
ժ Ҫ: ϸ���������ܹ���ϸ��Ĥ�������Եذ���ϸ���ڰб꣬�����нϺõ������ȶ��ԣ�����߱��ܺõij�ҩ�ԣ����ܵ��о���Խ��Խ��Ĺ�ע��Ŀǰ����Ҫ�����ַ�ʽ���ϸ�������ģ�������Ȼ�����л�úͶ����еĻ��Ļ������Ľ��л�ѧ���Ρ�������Ҫ��������������м�Ҫ�������ص��������ϸ����������Ȼ�����Լ�ͨ����ѧ���η����õ�ϸ�������ĵIJ��ԣ���̽��ϸ�������ĵĽṹ-���Թ�ϵ����ϸ����������
������ҩ�������˵�����ҩ���з���������ȫ�����Ѿ��������еĶ�����ҩ�ﳬ�� 50 ������С����ҩ��Ƚϣ�1��2�ݣ�������ҩ����н����������ð����Ը�ǿ������ȫ�������ø�С�������������ص����߷�Ӧ���ŵ��3��4�ݣ��Ѿ��㷺Ӧ���ڰ�֢����Ѫ�ܲ�����л��ʹ�Ⱦ�Բ��ȸ��༲�������ơ�
���ǣ������Ķ��ķ��Ӵ�������Խϸ��Ĥ��ø���ȶ��Բ�Ϳڷ��������öȵ͵������5�ݣ����´���������������õĶ���ҩ����ѽ���ϸ���ڷ���ҩЧ�������������ڽ����6�ݣ���ˣ�����ҩ����ʵ��Ӧ�����ܵ��˼�������ơ�Ϊ�˿˷�����ҩ����ڵ����⣬���Ƿ��ֲ���չ��һ��ϸ�������ķ��ӣ����Ǿ��д���ϸ��Ĥ�����������������Եذ���ϸ���ڰб꣬�����ȶ��Ըߣ��ڷ��������öȺã����߳�ҩǰ������һ���棬����ѧ�ϳ�����ȡ�õ�һϵ����Ҫ��չ��7 �� 15�ݣ��ٽ��о��߷�չ��һϵ�л�ѧ�ϳɻ��ĵķ�����16 �� 20�ݣ�Ϊ�о����컷�ĴӶ��õ�ϸ�������ĵ춨�˼�����������Ч�����ϸ�������ķ��ӵĻ�ȡ���⡣
�����ص����Ŀǰ�Ѿ����ֵ�ϸ�������ļ���Ĥ������ͬʱҲ�������ֻ��ϸ�������ĵĻ�ѧ���β��ԣ�������۸�������ڵ����⣬��չ��ϸ�������ĵ�Ӧ��ǰ����
2 ��Ȼ�����е�ϸ��������
��������Ȼ����һ��㷺���ڵ���Ȼ�������ͨ���������Ǻ�����;���ϳɣ����п�������ʹ���������������ߵ��������á����У�ijЩ���ľ��д�ϸ��Ĥ��������������������ϸ�������Ļ�����( cyclosporine A)��ֲ�ﻷ��( cyclotides)��
2. 1 �����ؼ�Ĥ����
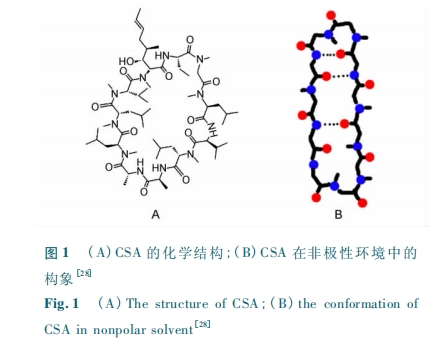
��ʮһ�� cyclosporine A(CSA)������һ��������ֲ������Ŀ���������ҩ�CSA �� 1969 ������� Tolypocladium inflatum �б����֣�ͨ���Ǻ�����;���ϳɣ�21�ݣ�����һ�� D �ͱ������һ������Ȼ�����ᣬ�����ڶ��ĹǼ��Ϻ��� 7 �� N-�����İ�����л�( ͼ 1A)��CSA ���Դ��ܰ�ϸ��Ĥ�����ܰ�ϸ���ڵ�����( cyclophilin) ��ϣ�22�ݣ�ͨ�����ƸƵ�����ø( calcineurin) �Ļ��ԣ�ʹ�ܰ�ϸ���İ�ϸ������ 2 ( IL-2) ���������٣��Ӷ����� T ϸ���ļ�����������ߵ����á��о��߽����� CSA�ľ���ṹ��23��24�ݣ����ö�ά �� �� �� �� �� �� �� CSA�ڼ��ԺͷǼ����ܼ������µĶ�̬�ṹ��24 �� 27�� ( ͼ1B)��ʵ�������CSA �ڷǼ����ܼ��;���ṹ���γ����Է����������ͬʱ�������� 7 �����ĹǼ��ϵ� N ���������������ֹ����� CSA ������������嶼���������ڱΣ����������ڷǼ��Ի����µ��ܽ⣬��������ϸ��Ĥ�еļ���˹�����ܡ�
2. 2 ֲ�ﻷ�ļ�Ĥ����
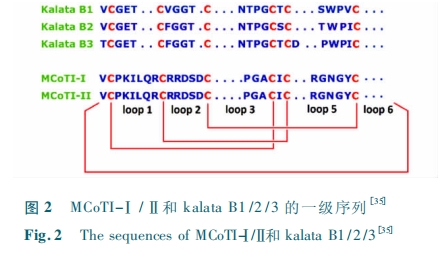
ֲ�ﻷ��Ҳ��һ�������Ҫҩ���ֵ�Ļ��ķ��ӡ�ֲ�ﻷ��ͨ��������;���ϳɣ�������л���һ���� 28 �� 37������Դ��ֲ�����Ȼ�����35�ݡ�Ŀǰ�Ѿ�����ֲ�ﻷ�ľ��кܶ�������ԣ����翹ϸ�������桢������������ø���Ƽ��ȣ�29 �� 31�ݡ�ֲ�ﻷ����Ҫ��Ϊ����: ��Ī����˹�͡�( the M��bius cyclotides)�͡������͡�( the bracelet cyclotides)����������Ϊ�ȵ���ø���Ƽ��ĵ� 3 ���࣬����ֲ�ﻷ�Ľ�����MCoTI-��� MCoTI-�� �������ġ���ͬ���͵�ֲ�ﻷ�Ķ��������ƵĽṹ(ͼ 2)�������� N �˺� C �������������ϣ�6 �����װ����γ� 3 �Զ���������� �Զ�������������Ե��м䴩�����γɻ��װ����( cyclic cystine knot��CCK) �Ľ� ������ �� �� CCK�ṹʹ��ֲ�ﻷ�ĵ����ȶ��ԡ���ѧ�ȶ��Ժ�ø�ȶ��Ժܸߣ����㾭�����Ҳ��Ȼ�߱��ܸߵ�������ԣ�32�ݡ�ֲ�ﻷ�İ��� 6 ������( loop)������ijЩ�����ǿɱ�ģ��ʶ�������ֲ�ﻷ���ϼӾ����������ԵĶ���Ƭ�Σ�����ֲ�ﻷ�������ڵ��ȶ��ԺͽϺõ��������öȣ���߶���Ƭ�������ڵ��ȶ��Ժʹ���ʱ���33�ݡ�
Craik ���״α����˿��Դ���ϸ��Ĥ��ֲ�ﻷ�� MCoTI-�� �� �� �� ͨ �� �� �� �� �� ��(macropinocytosis) �� ϸ �� �� ȡ��34�ݡ��� ��Camarero���ڻ�ϸ��ˮƽ��֤ʵ�� MCoTI-��Ҳ���Դ���ϸ��Ĥ��35�ݡ��� MCoTI-��MCoTI-���⣬Craik �ȷ���ֲ�ﻷ�� kalata B1 �� �� һ �� �� �� �� ֲ �� �� �� �� ��STFI-1 �����Դ���ϸ��Ĥ��36�ݡ�
�о����������ѷ��ֵ�ֲ�ﻷ����Դ�� 4 ��ϸ�������ĵĿ�Ĥ����������ͬ��MCoTI-��ͨ��ϸ��������ȡ�ķ�ʽ����ϸ����ӫ�������ټ���������MCoTI-�����������С���У�����ϸ����ȡ���̶��¶����У�˵�� MCoTI-�� �Ŀ�Ĥ�� �� �� �� �� ���̣���Ҫ����������36�ݡ�MCoTI-��Ŀ�Ĥ�������Ϊ���ӣ�����������������( fluid-phase endocytosis)������֬�ʵ��������ú����鵼���������õȹ�ͬ���õĽ����35�ݡ���ֲ�ﻷ�� kalata B1 ��ϸ�����ԣ�����һ��Ũ��ʱ�ᵼ��ϸ��Ĥ���ѣ�����ϸ���� ����ʵ �� �� �֣�kalata B1 �� �� ֬ �� �� �� ��( POPE ) �� �� �� �� �ã��� �� �� �� POPE �� ��( transbilayer movement)���Ӷ�����ϸ��Ĥ����IJ��Գơ�Craik �Ȳ²� kalata B1 ͨ����ϸ��Ĥ������ã��ı�ϸ��Ĥ�����ʣ��Ӷ�����ϸ�����������ã�36�ݡ���һ���棬kalata B1 ��ϸ��Ĥ��������֬����أ�������ĵ������ϸ��Ĥ�� kalata B1 ����ǿ����õ�ԭ���37�ݡ�
ֲ�ﻷ��ϸ�������̵��о������ڽϳ����ĽΡ�һ���棬��ṹ�ڿ�Ĥ�����е����û�������ȷ�����о�ֲ�ﻷ�ĵĿ�Ĥ����ʱ��ʵ�������IJ���ᵼ�²�ͬ��ʵ����������ʵ����ѡ���ϸ��ϵ�����̷�ʽ��ƫ�ó̶Ȳ�ͬ������ֲ�ﻷ�ĵ�Ũ�Ȳ�ͬ�ȣ�����Խ������Ӱ�졣ͬʱ����ǰֻ��ͨ���ı�л��ķ�ʽ��̽������λ���������Ĥ���̵Ĺ��ף�����ʵʱ���ĵĹ���仯�Կ�Ĥ���̵�Ӱ�졣���⣬�Կ�Ĥ�����е�ϸ��������Ϊ������ʮ�����ޡ��������鵼���������ú�ϸ��Ĥ��������( caveolae) �鵼��������������϶��⣬����ѧ�Ҷ��������͵�ϸ��������Ϊ�����Ⲣ�����롣��Ҳ�谭���о��߶�ֲ�ﻷ�Ŀ�Ĥ���̵Ľ�һ�����⡣��ˣ���չ���ͼ����о�ֲ�ﻷ�Ŀ�Ĥ���ԵĽṹ-���Թ�ϵ����һ���д�����Ŀ�ѧ���⡣
3 �˹������ϸ��������
ϸ�������ľ��кܺõij�ҩǰ�������ʹ�о������û�ѧ�ϳ��ֶζԶ��ķ��ӽ��и��죬���ڻ�ü��ǿ�������ԺͿ�Ĥ�����Ļ��ķ��ӣ��������Ŀ⡣���潫�� CSA �ĸ��졢ֲ�ﻷ�ĵĸ���������Ļ�������������Խ��н��ܡ�
3. 1 ���� CSA �ĸ������
���� CSA ��ϸ�������ĵ����˼·�������ڻ��ĵĹ���Ͷ��ĹǼ����� N-���������档
3. 1. 1 ���Ĺ������Ĥ����
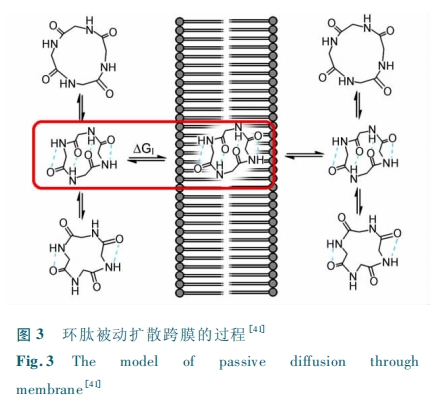
�ܵ� CSA �ڼ��ԺͷǼ����ܼ��й���仯���������о��߿�ʼ��ע���Ĺ�������ӿ�Ĥ����֮��� �� ϵ��38 �� 40�ݡ�Lokey �� ϵ ͳ �� �� �� �� �� �� ��Leu-Leu-Leu-Leu-Pro-Tyr �IJ�ͬ�Ƕ�ӳ�칹��Ĺ���� �� Ĥ �� �� ֮ �� �� �� ϵ��41�� ( ͼ 3 )��ʵ �� ʹ ��CDCl3 ģ��ϸ��Ĥ�������о���������ϸ��Ĥ�еĶ�̬�� ��H /D �� �� ʵ �� �� �� �� �� �� �� �� �� ��N��H �� �� �� �� �� �� �� ¶ �� �ȡ�ʵ �� �� �� �� ����N��H�����ܼ��ɼ���( solvent accessibility) �뻷�Ŀ�Ĥ�������������ԣ��ܼ��ɼ��Խϵ͵ķ��ӿ�Ĥ������ǿ��
Ϊ�˸��Ӷ������о���Ԥ��ı�����ɢ��Ĥ��������Lokey �� Jacobson �������һ���µ�Ԥ�ⷽ����42�ݡ����軷����ϸ��Ĥ�еĹ���(�ͽ�绷������ low-dielectric conformation��LDC) ֻ��һ�֣�����ָ�����������ɢģ�� Pe = KpD / d �е� KpӦ����Ϊ LDC ��ϸ��Ĥ��ˮ���еķ���ϵ����������ͨ��ʵ������õ����й����µ�ƽ��ֵ���������������ͨ�����Ӷ���ѧ����õ� ��GI ( ���ù�ʽ ��GI =�� ��TlnKp )���Ӷ��������ۼ�������ʡ�����ָ�������ģ�͵�Ԥ��������Ԥ�������Ŀ��Ԥ�⼫�Ա��������ȷ���������Ǽ������ ��GI��ʵ���õ� lnPe�����Թ�ϵ��������ϵ��ȴ����һ�£���Ϊ������û�п�����ЧӦ;���⣬���ķ��ӳߴ����״������Ҳû�п������ڣ�����ʵ����֤����С����ģ�͵���ȷ�Ի��д����顣
�Ծ�������ɢ��Ĥ�Ļ��Ķ��ԣ�Ԥ�Ⲣͨ����ѧ���εķ����ı� LDC ���Ӷ���С���Ĵ�ˮ����ϸ��ĤǨ���еļ���˹�����ܱ����о��Ĺؼ��㡣���ԣ���ʶ��ѧ���ζԻ��Ĺ����Ӱ����������Ҫ�Ŀ�ѧ���⡣Ȼ���������еı�����ɢ��Ĥ�Ļ���ʵ���У���л��������Dz�����ɲ����ǷǼ��Եģ�˵�����ֿ�Ĥ���Զ�����л���Ҫ��ϸߣ��Լ��Ժʹ���ɵIJл������̶Ƚϵ͡����������⣬�����ʵ��������̶Ƚϴ�ķ���Ȼ����������߱�����ɢ�����»��ĵĶ�����;��һ���棬��������ϸ������ pH ��ø���ԵIJ��죬��� pH ��Ӧ��ø��Ӧ�л�����ϸ�����伫�Ա��ڱΣ���ϸ����ȥ�ڱΣ�����ĵ������������ԡ�
3. 1. 2 N-�������Ĥ����
���˶��Ĺ���仯�Կ�Ĥ�����й��ף����ĹǼܵ����� N-�����Կ�Ĥ����Ҳ��Ӱ�졣�Ǽ����� N-������������Ȼ�����кܳ��������ض�λ������ N-��������ǿ����������������á���ǿ�� �� �� �� �� �� ѡ �� �� �� �� �� �� �� �� �� �� ��Ϊ��43 �� 45�ݡ�
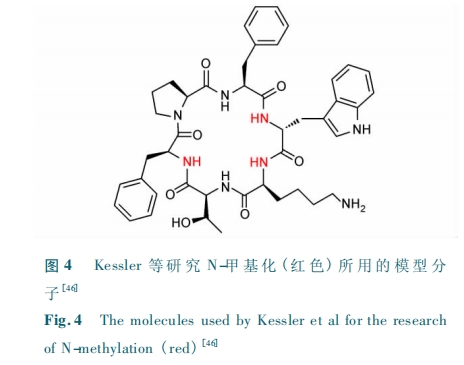
�� �� �� ˼ ·��Kessler �� Ϊ �� �� VeberHirschmann ������( cyclo ( -PFwKTF-) ) �Ŀڷ��������öȣ�ϵͳ���о��� N-���������е����ã�46��(ͼ 4)�����Ƿ��֣����ض�λ������ N-������ �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� �� ��9. 9% ����ԭ���� N-���������˻��ķ��ӵ�֬���ԣ�ʹ������״���ϸ��Ĥ��֮��Kessler ���ֱ����������������Ƽ������� MK678 �� N-��������߿ڷ����ԣ�����ʾ�ý�������� N-������ǿ���Ŀ�Ĥ������ɣ�47�ݡ������÷���ģ��ķ������ֿڷ�������ߵķ�����ˮ���еĹ����� CSA �������˺�Kessler �� Hoffman �ȷ��ּ���Ŀ���Ĥ����֮��û�б�Ȼ��ϵ��48�ݡ�
Jacobson �� Lokey �Ƚ�һ��ϵͳ���о��� N-�����Ի��Ĺ���Ϳ�Ĥ������Ӱ���49�� ( ͼ 5 )�������о��˻����� Leu-Leu-Leu-Leu-Pro-Tyr �IJ�ͬ�Ƕ�ӳ�칹������֬�ϵļ�����Ӧ�����Ƿ��֣�һ�������£�������֬�ϵ����� 6 �����͵Ļ����ĵļ�����Ӧ��������ѡ����(ѡ���Դ��� 95% )���ھ�������ѡ���ԵĻ��Ľ��� H /D ����ʵ��ʱ���֣�δ�����Ļ��ķ��ӵ����� N��H ��¶����Һ��;�����ּ����������� �� δ �� �� N-�� �� �� ��N��H ������¶����Һ�С�����ָ������������ѡ���Կ�����Դ�ڲ�����ӵ����� N��H ��λ��������������ƣ�������������ʵ����������ּ����Ķ��ĵĿ�Ĥ������ǿ������ͨ�����ۼ���ָ��������ѡ���Լ��������������������п�Ĥ������ǿ�ģ�����ڼ��Խϵ͵��ܼ������������еļ�����Ӧ���������ɿ�Ĥ������ǿ�����������һ��������ȴ���õ��������͵����ż���λ�㣬Ҳ����˵���ü��������µ�����ѡ�������䴩ϸ��Ĥ�ij�ֲ���Ҫ����������Ҳ���֣�����1��3��4 λ��������ֱ��Ϊ˿��������Ĥ��������½���˵�� N-��������ֻ������ijЩ������л�������˼���ǣ�����ָ�����ڸ�ʵ���з��ֵļ��� λ �� �� ǰ �� Kessler �� �� �� Veber-Hirschmann���õ��ļ���λ���46��һ�£����Dz²����е� ��-ת��(��-turn)�ṹָ���˿绷������������γɡ�
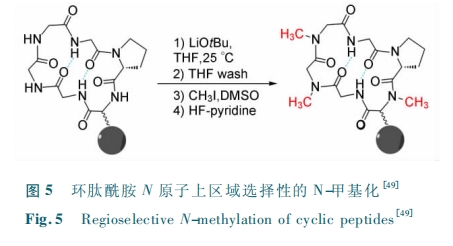
���Ϲ���������N-��������ͨ���ı价����ˮ���еĹ�����������Ĥ��������ǰ�˹����Ļ����ϣ�Kessler ��ϵͳ���о��� N-�����Ի��Ĺ����Ӱ���50�ݡ�����ѡ������ cyclo( -D-Ala-Ala5 -) ��Ϊ�о���������ͬ N-������������ˮ���еĶ�ά�˴Žṹ��������Ϊ N-�����������������Ӱ�컷�� ��-ת�ǽṹ���γɣ��Ӷ����¿绷����������ij���: ( 1 ) �ļ���˳����ʽ; ( 2 ) �ԹǼ���N��H ������ڱ�����;(3 ) Ť������ԭ���Ľṹ�����Ƿ��֣�ʵ�������õ���һЩ���͵� ��-ת�ǽṹ��ǰ�� �� �� �� N-�� �� �� ϸ �� �� �� �� �� �� �� Ӧ���22��4 6 ��47�ݡ����⣬����ָ��ijЩ�ṹ����İ�����������ڻ��������� N-������ D-��������ã����� ��-N �����ĸ�����;��жԳ��Եĸʰ��ᡣ
������Щ���ֶ�ϸ�������ĵ������ָ�����ã����������Դ���һЩ���⡣���磬�о�ϵͳ���ǻ����ģ�������֪����������绷����� N-����֮�����������Эͬ�������Ӱ�졣ͬʱ������λ�� N-������ ��-ת�ǵ��γ���������Ӱ��Ҳ������;ͬʱ�����������ɸѡ�� N-�������Խ���ø�Ϊ���ѣ���Ϊ��Ҫ�ϳɵ�������������ָ�����������ԣ���Ҫ��չ���Կ��ٵõ�����ϸ���� �� �� �� N-�� �� �� �� �� �� ����Jacobson ��Lokey �ȣ�49�ݱ�����ѡ���Լ����Ĺ����Դ˾���һ�������ԣ����Ǿ���Ŀ��ʵ�ֻ��нϴ��ࡣ
3. 2 ����ֲ�ﻷ�ĵĸ������
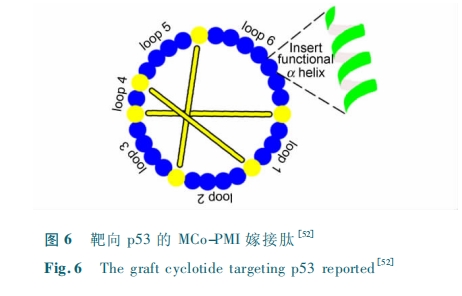
����ֲ�ﻷ�ĵ�ϸ�������ĵ����˼·�ǽ�����������ԵĶ������мӵ�ֲ�ﻷ�ĹǼ��ϡ�������о��߽�����������мӵ�ֲ�ﻷ��������������е������ȶ��ԡ����� Daly �Ƚ� VEGF-A�������� Kalata B��51�ݡ�֮���о��߷��ּӶ��Ŀ�����ϸ��Ĥ����߿ڷ��������öȣ�33�ݡ����� MCoTI-�� �� �� Ĥ �� �ԣ�Camarero �� �� �� ��Hdm2 �� HdmX �� �� �������в��� MCoTI-��� 6 �Ż��õ� MCo-PMI ����(ͼ 6)��MCo-PMI �ܹ����� Hdm2 �� HdmX ���ã������ְ����� p53 �Ļ��ԣ�52�ݡ���С��ģ����֤ʵ������ҩ���� 40 mg / kgʱ��С��������������������˵���Ӷ��� MCoPMI ȷʵ���Դ���ϸ��Ĥ��������ڵİбꡣ
��ȻĿǰ�Ѿ�����һЩֲ�ﻷ�Ŀ���ͨ��ϸ��Ĥ���������е�ijЩ������ MCoTI �� Kalata B ��Ϊ���ĹǼܱ����ڼӶ��ĵ��о�����������ֲ�ﻷ�ĵĿ�Ĥ��ʽ��Ϊϸ��������ȡ����Ҫ�������������ҹ��̸��ӣ�����������ͬʱ������̲�����ɢһ���㷺������С����ҩ�С���ˣ�ֲ�ﻷ�Ŀ�ĤЧ���Բ���ȷ���Բ�ͬϸ�������ܶ�Ҳ��������ʶ�����Ҫ��һ������鵼ֲ�ﻷ�Ŀ�Ĥ�ķ��ӻ���;ͬʱ������Ҫϵͳ���о����Խ���ϸ��Ĥ��ֲ�ﻷ���������ڵ�ҩ������ѧ���������ĤЧ�ʼ������ڵķֲ�״�����ڶ�ֲ�ﻷ�ĵĿ�Ĥ�ṹ������ҩ������ѧ��ʶ��������û�ѧ�ֶ��Ż����Ľṹ��ʹ�������ڵ���Ϊ�����ϳ�ҩ��Ҫ��
3. 3 ���������ĵĸ������
Ϊʹ�����Ļ�����õ����ľ���ϸ����������ͨ��ʹ�õIJ��������������ġ������ġ����Ժ��ڻ���������ϸ�������С�
3. 3. 1 ������
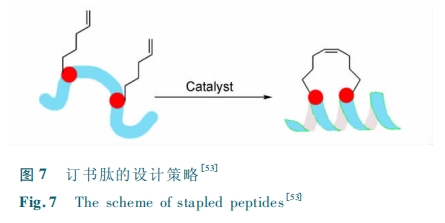
2000 �꣬Verdine С �� �� �� �� �� �� �� ( stapledpeptides)�ĸ���(ͼ 7)������ �� ����������������л��IJ�����һ�����ȵ�̼�����������Ӷ���Ч�ȶ������ĵ� �� �����ṹ��53�ݡ����øü�������С����2004 �걨����һ������ BCL-2 �鵼��ϸ���������̵Ķ�����( stabilized alpha-helix of BCL-2 domains��SAHBs)��54�ݡ����Ƿ��֣��� SAHB ������������ϸ�����̽鵼�´��� Jurkat ��Ѫ��ϸ����ϸ��Ĥ��֮���о��߷��ֲ��о���һϵ�п���ͨ��ϸ��Ĥ�Ķ����ģ�55 �� 58�ݡ���� N-������ǿ��Ĥ�����IJ��ԣ�Lin �Ƚ� N-�������붩�����У���һ������˶����ĵĿ�Ĥ������59�ݡ�������Ҫָ������������ˮ���еĽṹ��ȷ���ʵ������뻯ѧ���β�����ҵ�Ӱ����ṹ���ʶ���Ƚṹ������ͨ���ģ��������ڱ��ֶ���ԭ�л��Ի����ϣ����� N-���������λ�ѧ����������Ĥ������
3. 3. 2 ������ϸ������
ϸ������ ( cell penetrating peptides��CPP) ��һ �� �� �� �� �� ϸ �� Ĥ �� �� �� �ģ��� �� �� Ĥ ��( penetratin )��60�ݡ�ת ¼ �� �� �� �� ( trans-activator oftranscription��Tat ) Ƭ �Σ�61�ݡ�Pep-1��62�ݡ��� �� �� ��( polyarginines)��63�� �ȡ�CPP �� �� ͨ �� ϸ �� �� �� ��ȡ��64�ݻ������������ת�˹��̣�65��66�ݽ���ϸ�������ܽ���֮���ۻ�ǹ������ӵĵ����������ϸ����ϸ�����ij������൱��ľ����ᣬ��������磬�ܹ���ϸ��Ĥ����ĸ���ɷ����������ã���ǿ��ϸ��Ĥ������á�����ྫ����� CPP ( arginine richpeptides���ң�P) ͨ����ϸ��Ĥ�����ʹϸ��Ĥ�䱡������˲ʱ�״Ӷ�����ϸ��Ĥ������ CPP �ܹ������乲�����ӵķ��Ӵ���ϸ�����ص㣬Pei �Ƚ� CPP���۵����ӵ����ķ����ϣ��ɹ�ʵ���˻��ķ��ӿ�Ĥ��67�ݡ����ǣ�ֱ�����ε� �ң�P ����ϸ�����������ʱ����ϸ�����ԣ�����ϸ��������68 �� 70�ݡ�
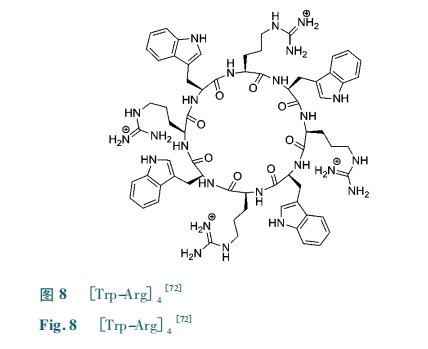
Ϊ��� CPP �Ŀ�ĤЧ�ʣ�Herce �� Cardoso �Ƚ�Tat �Ͷྫ���������β�������ң�P �Ļ���ʹ����ϸ��Ĥ���ʱ���ؼ�ЧӦ���ͣ���߿�ĤЧ�ʣ�71�ݡ����⣬�� �� �� �� �� �� �� �� �� �� �� Ҳ �� �� �� �� Ĥ��Parang �������һ������ CPP ��˼·:�߱��������ҽṹ���ԵĻ��Ŀ����ܹ�ͨ��ϸ��Ĥ��72�ݡ����ǻ�����һ�²����������ƺ�ɸѡ�����ֻ��ģ�TrpArg��4 (ͼ 8)�ͣ�Trp-Arg��5 �нϸߵĿ�ĤЧ�ʣ�����ʾ�� �� ϸ �� �� �� �� �� ( non-endocytosis ) �� �� �㡣��Trp-Arg��4 �������Խ��������ӵ�С����ҩ������ϸ����73�ݣ����������ữ�������γɸ����ﲢ��֮����Ĥ�ڣ�74�ݡ�
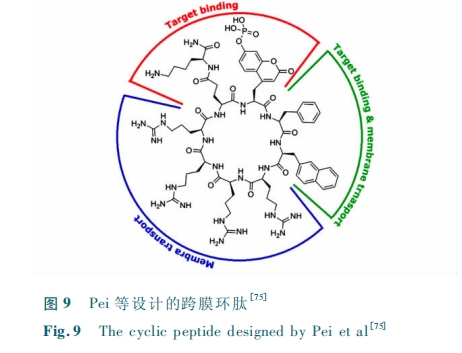
Pei �Ȼ��� �� �� �� CPP �� �� �� Ԫ �� �� �� �� ��(ͼ 9)�����ڻ��ߴ�Ϊ 7 �� 13 ���л��Ļ���������һ�� CPP СƬ�ΰ������Ľ���ϸ����75�ݡ���ʵ���У����Ƿ� �� �� Phe-��-Arg4 ( �� �� �� Ϊ L-2-�� �� �� ����)����л���Ϊ 7 �� 13 �Ļ��ĺ��ľ����̽���ϸ����������ǿ���������������Խ����Կ�Ĥ�����ữ������л�������ڡ�
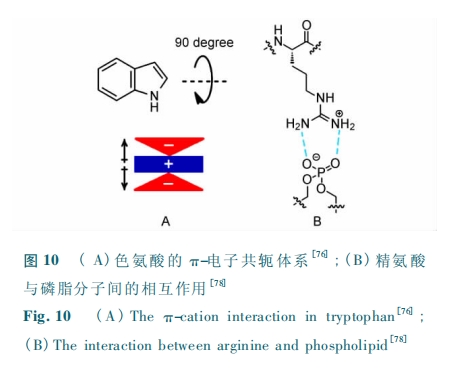
�������� CPP ���о����֣�����ɡ���������ˮ����������붼����ǿ������ϸ��Ĥ�������á����У��������ɫ�����ڶ�����Ĥ���������о�����Ҫ���ã�76��(ͼ 10)��ɫ����IJл����Ц�-���ӹ�����ϵ�������������ӷ���������-�� ����ã��ʶ�ɫ����һ�㴦��ˮ����ϸ��Ĥ֮��Ľ��紦������֬���Ӵ�����ɵĺ��������������á���������һ���˫��������壬������ϸ��Ĥ�������������γ�˫���������Ч�������Ĥ�������ã�77�ݡ�
4 ����
����ͨ���Զ�����ȣ�ϸ�������ľ��нϺ������ȶ��ԣ����ܹ�����ϸ��Ĥ�������Խ��ϸ���ڰб꣬�߱��ܺõij�ҩ�ԣ����ϸ���������Ѿ���Ϊ��ǰҩ���о������ȵ㡣���Ĵ���Ȼ������Դ���˹�������Դ������������ϸ��������������о���չ����չǰ�����ڻ�ѧ�ϳɻ�����軷�ķ��ӵĻ����ϣ�������Ȼ��Ҫ��һ������ķ��ӵĿ�Ĥ���������ķ����������Ĥ������õķ��ӻ��ƺ;�����̣�����ơ�����õ�����ϸ�������ԵĻ���ҩ�
��������������Ϊ��ҵ����ѧϰ����Ȩ��ԭ����ԭ��־���У�������Ȩ������ϵɾ�������±�ע���������³����������Ķ�ԭ�ļ��ο����ף����Ķ�ԭ��־��
