
’Σ ΓΓ“Σ ΓΓΕύκΡ‘ΎΜζΧε÷–ΨΏ”–÷Ί“ΣΒΡ…ζάμΙΠΡή ,≤Δ«“ΥϋΨΏ”–Μ·―ßΚœ≥…ΚΆ–ό ΈΒΡ±ψάϊ–‘ ,“ρ¥ΥΈϋ“ΐΝΥ‘Ϋά¥‘ΫΕύΒΡΜ·―ßΓΔ…ζΈο―ßΦΑΤδΫΜ≤φΝλ”ρ―–ΨΩ»Υ‘±ΒΡ―–ΨΩ–Υ»ΛΓΘ…ώΨ≠κΡ FF(NPFF) ΉςΈΣΑΔΤ§ΒςΫΎκΡ‘ΎΑΔΤ§ΡΆ ήΒ»“©άμ―ßΖΫΟφΨΏ”–÷Ί“ΣΒΡΒςΫΎΉς”ΟΓΘ»ΜΕχΤυΫώΈΣ÷Ι»‘»±ΖΠ NPFF ήΧεΗΏ―Γ‘ώ–‘ΒΡΦΛΕ·ΦΝΚΆόΉΩΙΦΝ ,¥”ΕχΉηΑ≠ΝΥNPFF “©άμ―ßΙΠΡήΦΑΤδΉς”ΟΜζ÷ΤΒΡ―–ΨΩΓΘ±ΨΈΡΦρ ωΝΥ NPFF ΒΡΖΔœ÷ ,Ήέ ωΝΥΫϋΦΗΡξά¥‘Ύ NPFF ΒΡ«ΑΧεΓΔ ήΧεΚΆ…ζάμ―ßΙΠΡήΒ»ΖΫΟφΥυ»ΓΒΟΒΡΉν–¬Ϋχ’ΙΓΘΫαΚœ±Ψ Β―ι “ΒΡΙΛΉς ,÷ΊΒψΫι…ήΝΥ NPFF ΙΙ–ßΙΊœΒΖΫΟφΒΡ―–ΨΩ ,≤Δ’ΙΆϊΝΥΗΟ―–ΨΩΖΫœρΫώΚσΒΡΖΔ’Ι«ς ΤΓΘ
ΨΏ”–ΒςΩΊΉς”ΟΒΡΕύκΡ”κΤδ ήΧε (“ΜάύΚ§”–10 % ΓΣ20 %ΕύΧ«ΒΡΧ«ΒΑΑΉ) ΒΡœύΜΞΉς”Ο «―–ΨΩ”κΤάΦέ…ζΈο¥σΖ÷Ή”(ΒΑΑΉ÷ ΓΔΕύΧ«ΚΆΚΥΥαΒ») ”κ–ΓΖ÷Ή” Ε±πΉς”ΟΒΡ÷Ί“ΣΉι≥…≤ΩΖ÷÷°“ΜΓΘœΗΑϊΙΠΡήΓΔΑϊΦδ–≈Κ≈ΉΣΒΦΚΆΟβ“Ώ”Π¥πΒΡΒςΩΊ¥σΕύ ΐΕΦ «Ά®ΙΐκΡ2ΒΑΑΉ÷ ÷°ΦδΒΡΖ«Ι≤ΦέΉς”ΟΡΘ Ϋά¥ Βœ÷ΒΡ ,ΜζΧε÷–ΡΎ‘¥–‘ΒΡ…ζΈοΜν–‘κΡΨΏ”–÷Ί“ΣΒΡ…ζΈο―ßΙΠΡή[1 ] ΓΘ»ΜΕχ ,≥ΐ“»ΒΚΥΊΓΔ¥Ώ≤ζΥΊΚΆΫΒ―ΣΗΤΥΊΒ»…Ό ΐκΡ÷°Άβ ,Ψχ¥σ≤ΩΖ÷ΡΎ‘¥–‘ΒΡΜν–‘κΡΕΦ≤ΜΡή÷±Ϋ”ΉςΈΣ“©Έοά¥ Ι”Ο ,Εχ‘ΎΝΌ¥≤…œΨ≠≥Θ Ι”ΟΒΡ“©ΈοΈΣ’β–©κΡάύ≈δΧεΒΡάύΥΤΈο ,Φ¥Τδœύ”ΠΒΡΦΛΕ·ΦΝΜρόΉΩΙΦΝ[1 ] ΓΘ“ρ¥Υ ,κΡάύ≈δΧεΒΡΙΙΫ®ΚΆ…Η―Γ“―≥…ΈΣΩΣΖΔ»Ϊ–¬ΒΡ“©ΈοΚΆ“©Έο«ΑΧεΓΔ…ζΈο―ßΚΆ“©άμ―ßΙΛΨΏΒΡ÷Ί“Σ≤Ώ¬‘÷°“ΜΓΘ
‘ΎΝΌ¥≤…œ ,ΑΔΤ§άύ“©ΈοΙψΖΚ”Ο”Ύ÷ΈΝΤΚΆΜΚΫβΗς÷÷÷–Ε»ΒΫ÷ΊΕ»ΧέΆ¥ ,ΒΪ¥Υάύ’ρΆ¥“©ΨΏ”–“Μ–©―œ÷ΊΗ±Ής”Ο(»γ±ψΟΊΓΔ≈ΜΆ¬ΓΔΚτΈϋ“÷÷ΤΓΔ’ρΨ≤ΚΆ“©Έο“άάΒ) ,ΦΪ¥σΒΊœό÷ΤΝΥΥϋ‘ΎΝΌ¥≤÷–ΒΡ”Π”Ο[2 ] ΓΘΫϋ 10 Ρξά¥ ,ΈΣΝΥ…ν»κΒΊΧΫΧ÷ΡΎ‘¥–‘ΑΔΤ§œΒΆ≥ΒΡΉς”ΟΜζ÷Τ ,Έ“Ο« Β―ι “≥ΛΤΎ¥” ¬ΡΎ‘¥–‘ΑΔΤ§κΡΒΡΙΙ–ßΙΊœΒΦΑΤδœύΙΊΒΡ“©άμ―ßΜν–‘―–ΨΩ[3 ΓΣ28 ],”»Τδ «‘ΎκΡάύ≈δΧεΒΡ…Η―ΓΖΫΟφ»ΓΒΟΝΥ“Μ–©ΫαΙϊ ,ΜώΒΟΝΥ“ΜœΒΝ– ΠΧΑΔΤ§ ήΧεΓΔΠΡΑΔΤ§ ήΧεΚΆΙ¬Ζ»κΡ ήΧεΒΡΗΏ―Γ‘ώ–‘ΒΡΦΛΕ·ΦΝΜρόΉΩΙΦΝ[18 ΓΣ28 ] ΓΘ
ΗυΨίΫϋΡξά¥“Μ–©Ήν–¬ΒΡ“©άμ―ß―–ΨΩΫαΙϊΆΤ≤β :ΑΔΤ§ ήΧεΒΡΦΛΜνΜα“ΐΤπΜζΧεΡΎΒΡΩΙΑΔΤ§œΒΆ≥≥ωœ÷œύ”ΠΒΡΜνΜ· ,≤Δ‘ΎΜζΧεΡΎ¥οΒΫ…ζάμΤΫΚβ[29 ] ΓΘΈ“Ο« ‘ΆΦΆ®Ιΐ…ν»κΒΊΝΥΫβΡΎ‘¥–‘ΩΙΑΔΤ§œΒΆ≥ΒΡΉς”ΟΜζ÷Τά¥»ΪΟφάμΫβΑΔΤ§ΒΡΆ¥ΨθΒςΫΎΓΔΡΆ ήΚΆ≥…ώΪΒ»“©άμ―ßΙΠΡήΓΘ…ώΨ≠κΡ FF(NPFF) «ΉνΨΏ¥ζ±μ–‘ΒΡΩΙΑΔΤ§κΡ÷°“Μ ,Έ“Ο«―–ΨΩΉιΉνΫϋ 3 Ρξά¥Έß»ΤΤδΙΙ–ßΙΊœΒΦΑΤδœύΙΊΒΡ“©άμ―ßΜν–‘Β»―–ΨΩΡΎ»ίΩΣ’ΙΝΥ“Μ–©≥θ≤ΫΒΡ―–ΨΩΓΘ”…”ΎΤΣΖυΒ»ΖΫΟφΒΡΩΦ¬« ,±ΨΈΡ÷ς“Σ≤ϊ ωΝΥ NPFF ΙΙ–ßΙΊœΒ―–ΨΩΖΫΟφΒΡΉν–¬Ϋχ’ΙΓΘ
1 ΓΓ…ώΨ≠κΡ FF ΦΑΤδ ήΧεΒΡΖΔœ÷
1. 1 RFa Φ“ΉεΦΑ…ώΨ≠κΡ FF ΒΡΖΔœ÷
1977 Ρξ , Price ΚΆ Greenberg ¥” »μ Χε Ε· Έο Ηρ(macrocallisa nimbosa) ΒΡΧεΡΎΖ÷άκ≥ωΨΏ”––Ρ‘ύ–ΥΖήΙΠΡήΒΡΥΡκΡ FMRF2NH2(Phe2Met2Arg2Phe2NH2)[30 ] ΓΘΖΔœ÷‘ΎœΏ≥φΓΔΙϊ”§ΚΆΜ»≥φΒ»ΈόΦΙΉΒΕ·ΈοΧεΡΎΕΦ¥φ‘ΎΉ≈FMRF2NH2 ―υΒΡΟβ“ΏΖ¥”Π ,Ϋχ“Μ≤ΫΖ÷άκΒΟΒΫ“ΜœΒΝ– CΡ©ΕΥΨυ“‘ RF2NH2 ΫαΙΙΫαΈ≤ΒΡ–¬κΡ ,”Ύ «ΫΪ’β–©ΫαΙΙœύάύΥΤΒΡκΡΕΦΙιΈΣ“ΜΗωΦ“Ήε ,Φ¥ FMRFa œύΙΊκΡ(FaRPs) Φ“Ήε ,ΫϋΤΎ―–ΨΩ»Υ‘±“≤ΫΪΥϋΟ«≥ΤΈΣ RFa κΡ(RFamide peptides)[31 ] ΓΘ
»ΜΕχ ,‘ΎΦΙΉΒΕ·ΈοΧεΡΎ―Α’“ RF2NH2 κΡΒΡΙΛΉςΫχ’Ι±»ΫœΜΚ¬ΐ ,÷±ΒΫ 1981 Ρξ ,Weber Β»[32 ] Ή¥Έ÷Λ ΒΝΥ‘ΎΦΙΉΒΕ·ΈοΧεΡΎ“≤¥φ‘ΎΉ≈άύΥΤ FMRFa ΒΡ RF2NH2κΡΓΘΥφΚσ ,Dockray Β»[33 ] ”Ύ 1983 Ρξάϊ”ΟΩΙ FMRFa ΒΡΩΙΧεΫχ––Ζ≈ΟβΖ÷Έω ,¥”ΦΠΡ‘÷–Ζ÷άκ≥ωΦΙΉΒΕ·ΈοΧεΡΎΒΡΒΎ“ΜΗω RF2NH2 κΡ ΓΣΓΣΓΣLRLRFaΓΘ”–»ΛΒΡ « ,ΥφΚσΖΔœ÷ΑΔΤ§κΡΦΉΤΏκΡ ( YGGFMRF) ΒΡ C Ρ©ΕΥΥΡκΡ–ρΝ–ΫαΙΙ”κ FMRF2NH2 ΒΡΑ±ΜυΥα≤–Μυ–ρΝ–Άξ»Ϊ“Μ÷¬ ,¥”ΕχΆΤ≤β FMRF2NH2 