
’Σ “Σ Ϋπ τ‘ΣΥΊ≥ΘΦϊ”ΎΧλ»ΜΒΑΑΉ÷ Ζ÷Ή”÷–Θ§≤ΔΑγ―ίΉ≈≤ΜΩ…Μρ»±ΒΡ÷Ί“ΣΫ«…ΪΓΘΕ‘Ϋπ τ”κΑ±ΜυΥαΒΡ≈δΈΜΖΫ ΫΒΡ≤ΜΕœ…ν»κάμΫβΘ§”–÷ζ”Ύ»ΥΙΛ…ηΦΤ–¬–ΆΒΡΫπ τ⁃ΕύκΡ≈δΈΜΡΘ ΫΘ§≤Δ‘ΎΕύκΡΫαΙΙΙΠΡή”≈Μ·ΓΔΖ÷Ή”ΉιΉΑΓΔ±ξΦ«”κ≥…œώ“‘ΦΑ“©Έο…ηΦΤΒ»Νλ”ρΜώΒΟΗϋΙψΖΚΒΡ―–ΨΩΚΆ”Π”ΟΓΘ±ΨΈΡ Ήœ»Φρ“ΣΫι…ήΒΑΑΉ÷ ÷–ΒΡ≤ΜΆ§ΜυΆ≈”κΫπ τάκΉ”÷°ΦδΒΡ≈δΈΜΡΘ Ϋ“‘ΦΑΨΏΧεΒΡ―–ΨΩΑΗάΐΘ§ΫχΕχΫι…ήΝΥΫπ τ≈δΈΜ‘ΎΕύκΡΖ÷Ή”ΒΡΕΰΦΕΫαΙΙΒςΩΊΓΔ≥§Ζ÷Ή”ΫαΙΙΉιΉΑΓΔΕύκΡΦΑΒΑΑΉ÷ ΒΡ”ΪΘ®ΝΉΘ©Ιβ±ξΦ«ΓΔΒΑΑΉ÷ ’έΒΰΖΫ ΫΖ÷ΈωΒ»Μυ¥Γ―–ΨΩ÷–ΒΡ”Π”ΟΓΘΉνΚσΘ§Ε‘…œ ω―–ΨΩΝλ”ρΈΣΫπ τ⁃ΕύκΡάύΖ÷Ή”ΧΫ’κΜρ“©ΈοΒΡ…ηΦΤ”≈Μ·ΥυΧαΙ©ΒΡΖΔ’Ι«±ΝΠΫχ––’ΙΆϊΓΘ
‘ΎΧλ»Μ…ζΈοΖ÷Ή”÷–Θ§Ϋπ τ‘ΣΥΊΑγ―ίΉ≈≤ΜΩ…Μρ»±ΒΡ÷Ί“ΣΫ«…ΪΓΘάΐ»γ‘Ύ“―÷ΣΫαΙΙΒΡΒΑΑΉ÷ Ζ÷Ή”Β±÷–Θ§¥σ‘Φ”–Ϋϋ“ΜΑκΒΡΖ÷Ή”Κ§”–“ΜΗωΜρΕύΗωΫπ τ‘ΣΥΊΓΘ’β–©Ϋπ τ‘ΣΥΊΆ®Ιΐ≈δΈΜœύΜΞΉς”ΟΘ§Ά§κΡΝ¥÷–ΒΡΏδΏρΜυΓΔέœΜυΓΔτ»ΜυΓΔΑ±ΜυΜρΤδΥϊΜυΆ≈–Έ≥…Έ»Ε®ΫαΙΙΘ§‘ΎΕύ÷÷÷Ί“Σ…ζΈο―ßœ÷œσΘ§»γΙβΚœΉς”ΟΓΔΟΗ¥ΏΜ·Ζ¥”ΠΓΔΘΡΘΈΘΝ Η¥÷ΤΓΔΘ“ΘΈΘΝ ΉΣ¬ΦΓΔΒΑΑΉ÷ Ζ≠“κΒ»÷–ΤπΉ≈ΙΊΦϋΉς”ΟΓΘΥφΉ≈ΕύΡξά¥ΒΡ―–ΨΩΘ§»ΥΟ«Ε‘“Μ–©≥ΘΦϊΫπ τ‘ΣΥΊ»γΧζΓΔΆ≠ΓΔΟΧΓΔ–ΩΓΔΗΤΓΔΟΨΘ§“‘ΦΑΉ‘»ΜΫγΫœ…ΌΦϊΒΡΫπ τ‘ΣΥΊ»γΡχΓΔΖΑΓΔ≤§ΓΔΫπΓΔ“ΩΜράύΫπ τ‘ΣΥΊ»γ…ιΒ»”κΒΑΑΉ÷ ΕύκΡΖ÷Ή”÷°ΦδΉς”ΟΜζάμΒΡΝΥΫβ≤ΜΕœΦ”…νΓΘ’β–©»œ ΕΫχ“Μ≤ΫΆΤΕ·ΝΥ–¬–ΆΫπ τ≈δΈΜΜ·ΚœΈοΒΡ»ΥΙΛ…ηΦΤ”κΚœ≥…Θ§≤Δ”Ο”ΎΒΑΑΉ÷ ΚΆΕύκΡΒΡΙΠΡή―–ΨΩΓΘ
Ϋπ τ‘ΣΥΊ‘ΎΧλ»Μ…ζΈοΖ÷Ή”ΧεœΒ÷–ΒΡΉς”Ο¥σ÷¬Ω…“‘Ζ÷ΈΣΝΫΗωΖΫΟφΘ§“ΜΖΫΟφ «Ϋπ τΕ‘ΒΑΑΉ÷ Ζ÷Ή”ΤπΒΫΝΥΫαΙΙ–‘ΒΡΈ»Ε®”κΈ§≥÷Ής”ΟΘ§άΐ»γ‘ΎΚΥΥαΉΣ¬Φ“ρΉ”÷–≥ΘΦϊΒΡ–Ω÷ΗΫαΙΙΘ§–Ω‘≠Ή”Ά®Ιΐ”κΒΑΑΉ÷ –ρΝ–÷–ΒΡΉιΑ±Υα”κΑκκΉΑ±ΥαΒΡΉιΚœ≈δΈΜΘ§–Έ≥…Ζ«≥ΘΧΊ’ςΒΡ Π¬ΉΣΫ«ΫαΙΙΘ§“‘¬ζΉψ”κ ΘΡΘΈΘΝ ΫαΙΙœύΫαΚœΒΡ–η“ΣΓΘΝμ“ΜΖΫΟφ «Ϋπ τΦΑΤδΗ¥ΚœΈο≤Έ”κΝΥ…ζΈοΙΐ≥Χ≤ΔΧαΙ©≤ΜΩ…Χφ¥ζΒΡΙΠΡή–‘Ής”ΟΘ§άΐ»γ‘Ύ―ΣΚλΒΑΑΉ÷–ΒΡΕΰΦέΧζάκΉ”Θ§Ά®Ιΐ”κ―θΖ÷Ή”ΒΡΫαΚœΚΆ ΆΖ≈Θ§≥–ΒΘΝΥ‘Ύ―Σ“Κ÷– δ―θΒΡΙΠΡήΓΘ‘Ύ»ΥΙΛ…ηΦΤΒΡΫπ τΘ®άύΫπ τΘ© ”κΕύκΡΒΑΑΉ÷ ΒΡ≈δΈΜœύΜΞΉς”Ο÷–Θ§Ϋπ τ“≤Ω…“‘¥”…œ ωΝΫΗωΖΫΟφΖΔΜ”Ής”ΟΘ§άΐ»γΧαΙ©’κΕ‘ΧΊΕ®ΕύκΡ–ρΝ–ΜρΒΑΑΉ÷ ΙΙœσΒΡΫαΙΙΈ»Ε®”κΉιΉΑΙΠΡήΘ§“‘ΦΑΧαΙ©Αϋά®”ΪΘ®ΝΉΘ©Ιβ≥…œώΓΔΖ≈…δ–‘÷ΈΝΤΓΔΚΥ¥≈‘λ”ΑΒ»ΡΎ‘ΎΙΠΡήΘ§≤ΔΩ…‘ΎΫχ“Μ≤Ϋ”≈Μ·…ηΦΤΚσΙΙΫ®ΨΏ”–Η¥ΚœΙΠΡήΒΡ…ζΈοΧΫ’κΜρ÷ΈΝΤΖ÷Ή”Θ§”Ο”Ύ…ζΈο“Ϋ―ßΒΡ―–ΨΩΓΘ
±ΨΈΡ Ήœ»ΜΊΙΥΚΆάΐΨΌΕύκΡ”κΒΑΑΉ÷ ÷–ΒΡ≤ΜΆ§ΜυΆ≈”κΫπ τΜράύΫπ τάκΉ”÷°ΦδΒΡ÷ς“Σ≈δΈΜΉς”ΟΖΫ ΫΘ§ΫχΕχΙιΡ…ΫϋΡξά¥‘ΎΗΟΝλ”ρ÷–±»ΫœΆΜ≥ωΒΡΦΗάύ”Π”Ο ΒάΐΘ§άΐ»γΫπ τ≈δΈΜΜ·ΚœΈο‘ΎΕύκΡΖ÷Ή”ΒΡΕΰΦΕΫαΙΙΒςΩΊΓΔ≥§Ζ÷Ή”ΫαΙΙΉιΉΑΓΔΕύκΡΦΑΒΑΑΉ÷ ΒΡ”ΪΘ®ΝΉΘ© Ιβ±ξΦ«”κ ΨΉΌΓΔΒΑΑΉ÷ ΫαΙΙ”ρ’έΒΰΖΫ ΫΖ÷ΈωΒ»Μυ¥Γ―–ΨΩΝλ”ρΒΡ≤ΜΆ§≥ΧΕ»ΒΡ”Π”ΟΘ§≤Δ’ΙΆϊΝΥ…œ ω―–ΨΩΈΣΫπ τ⁃ΕύκΡάύΖ÷Ή”ΧΫ’κΜρ“©ΈοΒΡ…ηΦΤ”≈Μ·ΥυΧαΙ©ΒΡΖΔ’Ι«±ΝΠΓΘ
Θ≤ ΕύκΡ–ρΝ–÷–ΒΡ≈δΈΜΜ·ΚœΈοΫαΚœΈΜΒψ
