιΨΏ”–±»ΫœΒΆΒΡΩΙ“©–‘, “ρ¥Υ“≤±Μ»œΈΣ «ΩΙ…ζΥΊΒΡΧφ¥ζΤΖΚΆ‘ω–ßΦΝ, ”Ο“‘ΫβΨω»’“φ―œ÷ΊΒΡœΗΨζΩΙ“©–‘Έ Χβ[6].
ΗυΨίΧλ»ΜΩΙΨζκΡΒΡΧΊ–‘Ϋχ––»Ϊ–¬ΒΡΩΙΨζκΡ…ηΦΤ «ΖΔœ÷–¬ΩΙΨζκΡΒΡ÷Ί“ΣΆΨΨΕ[7], Τδ÷–ΚήΕύΚ§”–ΕύΗωΜν–‘Τ§ΕΈΒΡΕύκΡΨΏ”–ΫœΗΏΒΡ…ζΈοΜν–‘. »γΚ§”– 2 ΗωΦν–‘»ήΫβΒΞΈΜΒΡ 24 ΗωΑ±ΜυΥα≤–ΜυΒΡΕύκΡ WLBU2 ±»ΨΏ”–ΒΞΗωΦν–‘»ήΫβΒΞΈΜΒΡΕύκΡ LBU ΦΑ…ΪΑ±Υα»Γ¥ζΒΡ WLBU ±μœ÷≥ωΗϋΗΏΒΡ…±ΨζΜν–‘ΦΑ―Γ‘ώ–‘[8]. FrecerΒ»[9]¥”ΆΖ…ηΦΤ≥ω–¬–ΆΒΡΨΏ”–ΥΪΜν–‘–ρΝ– HBHPHBH Μρ HBHBHBH(B: ¥χ’ΐΒγΒΡΑ±ΜυΥα; H: ηΥ°–‘Α±ΜυΥα; P: ΦΪ–‘Α±ΜυΥα)ΒΡΜΖΉ¥ΩΙΨζκΡ. Τδ÷– 1 ΗωΜΖΉ¥ΩΙΨζκΡ(V4, CVKVQVKVGSGVKVQVKVC)ΨΏ”–ΫœΗΏΒΡΩΙΨζ–‘, ΤδΥΪΜν–‘–ρΝ– ΙΝΫ«Ή–‘Φ”«Ω, ”–÷ζ”ΎΩΙΨζκΡ…ζΈοΜν–‘ΒΡ±μœ÷. ΒΪΗΟκΡ”…”ΎΚ§”–¥σΝΩ ηΥ°–‘γ”Α±Υα, Υ°»ή–‘≤ν, œό÷ΤΝΥΤδ”Π”Ο[10,11]. ≥ΐ WLBU2 ΚΆ V4 Άβ, ≤Μ…ΌΩΙΨζκΡ“≤¥φ‘ΎάύΥΤΒΡœ÷œσ. ΗυΨίΩΙΨζκΡ Tritrpticin …ηΦΤΒΡΕ‘≥ΤΡΘΡβκΡΨΏ”–±» Tritrpticin ΗΏ 2 ~ 8 ±ΕΒΡΩΙΨζκΡΜν–‘[12]. Sushi 3 ΩΙΨζκΡΒΡΕΰΨέΧε±»ΒΞΧεΨΏ”–ΗϋΗΏΒΡΤΤΜΒ÷§ΕύΧ«ΫΚΆ≈ΒΡΡήΝΠ[13]. Pexiganan(MSI-78) ΚΆ PG-1 ‘Ύ»ή“Κ÷– «ΒΞΧε, Β±Τδ”κ…ζΈοΡΛœύΫαΚœ ±, –Έ≥…ΝΫ«ΉΒΡΕΰΨέΧε, ±μΟςΕΰΨέΧε «Μν–‘ΙΙœσ[14]. ΕΰΨέΧε“≤Ω…Ω¥≥… «Ζ÷Ή”÷–Κ§”– 2ΗωΜν–‘–ρΝ–. ¥ΥΆβ, Ρ≥–©Χλ»ΜΩΙΨζκΡ–ρΝ–÷–“≤ΑϋΚ§”– 2 ΗωœύΆ§ΒΡΕύκΡΤ§ΕΈ, »γ Gramicidin S( CycloVOLdFPVOLdFP)”… 2 ΗωœύΆ§ΒΡΤ§ΕΈΜΖΜ·Εχ≥…[15]. “ρ¥Υ, ΨΏ”–ΥΪΜν–‘–ρΝ–ΒΡΕύκΡΩ…ΡήΨΏ”–«±‘ΎΒΡ…ζΈοΜν–‘. ±ΨΈΡ…ηΦΤΝΥΚ§”–ΥΪΜν–‘–ρΝ–ΒΡΩΙΨζκΡ≤ΔΩΦ≤λΝΥΤδΜν–‘, ―–ΨΩΝΥΥΪΜν–‘–ρΝ– «Ζώ”–÷ζ”ΎΧαΗΏΩΙΨζκΡΒΡΜν–‘. Φχ”ΎΕύκΡΒΡΜ·―ßΚœ≥…≥…±Ψ, ΨΏ±ΗΩΙΨζΜν–‘ΒΡΕΧκΡΨΏ”–¥σ≈ζΝΩ…ζ≤ζΒΡ«±ΝΠ, “ρ¥Υ―Γ»ΓΨΏ”–ΫœΕΧ–ρΝ–ΒΡΕύκΡΉςΈΣΜν–‘–ρΝ–, …ηΦΤΝΥΑϋΚ§ 2 ΗωœύΆ§Μν–‘–ρΝ–ΒΡΕύκΡ, ≤β ‘ΝΥΤδΩΙΨζΜν–‘ΦΑΕΨ–‘, ≤Δ‘Υ”ΟΦΤΥψΡΘΡβΒΡΖΫΖ®ΦΤΥψΝΥΥυ…ηΦΤΒΡΕύκΡ”κœΗΨζΒΡœΗΑϊΡΛ÷–“Μ÷÷÷Ί“ΣΒΡΝΉ÷§ΓΣΓΣΓΣΝΉ÷§θΘΗ ”Ά(DMPG)ΒΡΫαΚœΡή, ≤ΔΕ‘Εΰ’ΏΫχ––ΝΥ±»Ϋœ. ”…”ΎΕΰΝρΦϋΒΡΜΖΜ·”–÷ζ”ΎΈ»Ε®ΕύκΡΒΡΫαΙΙ, “ρ¥ΥΜΙ…ηΦΤΝΥΆ§ ±ΨΏ”–ΒΞΗωΕΰΝρΦϋΚΆΜΖΉ¥ΫαΙΙΒΡΕύκΡ, ―–ΨΩΝΥΤδΩΙΨζΜν–‘ΚΆΜΖΉ¥ΫαΙΙΒΡΙΊœΒ, ΈΣΨΏ”–ΜΖΉ¥ΫαΙΙΒΡΩΙΨζκΡΒΡΫαΙΙΚΆΜν–‘ΙΊœΒ―–ΨΩ“‘ΦΑΩΙΨζκΡΒΡ…ηΦΤΧαΙ©“άΨί.
1 Β―ι≤ΩΖ÷
1. 1 ‘ΦΝ”κ“«Τς
ΗοάΦ œ“θ–‘ΨζΑϋά®¥σ≥ΠΗΥΨζ(Escherichia coli)ΓΔ Ά≠¬ΧΦΌΒΞφΏΨζ(Pseudomonas aeruginosa)ΚΆ±Ϊ¬ϋ≤ΜΕ·ΗΥΨζ(Acinetobacter baumannii); ΗοάΦ œ―τ–‘ΨζΑϋά®ΫπΜΤ…ΪΤœΧ―«ρΨζ( Staphyloccocus aureus)ΚΆΧΌΜΤΈΔ«ρΨζ(Micrococcus luteus). Τδ÷–, ¥σ≥ΠΗΥΨζΓΔ ΫπΜΤ…ΪΤœΧ―«ρΨζΦΑΧΌΜΤΈΔ«ρΨζ”…Κ”Ρœ Γ“©ΤΖΦλ―ιΥυΧαΙ©, Τδ”ύΨζ÷ξ”…ΝΌ¥≤Ζ÷άκΒΟΒΫ. »Υ÷ßΤχΙή…œΤΛœΗΑϊ÷ξ(BEAS-2B œΗΑϊ)”…÷Θ÷ί¥σ―ßΈβΈάΕΪΫΧ ΎάΓ‘υ; Εύ’≥ΨζΥΊB(Polymyxin B, PB)ΚΆΥΡΦΉΜυ≈ΦΒΣ(MTT)ΙΚΉ‘ Sigma ΙΪΥΨ; ΤδΥϋΜ·―ß ‘ΦΝΨυΈΣΙζ≤ζΖ÷Έω¥Ω; Β―ι”ΟΥ°ΈΣΕΰ¥Έ’τΝσΥ°.
Spectra MR ΟΗ±ξ“«(ΟάΙζ DYNEX ΙΪΥΨ); IS10-OMNIL8 –ΆΚλΆβΈϋ ’ΙβΤΉ“«(ΟάΙζ Thermo Fisher ΙΪΥΨ); J-815 –Ά‘≤Εΰ…ΪΙβΤΉ“«(»’±Ψ JASCO ΙΪΥΨ)
1. 2 Β―ιΙΐ≥Χ
1. 2. 1 …±Ψζ Β―ι Χτ»Γ ΝΩΨζ÷÷‘ΎΙΧΧε Luria-Bertani(LB)≈ύ―χΜυ÷–Μ°œΏ, ÷Ο”Ύ 37 Ε»≈ύ―χœδ÷–Ιΐ“Ι≈ύ―χ. Χτ»Γ ΝΩ¥Π”ΎΕ‘ ΐ…ζ≥ΛΤΎΒΡœΗΨζ”Ύ…ζάμ―ΈΥ°÷–, ≈δ÷Τ 0.5 ~ 1 ¬σ œΉ«Ε»ΒΡΨζ–ϋ“Κ, ¥Υ ±œΗΨζΨζ¬δ ΐ‘ΦΈΣ 1X10^8cfu / mL, Ι”Ο ±œΓ Ά÷Ν 1X10^6cfu / mL. ≈δ÷Τ≈®Ε»ΈΣ 2 mg / mL ΒΡΕύκΡ»ή“Κ. ≤…”ΟΈΔΝΩ»βΧάΕΰ±ΕœΓ ΆΖ®≤βΕ®Κœ≥…ΕύκΡΕ‘ΗςœΗΨζΒΡΉν–Γ“÷Ψζ≈®Ε»(Minimum inhibitory concentration, MIC)[16], Ά§ ±Ϋχ––»ήΦΝΓΔ ÷ΜΦ”Ψζ“Κ≤ΜΦ”ΕύκΡΒΡ“θ–‘Ε‘’’ΚΆΦ”»κΕύ’≥ΨζΥΊ B ΒΡ―τ–‘Ε‘’’, ΟΩΗωΕύκΡΤΫ––Ϋχ–– 3 ¥Έ. ”Ύ 37Ε»ΚψΈ¬œδ÷–≈ύ―χ 12 h, Ιέ≤λΩΉΡΎ»ή“ΚΒΡΜλΉ««ιΩω, 96 ΩΉΑεΆ§≈≈÷–»β―έΥυΦϊ≥Έ«εΩΉΥυΕ‘”ΠΒΡΉν–Γ≈®Ε»Φ¥ΈΣΗΟΕύκΡΒΡ MIC ÷Β. ≤…”ΟΑκ ΐ”––ßΦΝΝΩ(ED50 )ΤάΦέΕύκΡΒΡ…±Ψζ–߬ . ≤…”ΟΟΗ±ξ“«≤βΕ®…œ ω 96 ΩΉΑε600 nm ¥ΠΒΡΙβΟήΕ»÷Β(OD600 ). Α¥œ¬ ΫΦΤΥψ…±ΨζΑΌΖ÷ ΐ, ΉςΆΦΒΟ≥ωΕύκΡ…±…ΥœΗΨζΒΡΑκ ΐ”––ßΦΝΝΩ.