Ω…Ρή «”κΡ‘Ζ»κΡ“ΜΤπ–≠Ά§ΫχΜ·Εχά¥ΒΡ[34],»ΜΕχ“©άμ―ß―–ΨΩΫαΙϊ»¥œ‘ ΨΦΉΤΏκΡΚΆ FMRFa±μœ÷≥ωΆξ»ΪœύΖ¥ΒΡ…ζάμΙΠΡή[34 ,35] ΓΘΈΣΝΥΫχ“Μ≤ΫΧΫΧ÷FMRF2NH2 ―υΈο÷ ΒΡ…ζΈοΜν–‘ ,1985 Ρξ , Yang Β»[36] ‘Υ”ΟΟβ“Ώ«ΉΚœΒΡΦΦ θ ,”ΟΩΙ FMRF2NH2 ΒΡ―Σ«ε¥”≈ΘΡ‘÷–Ζ÷άκΒΟΒΫΝΥΝΫ÷÷–¬ΒΡΨΏ”–ΩΙΑΔΤ§Μν–‘ΒΡ RF2NH2 κΡ ,Φ¥ΑΥκΡΫαΙΙΒΡ NPFF(FLFQPQRF2NH2) ΚΆ °ΑΥκΡΫαΙΙΒΡ NPAF(AGEGLSSPFWSLAAPQRF2NH2 ) ΓΘ
Υδ»Μ NPFF ΚΆ NPAF «ΉςΈΣ¬πΖ»ΒΡΒςΫΎκΡ±ΜΖΔœ÷ΒΡ ,ΤδΙΠΡή≤Δ≤ΜΫωΫωΨ÷œό”ΎΕ‘ΑΔΤ§œΒΆ≥ΒΡΒςΫΎΓΘ“―”–ΒΡ±®Βά±μΟς , NPFF ≤ΜΫωΨΏ”–ΩΙΑΔΤ§ΒΡΜν–‘(anti2opioid) ,ΥϋΜΙΨΏ”–Φ”«ΩΑΔΤ§Ής”Ο ,Φ¥‘≠ΑΔΤ§Μν–‘(pro2opioid) ,“ρ¥Υ‘γΤΎΒΡΩΙΑΔΤ§κΡ“≤ΕΦ±ΜΙιΈΣΑΔΤ§ΒςΫΎκΡ (opioid2modulating peptides)[29 ] ΓΘ¥σΝΩΒΡ―–ΨΩ±μΟς ,NPFF ΦΑΤδœύΙΊκΡ‘ΎΆ¥ΨθΓΔΑΔΤ§ΡΆ ήΚΆ≥…ώΪΓΔ–Ρ―ΣΙήœΒΆ≥ΓΔ…ψ ≥ΓΔΧεΈ¬ΓΔœϊΜ·Βά‘ΥΕ·ΓΔΥ°ΤΫΚβΚΆΡΎΖ÷ΟΎΒ»ΖΫΟφΕΦΨΏ”–÷Ί“ΣΒΡΒςΫΎΉς”Ο[37 ] ΓΘ
1. 2 ΓΓ…ώΨ≠κΡ FF «ΑΧεΒΡΦχΕ®
Ή‘¥” NPFF ΚΆ NPAF ΖΔœ÷÷°Κσ ,Ϋχ“Μ≤ΫΒΡ Β―ιΖΔœ÷‘ΎΤδΥϋΦΗ÷÷≤Η»ιΕ·ΈοΒΡ÷– ύ…ώΨ≠œΒΆ≥÷–“≤Ά§―υ¥φ‘ΎΉ≈ NPFF ―υΒΡΟβ“ΏΖ¥”Π ,ΒΪ «Έ¥Ρή¥”ΥϋΟ«ΧεΡΎΖ÷άκ¥ΩΜ·≥ω–¬ΒΡ NPFF œύΙΊκΡΓΘ÷±ΒΫ 1995 Ρξ ,≤≈¥”¥σ σΒΡΦΙΥηΚΆΡ‘÷–Ζ÷άκ≥ω“Μ÷÷ NPFF ―υΈο÷ ΓΣΓΣΓΣNPSF[38 ] ΓΘΥφΉ≈ΫϋΡξά¥Ζ÷Ή”…ζΈο―ßΦΦ θΒΡΖΔ’Ι ,‘Ύ»ΥΓΔ≈ΘΓΔ¥σ σΚΆ–Γ σΧεΡΎΖ÷±πΩΥ¬ΓΒΟΒΫΝΥ±ύ¬κ NPFFΠΑAFΠΑSF ΒΡΜυ“ρ (pro2NPFFA «ΑΧεΒΡΜυ“ρ)[39 ,40 ] ΓΘ”κ¥ΥΆ§ ± ,Hinuma Β»(2000 Ρξ) ΚΆ Liu Β»(2001 Ρξ) ΜΙΖ÷±πάϊ”Ο±μ¥ο–ρΝ–±ξ«© ( ESTs ,expressed sequence tags) ‘ΎGen2Bank ΐ Ψί Ωβ ÷– ―Α ’“ –¬ ΒΡ RF2NH2 œύ ΙΊ κΡ(FaRPs) ,‘Ύ¥ΥΙΐ≥Χ÷– ,ΖΔœ÷ΝΥ“ΜΗω–¬ΒΡ NPFF ΒΡ«ΑΧε ΓΣΓΣΓΣpro2NPFFB «ΑΧε[41 ,42 ] ΓΘΫχ“Μ≤ΫΒΡΖ÷άκΦχΕ®ΖΔœ÷ ,‘Ύ≤ΜΆ§ΒΡ÷÷ τ÷– ,NPFF ΒΡΝΫΗω«ΑΧεΒΑΑΉΒΡΫαΙΙΚΆΟΗ«–ΖΫ Ϋ“≤”–Υυ≤ΜΆ§ ,¥”ΕχΒΟΒΫΝΥ“ΜœΒΝ– NPFFœύΙΊκΡΓΘ«ΑΧε pro2NPFFA Ψ≠ΟΗ«–ΚσΒΟΒΫΝΫ÷÷≤ΜΆ§–ρΝ–ΒΡ…ώΨ≠κΡ : (1) Κ§”– NPFF(FLFQPQRF2NH2) –ρΝ–ΒΡκΡ ; (2) Κ§”– NPSF (SLAAPQRF2NH2) –ρΝ–ΒΡκΡΓΘΕχpro2NPFFB «ΑΧε“≤Ρή…ζ≥…ΝΫ÷÷άύ–ΆΒΡ RF2NH2 κΡ : (1)Κ§”– LPQRF2NH2 –ρΝ–ΒΡ…ώΨ≠κΡ ; (2) Κ§”– LPLRF2NH2 –ρΝ–ΒΡ…ώΨ≠κΡ[43 ΓΣ45 ] ΓΘ’β–©…ώΨ≠κΡΒΡΨΏΧε–ρΝ–ΫαΙΙ»γ±μ 1 Υυ Ψ ,ΒΪ±μ÷–ΥυΝ–≥ωΒΡ“Μ–©κΡΒΡΟΗ«–ΈΜΒψ»‘–ηΫχ“Μ≤ΫΒΡ Β―ι÷Λ ΒΓΘ

1. 3 ΓΓ…ώΨ≠κΡ FF ήΧεΒΡΦχΕ®
Υδ»ΜΫœ‘γΨΆΖΔœ÷ΝΥ NPFF ,ΒΪ ήΒΫ‘γΤΎ―–ΨΩ ÷ΕΈΒΡœό÷Τ ,Ε‘ NPFF ήΧεΒΡ―–ΨΩœύΕ‘±»Ϋœ÷ΆΚσΓΘΉν≥θ ,÷ΜΡή”ΟΖ≈ΟβΫαΚœ Β―ι―Α’“ NPFF ΒΡΫαΚœΈΜΒψά¥―–ΨΩΤδ ήΧεΒΡΩ…ΡήΖ÷≤Φ«χ”ρ[46 ,47 ] ΓΘ2000 Ρξ ,άϊ”ΟΖ÷Ή”…ζΈο―ßΦΦ θ ,»’±ΨΓΔ”ΔΙζΚΆΟάΙζΒΡ 3 Φ“ Β―ι “ΦΗΚθΆ§ ±ΖΔœ÷ ΝΥ NPFF ΒΡ ΝΫ ÷÷ ≤Μ Ά§ ΒΡ G ΒΑ ΑΉ ≈Φ ΝΣ ή Χε(GPCR) ,Φ¥ NPFF1 ΚΆ NPFF2 ήΧε (ΨΏΧεΦϊ±μ 2) ,«“NPFF1 ”κ NPFF2 Φδ ÷Μ ”– 50 % ΒΡ Α± Μυ Υα –ρ Ν– œύΆ§[42 ,48 ,49 ] ΓΘ‘≠ΈΜ‘”ΫΜ Β―ι±μΟςΥϋΟ«‘ΎΧεΡΎΒΡΖ÷≤Φ“≤¥φ‘Ύ“ΜΕ®ΒΡ≤ν“λ ,Τδ÷– NPFF1 ήΧε÷ς“ΣΖ÷≤Φ”Ύ«πΡ‘ΒΡ χ≈‘ΚΥ…œ ,Εχ NPFF2 ήΧ廥÷ς“ΣΖ÷≤Φ”ΎΦΙΥηΚΆ«πΡ‘ΒΡΆβ≤ύ«χ”ρ[41 ] ΓΘ
”κ“―ΦχΕ®ΒΡ GPCRs œύ±»Ϋœ ,ΖΔœ÷ NPFF ήΧε”κ“Μ–©ΫιΒΦ…ψ ≥ΙΠΡήΒΡ GPCRs ΫαΙΙœύάύΥΤ ,»γ NPFF1 ήΧε”κ»ΥάύΒΡ 26RFa ΒΡ ήΧε ( GPR103 ,“≤±ΜΟϋΟϊΈΣSP9155ΠΑAQ27 , 49 %)[50 ] ΓΔ ≥ ”ϊ ΥΊ ή Χε 1 ( orexin1 ,37 %) ΓΔ ≥”ϊΥΊ ήΧε 2 (orexin2 ,35 %) ΓΔ…ώΨ≠κΡ Y Y2 ήΧε (NPY Y2 , 34 %) ΓΔΒ®Ρ“ ’ΥθΥΊ A ήΧε (CCKA ,34 %) ΓΔ…ώΨ≠κΡ Y Y1 ήΧε(NPY Y1 ,32 %) ΓΔ¥Ό»ιœΌΥΊ ΆΖ≈κΡ ήΧε( GPR10 ,32 %) ΚΆ…ώΨ≠κΡ Y Y4 ήΧε(NPYY4 ,31 %) Β» ήΧε÷°ΦδΕΦΨΏ”–“ΜΕ®ΒΡΆ§‘¥–‘[49 ] ΓΘ”–»ΛΒΡ « ,ΉνΫϋ–¬ΖΔœ÷ΒΡΙ¬Ευ GPCRs “≤Ρή±Μ NPFFœύΙΊκΡΫαΚœ≤ΔΦΛΜν ,»γΉνΫϋ–¬ΖΔœ÷ΒΡ“ΜΗω‘Ύ≤Η”ΐΕ·ΈοΧεΡΎΧΊ“λ–‘±μ¥ο Mrgs (mas2relatedgenes) ήΧεΦ“Ήε (2001 Ρξ) ,ΑϋΚ§”– 50Εύ÷÷Ι¬Ευ GPCRs ,ΥϋΟ«ΕΦΧΊ“λ–‘ΒΊΖ÷≤Φ”Ύ»ΥΚΆ σΒΡ±≥Ηυ…ώΨ≠ΫΎ–Γ…ώΨ≠‘ΣΚΆ»ΐ ≤φ …ώ Ψ≠ ‘Σ Β» Η– Ψθ …ώ Ψ≠ ‘Σ œΗ Αϊ…œ[51 ,52 ], Εχ ±μ ¥ο MrgA1ΓΔMrgA4 ΚΆMrgC11 ήΧεΒΡ HEK œΗΑϊΡή±Μ NPFFΚΆ FMRFa ΦΛΜν[52 ΓΣ54 ] ΓΘ
Ά®ΙΐΖ≈…δ–‘≈δΧεΫαΚœ Β―ι ,ΖΔœ÷NPFF ”κΝΫ÷÷ ήΧε”–Ή≈ΫœΗΏΒΡ«ΉΚœΝΠ ,ΒΪ≤ΜΆ§ΒΡ NPFF œύΙΊκΡΕ‘ΝΫΗω ήΧεΒΡ«ΉΚœΝΠ”–Υυ≤ΜΆ§[41 ,45 ] ΓΘάϊ”ΟΖ≈…δ–‘≈δΧεΫαΚœ Β―ιΚΆΧεΆβΙΠΡή–‘Φλ≤β Β―ι≥θ≤Ϋ―–ΨΩΖΔœ÷ ,”… pro2NPFFA «ΑΧεΟΗ«–ΒΟΒΫΒΡ NPFF œύΙΊκΡ (»γ :NPA2NPFF ,NPAF , EFW2NPSW) ”κ NPFF2 ήΧε”–ΫœΗΏΒΡ«ΉΚœ–‘ ,≤ΔΨΏ”–“ΜΕ®ΒΡ―Γ‘ώ–‘(‘Φ 100 ±Ε) ;œύΖ¥ ,”… pro2NPFFB «ΑΧεΦτ«–Εχά¥ΒΡ NPVF Ε‘ NPFF1 ήΧεΨΏ”–«αΈΔΒΡ―Γ‘ώ–‘(35 ±ΕΉσ”“)[45 ] ΓΘ
2 ΓΓ…ώΨ≠κΡ FF Μ·―ß–ό Έ
NPFF ΉςΈΣ“Μ÷÷÷Ί“ΣΒΡΡΎ‘¥–‘ΑΔΤ§ΒςΫΎκΡ ,Τδ“©άμ―ßΉς”ΟΦΑΤδΜζ÷ΤΒΡ―–ΨΩΕ‘”ΎΫ“ ΨΑΔΤ§œΒΆ≥ΒΡΉς”ΟΜζ÷Τ“≤”–Ή≈ΦΪΤδ÷Ί“ΣΒΡΒΊΈΜΓΘ»ΜΕχ ,ΤυΫώ»‘»±ΖΠ―–ΨΩ NPFF ήΧεΒΡ”––ß“©άμ―ßΙΛΨΏΓΘΧΊ±π «NPFF1 ΚΆ NPFF2 ήΧε≥ωœ÷Κσ ,’βΝΫ÷÷ ήΧεΒΡΗΏ―Γ‘ώ–‘ΦΛΕ·ΦΝΚΆόΉΩΙΦΝΫΪ≥…ΈΣ≤ϊ Ά NPFF ΦΑΤδ ήΧε“©άμ―ßΉς”ΟΒΡ”–ΝΠΙΛΨΏΓΘΥδ»Μ“―”–ΒΡ NPFF ΦΑΤδœύΙΊκΡΒΡΙΙ–ßΙΊœΒ―–ΨΩΖΔœ÷ΝΥ“Μ–© NPFF ήΧεΒΡκΡάύΚΆΖ«κΡάύΒΡ≈δΧε[55 ],ΒΪ»‘»±ΖΠΕ‘”Ύ NPFF1 ΚΆ NPFF2 ήΧεΨΏ”–ΗΏ―Γ‘ώ–‘«“ΗΏ«ΉΚœΝΠΒΡάμœκ≈δΧε ,ΧΊ±π «ΗΏ―Γ‘ώ–‘ΒΡόΉΩΙΦΝΓΘ