‘Ύ–μΕύ…ζΟϋΜνΕ·÷–Θ§Ϋπ τάκΉ”ΒΡ≈δΈΜΩ…“‘”––ßΒΊ”ΑœλΕύκΡΒΡΕΰΦΕΫαΙΙΘ§«ΐΕ·ΒΑΑΉ÷ ’έΒΰΚΆΉιΉΑΘ§¥”ΕχΒςΩΊΙΠΡήΒΑΑΉ÷ ΒΡ…ζΈοΜν–‘ΓΘΝΥΫβΕύκΡΝ¥÷–ΗςΜυΆ≈”κ≤ΜΆ§Ϋπ τάκΉ”ΒΡ≈δΈΜΖΫ ΫΚΆΡήΝΠΫΪ”–÷ζ”Ύ»ΥΙΛ…ηΦΤΚΆ”≈Μ·ΕύκΡ”κΫπ τΒΡ≈δΈΜΫαΙΙΓΘΕχΕ‘Χλ»ΜΒΡΙΠΡήΕύκΡΤ§ΕΈΘ§‘ρ”–ΆϊΆ®Ιΐ≈δΈΜΜ·ΚœΈοΒΡΫαΚœΘ§ΡΘΡβΧλ»ΜΒΡΫαΙΙΘ§ΜώΒΟ”≈Μ·ΒΡ…ζΈοΜν–‘ΓΘΉςΈΣΕύκΡΒΡΫαΙΙΒΞ‘ΣΘ§≥ΘΦϊΒΡ Θ≤ΘΑ ÷÷Α±ΜυΥαΨυ”… ΘΟΓΔΘ»ΓΔΘœΓΔΘΈΓΔΘ” Β»‘ΣΥΊΉι≥…ΓΘΤδ÷–Θ§“‘τ»ΜυΘ®ΓΣΘΟΘœΘœΘ»Θ©÷–ΒΡ―θ‘≠Ή”ΓΔΏδΏρΜυΆ≈ΥυΚ§ΒΡΒΣ‘≠Ή”“‘ΦΑέœΜυΘ®ΓΣΘ”Θ»Θ©ΥυΚ§ΒΡΝρ‘≠Ή”ΒΡΗχΒγΉ”ΡήΝΠΉν«ΩΘ§Ήν“Ή≤Έ”κ≈δΈΜΖ¥”ΠΓΘάϊ”ΟΒΑΑΉ÷ ΐΨίΩβΘ®Θ–ΘρΘοΘτΘεΘιΘν ΘΡΘαΘτΘα Θ¬ΘαΘνΘκΘ§ Θ–ΘΡΘ¬Θ©Ζ÷ΈωΖΔœ÷Θ§‘Ύ…ζΈοœΒΆ≥÷–Ης÷÷Α±ΜυΥα≤–Μυ≤Έ”κΫπ τάκΉ”≈δΈΜΒΡΤΒ¬ ΉνΗΏΒΡ «Ι»Α±ΥαΘ®Θ«ΘλΘθΘ©ΚΆΧλΕ§Α±ΥαΘ®ΘΝΘσΘπΘ©ΒΡτ»ΜυΘ§ΉιΑ±ΥαΘ® Θ»ΘιΘσΘ© ΒΡ Ώδ Ώρ Μυ Ά≈ “‘ ΦΑ Ακ κΉ Α± Υα Θ® ΘΟΘυΘσΘ© ΒΡ έœΜυΘέΘ±Θ§Θ≤ΘίΓΘ
≈δΈΜΖ¥”Π÷–Ϋπ τάκΉ”ΒΡ―Γ‘ώΤΪΚΟΘ§‘ρ“άΨίΥυ―ΓΕ®ΒΡΕύκΡΟΣΕ®ΈΜΒψΕχ”–Υυ≤ΜΆ§Θ§÷ς“ΣΉώ―≠ΒΡ‘≠άμ «»μ”≤ΥαΦνάμ¬έΓΘάΐ»γΘ§έœΜυ τ”Ύ»μΦνΘ§“ρ¥ΥΒ±ΫαΚœΈΜΒψΈΣ ΘΟΘυΘσ ±Θ§ΘΟΘθΘ®ΔρΘ©ΓΔΘ»ΘγΘ®ΔρΘ©ΓΔΘΟΘδΘ®ΔρΘ© Β»»μΥαΫπ τάκΉ”≤Έ”κ≈δΈΜΒΡΡήΝΠΗϋ«ΩΓΘΕχΕ‘”ΎΚ§”–”≤Φντ»ΜυΘ®ΓΣΘΟΘœΘœΘ»Θ© ΒΡ ΘΝΘσΘπ Θ· Θ«ΘλΘθ Εχ ―‘Θ§ ”≤ Υα Ϋπ τ άκ Ή” ΘΆΘνΘ®ΔρΘ©ΓΔΘΆΘγΘ®ΔρΘ©ΓΔΘΟΘαΘ®ΔρΘ© Ω…“‘ΧαΙ©Ηϋ«ΩΒΡ≈δΈΜΡήΝΠΒ»ΓΘΕ‘”ΎΉιΑ±ΥαΒΡΏδΏρΜυΆ≈Εχ―‘Θ§Τδ”κΗςάύ»μΓΔ”≤Ϋπ τάκΉ”÷°ΦδΕΦΨΏ”–”≈‘ΫΒΡ≈δΈΜΉς”ΟΡήΝΠΘ§“ρ¥ΥΤδΨΏ”–Ήν«ΩΒΡΆ®”Ο–‘Θ®±μΘ±Θ©ΓΘ
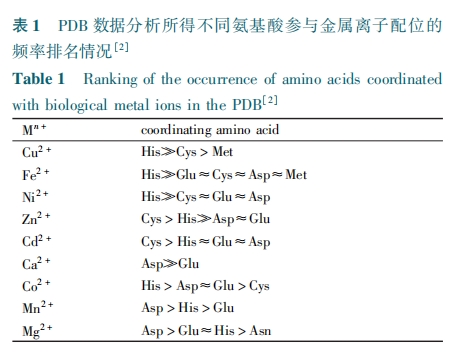
Θ≤.Θ± τ»Μυ
τ»Μυ‘Ύ…ζάμ ΘπΘ» ΧθΦΰœ¬Άξ»Ϊ»Ξ÷ Ή”Μ·Θ§ΧαΙ©ΨΏ”–Κή«Ω≈δΈΜΡήΝΠΒΡΓΣΘΟΘœΘœΘ≠ ΜυΆ≈Θ§‘Ύ”κΫπ τάκΉ”ΒΡœύΜΞΉς”Ο÷–Αγ―ί÷Ί“ΣΒΡΫ«…ΪΓΘάΐ»γΦΙΉΒΕ·ΈοΒΡΦΓ»β ’ΥθΨΆ–η“ΣΆ®ΙΐΦΓΗΤΒΑΑΉ”κ ΘΟΘαΘ®ΔρΘ© ΒΡœύΜΞΉς”Οά¥Ϋχ––ΒςΩΊΘέΘ≥ΘίΓΘ“ΜΑψά¥ΥΒΘ§ΦΓΗΤΒΑΑΉ÷– ΘΟΘαΘ®ΔρΘ© ΫαΚœΈΜΒψ”–ΕύΗωΘ§Εχ‘ΎΈόΦΙΉΒΕ·Έο÷–Θ§»¥Μα“ρΈΣΙΊΦϋΫαΚœΈΜΒψΒΡΑ±ΜυΥα≤ΜΆ§Θ§ΕχΕΣ ß≤ΩΖ÷ ΘΟΘαΘ®ΔρΘ© ΒΡρϋΚœΡήΝΠΘέΘ¥ΘίΓΘ≤Έ”κ ΘΟΘαΘ®ΔρΘ© ≈δΈΜΒΡΙΊΦϋΑ±ΜυΥα≤–Μυ±ψ «ΘΝΘσΘπ ΚΆ Θ«ΘλΘθ ΒΡ τ» ΜυΘέΘΒΘίΓΘΫϋ ΤΎ Έß »Τ ≈Ν Ϋπ …≠ ≤ΓΘ®Θ–ΘαΘρΘκΘιΘνΘσΘοΘν̓Θσ ΘδΘιΘσΘεΘαΘσΘεΘ§ Θ–ΘΡΘ©ΒΡ―–ΨΩ±μΟςΘ§Θ–ΘΡ œύΙΊΜυ“ρΘ–ΘΝΘ“ΘΥΘΙ ±ύ¬κΒΡΒΑΑΉ÷ ÷ς“ΣΆ®Ιΐ ΘΝΘσΘπ ΚΆ Θ«ΘλΘθ ΒΡτ»Μυ“‘ΦΑ Θ»ΘιΘσ ΒΡΏδΏρΜυΆ≈ρϋΚœ ΘΆΘνΘ®ΔρΘ©Θ§ά¥Ζά÷Ι ΘΆΘνΘ®ΔρΘ©“ΐΖΔΒΡ Θ–ΘΡ œύΙΊ÷ΔΉ¥ΘέΘΕΘίΓΘ≥ΘΦϊΒΡΆ®Ιΐ ΘΝΘσΘπ ΚΆ Θ«ΘλΘθ ”κΕύκΡ≈δΈΜΫαΚœΒΡΙΐΕ…Ϋπ τ÷ς“ΣΑϋά®ΘΚΘΟΘα Θ® ΔρΘ©ΓΔΘΆΘγΘ®ΔρΘ©ΓΔΘΆΘνΘ®ΔρΘ©Β»ΘέΘΖΘίΓΘ
Θ≤.Θ≤ Α±ΜυΚΆΏδΏρΜυΆ≈
Ά®Ιΐ ΘΈ ‘≠Ή”ΧαΙ©≈δΈΜΒγΉ”ΒΡΜυΆ≈÷ς“ΣΑϋά®ΓΣΘΈΘ»Θ≤ΚΆΏδΏρΜυΆ≈ΓΘΈΜ”ΎκΡΝ¥÷ςΝ¥ ΘΈ Ρ©ΕΥΒΡ≤°ΑΖΜυΆ≈Θ®ΠΝ⁃ΘΈΘ»Θ≤ Θ§ΘπΘΥΘα ΘΫ ΘΗ.ΘΙΘ©ΨΏ”–ΫœΚΟΒΡ≤Έ”κ≈δΈΜΒΡΡήΝΠΘ§œύΕ‘Εχ―‘άΒΑ±Υα≤ύΝ¥Ρ©ΕΥΒΡΑ±Μυ Θ® Π≈⁃ΘΈΘ»Θ≤ Θ§ ΘπΘΥΘα ΘΫΘ±ΘΑ.ΘΖΘΙΘ©‘Ύ÷––‘ΧθΦΰœ¬ΒΡ≈δΈΜΡήΝΠ‘ρΚή»θΓΘ‘Ύ≥ΘΦϊΒΡ Θ≤ΘΑ ÷÷Α±ΜυΥα÷–Θ§ΉιΑ±Υα”κΫπ τάκΉ”÷°ΦδΒΡ≈δΈΜΉς”ΟΘ§‘ΎΧλ»Μ…ζΟϋΜνΕ·Ιΐ≥Χ÷–ΨΏ”– °Ζ÷÷Ί“ΣΒΡ“β“εΓΘΉιΑ±ΥαΒΡΏδΏρΜυΆ≈…œΒΡ Θ≥ ΈΜΒΣ‘≠Ή”‘Ύ÷––‘ΧθΦΰœ¬Θ®ΘπΘΥΘα ΘΫ ΘΕ.ΘΑΘ¥Θ©Φ¥Ω…»Ξ÷ Ή”Μ·Θ§ΈΣΫπ τάκΉ””κΒΑΑΉ÷ ΒΡΫαΚœΑσΕ®ΧαΙ©ΝΥ÷Ί“ΣΒΡΫ”»κΈΜΒψΘ§ΒςΩΊΗς÷÷ΒΑΑΉ÷ ΒΡΙΠΡήΓΘ
άΐ»γΘ§‘Ύ”κ―ΣΧ«ΒςΫΎΟή«–œύΙΊΒΡ“»ΒΚΥΊΙΠΡήΒςΩΊ÷–Θ§ΘΎΘνΘ®ΔρΘ© ΚΆΉιΑ±ΥαΏδΏρΜυΆ≈ΒΡ≈δΈΜΤπΉ≈ΙΊΦϋΒΡΒςΩΊΉς”ΟΓΘ“»ΒΚΥΊΉν≥θΖ÷ΟΎ ±Θ§“‘ ΘΎΘνΘ®ΔρΘ© Έ»Ε®ρϋΚœΒΡΖ«Μν–‘ΝυΨέΧε–Έ Ϋ¥φ‘ΎΘέΘΗΘίΘ®ΆΦ Θ±ΘαΘ©Θ§ΕχΚσ“‘ΤδΨΏ”–…ζΈοΜν–‘ΒΡΒΞΧε–Έ Ϋ”κœ¬”Έ ήΧεœύΜΞΉς”ΟΘ§ΒςΩΊœ¬”Έ–≈Κ≈Ά®¬ΖΙΊΦϋΖ÷Ή”ΒΡΝΉΥαΜ·Υ°ΤΫΘ§¥”ΕχΩΊ÷ΤœΗΑϊΕ‘Χ«άύΒΡ¥ζ–ΜΓΘ―–ΨΩ±μΟςΘέΘΙΘ§Θ±ΘΑΘίΘ§¥”»ΥάύΡρ“Κ÷–Ζ÷άκΒΟΒΫΒΡΥΡκΡ–ρΝ– Θ«Θ»Θ‘ΘΡΘ®Θ«ΘλΘυ⁃Θ»ΘιΘσ⁃Θ‘ΘηΘρ⁃ΘΝΘσΘπΘ©Θ§ΨΏ”–ΫΒΒΆ―ΣΧ«ΒΡΙΠ–ßΓΘΫχ“Μ≤Ϋ≤ϊ ΆΤδΜζάμΖΔœ÷Θ§ΨΏ”–Θ»Θ‘ΘΡ Μυ–ρΒΡΕΧκΡΩ…“‘Ά®Ιΐ ΘΈ ΕΥ”ΈάκΒΡΑ±ΜυΚΆΝΎΫϋΘΈ Ρ©ΕΥΒΡΉιΑ±ΥαρϋΚœ ΘΎΘνΘ®ΔρΘ©Θ§¥Ό Ι“»ΒΚΥΊΖ÷…Δ≥…ΒΞΧε–Έ ΫΘ§“‘ΦΛΜν“»ΒΚΥΊΫΒΒΆ―ΣΧ«ΒΡ…ζΈοΙΠΡήΓΘ
ΝμΨί±®ΒάΘ§ΘΟΘθΘ®ΔρΘ© ΚΆ ΘΎΘνΘ®ΔρΘ© ΕΦΩ…“‘”κ Π¬⁃ΒμΖέ―υΒΑΑΉΕύκΡ Θ® ΘαΘμΘυΘλΘοΘιΘδ Π¬Θ§ ΘΝΠ¬Θ© ΫαΚœΘ§ΒΦ÷¬ ΘΝΠ¬ ΨέΦ·Θ§”κάœΡξ≥’¥τ÷Δ Θ®ΘΝΘλΘζΘηΘεΘιΘμΘεΘρ.Θσ ΘδΘιΘσΘεΘαΘσΘεΘ© ΒΡΫχ≥ΧΟή«–œύΙΊΘέΘ±Θ±ΘίΓΘΘΟΘθΘ®ΔρΘ© Μρ ΘΎΘνΘ®ΔρΘ© ΒΡ≈δΈΜΜαΒΦ÷¬ ΘΝΠ¬ ΕύκΡΒΡΫαΙΙ±δΜ·ΘέΘ±Θ≤Θ§Θ±Θ≥ΘίΘ§≤Έ”κ≈δΈΜΒΡΙΊΦϋΈΜΒψ’ΐ «ΕύκΡ–ρΝ–÷– ΘΈ Ρ©ΕΥΒΡΑ±ΜυΚΆ»ΐΗωΉιΑ±ΥαΒΡΏδΏρΜυΆ≈Θ®ΆΦΘ± ΘβΘ©ΓΘ
’β–©άΐΉ”ΕΦ±μΟςΘ§’κΕ‘ΧΊΕ®Ϋπ τάκΉ”Υυ…ηΦΤΒΡΕύκΡάύρϋΚœΦΝΘ§‘ΎœύΙΊΦ≤≤ΓΒΡ÷ΈΝΤ÷–ΨΏ”–ΙψΖΚΒΡ”Π”Ο«ΑΨΑΓΘΕχΝμ“ΜΖΫΟφΘ§“≤œ‘ Ψ≥ωΉιΑ±ΥαΒΡΏδΏρΜυΆ≈ΉςΈΣΙΊΦϋΑ±ΜυΥα≤–ΜυΘ§≤Έ”κ≈δΈΜΖ¥”ΠΒΡΡήΝΠΚΆ”Π”ΟΒΡΙψΖΚ–‘ΓΘ“ρ¥ΥΘ§‘ΎΕύκΡΒΡ…ηΦΤ÷–Θ§ΉιΑ±ΥαΩ…“‘ΚήΚΟΒΡΈΣ≈δΈΜΜ·ΚœΈοΒΡΫαΚœΧαΙ©ΈΜΒψΓΘ≥Θ”ΟΒΡΆ®ΙΐΉιΑ±Υα”κΕύκΡ≈δΈΜΫαΚœΒΡΙΐΕ…Ϋπ τ÷ς“ΣΑϋά®ΘΚΘΎΘνΘ®ΔρΘ©ΓΔΘΟΘθΘ®ΔρΘ©ΓΔΘΈΘιΘ®ΔρΘ©ΓΔΘΟΘοΘ®ΔρΘ©ΓΔΘ–ΘδΘ®ΔρΘ©ΦΑ Θ“ΘθΘ®ΔρΘ©ΘέΘ±Θ¥Θί ΚΆΘ“ΘθΘ®ΔσΘ©ΘέΘ±ΘΒΘίΒ»ΓΘ
Θ≤.Θ≥ έœΜυ
‘ΎΒΑΑΉ÷ ’έΒΰΒΡΙΐ≥Χ÷–Θ§ΦΗΚΈΩ’ΦδΙΙœσ…œœύΝΎΒΡΑκκΉΑ±Υα÷°ΦδΆ®ΙΐέœΜυ–Έ≥…ΒΡΕΰΝρΦϋ «Έ»Ε®ΒΑΑΉ÷ ΧΊ βΫαΙΙΒΡ÷Ί“ΣΖΫΖ®ΓΘ¥ΥΆβΘ§έœΜυ“≤Ω…“‘Ά®Ιΐ”κΫπ τ≈δΈΜ Βœ÷Ε‘ΒΑΑΉ÷ ΫαΙΙΒΡΈ»Ε®Θ§άΐ»γ ΘΎΘνΘ®ΔρΘ©”κ«ΑΈΡΧαΒΫΒΡ–Ω÷ΗΫαΙΙΒΑΑΉ–ρΝ–÷–ΧΊΕ®ΈΜΒψΒΡ Θ¥ ΗωΑκκΉΑ±ΥαΜρ Θ≤ ΗωΑκκΉΑ±ΥαΚΆ Θ≤ ΗωΉιΑ±Υα≈δΈΜΘ§«ΐΕ·ΒΑΑΉ÷ ’έΒΰ–Έ≥…ΨΏ”–ΙΠΡήΜν–‘ΒΡ ÷÷ΗΉ¥ΫαΙΙΘ®ΆΦ Θ≤Θ©ΓΘ

Ρ≥–©ΗΜΚ§ΑκκΉΑ±ΥαΓΔ”–ΆΜ≥ωΒΡΫπ τ≈δΈΜΡήΝΠΒΡΒΑΑΉ÷ ±ΜΆ≥≥ΤΈΣΫπ τΝρΒΑΑΉΘ®ΘμΘεΘτΘαΘλΘλΘοΘτΘηΘιΘοΘνΘεΘιΘνΘσΘ©ΘέΘ±ΘΗΘίΓΘ’ΐ «έœΜυ÷–ΝρΘ® Θ”Θ©‘≠Ή”ΝΦΚΟΒΡΗχΒγΉ”ΡήΝΠΘ§ ΙΤδΩ…“‘ΉςΈΣΝΦΚΟΒΡΒγΉ”Ι©Χε≤Έ”κ≈δΈΜΖ¥”ΠΘ§¥”Εχ»ΟΑκκΉΑ±Υα≥…ΈΣΕύκΡ–ρΝ–÷–ΝΦΚΟΒΡΟΣΕ®ΈΜΒψΘ§±ΜΙψΖΚ”Π”Ο”Ύ≈δΈΜΜ·ΚœΈο≤Έ”κΒΡΕύκΡΫαΙΙΈ»Ε®ΒΡ…ηΦΤΝλ”ρΓΘ”κΚ§”–ΑκκΉΑ±ΥαΒΡΕύκΡ≈δΈΜΫαΚœΡήΝΠΫœ«ΩΒΡΫπ τάκΉ”ΜΙΑϋά®ΘΚΘ–Θβ Θ®ΔρΘ©ΘέΘ±ΘΙΘίΘ§ΘΟΘδ Θ®ΔρΘ©ΘέΘ≤ΘΑΘ§Θ≤Θ±ΘίΘ§ Θ»Θγ Θ®ΔρΘ©ΘέΘ≤Θ≤ΘίΘ§ΘΈΘιΘ®ΔρΘ©ΘέΘ≤Θ≥ΘίΘ§“‘ΦΑάύΫπ τ‘ΣΥΊάΐ»γœ¬ΈΡΫΪΧαΒΫΒΡ ΘΝΘσΘ®ΔσΘ©Β»ΓΘ
Θ≥ Ϋπ τ≈δΈΜΕ‘ΕύκΡΫαΙΙΒΡ”Αœλ
Χλ»ΜΕύκΡ–ρΝ–”…”ΎΫαΙΙ…œΙΐΕ»»α–‘Θ§ΒΦ÷¬Έ»Ε®–‘≤νΘ§ΟφΝΌΉ≈“Ή±ΜΧεΡΎ”»Τδ «ΈΗ≥ΠΒάΒΡΗςάύΟΗΥ°ΫβΘ§ΧεΡΎ―≠ΜΖ÷ΆΝτ ±ΦδΕΧΒ»Έ ΧβΓΘΙΠΡήΕύκΡΒΡ…ζΈοάϊ”Ο¬ ΒΆΘ§œό÷ΤΝΥΕύκΡΉςΈΣ“©Έο÷ΤΦΝΒΡ ΒΦ ΝΤ–ßΘέΘ≤Θ¥ΘίΓΘ¥σ≤ΩΖ÷¥Π”ΎΒΑΑΉ÷ ”κΒΑΑΉ÷ œύΜΞΉς”ΟΘ®Θ–Θ–Θ…ΘσΘ©ΙΊΦϋΫγΟφΒΡΕύκΡ–ρΝ–ΒΡΫαΙΙΧΊ’ς”κΕύκΡΒΡ…ζΈοΜν–‘÷°Φδ¥φ‘ΎΉ≈±Ί»ΜΒΡΝΣœΒΓΘ≈δΈΜΜ·ΚœΈοΒΡΑσΕ®Θ§Ε‘ΕύκΡΫαΙΙΨΏ”–œ‘÷χΒΡ”ΑœλΘΚ“ΜΖΫΟφΘ§Χεœ÷‘ΎΕ‘ΕύκΡΫαΙΙ»α–‘ΒΡΗΡ…ΤΘ§ ΙΤδΜώΒΟ ήΒΫœό÷ΤΒΡΈ»Ε®ΙΙœσΘ§”––ßΒΊΧαΗΏΩΙΟΗΫβΡήΝΠΚΆ Ε±π ήΧεΒΡ―Γ‘ώ–‘ΘΜΝμ“ΜΖΫΟφΘ§’β÷÷≈δΈΜΩ…“‘«ΐΕ·ΫαΙΙΥ……ΔΒΡΕύκΡ–ρΝ–’έΒΰΜρΉιΉΑ–Έ≥…ΙΧΕ®ΒΡΫαΙΙΘ§¥”ΕχΨΏ”–ΗϋΚΟΒΡ…ζΈοΜν–‘ΓΘ
“ρ¥ΥΘ§»ΥΙΛ…ηΦΤΒΡΙΠΡήΕύκΡ–ρΝ–«ΑΧεΘ§Ά®Ιΐ‘ΎΧΊΕ®ΈΜΒψ“ΐ»κΒΡΧΊ βΑ±ΜυΥα≤–ΜυΈΣ≈δΈΜΜ·ΚœΈοΒΡΑσΕ®ΧαΙ©ΈΜΒψΘ§Φ¥Ω…“‘≈δΈΜΫαΚœΒΡΖΫ Ϋœό÷ΤΕύκΡΘ§ ΙΤδΈ§≥÷Έ»Ε®ΒΡΓΔΨΏ”–ΗϋΚΟ…ζΈοΜν–‘ΒΡΧΊ βΫαΙΙΓΘ»ΟΕύκΡΡΘΡβΤδ‘ΎΧλ»ΜΒΑΑΉ÷ ÷–Έ»Ε®ΒΡΙΙœσΘ§”––ßΒΊΧαΗΏΤδΈ»Ε®–‘ΚΆΩΙΦτ«–ΡήΝΠΓΘΆ§ ±Θ§’β÷÷ΡΘΡβΩ…“‘ΉΣΜ·≥…ΕύκΡ“©Έο«ΑΧε–¬≥¬¥ζ–ΜΒΡΈ»Ε®–‘ΓΔ Ε±πΉ®“Μ–‘“‘ΦΑ…ζΈοΜν–‘Θ§»ΟΕύκΡ’Ι Ψ≥ωΗϋΚΟΒΡ…ζΈο―ßΙΠΡήΘ§≤Δ±ΘΝτΤδ”≈ΝΦΒΡ…ζΈοœύ»ί–‘ΚΆΑ≤»Ϊ–‘ΘέΘ≤ΘΒΘίΓΘ¥σΝΩΒΡ±®Βά±μΟςΘ§Ϋπ τάκΉ”ΒΡρϋΚœΩ…“‘“ΐΖΔΜρ”ΑœλΕύκΡΒΡ’έΒΰΘ§’βάοΫΪ÷ς“ΣΫι…ή“Μ–©άϊ”Ο≈δΈΜΜ·ΚœΈο ΙΕύκΡΜώΒΟΙΧΕ®ΒΡΙΙœσΚΆ”≈Μ·ΒΡ…ζΈοΙΠΡήΒΡΨΏΧε…ηΦΤΖΫΑΗΓΘ
Θ≥. Θ± ΕύκΡΫαΙΙΒΡΈ»Ε®
’κΕ‘ Θ–Θ–Θ…Θσ ΒΡΖ÷Έω±μΟςΘ§–μΕύ¥Π”ΎΙΊΦϋΫΜΜΞΫγΟφΒΡΕύκΡΈΣΜΖΉ¥Θ®ΘλΘοΘοΘπΘ©ΫαΙΙΘέΘ≤ΘΕΘίΓΘΕύκΡΒΡΜΖΜ·Θ§ «Ε‘Χλ»ΜΒΑΑΉ÷ ΙΠΡήΫαΙΙΒΡΡΘΡβΘ§Ω…“‘»ΟΕύκΡΨΏ”–‘ΎΒΑΑΉ÷ ÷–Ος»ΖΒΡΙΙœσΓΘΡ≥–©ΨΏ”–Α–œρ Ε±πΙΠΡήΒΡΕΧκΡ–ρΝ–Θ§ΫΪΤδ ΉΈ≤Ν§Ϋ”–Έ≥…ΜΖΉ¥ΫαΙΙΘ§Ω…“‘”––ßΒΊΧαΗΏΤδΫαΙΙΒΡΈ»Ε®–‘ΚΆ ήΧεΒΡ«ΉΚœΝΠΘ§’Ι Ψ≥ωΗϋ”≈‘ΫΒΡ…ζΈοΜν–‘ΓΘάΐ»γ’ϊΚœΥΊΑ–œρΕΧκΡ–ρΝ– Θ“Θ«ΘΡΘέΘ≤ΘΖΘίΓΔΘΈ⁃ΗΤ’≥ ΒΑ ΑΉ Α– œρ κΡ Θ»ΘΝΘ÷ΘέΘ≤ΘΗΘίΓΔ ΚΎ …Ϊ ΥΊ Νω Α– œρ κΡ ΠΝ⁃ΘΆΘ”Θ»ΘέΘ≤ΘΙ ΓΪ Θ≥Θ±ΘίΓΔΘψΘοΘνΘοΘτΘοΘχΘιΘνΒ»ΓΘ
≥ΐΝΥ¥ΪΆ≥ΒΡΆ®ΙΐΕΰΝρΦϋΜρ ΉΈ≤θΘΑΖΥθΚœΒΡΖΫΖ®»ΟΕύκΡ≥…ΜΖ÷°ΆβΘ§άϊ”ΟΫπ τ≈δΈΜΜ·ΚœΈο”κΕύκΡœύΜΞΉς”Οά¥ΙΙΫ®ΜΖΜ·ΫαΙΙΒΡΖΫΖ®Θ§ΈΣΕύκΡΒΡΜΖΜ·…ηΦΤΉΔ»κΝΥ–¬ΒΡΥΦ¬ΖΓΘάϊ”ΟΫπ τοΣΘ® ΘρΘηΘεΘνΘιΘθΘμΘ§ Θ“ΘεΘ© ΚΆοΫΘ®ΘτΘεΘψΘηΘνΘεΘτΘιΘθΘμΘ§ Θ‘ΘψΘ©≈δΈΜΜ·ΚœΈο”κΑκκΉΑ±Υα≈δΈΜΘ§―–ΨΩ»Υ‘±≥…ΙΠΒΊ Ι ΠΝ ¥ΌΚΎΦΛΥΊκΡΘ®ΠΝ⁃ΘΆΘ”Θ»Θ©–Έ≥…Έ»Ε®ΒΡΜΖΉ¥ΫαΙΙΘ§”––ßΧα…ΐΝΥΤδΑ–œρΙΠΡήΘέΘ≤ΘΙΘίΓΘΜυ”Ύ”–ΜζΜΖΫπ τ Θ…ΘρΘ®ΔσΘ© ≈δΈΜΜ·ΚœΈο”κΉιΑ±ΥαΒΡ≈δΈΜΖΫΑΗΘ§“≤±Μ≥…ΙΠ”Ο”Ύ Θ»Θ“Θ«ΘΡΘ» ΕύκΡΒΡΜΖΜ·Θ§”––ßΧα…ΐΝΥΑ–œρΕύκΡΒΡ―Γ‘ώ–‘ΘέΘ≥Θ≥ΘίΓΘ