1. 2. 2 œΗΑϊΕΨ–‘ Β―ι ≤…”Ο MTT Ζ®≤βΕ®œΗΑϊΕΨ–‘[17]. ”ΟΚ§ 10% (ΧεΜΐΖ÷ ΐ)ΧΞ≈Θ―Σ«εΒΡ RPMI1640(Κ§ 100 U/ mL «ύΟΙΥΊΚΆ 100 U/ mL Ν¥ΟΙΥΊ)≈ύ―χΜυ≥ΘΙφ≈ύ―χ BEAS-2B œΗΑϊ. ≤…”ΟΕ‘ ΐ…ζ≥ΛΤΎΒΡ BEAS-2B œΗΑϊ, ”ΟΚ§ 10% ΧΞ≈Θ―Σ«εΒΡ RPMI 1640 ≈ύ―χ“ΚΒςΫΎœΗΑϊ≈®Ε»ΈΣ 5X10^4Cell / mL, Ϋ”÷÷ΒΫ 96 ΩΉΑε…œ,ΟΩΩΉ 200 uL. ≈ύ―χ 12 h Κσ, Φ”»κ≤ΜΆ§≈®Ε»ΒΡΕύκΡ(÷’≈®Ε»Ζ÷±πΈΣ 2.5, 5, 10, 20, 40, 80, 160 ΚΆ 320ug / mL), ΟΩ÷÷ΕύκΡ≈®Ε»…η 4 ΗωΗ¥ΩΉ, ≤Δ…η≤ΜΦ”œΗΑϊ“ΚΒΡΒςΝψΩΉΚΆ÷ΜΦ”œΗΑϊ“ΚΓΔ ≤ΜΦ”κΡ“ΚΒΡΩ’ΑΉΕ‘’’ΩΉ. ≈ύ―χ 4 h Κσ, ΟΩΩΉΦ”»κ 180 uL ≤ΜΚ§―Σ«εΒΡ RPMI 1640 ≈ύ―χ“ΚΚΆ 20 uL 5 g / L ΒΡ MTT ‘ΦΝ, ΦΧ–χΖθ”ΐ 4 h, Τζ»Ξ…œ«ε“Κ, Φ”»κ 180 uL DMSO, «α«α’ώΒ¥ 10 min, ”Ύ 490 nm ≤®≥Λœ¬≤βΕ®ΙβΟήΕ»÷Β.
1. 2. 3 ΚλΆβΙβΤΉ≤βΕ® »ΓΕύκΡ ‘―υ‘Φ 1 mg ”κ KBr Μλ‘», ―ΙΤ§, ≤βΕ®ΕύκΡΒΡΚλΆβΙβΤΉ.
1. 2. 4 ‘≤Εΰ…ΪΙβΤΉ≤βΕ® ‘Ύ “Έ¬œ¬, ≤…”Ο 0.1 mm ·”Δ±»…ΪΟσ≤βΕ®ΕύκΡ‘ΎΥ°»ή“Κ÷–ΒΡ‘ΕΉœΆβ‘≤Εΰ…ΪΙβΤΉ. …®ΟηΖΕΈß 175 ~ 260 nm, ΟΩΧθΕύκΡ…®Οη 3 ¥Έ, …®ΟηΥΌΕ» 1000 nm / min, ΕύκΡ≈®Ε»ΈΣ 1 mg / mL.
1. 2. 5 ΦΤΥψΖΫΖ® ΝΉ÷§θΘΗ ”Ά(DMPG) «œΗΨζΒΡœΗΑϊΡΛ÷–ΒΡ“Μ÷÷÷Ί“ΣΉι≥…≥…Ζ÷, ¥χ”–ΗΚΒγΚ…, ‘ΎΩΙΨζκΡΚΆœΗΨζœΗΑϊΡΛΫαΚœΙΐ≥Χ÷–ΨΏ”–÷Ί“ΣΉς”Ο. ≤…”Ο DMPG ΡΘΡβœΗΨζœΗΑϊΡΛ, Ε‘ΕύκΡΚΆ DMPG ΒΡΫαΚœΡήΫχ––ΝΥΦΤΥψ. ΕύκΡ”κ DMPG Ζ÷Ή”ΒΡ≥θ ΦΙΙ–Ά”… Gaussian View ΚΆ Amber »μΦΰ÷–ΒΡ leap ΡΘΩιΙ≤Ά§ΙΙΫ®, ΦΤΥψ Ι”Ο Gaussian 09 »μΦΰ[18], ≤…”Ο PM3 ΑκΨ≠―ιΥψΖ®Ε‘ DMPG ΦΑΗςΗωΕύκΡΖ÷Ή”Ϋχ––ΙΙ–Ά”≈Μ·. »ΜΚσΕ‘ΕύκΡ”κ DMPG Ζ÷Ή”ΒΡΗ¥ΚœΈοΫχ––”≈Μ·ά¥ΡΘΡβΕύκΡ”κ DMPG Ζ÷Ή”ΒΡœύΜΞΉς”Ο, ”…”ΎΕύκΡ”κ DMPG Ζ÷Ή”ΒΡΗ¥ΚœΈοΖ÷Ή”ΝΩΫœ¥σ, ΡΘΡβ ±»‘»Μ≤…”Ο PM3 ΑκΨ≠―ιΥψΖ®Ϋχ––”≈Μ·. ΕύκΡ”κ DMPG Ζ÷Ή”÷ς“ΣΆ®ΙΐΨ≤ΒγΉς”ΟΫαΚœ‘Ύ“ΜΤπ, Ά§ ±ΜΙΜα–Έ≥…«βΦϋ, ΦΤΥψ ±–η“ΣΆ§ ±ΩΦ¬« 2 ÷÷Ής”Ο. Ά®ΙΐΫαΚœΡήά¥ΚβΝΩΕύκΡ”κ DMPG Ζ÷Ή”ΦδΒΡœύΜΞΉς”Ο≥ΧΕ», ΫαΚœΡήΦΤΥψΖΫΖ®»γœ¬:
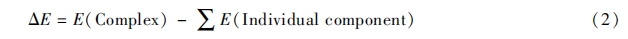
Ϋ÷–, E ΈΣ”≈Μ·ΚσΗςΉιΖ÷ΒΡΒΞΒψΡή. ΦΤΥψ ±ΨΓΝΩΩΦ¬«≤ΜΆ§ΒΡΫαΚœΡΘ Ϋ, “‘ ΙΕύκΡ”κ DMPG ¥οΒΫΉνΚΟΒΡΫαΚœ.