NPFFΒΡΖΔœ÷“―Ψ≠”– 20 ΕύΡξΒΡάζ Ζ ,ΒΪΤδΙΙ–ßΙΊœΒΖΫΟφΒΡ―–ΨΩœύΕ‘”––©ΜΚ¬ΐ ,Τδ‘≠“ρ «ΕύΖΫΟφΒΡ ,»γ»±ΖΠ NPFF ήΧεΒΡΆΜ±δΚΆΨßΧεΫαΙΙΒ» ήΧεΙΙ–ßΙΊœΒΒΡ ΐΨί ,’β–©ΉηΑ≠ΝΥ≈δΧεΚΆ ήΧεΫαΚœΙΊœΒΒΡ―–ΨΩ ;NPFF ήΧεΒΡΖΔœ÷œύΕ‘÷ΆΚσ ;≤Δ«““―”–ΒΡ NPFF ΙΙ–ßΙΊœΒ―–ΨΩ“≤œύΕ‘”–œόΓΘ‘Ύ¥ΥΈΡ÷–ΫΪΑ¥’’κΡάύΦΛΕ·ΦΝΚΆόΉΩΙΦΝ…ηΦΤΒΡ“ΜΑψΥΦ¬Ζ ,ΫΪ“―±®ΒάΒΡ NPFF ΒΡΙΙ–ßΙΊœΒ ΐΨίΉω“Μ≥θ≤ΫΒΡΙιΡ…ΚΆΖ÷Έω ,œΘΆϊ”–÷ζ”Ύ…ηΦΤΚΆ…Η―Γ≥ω–¬ΒΡ NPFF ήΧεΗΏ–ßΒΡκΡάύΚΆΖ«κΡάύ≈δΧεΓΘ
2. 1 ΓΓ…ώΨ≠κΡ FF ΒΡΤ§ΕΈ―–ΨΩΦΑΤδΧΊ βΑ±ΜυΥα…®Οη
Ά®Ιΐ”–ΡΩΒΡΓΔ”–Ιφ¬…ΒΊΥθΕΧΤδ–ρΝ– ,“‘ΤΎ―Α’“Ήν–ΓΜν–‘Τ§ΕΈ «ΕύκΡΙΙ–ßΙΊœΒ―–ΨΩ≥θΤΎΒΡ“Μ÷÷≥ΘΙφΖΫΖ®ΓΘNPFF ΚΆ NPVF Ζ÷ ±π ά¥ Ή‘ pro2NPFFA ΚΆ pro2NPFFB «ΑΧε ,≤Δ«“Ε‘ NPFF1 ΚΆ NPFF2 ήΧεΒΡ―Γ‘ώ–‘≤ΜΆ§ΓΘ÷ΒΒΟΉΔ“βΒΡ «ΥϋΟ«ΨΏ”–œύΆ§ΒΡ C Ρ©ΕΥΥΡκΡ–ρΝ–ΚΆθΘΑ±ΫαΙΙ ,≤Δ«“ C Ρ©ΕΥ–ρΝ–ΈΣ NPFF œύΙΊκΡΒΡ±Θ Ί–ρΝ– ,Ζ÷±π¥”Τδ N Ρ©ΕΥΩΣ ΦΆ®Ιΐ÷πΗωΑ±ΜυΥα…ΨΦθά¥―–ΨΩΤδΙΙ–ßΙΊœΒ ,ΨΏΧεΒΡ Β―ιΫαΙϊ»γ±μ 3Υυ Ψ[43 ] ΓΘ
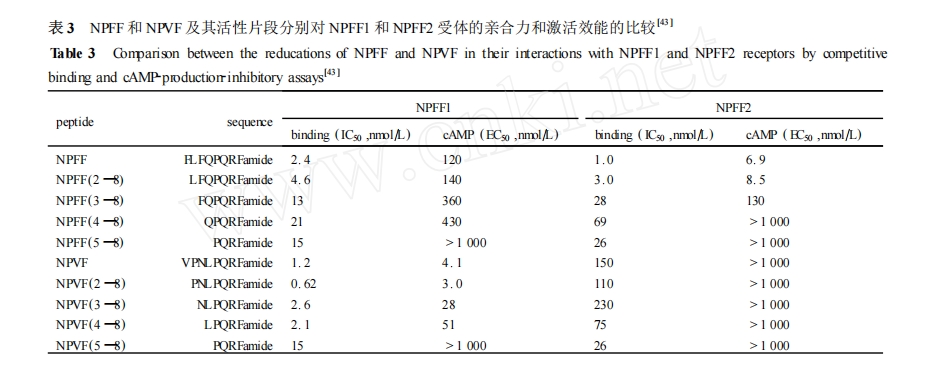
Ε‘”Ύ NPFF ,ΟΩ…Ψ≥ΐ N Ρ©ΕΥΒΡ“ΜΗωΑ±ΜυΥα≤–Μυ ,Υϋ”κ NPFF1 ΚΆ NPFF2 ήΧεΒΡ«ΉΚœΝΠΦΑΤδΧεΆβ“©άμ―ßΜν–‘ΨΆ≥ωœ÷œύ”ΠΒΡΫΒΒΆ ,‘Ύ…Ψ≥ΐΒΎ 1 ΈΜ Phe ΚσΤδ ήΧε«ΉΚœΝΠΚΆ…ζΈοΜν–‘Έ¥ ήΟςœ‘ΒΡ”Αœλ ,ΒΪ «…Ψ÷ΝΒΎ 3 ΈΜΚΆΒΎ 4 ΈΜΑ±ΜυΥα ± ,Τδ ήΧε«ΉΚœΝΠΚΆ…ζΈοΜν–‘ΕΦ≥ωœ÷Οςœ‘ΒΡΫΒΒΆΓΘΝν»Υ≤ο“λΒΡ « NPVF …Ψ≥ΐ NΡ©ΕΥΒΡΒΎ“ΜΗωΑ±ΜυΥα≤–ΜυΚσ ,Υϋ”κ NPFF1 ΚΆ NPFF2 ήΧεΒΡ«ΉΚœΝΠΚΆ…ζΈοΜν–‘Ζ¥ΕχΕΦ≥ωœ÷ΝΥ…ΐΗΏ ,≤Δ«“…ΨΒτΒΎ 2 ΈΜΚΆΒΎ 3 ΈΜΑ±ΜυΥαΚσ ,ΥϋΒΡ ήΧε«ΉΚœΝΠΚΆ…ζΈοΜν–‘ΕΦœ¬ΫΒΒΟ≤ΜΧΪœ‘÷χΓΘΕχΒΎ 4 ΈΜΒΡ Leu ΒΡ…Ψ≥ΐΜαΒΦ÷¬ΤδΕ‘ NPFF1 ήΧεΒΡ«ΉΚœΝΠΚΆ…ζΈοΜν–‘ΒΡ¥σ¥σΫΒΒΆΓΘ’β–© ΐΨί±μΟς NPFF ΚΆ NPVF ΒΡ N Ρ©ΕΥΥΡκΡΫαΙΙ «”ΑœλΥϋΟ«ΫαΚœ≤ΔΦΛΜν NPFF1 ΚΆ NPFF2 ήΧεΒΡΙΊΦϋ«χ”ρ ,Τδ÷–ΨωΕ®ΥϋΟ« ήΧε―Γ‘ώ–‘ΒΡ «NPFF ΒΡ Phe3 ΚΆ Gln4,NPVF ΒΡ Leu4 ΓΘ