¬ Β…œΘ§Έό¬έ «ΜΖΜ·Θ§“ύΜρΤδΥϊΩ…“‘‘Φ χΕύκΡΫαΙΙΒΡ…ηΦΤΘ®Φθ–ΓΕύκΡΒΡ»α–‘Θ©Θ§ΕΦΩ…“‘Ά®ΙΐΕύκΡΫαΙΙΒΡΙΧΕ®Θ§ά¥ΧαΗΏΤδ”κΡΩ±ξΖ÷Ή”œύΜΞΉς”ΟΒΡΡήΝΠΘ®«ΉΚΆΝΠΘ©ΚΆΉ®“Μ–‘ΓΘάϊ”Ο≈δΈΜΜ·ΚœΈο”κΕύκΡΒΡΧΊ βΈΜΒψ≈δΈΜΫαΚœΘ§Ω…“‘”––ß«ΐΕ·ΕύκΡ’έΒΰ–Έ≥…ΧΊ βΒΡΕΰΦΕΫαΙΙΘ§ΗϋΚΟΒΊ”κΒΉΈοΫαΚœΘ§”––ßΧαΗΏΫαΚœΒΡ«ΉΚœΝΠΓΘ―–ΨΩ»Υ‘±“‘ Θ“ΘθΘ®ΔρΘ©ΚΆ Θ–ΘδΘ®ΔρΘ©≈δΈΜΜ·ΚœΈο”κΕύκΡ÷–ΧΊΕ®ΈΜΒψΒΡΉιΑ±ΥαΜρΑκκΉΑ±ΥαΫχ––≈δΈΜΘ§ΖΔœ÷ΕύκΡΒΡ ΠΝ ¬ί–ΐΒΟΒΫΝΥΟςœ‘ΒΡΧα…ΐΘέΘ±Θ¥ΘίΓΘΘ“ΘθΘ®ΔσΘ©≈δΈΜΜ·ΚœΈοΆ§―υ“≤Ω…“‘Ά®Ιΐ”κΕύκΡ≈δΈΜά¥Η≥”ηΕύκΡΗΏΈ»Ε®–‘ΒΡ ΠΝ ¬ί–ΐΫαΙΙΘ§¥”Εχ”Π”Ο”Ύ…ζΈοΜν–‘ΕύκΡΒΡ…ηΦΤΘέΘ≥Θ¥ΘίΓΘΕχ Θ”ΘψΘηΘνΘεΘιΘδΘεΘρ ―– ΨΩ Ά≈ Ε” ‘ρ ≥… ΙΠ ΒΡ άϊ ”Ο ΘΝΘσΘ®ΔσΘ©”κΕύκΡ≤ΜΆ§ΈΜΒψΒΡΑκκΉΑ±ΥαΖΔ…ζ≈δΈΜ¥”Εχ«ΐΕ·ΜρΗ…»≈ΕύκΡ–Έ≥… ΠΝ ¬ί–ΐΫαΙΙΘέΘ≥ΘΒΘίΚΆ Π¬ ΖΔΦ–ΫαΙΙΘέΘ≥ΘΕΘίΘ®ΆΦ Θ≥Θ©ΓΘΘΟΘθΘ®ΔρΘ©“≤±Μ±®ΒάΩ…“‘Ά®Ιΐ≈δΈΜ ΙΨΏ”–ΒμΖέΟΗ“÷÷ΤΉς”ΟΒΡΥΡκΡ–ρΝ––Έ≥…”κΒΉΈοΨΏ”–ΗϋΗΏ«ΉΚœΝΠΒΡ Π¬ ΉΣΫ«ΫαΙΙΘ§”––ßΧαΗΏΝΥΤδ“÷÷Τ–ßΙϊΘέΘ≥ΘΖΘίΓΘ
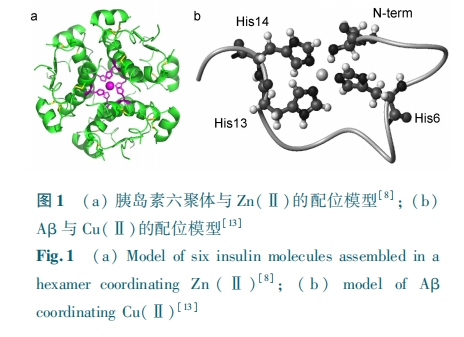
Θ≥.Θ≤ ΕύκΡΒΡΫχ“Μ≤ΫΉιΉΑ
Ϋπ τ‘ΣΥΊ”κΕύκΡΒΡΕύΦέΜ·≈δΈΜΫαΚœΡΘ ΫΨωΕ®ΝΥΤδ≤ΜΒΪΩ…“‘‘ΎΖ÷Ή”ΡΎΧαΙ©ΕύκΡ’έΒΰ≥…ΨΏ”–ΧΊ βΕΰΦΕΫαΙΙΘ§ΜΙΩ…“‘ΫιΒΦΖ÷Ή”ΦδΒΡœύΜΞΉς”ΟΘ§ΙΙΫ®≥ωάμœκΒΡ»ΐΦΕΜρΥΡΦΕΫαΙΙΘ§”–ΆϊΫχ“Μ≤Ϋ”Ο”Ύ≥§Ζ÷Ή”ΙΠΡήΕύκΡΘ·ΒΑΑΉ÷ ΦήΙΙΉ‘ΉιΉΑΒΡ…ηΦΤΚΆΒςΩΊΓΘ
Ε‘”Ύ»ΥΙΛ…ηΦΤΒΡΙΠΡήΒΑΑΉ÷ ΫαΙΙΕχ―‘Θ§Ήν÷’…ζΈοΜν–‘ΒΡ”–Έό”κΤδ’έΒΰΚΆΉιΉΑΒΡΖΫ ΫΟή«–œύΙΊΓΘ”…”ΎΫαΙΙœύΕ‘ΦρΒΞΘ§ΉιΉΑΫαΙϊ“ΉΆ®ΙΐΑ±ΜυΥα–ρΝ–Ής≥ω‘ΛΙάΚΆΒς’ϊΓΘΠΝ ¬ί–ΐΨμ«ζκΡ «“Μάύ”Π”ΟΉνΈΣΙψΖΚΒΡΉΑΡΘ–ΆΓΘΕχ«ΐΕ·’β÷÷ΕύκΡΖ÷Ή”ΦδΉιΉΑΒΡΘ§±ψ «”…≈δΈΜΜ·ΚœΈο”κΧΊΕ®ΈΜΒψΒΡΑ±ΜυΥα≤–Μυ÷°ΦδΒΡ≈δΈΜΘ§άΐ»γ ΘΝΘσΘ®ΔσΘ©⁃ΘΟΘυΘσΘ®ΆΦ Θ¥ΘαΘ©Θ§ΘΎΘνΘ®ΔρΘ©⁃Θ»ΘιΘσΘ®ΆΦ Θ¥ ΘβΘ©Θ§Θ»ΘγΘ®ΔρΘ©⁃ΘΟΘυΘσΘ®ΆΦ Θ¥ΘδΘ©Β»–Έ≥…ΒΡ ΠΝ ¬ί–ΐ»ΐΨέΧεΫαΙΙΘ§“‘ΦΑ”… ΘΟΘαΘ®ΔρΘ© ”κ Θ«ΘλΘθ ΚΆ Θ«ΘλΘν ≈δΈΜ«ΐΕ·ΒΡ ΠΝ ¬ί–ΐΥΡΨέΧεΫαΙΙΒΡ–Έ≥…ΘέΘ≤ΘίΘ®ΆΦ Θ¥ΘψΘ©ΓΘ÷±Ϋ”‘ΎΒΑΑΉ÷ …œ–ό ΈΖ«Χλ»ΜΒΡ Θ≤Θ§Θ≤Γδ⁃ΘβΘιΘπΘυΘρΘιΘδΘιΘνΘε Θ® Θ¬ΘιΘπΘυΘ©Θ§ Θ±Θ§Θ±ΘΑ⁃ΘπΘηΘε⁃ΘνΘαΘνΘτΘηΘρΘοΘλΘιΘνΘεΘ®Θ–ΘηΘεΘνΘ©Β»ΜυΆ≈Θ§Ω…“‘Ηϋ÷±Ϋ”ΒΊΈΣ≈δΈΜΜ·ΚœΈοΒΡΫαΚœΧαΙ©ΈΜΒψΘ§ΫΪΙΠΡήΒΑΑΉ÷ ΉςΈΣΒΞ‘Σ”Ο”ΎΗϋΗ¥‘”ΒΡΫαΙΙΉιΉΑΘέΘ≥ΘΗΘίΓΘ
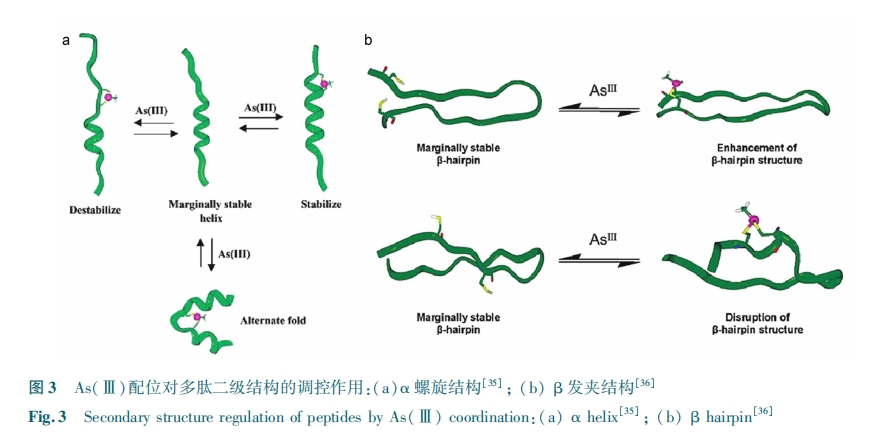
Ά®Ιΐ‘ΎΕύκΡΫαΙΙ…œΫχ––≈δΈΜΑ±ΜυΥαΒΡΕύΦέΈΜ…ηΦΤΘ§Ω…“‘Ϋχ“Μ≤Ϋάϊ”ΟΫπ τ≈δΈΜΘ§ Βœ÷Ε‘ΕύκΡΉ‘ΉιΉΑΒΡ”’ΒΦ”κΫαΙΙ«ΩΜ·ΓΘάΐ»γΘ§άϊ”Ο ΘΎΘνΘ®ΔρΘ© ”κΉιΑ±Υα≈δΈΜΘ§ Β œ÷ Π¬ Βμ Ζέ ―υ Εύ κΡ Ήι ΉΑ ΒΡ ΩΊ ÷ΤΘέΘ≥ΘΙΘίΘΜάϊ ”ΟΘ–ΘβΘ®ΔρΘ©Θ§ ΘΎΘνΘ®ΔρΘ©Θ§ ΘΟΘδΘ®ΔρΘ©Θ§ Θ»ΘγΘ®ΔρΘ©”κ Π¬ ’έΒΰΕύκΡΡΎ…ηΦΤΒΡΑκκΉΑ±ΥαΒΡ≈δΈΜΘ§Ω…“‘Ά§ ± Βœ÷ Π¬ ’έΒΰΫαΙΙΒΡΈ»Ε®“‘ΦΑΕύκΡΥ°ΡΐΫΚΒΡ–Έ≥…ΘέΘ¥ΘΑΘίΘΜΆ®Ιΐ‘Ύ ΠΝ ¬ί–ΐκΡΒΡ ΘιΘ§ Θι ΘΪ Θ¥Θ§ Θι ΘΪ ΘΖΘ§ Θι ΘΪ Θ±Θ± ΈΜ÷Ο…œ“ΐ»κτ«ΜυΙ»Α±ΥαΘ§Ω…“‘ ΙΤδ‘Ύ ΘΟΘαΘ®ΔρΘ©άκΉ”¥φ‘ΎΒΡ«ιΩωœ¬Θ§≤ζ…ζ¬ί–ΐΘ§≤ΔΉ‘ΉιΉΑ–Έ≥…ΤΫ––ΜρΖ¥ΤΫ––ΒΡΫαΙΙΉιΚœΘέΘ¥Θ±Θ§Θ¥Θ≤ΘίΓΘΫπ τ‘ΣΥΊΆ®Ιΐ≈δΈΜΖ¥”ΠΩΊ÷ΤΕύκΡ≤ΡΝœΒΡΉιΉΑΚΆΙΠΡήΗ¥ΚœΜ·ΫΪ”–Άϊ”Π”Ο”ΎΉι÷·–όΗ¥”κ‘Ό…ζΒ»“Ϋ―ßΝλ”ρΓΘ
Θ¥ Μυ”Ύ≈δΈΜΉς”ΟΒΡΕύκΡ”ΪΙβ±ξΦ«‘Ύ…ζΈο―ß÷–ΒΡ”Π”Ο
±®ΒάΖΔœ÷”–“Μ–©≈δΈΜΜ·ΚœΈο”κΑ±ΜυΥα≤–ΜυΖΔ…ζ≈δΈΜΚσΘ§≈δΈΜ≤ζΈοΩ…≤ζ…ζ”ΪΙβ–≈Κ≈ΓΘ’βάύΜυ”Ύ≈δΈΜΡΘ ΫΒΡΕύκΡ”ΪΙβ±ξΦ«ΖΫΖ®Θ§ΨΏ”–ΒΆ±≥ΨΑΓΔΧΊ“λ–‘ΗΏΓΔΖ¥”Π–ß¬ ΗΏΒ»”≈ ΤΓΘ¥ΥΆβΘ§≈δΈΜ≤ζΈοΒΡ”ΪΙβ–≈Κ≈≤ΜΒΪΈΣ≈δΈΜΖ¥”ΠΒΡœύΙΊΕ·ΝΠ―ß±μ’ςΙΛΉςΧαΙ©ΝΥ±ψάϊΘ§‘ΎΕύκΡΒΡ±ξΦ«ΚΆΑϊΡΎ ΨΉΌΘ§“‘ΦΑΒΑΑΉ÷ ’έΒΰΚΆœύΜΞΉς”ΟΒ»―–ΨΩΝλ”ρ“≤ΕΦ’Ιœ÷≥ωΝΥΝΦΚΟΒΡ”Π”Ο«ΑΨΑΓΘ
άΐ»γΘ§«°”άΫΓΒ»άϊ”Ο÷§»ή–‘ ΘΝΘσΘ®ΔσΘ© ≈δΈΜΜ·ΚœΈο”κΒΑΑΉ÷ ΝΎΫϋΈΜΒψΒΡ Θ¥ ΗωœΏ–‘≈≈Ν–ΒΡΑκκΉΑ±Υα≈δΈΜΚσ≤ζ…ζΒΡ”ΪΙβ–≈Κ≈Θ§ Βœ÷ΝΥΜνœΗΑϊΡΎΒΡΒΑΑΉΕ®Βψ±ξΦ«ΘέΘ¥Θ≥ΘίΓΘΕΧκΡΡΎΑϋΚ§ΒΡ Θ¥ Ηω ΘΟΘυΘσ Ω…“‘”κΥΪ…ι»ΨΝœΘΤΘλΘΝΘσΘ»⁃Θ≈ΘΡΘ‘Θ≤ Μρ Θ“ΘεΘΝΘσΘ»⁃Θ≈ΘΡΘ‘Θ≤ ΫαΚœΘ§ΖΔ≥ω¬Χ…ΪΜρΚλ…ΪΒΡ”ΪΙβΓΘ―–ΨΩ»Υ‘±άϊ”Ο¥χ”–ΥΡΑκκΉΑ±ΥαΕΧκΡΘ® Θ‘ΘΟΘ©±ξ«©ΒΡΝ§Ϋ”ΒΑΑΉ ΘΟΘοΘνΘνΘεΘχΘιΘνΘ¥Θ≥⁃Θ‘ΘΟΘ§œ»Κσ”Ο ΘΤΘλΘΝΘσΘ» ΚΆΘ“ΘεΘΝΘσΘ» Ϋχ––±ξΦ«Θ§ Βœ÷‘ΎΜνœΗΑϊΡΎΕ‘–¬Ψ…Ν§Ϋ”ΒΑΑΉΒΡΗϋΧφΓΔΖ÷≤ΦΚΆ‘Υ δΒΡ Β ±Ιέ≤βΓΘ¥ΥΆβΘ§Θ“ΘεΘΝΘσΘ» Ε‘ΕΰΑ±ΜυΝΣ±ΫΑΖΘ®ΘΡΘΝΘ¬Θ©ΨΏ”–ΙβΉΣΜ·Ής”ΟΘ§Ψ≠Ιβ―θΜ·Ής”Ο¥ΠάμΚσΩ…‘ΎΒγΨΒœ¬Ε‘œΗΑϊΒΡ≥§ΈΔΫαΙΙΫχ––Ιέ≤λΘέΘ¥Θ¥ΘίΓΘ»γΆΦ ΘΒ ΘΝ Υυ ΨΘ§±ξΦ«ΚσΝΣΫ”ΑΏΩιΒΡΥΪΡ©ΕΥΒΡ…ζ≥…ΒΡ ΘψΘοΘνΘνΘεΘχΘιΘνΘ¥Θ≥⁃Θ‘ΘΟ ≥ œ÷Κλ…ΪΕχ÷–≤ΩΈΣ¬Χ…ΪΘ§ΆΦΘΒΘ¬⁃ΘΟ ΒΡΒγΨΒΆΦ“≤œ‘ ΨΝΥΝ§Ϋ”Άβ≤ύ≤ΩΖ÷’κΕ‘–¬…ζΘψΘοΘνΘνΘεΘχΘιΘνΘ¥Θ≥ ΒΑΑΉΒΡ»Ψ…ΪΓΘΗΟΖΫΖ®ΫαΚœ”ΪΙβœ‘ΈΔΨΒΚΆΒγΨΒΩ…“‘Ε‘ΡΩΒΡΒΑΑΉΫχ––¬ω≥ε ΫΒΡΉΖΉΌ±ξΦ«ΚΆΙέ≤βΘ§ΈΣΒΑΑΉ÷ ÷ήΤΎΒΡΧΫΥςΧαΙ©ΝΥ–¬–ΆΚΆ÷±ΙέΒΡ±μ’ς ÷ΕΈΓΘ
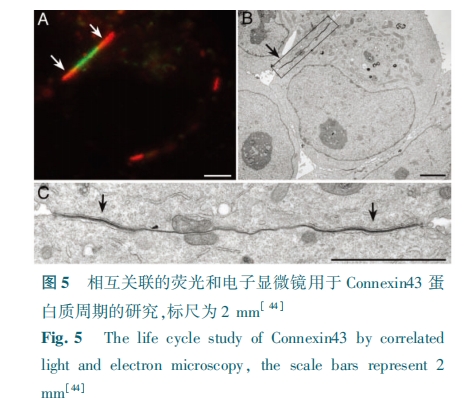
ΫϋΡξά¥Θ§ΗΟ≈δΈΜΖΫΖ®ΙψΖΚ”Π”Ο”ΎΒΑΑΉ÷ ΜρΕύκΡΖ÷Ή”ΡΎΒΡ’έΒΰΚΆΖ÷Ή”ΦδΒΡœύΜΞΉς”ΟΘ®ΕΰΨέΜ·Θ§ΙΙœσΩΣ±’Β»Θ©ΒΡΧΫ≤βΘ§Ϋ®ΝΔΝΥ“ΜœΒΝ–Μυ”Ύ ΘΝΘσΘ®ΔσΘ©⁃ΥΡΑκκΉΑ±ΥαΕΧκΡ±ξ«©ΙΙΫ®ΒΡΒΑΑΉ÷ ΫαΙΙ”κΙΠΡήΒΡΦλ≤βΤΫΧ®ΘέΘ¥ΘΒ ΓΪ Θ¥ΘΗΘίΘ®ΆΦ ΘΕΘ©ΓΘΡΩ«ΑΘ§ΘΝΘσΘ®ΔσΘ©⁃ΥΡΑκκΉΑ±ΥαΫαΚœΡΘ–ΆΉςΈΣΒΑΑΉ÷ ΒΡΖ÷Ή”±ξΦ« ÷ΕΈΘ§ΨΏ”–Ω’Φδ’ΦΈΜ–ΓΘ§≤Μ”ΑœλΒΑΑΉ÷ ±Ψ…μ––ΈΣΒΡ”≈ ΤΓΘ’β÷÷≈δΈΜΚσ≤ζ…ζ”ΪΙβ–≈Κ≈ΒΡ±ξΦ«ΖΫΖ®ΫαΚœ»ΪΡΎΖ¥…δ”ΪΙβΒΞΖ÷Ή”≥…œώΦΦ θΘ®Θ‘Θ…Θ“ΘΤΘ©Θ§Ω…“‘‘ΎΒΑΑΉ÷ ΒΡΒΞΖ÷Ή”Υ°ΤΫΫχ––Ω… ”Μ·ΒΡΕ®ΝΩ±μ’ςΘέΘ¥ΘΙΘίΘ§“≤Ω…άϊ”Ο”ΪΙβΡήΝΩΙ≤’ώΉΣ“ΤΘ® ΘΤΘ“Θ≈Θ‘Θ©Φλ≤βΒΑΑΉ÷ œύΜΞΉς”ΟΘέΘΒΘΑΘ§ΘΒΘ±ΘίΘ§≤Δ”–―–ΨΩΫΪΗΟ±ξΦ«≥…œώΖΫΖ®”Ο”ΎΜνœΗΑϊΒΡ Θ–ΘΝΘΧΘΆ ≥§ΗΏΖ÷±φ¬ Ιβ―ß≥…œώΘέΘΒΘ≤ΘίΓΘ
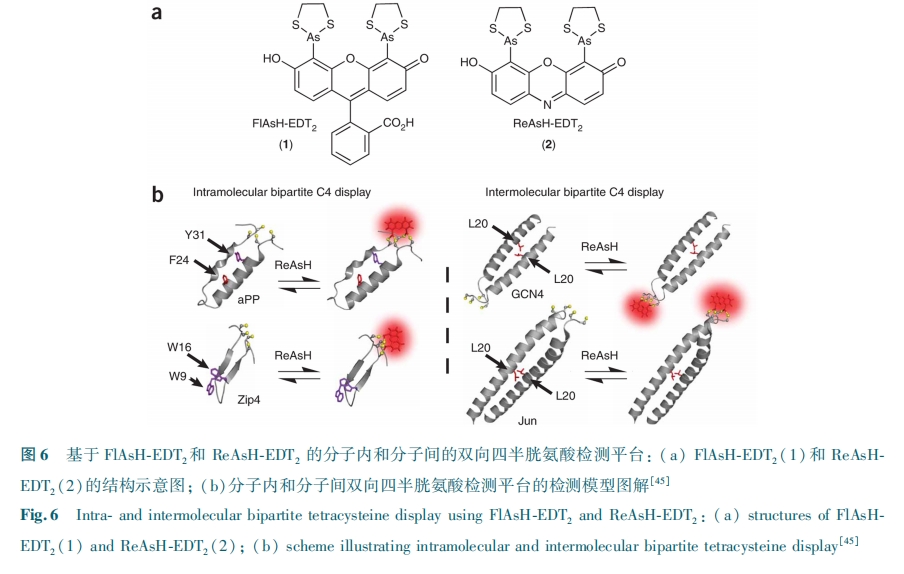
Νμ“Μάύ≈δΈΜΖ÷Ή”…φΦΑΫπ τ”–Μζ≈δΈΜΜ·ΚœΈοΘ§“ρΤδΉςΈΣΝΉΙβ»ΨΝœΨΏ”–ΝΩΉ”–߬ ΗΏΘ§ΥΙΆ–ΩΥΥΙΈΜ“Τ¥σΘ§≤Μ“ΉΙβΤ·ΑΉΒ»ΧΊΒψΘ§ΫϋΡξά¥‘Ύ…ζΈο―ß≥…œώΒ»Νλ”ρΒΡ”Π”Ο ήΒΫ÷Ί ”ΘέΘΒΘ≥ ΓΪ ΘΒΘΒΘίΓΘ―–ΨΩ±μΟςΘ§ΙΐΕ…Ϋπ τ“ΩΘ®ΔσΘ© ≈δΚœΈο“≤Ω…“‘Ά®Ιΐ”κΕύκΡΧΊ βΈΜΒψΒΡΉιΑ±Υα≈δΈΜΘ§≤ζ…ζ¬Χ…ΪΝΉΙβΓΘ―–ΨΩ»Υ‘±ΖΔœ÷ΙΐΕ…Ϋπ τ“ΩΘ®ΔσΘ© ≈δΚœΈοΘ® ΘβΘιΘσΘ®Θ≤⁃ΘπΘηΘεΘνΘυΘλΘπΘυΘρΘιΘδΘιΘνΘε⁃ΘΟΘ≤Θ§ ΘΈΘ©⁃ΘβΘιΘσ Θ® ΘαΘώΘθΘοΘ© ΘιΘρΘιΘδΘιΘθΘμ Θ®ΔσΘ©Ω…”κΕύκΡ–ρΝ– ΘΈ Ρ©ΕΥΒΡ“ΜΗωΉιΑ± Υα Θ® ΘμΘοΘδΘεΘλ Θ± Θ© Μρ –ρ Ν– ÷– Φδ ΒΡ ΝΫ Ηω Ήι Α± ΥαΘ®ΘμΘοΘδΘεΘλ Θ≤ Θ© Ά® Ιΐ ≈δ ΈΜ Φϋ Ϋα Κœ ≤Δ ≤ζ …ζ ¬Χ …Ϊ ΝΉ ΙβΓΘΘ…ΘρΘ®ΔσΘ©Ϋπ τ≈δΚœΈοΒΡΜ·―ßΫαΙΙ ΫΦΑΤδ”κΉιΑ±ΥαΒΡ≈δΈΜΡΘ Ϋ»γΆΦ ΘΖ Υυ ΨΓΘάϊ”Ο≤ζΈοΒΡ¬Χ…ΪΝΉΙβ–≈Κ≈Θ§ΗΟΙΐΕ…Ϋπ τ Θ…ΘρΘ®ΔσΘ© ≈δΚœΈοΩ…“‘±ξΦ«¥χ”–ΉιΑ±ΥαΒΡ¥©ΡΛκΡΘ§”Ο”ΎΜνœΗΑϊ≥…œώΘέΘΒΘΕΘίΘ®ΆΦ ΘΗΘ©ΓΘ
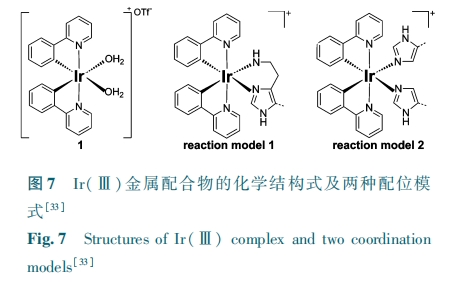
’β÷÷ Θ…ΘρΘ®ΔσΘ© ≈δΚœΈο”κΉιΑ±Υα÷°ΦδΒΡΒΎΕΰ÷÷≈δΈΜΡΘ–Ά±ΜΫχ“Μ≤Ϋ”Π”Ο”ΎΕύκΡΫαΙΙ…œΒΡΈ»Ε®ΜΖΜ·Θ§Ά§ ±Η≥”ηΕύκΡΗ¥ΚœΈοΝΦΚΟΒΡΕύ÷Ί…ζΈο―ß–ß”ΠΑϋά®”≈Μ·ΒΡ»κΑϊ–߬ ΓΔΑ–œρ–‘“‘ΦΑ”κΕΨ–‘ΕύκΡ ΘΥΘΧΘΝ ώνΝΣΚσ’Ι Ψ≥ωΒΡ―Γ‘ώ–‘ΩΙΑ©Μν–‘Β»ΘέΘ≥Θ≥ΘίΓΘ
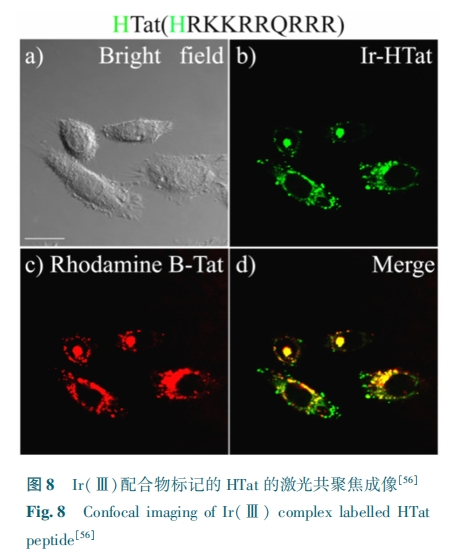
Ήœ»Θ§ΉςΈΣ“Μ÷÷–¬–ΆΒΡΝΉΙβΕύκΡΜΖΜ·ΖΫΖ®Θ§Τδ≈δΈΜΙΐ≥ΧΨΏ”–ΗΏ–ßΓΔ±ψΫίΓΔΧθΦΰΈ¬ΚΆΒ»ΧΊΒψΓΘΕύκΡΥυ≥…ΜΖΒΡ¥σ–ΓΕ‘≈δΈΜΖ¥”ΠΒΡ«ΉΚœΝΠ¥φ‘Ύ“ΜΕ®ΒΡ”ΑœλΘ§Β±ΕύκΡ–ρΝ–÷–ΝΫΗω Θ»ΘιΘσ ÷°ΦδΦδΗτ Θ≤ ΓΪ Θ≥ ΗωΑ±ΜυΥα ±Θ§«ΉΚœΝΠΉνΗΏΩ…¥ο Θ≤ ΘΙΘ≥ ΠΧΘΆΓΘΤδ¥ΈΘ§ΗΟΜΖΜ·ΕύκΡ≤ζΈο‘Ύ…ζάμΧθΦΰœ¬Έ»Ε®–‘ΝΦΚΟΘ§ΡήΒ÷ΩΙ―Σ“Κ÷–Ά§≈®Ε»ΒΡΘΎΘνΘ®ΔρΘ©ΓΔΘΟΘθΘ®ΔρΘ©ΓΔΘΟΘαΘ®ΔρΘ©ΓΔΘΆΘγΘ®ΔρΘ© Β»ΕΰΦέ―τάκΉ”Θ§ΑΉΒΑΑΉ“‘ΦΑœΗΑϊΡΎΗΏ≈®Ε»ΒΡΙ»κΉΗ κΡΒ»ΒΡΨΚ’υΓΘΜυ”ΎΫπ τ Θ…ΘρΘ®ΔσΘ© ≈δΚœΈο”κΉιΑ±ΥαΒΡΕύκΡΜΖΜ·≤ΜΒΪ‘ΎΫαΙΙ…œΗ≥”ηΝΥΕύκΡΝΦΚΟΒΡΈ»Ε®–‘Θ§‘ΎΙΠΡή…œ“≤ΈΣΜΖΜ·Εύ κΡ ΙΠ Ρή Η¥ Κœ Έο ¥χ ά¥ ΝΥ œ‘ ÷χ ΒΡ ”≈ Μ·ΓΘΕύ κΡΘ»Θ“Θ«ΘΡΘ» ”κ Θ…ΘρΘ®ΔσΘ© ≈δΚœΈο≈δΈΜΚσΘ§Χεœ÷≥ωΝΥΜΖΜ·ΕύκΡ”≈”ΎΤδœΏ–‘ΫαΙΙΒΡ”κΑ–œΗΑϊπΛΗΫΒΡ«ΉΚœΝΠΚΆΑ–œρ―Γ‘ώ–‘ΓΘ’ϊΚœΥΊ ΠΝΠΆ Π¬Θ≥ ΗΏ±μ¥οΒΡ ΘΝΘΒΘ¥ΘΙ œΗΑϊΕ‘ΜΖΜ·ΕύκΡ Θ…ΘρΘ®ΔσΘ©⁃Θ»Θ“Θ«ΘΡΘ» ΒΡ…ψ»ΓΡήΝΠΟςœ‘”≈”ΎœΏ–‘ΕύκΡ Θ“Θ«ΘΡΘ»Θ»⁃Θ…ΘρΘ®ΔσΘ©Θ§Ά§ ±’ϊΚœΥΊ ΠΝΠΆΠ¬Θ≥ ±μ¥οΝΩΫœΒΆΒΡΘΆΘΟΘΤ⁃ΘΖ œΗΑϊΕ‘ Θ…ΘρΘ®ΔσΘ©⁃Θ»Θ“Θ«ΘΡΘ» ΒΡ…ψ»ΓΡήΝΠ“≤Ϋœ»θΘ§Χεœ÷≥ωΝΥΗυΨί’ϊΚœΥΊ ΠΝΠΆΠ¬Θ≥ ±μ¥οΝΩ≤ΜΆ§ΒΡ»κΑϊΡήΝΠΒΡ―Γ‘ώ–‘ΓΘΆ§ ±Θ§ Θ…Θρ Θ®ΔσΘ© ≈δΚœΈοΒΡ«Ή÷§–‘“Σ”≈”ΎΘΤΘ…Θ‘ΘΟ ΚΆ ΘΝΘλΘεΘχΘα ΘΤΘλΘθΘοΘρ Β»≥Θ”ΟΒΡ”–Μζ”ΪΙβ»ΨΝœΘ§Ω…“‘ΈΣ’ϊΗωΕύκΡΗ¥ΚœΈο¥χά¥ΗϋΚΟΒΡœΗΑϊΆ®ΆΗΡήΝΠΓΘΒ±»κΑϊΡήΝΠΫœ≤νΒΡ―τάκΉ”œΗΑϊΕΨ–‘ΕύκΡ ΘΥΘΧΘΝ ”κ Θ…ΘρΘ®ΔσΘ©⁃Θ»Θ“Θ«ΘΡΘ» œύΝ§ΚσΘ§ΤδœΗΑϊΕΨ–‘ΒΟΒΫΝΥœ‘÷χΒΡΧα…ΐΓΘ”κ¥ΥΆ§ ±Θ§ΫαΚœΜΖΜ· Θ“Θ«ΘΡ ΒΡΑ–œρΙΠΡήΘ§’ϊΗωΕύκΡΗ¥ΚœΈοΧεœ÷≥ωΝΥΝΦΚΟΒΡ―Γ‘ώ–‘œΗΑϊ…±…ΥΙΠΡήΓΘ»γΆΦ ΘΙΥυ ΨΘ§ΫΪ–Γ–ΆΥσΉ¥»ΥΖΈ≥…œΥΈ§œΗΑϊ Θ»ΘΧΘΤΘ®±μ¥οΚλ…Ϊ”ΪΙβΒΑΑΉΘ©”κΫœ¥σΒΡΕύ±Ώ–ΈΖ«–ΓœΗΑϊΖΈΑ©œΗΑϊ ΘΝΘΒΘ¥ΘΙΙ≤≈ύ―χΚσΘ§ΕύκΡ Θ…ΘρΘ®ΔσΘ©⁃Θ»Θ“Θ«ΘΡΘ»⁃ΘΥΘΧΘΝΘ®¬Χ…ΪΝΉΙβΘ©ΩλΥΌ”–―Γ‘ώ–‘ΒΊ”Ω»κ ΘΝΘΒΘ¥ΘΙ œΗΑϊΘ§≤Δ‘λ≥…ΝΥœΗΑϊΙΡ≈ίΚΆΚΥ≈®ΥθΒ»œΗΑϊΒρΆωΒΡ±ξ÷Ψ–‘œ÷œσΘ§≥δΖ÷œ‘ Ψ≥ωΟς»ΖΒΡ―Γ‘ώ–‘÷ΉΝωœΗΑϊ…±…ΥΙΠΡήΓΘ