2 ΫαΙϊ”κΧ÷¬έ
2. 1 ΕύκΡΒΡ…ηΦΤ
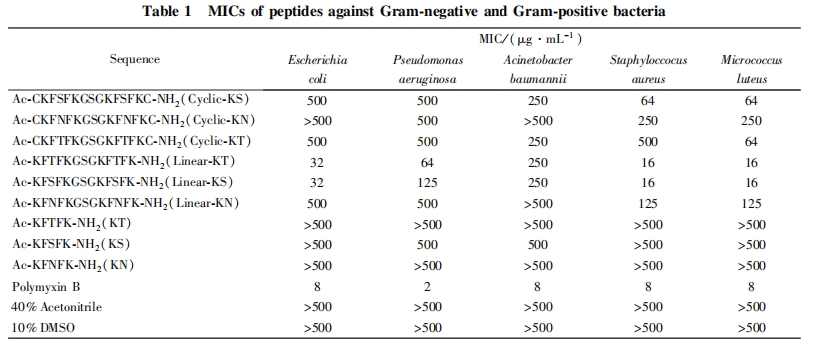
ΡΎΕΨΥΊ”÷≥Τ÷§ΕύΧ«, «ΗοάΦ œ“θ–‘ΨζΆβ≤ψœΗΑϊΡΛΒΡ÷Ί“Σ≥…Ζ÷÷°“Μ, ÷ς“ΣΈΜ”ΎΡΛΒΡΆβ≤ύ, «”κΗς÷÷Άβ‘¥–‘Έο÷ Ήνœ»Ϋ”¥ΞΒΡ≤ΩΖ÷. Lipid A «ΡΎΕΨΥΊ÷–ΨΏ”–…ζΈοΜν–‘ΒΡ±Θ Ί≤ΩΖ÷. ―–ΨΩ±μΟς[9], ”κΡΎΕΨΥΊΫαΚœΒΡΥό÷ςΖά”υΒΑΑΉ”…ΗΜΚ§¥χ’ΐΒγΒΡΑ±ΜυΥαΉι≥…ΝΫ«Ή–‘ΫαΙΙ, ΨΏ”–…±ΨζΙΠΡή, “ρ¥ΥΜυ”ΎΥό÷ςΖά”υΒΑΑΉΒΡΫαΚœΜυ‘Σ…ηΦΤΚœ≥…ΒΡΕύκΡΩ…Ρή±ΘΝτΤδ…±ΨζΜν–‘, ΨΏ”–«±‘ΎΒΡ”Π”ΟΦέ÷Β. Frecer Β»[19]Μυ”Ύ¥ΥΦΤΥψΝΥΕύΗωΕύκΡΤ§ΕœΚΆ Lipid A ΒΡΫαΚœΡή, ΖΔœ÷ KFNFK, KFTFK ΦΑ KFSFK –ρΝ–ΕύκΡ”κ Lipid A ΫαΚœ ΆΖ≈ΝΥΫœΗΏΒΡΫαΚœΡή, ΙΙœσΫœΈ»Ε®, ¥”άμ¬έ…œ‘Λ≤βΝΥ’β 3 ΗωΕύκΡΤ§ΕœΩ…ΡήΨΏ”–ΩΙΨζΜν–‘. KFNFK, KFTFK ΚΆ KFSFKΒΡΦΪ–‘ΚΆΖ«ΦΪ–‘Α±ΜυΥαΫΜ≤φ≈≈Ν–, Ζ÷±π–Έ≥… ηΥ°ΟφΚΆ«ΉΥ°Οφ, Α±ΜυΥα–ρΝ–ΖϊΚœ BHPHB Ζ÷≤ΦΙφ¬…. ΨΏ”–”κ Lipid A ΫœΗΏΒΡΫαΚœΡή±μΟς’β 3 ΗωΕύκΡΩ…ΡήΗϋ»ί“ΉΚΆœΗΨζΒΡœΗΑϊΡΛΫαΚœ. “ρ¥Υ±ΨΈΡΫΪ’β 3 ΗωΕύκΡ ”ΈΣ“ΜΗωΜν–‘–ρΝ–, ≤Έ’’ V4 ΩΙΨζκΡΒΡ…ηΦΤ, ≤…”Ο GSG ΫΪ 2 ΗωΜν–‘–ρΝ–Ν§Ϋ”–Έ≥…Κ§”–ΥΪΜν–‘–ρΝ–ΒΡœΏ–‘ΕύκΡ Linear-KT, Linear-KS ΚΆ Linear-KN. ΈΣΫχ“Μ≤ΫΙΧΕ®ΕύκΡΒΡΙΙœσ, Ι ηΥ°ΟφΗϋ ηΥ°, «ΉΥ°ΟφΗϋ«ΉΥ°,≤…”ΟΕΰΝρΦϋΜΖΜ·œΏ–‘ΕύκΡ, –Έ≥…ΜΖΉ¥ΒΡΚ§”–ΥΪΜν–‘–ρΝ–ΒΡΕύκΡ Cyclic-KS, Cyclic-KN ΚΆ Cyclic-KT(ΨΏΧε–ρΝ–Φϊ±μ 1).
2. 2 ΕύκΡΒΡΉν–Γ“÷Ψζ≈®Ε»ΒΡ»ΖΕ®
≤…”ΟΡΩ ”Ζ®, Ιέ≤λ»β―έΥυΦϊΒΡ≥Έ«εΩΉΥυΕ‘”ΠΒΡΉν–Γ≈®Ε», ΩΦ≤λΝΥΕύκΡΕ‘Εύ÷÷ΗοάΦ œ“θ–‘ΨζΚΆ―τ–‘ΨζΒΡΩΙΨζΜν–‘, ΫαΙϊΝ–”Ύ±μ 1. ”…±μ 1 ΐΨίΩ…“‘Ω¥≥ω, ΨΏ”–ΒΞΗωΜν–‘–ρΝ–ΒΡΕύκΡ KT, KS ΦΑ KN Ε‘ 5 ÷÷œΗΨζΒΡ…±ΨζΜν–‘Ψυ≤Μάμœκ, Ήν–Γ“÷Ψζ≈®Ε»Ψυ≤ΜΒΆ”Ύ 500 mg / mL, ΥΒΟςΨΏ”–ΒΞΗωΫαΚœ–ρΝ–ΒΡΕύκΡΜν–‘≤ΜΗΏ. ΨΏ”–ΥΪΜν–‘–ρΝ–ΒΡœΏ–‘ΕύκΡΕ‘œΗΨζΒΡ…±ΨζΜν–‘ΫœΨΏ”–ΒΞΗωΜν–‘–ρΝ–ΒΡΕύκΡ”–ΥυΧαΗΏ, ΧΊ±π « Line-ar-KT ΚΆ Linear-KS, Ε‘ΫπΜΤ…ΪΤœΧ―«ρΨζΦΑΧΌΜΤΈΔ«ρΨζΒΡΉν–Γ“÷Ψζ≈®Ε»ΒΆ÷Ν 16 mg / mL. ΕχΜΖΉ¥ΒΡΕύκΡΕ‘œΗΨζΒΡ…±ΨζΜν–‘Τ’±ιΫœΒΆ, Ϋι”ΎΨΏ”–ΒΞΗωΜν–‘–ρΝ–ΚΆΥΪΜν–‘–ρΝ–ΒΡœΏ–‘ΕύκΡ÷°Φδ. ‘ΎΕ‘ 5 ÷÷œΗΨζΒΡ…±ΨζΜν–‘Φλ―ι÷–, Κ§”–ΥΪΜν–‘–ρΝ–ΒΡΕύκΡ Linear-KT ΚΆ Linear-KS Ψυ±» Linear-KN ±μœ÷≥ωΫœΗΏΒΡ…±ΨζΜν–‘,Εχ Linear-KT ΚΆ Linear-KS ΒΡΜν–‘‘ρœύΫϋ. ΨΏ”–“ΜΕ®Μν–‘ΒΡΕύκΡΕ‘ΗοάΦ œ―τ–‘ΨζΒΡ…±ΨζΜν–‘Ψυ¬‘«Ω”ΎΗοάΦ œ“θ–‘Ψζ
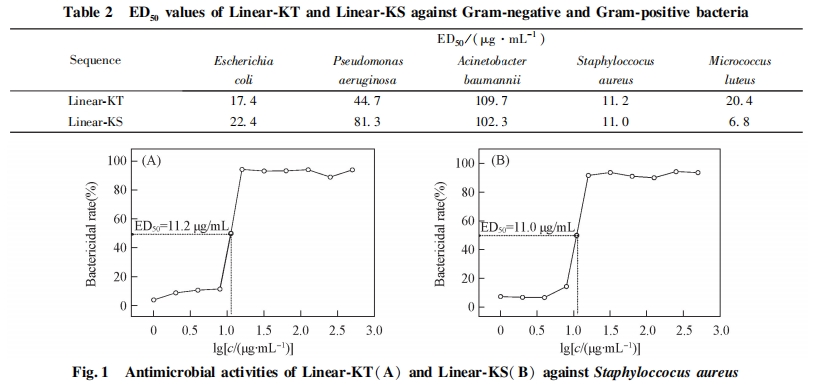
2. 3 ΕύκΡΕ‘œΗΨζΒΡΑκ ΐ”––ßΦΝΝΩΒΡ»ΖΕ®
”…ΙΪ Ϋ(1)ΦΤΥψΒΟ≥ωΕύκΡ‘ΎΗς≈®Ε»œ¬ΒΡ…±ΨζΑΌΖ÷ ΐ, Μφ÷ΤΤδ…±Ψζ¬ «ζœΏ≤ΔΜώΒΟΑκ ΐ”––ßΦΝΝΩED50 , ΫαΙϊΝ–”Ύ±μ 2. ΆΦ 1 ΈΣ Linear-KT ΚΆ Linear-KS Ε‘ΫπΜΤ…ΪΤœΧ―«ρΨζΒΡ…±Ψζ¬ «ζœΏ.
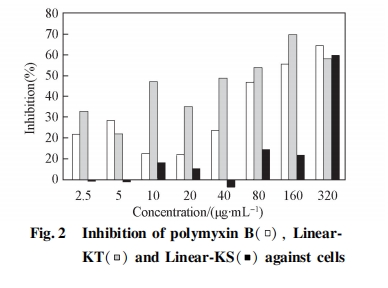
2. 4 œΗΑϊΕΨ–‘ Β―ι
―Γ»ΓΥυ…ηΦΤΕύκΡ÷–…±Ψζ–ßΙϊΫœΚΟΒΡ Linear-KT ΚΆ Linear-KS Ϋχ––œΗΑϊΕΨ–‘ Β―ι, ΫαΙϊΦϊΆΦ 2. ―τ–‘Ε‘’’ΕύκΡάύΩΙΨζΥΊΕύ’≥ΨζΥΊ B ΥδΨΏ”–Ϋœ«ΩΒΡ…±ΨζΜν–‘(±μ 1), ΒΪ“≤“ΐΤπ≤ΩΖ÷’ΐ≥ΘœΗΑϊΒΡΥάΆω, ΧΊ±π «Τδ≈®Ε»ΗΏ”Ύ 80 mg / mL ±, Εύ’≥ΨζΥΊ B Ω…ΒΦ÷¬ΫœΗΏΒΡœΗΑϊΥάΆω¬ . Linear-KT ΒΡœΗΑϊΕΨ–‘Ϋœ¥σ, ‘Ύ¥σ≤ΩΖ÷ΒΡ―–ΨΩ≈®Ε»œ¬ΨυΕ‘œΗΑϊ”–ΕΨ–‘. œύ±»÷°œ¬,Linear-KS Ε‘œΗΑϊΒΡΕΨ–‘Ϋœ–Γ, ‘Ύ≈®Ε»≤ΜΗΏ”Ύ 160ug / mL ±, ΤδΕ‘œΗΑϊΒΡ“÷÷Τ¬ Ψυ≤Μ≥§Ιΐ 15% , ΒΆ≈®Ε» ±ΕΨ–‘ΗϋΒΆ. Linear-KS ΨΏ”–ΫœΗΏΒΡ…±ΨζΜν–‘«“œΗΑϊΕΨ–‘ΫœΒΆ, “ρ¥Υ Linear-KS ΨΏ”–Ϋχ“Μ≤Ϋ―–ΨΩΒΡΦέ÷ΒΚΆ«±‘ΎΒΡ”Π”ΟΦέ÷Β.
2. 5 ΚλΆβΙβΤΉΫβΈω
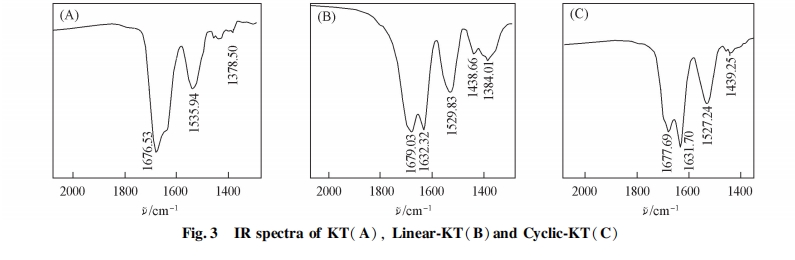
ΚλΆβΙβΤΉ «ΫβΈωΕύκΡ“‘ΦΑΒΑΑΉ÷ ΕΰΦΕΫαΙΙΒΡ”–ΝΠ ÷ΕΈ. ΕύκΡΚΆΒΑΑΉ÷ ‘ΎΚλΆβΙβΤΉ«χ”–»τΗ…ΧΊ’ςΈϋ ’¥χ, ÷ς“Σ”–θΘΑΖI¥χ(1600 ~ 1700 cm-1)ΚΆθΘΑΖII¥χ(1600 ~ 1500 cm-1). θΘΑΖI¥χ÷ς“Σ”…ΕύκΡΙ«ΦήκΡΝ¥ C O ΒΡ…λΥθ’ώΕ·“ΐΤπ, Ε‘―–ΨΩΕΰΦΕΫαΙΙΉν”–Φέ÷Β[20]. ΆΦ3 ΈΣ KT, Linear-KT ΚΆ Cyclic-KT ΒΡΚλΆβΙβΤΉΆΦ. Ω…Φϊ, KT ‘Ύ 1600 ~ 1700 cm-1¥Π”– 1 ΗωΟςœ‘ΒΡ«ΩΖε, Ζε–ΈΫœΩμ; Linear-KT ‘Ύ 1679 ΦΑ 1632 cm-1¥Π”– 2 ΗωΚή«ΩΒΡΈϋ ’Ζε; Εχ Cyclic-KT ‘Ύ 1677 ΦΑ 1631 cm-1¥Π“≤”– 2 ΗωΚή«ΩΒΡΈϋ ’Ζε. “ρ¥Υ, Linear-KTΚΆ Cyclic-KT ΨυΩ…Ρή–Έ≥…’έΒΰΫαΙΙ. ≤…”ΟΚλΆβΙβΤΉΥδ»ΜΩ…¥”“ΜΕ®≥ΧΕ»…œΧαΙ©ΕύκΡΒΡΕΰΦΕΫαΙΙ–≈œΔ, ΒΪ”…”Ύ 1600 ~ 1700 cm-1≤®ΕΈ ήΗ…»≈“ρΥΊΫœΕύ, «“ΚλΆβΙβΤΉ≤β ‘÷–ΕύκΡ¥Π”ΎΙΧΧεΉ¥Χ§, ≤ΜΡήΖ¥”≥≥ωΕύκΡ‘ΎΥ°»ή“Κ÷–ΒΡΉ¥Χ§, “ρ¥ΥΫχ“Μ≤Ϋ≤…”Ο‘≤Εΰ…ΪΙβΤΉΩΦ≤λΝΥΕύκΡ‘ΎΥ°÷–ΒΡΕΰΦΕΫαΙΙ.
2. 6 ‘≤Εΰ…ΪΙβΤΉ≤βΕ®
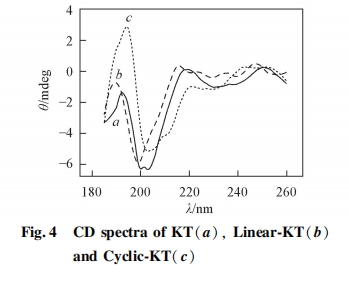
ΆΦ 4 ΈΣΕΧΝ¥ΕύκΡ KTΓΔ œΏ–‘ΕύκΡ Linear-KT ΦΑΜΖΉ¥ΕύκΡ Cyclic-KT ΒΡ‘≤Εΰ…ΪΙβΤΉ. ΕΧΝ¥ΕύκΡ KT ‘Ύ200 nm ¥Π”– 1 ΗωΗΚΖε, ΈΣΈόΙφΨμ«ζΫαΙΙΒΡΧΊ’ςΖε[21,22]. KT ÷ΜΑϋΚ§ 5 ΗωΑ±ΜυΥα≤–Μυ, κΡΝ¥≥ΛΕ»ΫœΕΧ, ≤ΜΉψ“‘–Έ≥…ΕΰΦΕΫαΙΙ, “ρ¥Υ“‘ΈόΙφΫαΙΙ¥φ‘Ύ”Ύ»ή“Κ÷–. ΕύκΡ Linear-KT ‘Ύ 200 nm ¥Π“≤”– 1 ΗωΗΚΖε, ±μΟς Linear-KT‘ΎΥ°»ή“Κ÷–“≤≥ ΈόΙφΫαΙΙ. ΜΖΉ¥ΕύκΡ Cyclic-KT ‘Ύ 202ΦΑ 213 nm ¥Π”– 2 ΗωΗΚΖε, «“‘Ύ 195 nm ¥Π”– 1 Ηω’ΐΖε.Βδ–ΆΒΡ B-sheet ΫαΙΙ‘Ύ 216 nm ¥Π”– 1 ΗωΗΚΖε, 195 nm ¥Π”– 1 Ηω’ΐΖε, “ρ¥Υ Cyclic-KT ‘Ύ»ή“Κ÷–Ω…Ρή“‘ΈόΙφΦΑ≤ΩΖ÷ B-sheet ΫαΙΙ¥φ‘Ύ. Cyclic-KT “‘ΕΰΝρΦϋΫΪΕύκΡΝ¥ΜΖΜ·, ΙΒΟΕύκΡΒΡΙΙœσœύΕ‘ΙΧΕ®, “ρ¥Υ±μœ÷≥ω≤ΩΖ÷ B-sheet ΕΰΦΕΫαΙΙ. ±Ψ―–ΨΩ÷–Υυ”ΟΕύκΡ≥ΛΕ»ΫœΕΧ, “ρ¥Υ≤ΜΉψ“‘–Έ≥…Οςœ‘ΒΡΕΰΦΕΫαΙΙ, ‘≤Εΰ…ΪΙβΤΉ–≈Κ≈ΉήΧεΫœΒΆ.
2. 7 ΡΘΡβΦΤΥψ
Linear-KT ΚΆ Linear-KS Ε‘ΗοάΦ œ“θ–‘ΨζΚΆ―τ–‘ΨζΨυ±μœ÷≥ωΫœΚΟΒΡ…±ΨζΜν–‘, ΧΊ±π «Ε‘ΗοάΦ œ―τ–‘Ψζ. ’β 2 ΧθΕύκΡΨυΚ§”–ΥΪΜν–‘–ρΝ–, «”… 2 ΧθΨΏ”–ΒΞΜν–‘–ρΝ–ΒΡΕύκΡΆ®ΙΐΦρΒΞΒΡΑ±ΜυΥα≤–ΜυœύΝ§Εχ–Έ≥…, ΒΪ±»œύ”ΠΒΡΨΏ”–ΒΞΜν–‘–ρΝ–ΒΡΕύκΡ±μœ÷≥ωœ‘÷χΧαΗΏΒΡ…±ΨζΜν–‘. “ρ¥Υ±ΨΈΡ≤…”ΟΦΤΥψΡΘΡβΒΡΖΫΖ®ΦΤΥψΝΥΥυ…ηΦΤΒΡΕύκΡ”κΝΉ÷§œύΜΞΉς”Ο ±ΒΡΫαΚœΡή, “‘ΤάΦέΕύκΡΚΆœΗΑϊΡΛΒΡΫαΚœ≥ΧΕ». ΝΉ÷§ «œΗΑϊΡΛΒΡ÷ς“ΣΉι≥…≥…Ζ÷, œΗΨζΒΡœΗΑϊΡΛΚ§”–ΫœΗΏΚ§ΝΩΒΡ DMPG, Εχ≤Η»ιΕ·ΈοΒΡœΗΑϊΡΛ÷– DMPG ΒΡΚ§ΝΩ‘ρΚή…Ό, “ρ¥Υ±ΨΈΡ≤…”Ο DMPG ΈΣ―–ΨΩΕ‘œσΩΦ≤λΤδ”κΕύκΡΒΡΫαΚœΡήΝΠ. ≤…”Ο Gaussian 09 ÷–ΒΡ PM3 ΑκΨ≠―ιΥψΖ®”≈Μ·DMPG ΦΑΗςΕύκΡΒΡΖ÷Ή”ΙΙ–Ά. ΚλΆβΙβΤΉΆΦΧα ΨœΏ–‘ΚΆΜΖΉ¥ΒΡΕύκΡ–Έ≥…’έΒΰΫαΙΙ, ‘≤Εΰ…ΪΙβΤΉ“≤±μΟςΜΖΉ¥ΕύκΡΩ…ΡήΨΏ”–≤ΩΖ÷ B-sheet ΕΰΦΕΫαΙΙ, “ρ¥Υ“‘’έΒΰΫαΙΙ”≈Μ·œΏ–‘ΦΑΜΖΉ¥ΕύκΡ, ‘Ύ”≈Μ·Ιΐ≥Χ÷–ΫΪΕύκΡ”κDMPG Ζ÷Ή”ΒΡΙ«Φή≤ΩΖ÷Ϋχ––ΙΧΕ®“‘ΥθΕΧ”≈Μ· ±Φδ, ΫαΙϊΦϊΆΦ 5. ΗυΨίΕύκΡΚΆ DMPG ΒΡΙΙ–Ά”≈Μ·ΫαΙϊΙΙΫ®ΕύκΡΚΆ DMPG –Έ≥…ΒΡΗ¥ΚœΈο, ≤ΔΕ‘Η¥ΚœΈοΫχ“Μ≤Ϋ”≈Μ·, Η¥ΚœΈοΒΡΙΙ–ΆΦϊΆΦ 6. ΦΤΥψ ±÷ς“ΣΩΦ¬«ΕύκΡ”κ DMPG Ζ÷Ή”ΒΡΨ≤ΒγΚΆ«βΦϋΫαΚœ, Ά§ ±ΩΦ¬«ΨΓΩ…ΡήΕύΒΡΫαΚœΡΘ Ϋ. “‘ΫαΚœΡήΉνΒΆΈΣ‘≠‘ρ[23], ΗυΨίΙΪ Ϋ(2)ΦΤΥψ≥ωΕύκΡ”κ DMPG –Έ≥…ΒΡΗ¥ΚœΈοΚΆΗςΒΞΧε÷°ΦδΒΡΡήΝΩ≤νΦ¥ΒΟΒΫΫαΚœΡή, ΐΨίΝ–”Ύ±μ 3. ”…±μ 3 Ω…Φϊ, Υυ”–ΕύκΡ”κ DMPG Ζ÷Ή”ΫαΚœΙΐ≥ΧΒΡΡήΝΩ≤νΨυΈΣΗΚ÷Β, ΥΒΟςΕύκΡ”κ DMPG ΒΡœύΜΞΉς”ΟΙΐ≥Χ ΆΖ≈ΡήΝΩ, ΡήΝΩ≤νΒΡΨχΕ‘÷Β‘Ϋ¥σΥΒΟςΕΰ’ΏΒΡΫαΚœΡή‘Ϋ¥σ. ≥ΛΝ¥κΡΚΆΜΖΉ¥κΡ”κ DMPG Ζ÷Ή”ΫαΚœ ±ΨΏ”–Ϋœ¥σΒΡΫαΚœΡή, ‘ΕΗΏ”ΎΕΧΝ¥κΡ. ΫαΚœΡή‘Ϋ¥σ, ΥΒΟςΗΟάύΕύκΡΗϋ“Ή”κ DMPG Ζ÷Ή”≤ζ…ζœύΜΞΉς”Ο, ΫχΕχ≤ζ…ζ…±Ψζ–ßΙϊ. DMPG Ζ÷Ή”ΆΖΜυ¥χΗΚΒγ, Ω…”κ¥χ’ΐΒγΒΡΑ±ΜυΥα≤ζ…ζΨ≤ΒγΉς”Ο. DMPG ΒΡΗΚΒγΆΖΜυ‘Ύ”κ≥ΛΝ¥κΡΚΆΜΖκΡΫαΚœΒΡΙΐ≥Χ÷–, Ω…Ά§ ±”κ 2 Ηω¥χ’ΐΒγΒΡάΒΑ±Υα≤ζ…ζΨ≤ΒγΉς”Ο, Εχ‘Ύ”κΕΧΝ¥κΡΒΡΫαΚœΙΐ≥Χ÷–÷ΜΡή”κ 1Ηω¥χ’ΐΒγΒΡάΒΑ±Υα≤ζ…ζΨ≤ΒγΉς”Ο. ’β «‘λ≥…ΕΧΝ¥κΡΒΡΫαΚœΡή‘Ε–Γ”Ύ≥ΛΝ¥κΡ”κΜΖΉ¥κΡΒΡ÷ς“Σ‘≠“ρ. ¥ΥΫαΙϊΚΆ Β―ιΥυΒΟΒΡ…±ΨζΜν–‘ ΐΨίΜυ±Ψ“Μ÷¬, Φ¥ΨΏ”– 2 ΗωΜν–‘–ρΝ–ΒΡ≥ΛΝ¥κΡΚΆΜΖΉ¥κΡ±»ΨΏ”–ΒΞΗωΜν–‘–ρΝ–ΒΡΕΧΝ¥κΡ…±ΨζΜν–‘Ηϋ«Ω.
‘Ύ≥ΛΝ¥κΡΚΆΜΖΉ¥κΡ÷–, –ρΝ–÷–Κ§”–ΥΩΑ±ΥαΚΆΥ’Α±ΥαΒΡΕύκΡ±»œύ”ΠΒΡΚ§”–ΧλΕ§θΘΑΖΒΡΕύκΡΫαΚœΡή¥σ, ’β «“ρΈΣΥΩΑ±ΥαΚΆΥ’Α±ΥαΒΡ Ή¬ ΧΦ…œΒΡτ«ΜυΩ…”κΝΉ÷§Ζ÷Ή” 2 ΧθΧΦΝ¥…œΒΡτ Μυ–Έ≥…«βΦϋ, ¥”Εχ‘ω¥σΝΥΫαΚœΡή. …±Ψζ Β―ιΫαΙϊ“≤÷ΛΟςΚ§”–ΥΩΑ±ΥαΚΆΥ’Α±ΥαΒΡΕύκΡΜν–‘ΗΏ”ΎΚ§ΧλΕ§θΘΑΖΒΡΕύκΡ. Frecer Β»[19]ΦΤΥψ±μΟς, KFTFK, KFSFK ΚΆ KFNFK ”κΡΎΕΨΥΊΒΡΜν–‘≤ΩΖ÷ Lipid A ΫαΚœΒΡ Gibbs Ή‘”…ΡήΖ÷±πΈΣ-67, -75ΚΆ-55 kJ/ mol; KFTFK ΚΆ KFSFK –ρΝ–”κ Lipid A ΒΡΫαΚœΡήΗϋ¥σ, ΗΟΫαΙϊ”κ±ΨΈΡΫαΙϊ“Μ÷¬.
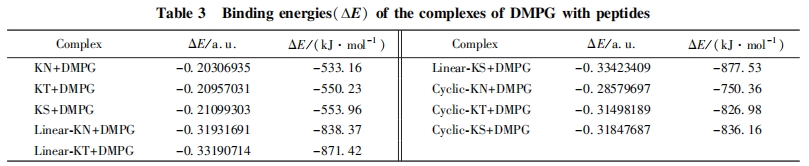
”κΜΖΉ¥κΡœύ±», ≥ΛΝ¥κΡ”κ DMPG Ζ÷Ή”ΒΡΫαΚœΡή¬‘ΗΏ”ΎΜΖΉ¥κΡ, ’βΩ…Ρή «”…”ΎΕύκΡΫαΙΙΒΡ”Αœλ. ≥ΛΝ¥κΡ‘ΎΥ°»ή“Κ÷–Μυ±Ψ“‘ΈόΙφΫαΙΙ¥φ‘Ύ, “ρ¥ΥΕύκΡΫαΙΙ»α–‘Ϋœ¥σ, Ω…ΗυΨίΥυΫαΚœΒΡΕ‘œσ≤…»ΓΗϋ ΚœΒΡΫαΙΙ”κ÷°ΫαΚœ. ΧΊ±π « Linear-KT ΚΆ Linear-KS, Τδ»α–‘ΫαΙΙ ΙΥΩΑ±ΥαΚΆΥ’Α±ΥαΒΡ BΧΦ…œΒΡτ«ΜυΨυΩ…”κDMPG Ζ÷Ή”÷– 2 ΧθΧΦΝ¥…œΒΡτ Μυ–Έ≥…«βΦϋ, ‘ω¥σΫαΚœΡή. ΕχΜΖΉ¥κΡ”…”Ύ ή’έΒΰΫαΙΙΚΆΕΰΝρΦϋΒΡœό÷Τ,κΡΒΡΫαΙΙœύΕ‘ΙΧΕ®, κΡΜΖΒΡ¥σ–Γ ήœό, ΙΝΉ÷§Ζ÷Ή”≤ΜΡήΆξ»Ϊ≤ε»κκΡΜΖ÷–, ÷ΜΡή‘ΎκΡΜΖΤΫΟφΒΡ≤ύΟφ”κΤδΫαΚœ, ÷Μ”κ DMPG Ζ÷Ή”÷– 1 ΧθΧΦΝ¥…œΒΡτ Μυ–Έ≥…«βΦϋ, «βΦϋ ΐΡΩΦθ…Ό, ΫαΚœΡήΫΒΒΆ, “ρ¥ΥΜΖΉ¥κΡΒΡΜν–‘≤ΜΦΑ≥ΛΝ¥κΡ.
3 Ϋα ¬έ
…ηΦΤΚœ≥…ΝΥΕύΗωΨΏ”– 2 ΗωΜν–‘–ρΝ–ΒΡœΏ–‘ΚΆΜΖΉ¥ΕύκΡΦΑΨΏ”–ΒΞΗωΜν–‘–ρΝ–ΒΡΕΧΝ¥ΕύκΡ, ―–ΨΩΝΥΤδ…±ΨζΜν–‘, ≤Δ≤…”ΟΦΤΥψΡΘΡβΒΡΖΫΖ®ΦΤΥψΝΥΕύκΡ”κœΗΨζœΗΑϊΡΛ÷–÷Ί“Σ≥…Ζ÷ΝΉ÷§θΘΗ ”ΆΒΡΫαΚœΡή, ¥”άμ¬έ…œΫβ ΆΝΥΤδΜν–‘. ΫαΙϊ±μΟς, Ψ≤ΒγΉς”ΟΚΆ«βΦϋ‘ΎΕύκΡ”κΝΉ÷§ΒΡΫαΚœΙΐ≥Χ÷–Τπ÷Ί“ΣΉς”Ο. œΏ–‘ΒΡ Linear-KT ΚΆ Linear-KS ΨΏ”–ΥΪΜν–‘–ρΝ–, Ω…Ά§ ±ΧαΙ© 2 ΗωΚ…’ΐΒγΑ±ΜυΥα”κΝΉ÷§ΫαΚœ, …±ΨζΜν–‘‘Ε‘Ε¥σ”ΎΒΞΜν–‘–ρΝ–. ΫΪΥΪΜν–‘–ρΝ–“ΐ»κΆ§“ΜΖ÷Ή”, Ω…ΤπΒΫΧαΗΏ…±ΨζΜν–‘ΒΡΉς”Ο. Ά§ ± Linear-KT ΚΆ Linear-KS ΒΡ»α–‘ΫαΙΙΡήΙΜ ΙΕύκΡ”κΝΉ÷§…œΒΡτ Μυ–Έ≥…ΕύΗω«βΦϋ, “ρ¥ΥΫαΚœΡήΫœ¥σ, ΨΏ”–Ϋœ«ΩΒΡ…±ΨζΜν–‘. ΦΤΥψΡΘΡβΒΡΖΫΖ®ΈΣΩΙΨζκΡΒΡ…±ΨζΜν–‘‘Λ≤β¥”άμ¬έ…œΧαΙ©ΝΥ“ΜΕ®ΒΡ“άΨί. άϊ”ΟΗΟΖΫΖ®Ω…‘Ύ“ΜΕ®≥ΧΕ»…œ‘Λ≤βΩΙΨζκΡΒΡ…±ΨζΜν–‘, ΧαΗΏΩΙΨζκΡΒΡ―–ΖΔ–ß¬ , ΈΣΩΙΨζκΡΒΡ…ηΦΤ”κΩΣΖΔΧαΙ©ΝΥ–¬ΆΨΨΕ.
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷ΨΓΘ