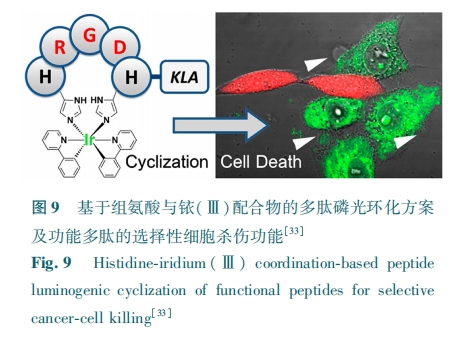
ΉήΒΡά¥ΥΒΘ§’β÷÷–¬–ΆΒΡΝΉΙβΕύκΡΜΖΜ·ΖΫΖ® «ΕύκΡ±ξΦ«ΚΆΫαΙΙ”≈Μ·ΒΡάμœκΫαΚœΓΘ≈δΈΜΖ¥”Π¥χά¥ΒΡΝΉΙβΈΣΙΠΡήΕύκΡΒΡΉΖΉΌΧαΙ©ΝΥ±Ί“ΣΒΡΦύ≤β–≈Κ≈Θ§ΈΣΚσ–χΒΡœΗΑϊ––ΈΣΙέ≤βΚΆΜζάμΖ÷ΈωΧαΙ©ΝΥΦΪ¥σΒΡ±ψάϊΓΘΕχ≈δΈΜΗ≥”ηΕύκΡΜΖΉ¥ΫαΙΙΒΡΆ§ ±ΧαΙ©ΝΥΝΦΚΟΒΡ«Ή÷§–‘Θ§Ε‘ΕύκΡ±Ψ…μΒΡ…ζΈοΙΠΡή“‘ΦΑ»κΑϊΡήΝΠΕΦ”–œ‘÷χΒΡ”≈Μ·–ßΙϊΓΘ
ΘΒ ’ΙΆϊ
≤ΜΆ§ΒΡ≈δΈΜΜ·ΚœΈο”κΕύκΡΒΡΫαΚœΘ§ΨΏ”–ΗςΉ‘ΧΊ βΒΡΑ±ΜυΥαΤΪΚΟΓΘΕχ’β÷÷≈δΈΜΖ¥”ΠΥυΨΏ±ΗΒΡ―Γ‘ώ–‘Θ§»ΟΈ“Ο«Ω…“‘ΗυΨί≤ΜΆ§ΒΡ«ιΩωΚΆ–η«σ…ηΦΤΕύκΡ–ρΝ–Θ§»Ο≈δΈΜΖΔ…ζ‘ΎΈ“Ο«‘ΛΤΎΒΡΈΜΒψΘ§¥”Εχ”Ο”Ύœό÷ΤΕύκΡΒΡΕΰΦΕΫαΙΙΘ§«ΐΕ·ΕύκΡΒΡ’έΒΰΘ§”≈Μ·Τδ…ζΈοΜν–‘Β»ΓΘΥδ»Μ”–¥σΝΩΒΡ±®Βά≤ΜΕœΫ“ ΨΉ≈Ης÷÷Α±ΜυΥα”κ≤ΜΆ§Ϋπ τάκΉ”÷°ΦδΒΡ≈δΈΜΡΘ–ΆΘ§ΒΪ’φ’ΐΫΪ’β÷÷œύΜΞΉς”Ο…ν»κ”Π”ΟΒΫ…ζΈο―ß÷–ΒΡ≥…ΙΠΑΗάΐ≤Δ≤ΜΕύΦϊΓΘ±ΨΈΡΉ≈÷ΊΫι…ήΒΡΝΫάύΨΏ”–¥ζ±μ–‘ΒΡ¥χ”–”ΪΘ· ΝΉΙβΒΡ≈δΈΜΡΘ ΫΘ§Τδ≈δΈΜΖ¥”ΠΜζάμΟς»ΖΘ§―Γ‘ώ–‘«ΩΘ§”»Τδ÷Ί“ΣΒΡ «≈δΈΜ≤ζΈοΥυ≤ζ…ζΒΡ”ΪΘ· ΝΉΙβ–≈Κ≈ΓΘ’βάύ≈δΈΜΧεœΒΨΏ”–ΕάΧΊΒΡ”≈ ΤΘΚΘ®Θ±Θ©”κ¥ΪΆ≥ΒΡάϊ”Ο Θ≈ΘΡΘΟΓΔγζγξθΘ―«ΑΖΒ»Ι≤ΦέΝ§Ϋ””ΪΙβΥΊΖ÷Ή”ΒΡΕύκΡ”ΪΙβ±ξΦ«ΖΫΖ®œύ±»Θ§’β÷÷Μυ”Ύ≈δΈΜΜ·ΚœΈο”κΕύκΡΒΡ≈δΈΜΖ¥”ΠΨΏ”–Ζ¥”ΠΙΐ≥ΧΦρΒΞΩλΥΌΓΔΧθΦΰΈ¬ΚΆΓΔΖ¥”Π–ß¬ ΗΏΓΔ―Γ‘ώ–‘ΗΏΒ»”≈ΒψΘΜΘ®Θ≤Θ© ≈δΈΜΖ¥”ΠΖΔ…ζ”κ”ΪΘ· ΝΉΙβ–≈Κ≈ΒΡ≤ζ…ζ÷±Ϋ”ΙΊΝΣΘ§Ω…“‘”Ο”Ύ±φΈω≈δΈΜΖ¥”ΠΒΡΖΔ…ζΘ§Ά§ ±”––ßΫΒΒΆΕύκΡΒΑΑΉ÷ ±ξΦ«ΒΡ±≥ΨΑ”ΪΙβ–≈Κ≈ΘΜΘ®Θ≥Θ© ≈δΈΜΖ¥”Π“ΜΑψΈΣΕύΦέΒΡ≈δΈΜΡΘ ΫΘ§Φ¥Ά§ ±–η“ΣΕύΗωΧΊ βΈΜΒψΒΡΑ±ΜυΥαΧαΙ©ΟΣΕ®ΈΜΒψΘ§”––ßΒΊ±Θ÷ΛΝΥ±ξΦ«Ζ¥”ΠΒΡΧΊ“λ–‘ΓΘ“ΜΑψά¥ΥΒΘ§≈δΈΜœύΙΊΒΡΈΜΒψΘ®ΦέΧ§Θ©‘ΫΕύΘ®ΗΏΘ©Θ§≈δΈΜΖ¥”ΠΖΔ…ζΒΡΧΊ“λ–‘ΨΆ‘Ϋ«ΩΓΘ‘ΎΜνœΗΑϊΒΡΒΑΑΉ÷ Θ· ΕύκΡ±ξΦ«ΒΡ”Π”Ο÷–Θ§Ά®≥Θ–η“ΣΗϋΗΏΒΡΧΊ“λ–‘Θ§Εχ‘ΎΒΑΑΉ÷ ’έΒΰΚΆœύΜΞΉς”ΟΖ÷ΈωΒΡ”Π”Ο÷–Θ§ΫœΒΆΒΡ≈δΈΜΦέΧ§Ω…“‘¥χά¥ΗϋΤ’±ιΒΡΫαΙΙ ”Ο–‘ΓΘ
¥Υάύ≈δΈΜΜ·ΚœΈο”κΧΊΕ®ΒΡΕύκΡ–ρΝ–≈δΈΜ≤ζ…ζΒΡ”ΪΙβ–≈Κ≈Θ§Ω…“‘”Ο”ΎΒΑΑΉΕ®ΝΩΖ÷ΈωΓΔœΗΑϊ≥…œώ”κΕύκΡάύ“©Έο ΨΉΌΘ§“‘ΦΑΒΑΑΉ÷ ’έΒΰΒΡΦλ≤βΓΔ«ΐ ΙΈ¥±’ΚœΒΡΜΖΉ¥ΕύκΡ≤Έ”κœύΙΊ Θ–Θ–Θ…Θσ “‘ΦΑ’κΕ‘ΡΩ±ξΖ÷Ή”«ΉΚœΝΠΒΡΗΏΆ®ΝΩ…Η―ΓΓΘΈ“Ο«ΤΎΆϊΆ®Ιΐ±ΨΈΡΈΣΜ·―ß”κ…ζΈο―ßΝλ”ρΒΡ―–ΨΩ’ΏΧαΙ©“ΜΗωΕύκΡ”κΫπ τ≈δΈΜ―–ΨΩΒΡΗ≈ ωΘ§ΤΎ¥ΐΈ¥ά¥Μ·―ß…ζΈο―ßΝλ”ρΒΡ―–ΨΩΦΧ–χΖΔ’ΙΗΏ÷ ΝΩΖ÷Ή”ΧΫ’κΘ§»Ο≈δΈΜΜ·―ß‘Ύ…ζΈο―ß÷–’Ι ΨΗϋ¥σΒΡς»ΝΠΚΆ Β”ΟΦέ÷ΒΓΘ
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷ΨΓΘ