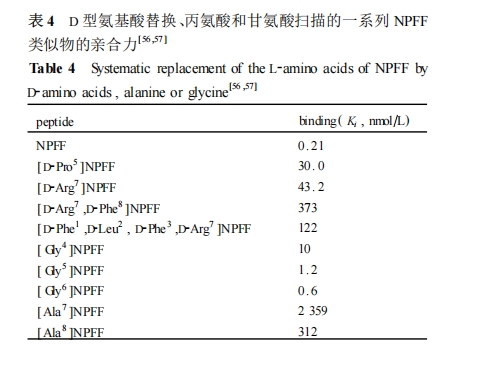
άϊ”ΟΧΊ βΑ±ΜυΥα (»γ D2–ΆΑ±ΜυΥαΓΔΗ Α±ΥαΚΆ±ϊΑ±ΥαΒ») ΧφΜΜ…®Οη «“ΜΗω…ζΈοΜν–‘κΡ±ΜΖΔœ÷ΚσΉν≥θ”Ο”Ύ…Η―ΓΤδΙΙ–ßΙΊΦϋΈΜΒψΒΡ÷Ί“ΣΖΫΖ®÷°“ΜΓΘNPFFΒΡΑ±ΜυΥα…®ΟηΧφΜΜΒΡΫαΙϊ»γ±μ 4 Υυ Ψ[56 ,57 ] ΓΘΆ®ΙΐΕ‘NPFF Ϋχ–– D –ΆΑ±ΜυΥα…®ΟηΚσΖΔœ÷ΒΎ 5ΓΔ7 ΚΆ 8 ΈΜΒΡΑ±ΜυΥα≤–Μυ±Μ»Γ¥ζ÷°ΚσΕ‘ NPFF ήΧεΒΡ«ΉΚœΝΠ¥σ¥σΫΒΒΆ ,Εχ 3 ΈΜΒΡΧφΜΜ“≤ ΙΤδ«ΉΚœΝΠ”–ΥυΫΒΒΆΓΘ”ΟΗ Α±Υα( Gly) ΧφΜΜ…®ΟηΒΡΫαΙϊ±μΟς ,ΒΎ 4 ΈΜΑ±ΜυΥαΧφΜΜΈΣ Gly ΙΒΟ«ΉΚœΝΠΫΒΒΆΫϋ 50 ±Ε ,ΒΎ 5 ΈΜΒΡΧφΜΜ ΙΤδ«ΉΚœΝΠœ¬ΫΒ‘Φ 6 ±ΕΉσ”“ ,ΕχΒΎ 6 ΈΜΧφΜΜΫωΫωΫΒΒΆ«ΉΚœΝΠ 3 ±ΕΓΘΆ®Ιΐ±ϊΑ±Υα(Ala) ΧφΜΜ…®ΟηΖΔœ÷ΒΎ 7 ΚΆ 8ΈΜΒΡΑ±ΜυΥα≤–Μυ±ΜΧφΜΜΚσΕΦ‘λ≥… NPFF άύΥΤΈο«ΉΚœΝΠ¥σ¥σœ¬ΫΒΓΘ’β–©Α±ΜυΥα…®ΟηΒΡΫαΙϊ”κ NPFF Τ§ΕΈ―–ΨΩΒΡΫαΙϊΜυ±Ψ“Μ÷¬ ,Φ¥ Phe3 ΚΆ Gln4 ΦΑΤδ≤ύΝ¥ΜυΆ≈‘Ύ ήΧεΫαΚœ÷–ΨΏ”–“ΜΕ®ΒΡΉς”Ο ,Εχ 5 ΚΆ 6 ΈΜΑ±ΜυΥα≤–ΜυΒΡ≤ύΝ¥ΫαΙΙΕ‘”ΎΫαΚœ NPFF ήΧεΒΡ”Αœλ≤Μ «Κή¥σ(ΒΪ 5 ΈΜ”Ο D –ΆΑ±ΜυΥαΧφΜΜΚσΕ‘Τδ«ΉΚœΝΠ”ΑœλΫœ¥σ ,Ω…Ρή « D2Pro ΒΡ“ΐ»κΗΡ±δΝΥ N Ρ©ΕΥΥΡκΡΒΡΩ’Φδ…λ’ΙΖΫœρ) ,Εχ NPFF ΫαΚœ ήΧεΒΡΙΊΦϋΈΜΒψ « Arg7 ΚΆPhe8,ΥϋΟ«≤ύΝ¥ΙΙœσΒΡ«αΈΔΗΡ±δΕΦΩ…ΡήΒΦ÷¬ ήΧεΫαΚœΡήΝΠΒΡ¥σ¥σΫΒΒΆΓΘ
ΝΫ÷÷ NPFF ήΧε―«–ΆΒΡ≥ωœ÷ ,±Ί»ΜΗχΙΙ–ß―–ΨΩ¥χά¥–¬ΒΡΈ Χβ : «Ζώ¥φ‘Ύ NPFF ήΧεΒΡ―Γ‘ώ–‘≈δΧε ?≥θ≤ΫΒΡ―–ΨΩΖΔœ÷ ,”…≤ΜΆ§«ΑΧεΟΗ«–Εχά¥ΒΡ NPFF œύΙΊκΡΕ‘ NPFF1 ΚΆ NPFF2 ήΧεΒΡ―Γ‘ώ–‘“≤”–Υυ≤ΜΆ§ΓΘ¥ΥΆβ ,¥” NPFF1 ΚΆ NPFF2 ήΧεΒΡ–ρΝ–ΫαΙΙΖ÷ΈωΖΔœ÷ ,ΥϋΟ«ΑϊΆβΜΖ–ΈΫαΙΙ…œΒΡΑ±ΜυΥα≤–Μυ≤ν“λ–‘Ϋœ¥σ ,ΕχΩγΡΛ«χ”ρΒΡΫαΙΙœύΕ‘±Θ Ί ,¥”ΕχΧα ΨΝΥ NPFF1 ΚΆNPFF2 ήΧεΒΡ≈δΧεΫαΙΙ≤ΜΆ§ ,Μρ’ΏΆ§“Μ≈δΧεΕ‘ΝΫ÷÷ ήΧεΒΡΫαΚœΡήΝΠ¥φ‘Ύ≤ν±πΓΘ
¥ΥΆβ ,”… NPFF ΒΡΝΫΗω«ΑΧε―ή…ζΕχά¥ΒΡ“ΜœΒΝ–NPFFœύΙΊκΡ ,Ά®ΙΐΦλ≤βΥϋΟ«”κ NPFF1 ΚΆ NPFF2 ήΧεΒΡ«ΉΚœΡήΝΠ ,Ω…“‘ΆΤ≤β≥ω NPFF œύΙΊκΡΒΡ N Ρ©ΕΥ«χ”ρΒΡΑ±ΜυΥα–ρΝ–ΫαΙΙΨωΕ®ΝΥΥϋΖ÷±π”κ NPFF1 ΚΆNPFF2 ήΧε«ΉΚœΝΠΒΡ≤ΜΆ§ ,¥”Εχ‘λ≥…ΝΥΥϋΕ‘ NPFF ήΧε―«–Ά―Γ‘ώ–‘ΒΡ≤ν“λΓΘNPFFΓΔNPSF ΚΆ NPVF ΈΣ 3 ΗωΫαΙΙάύΥΤΒΡΑΥκΡ ,ΥϋΟ«ΨΏ”–œύΆ§ΒΡ C Ρ©ΕΥΥΡκΡΫαΙΙ ,ΒΪ «ΥϋΟ«”κ NPFF1 ΚΆ NPFF2 ήΧεΒΡ«ΉΚœΝΠ¥φ‘ΎΚή¥σΒΡ≤ΜΆ§[45 ,58 ] ΓΘΕ‘”Ύ NPSF κΡ ,Ω…“‘Ά®Ιΐ―”≥ΛΤδ NΡ©ΕΥΑ±ΜυΥα–ρΝ–ά¥ΧαΗΏΤδΫαΚœ NPFF2 ήΧεΒΡΡήΝΠ ,ΕχΫαΚœ NPFF1 ΒΡΡήΝΠΗΡ±δ≤Μ¥σ ,¥”Εχ ΙΒΟ”… NPSFΒΡ N Ρ©ΕΥ―”≥ΛΕχά¥ΒΡ QFW2NPSF ΚΆ EFW2NPSF Ρή―Γ‘ώ–‘ΒΊΫαΚœ NPFF2 ήΧεΓΘΆ§―υ―”≥Λ NPFF ΒΡ N Ρ©ΕΥ“≤Ρή―Γ‘ώ–‘ΒΊΧαΗΏΤδΫαΚœ NPFF2 ήΧεΒΡΡήΝΠ ,“ρ¥Υ”… NPFF ΒΡ N Ρ©ΕΥ―”≥ΛΒΡ NPA2NPFFΓΔSQA2NPFF ΚΆSPA2NPFF Ε‘ NPFF2 ήΧε“≤ΨΏ”–ΫœΗΏΒΡ―Γ‘ώ–‘[45 ,58 ] ΓΘ÷ΒΒΟΉΔ“βΒΡ « ,NPAF “≤Ω…“‘Ω¥Ής NPFF ΒΡ N Ρ©ΕΥ―”≥ΛΕχά¥ΒΡ °ΑΥκΡ ,ΒΪ «Τδ N Ρ©ΕΥΒΡ―”≥Λ≤ΜΫωΟΜ”–ΧαΗΏΥϋΫαΚœ NPFF2 ήΧεΒΡΡήΝΠ ,Ζ¥Εχ ΙΥϋΫαΚœ NPFF1 ήΧεΒΡΡήΝΠΫΒΒΆΝΥ[45 ,58 ] ΓΘ’β–©ΆΤ¬έ»‘–η‘Υ”ΟœΒΆ≥ΒΡΙΙ–ßΙΊœΒ―–ΨΩά¥Ϋχ“Μ≤ΫΧΫΧ÷ΓΘ
2. 2 ΓΓ…ώΨ≠κΡ FF ΒΡ N Ρ©ΕΥ«χ”ρΒΡΜ·―ß–ό Έ
Ά®ΙΐΕ‘ NPFF ΒΡ N Ρ©ΕΥ«χ”ρΫχ––Α±ΜυΥαΧφΜΜΚΆΜ·―ß–ό ΈΕχ…ηΦΤΚœ≥…ΝΥ“ΜœΒΝ–ΒΡ NPFF άύΥΤΈο ,Ά®ΙΐΦλ≤βΥϋΟ«”κ NPFF ήΧεΒΡ«ΉΚœ–‘ΖΔœ÷ (ΨΏΧεΒΡ«ΉΚœ ΐΨίΦϊ±μ 5)[56 ,57 ]: (1) NPFF ΒΡ N Ρ©ΕΥΑ±ΜυΒΡ““θΘΜ·Β»Μ·―ß–ό Έ ,Ε‘ΤδΜν–‘ΒΡ”Αœλ≤ΜΧΪΟςœ‘ ; (2) N ΕΥ«χ”ρΒΡΒΎ 1 ΈΜΚΆ 2 ΈΜΑ±ΜυΥα≤–ΜυΒΡΙΙœσ≤ΜΧΪ”ΑœλΤδ ήΧεΫαΚœΡήΝΠ ,’β”κ«Α“Μ≤ΩΖ÷ΒΡ―–ΨΩΫαΙϊœύ“Μ÷¬ ,Φ¥ΫΪΤδκΡΝ¥Φθ÷ΝΈΣ NPFF(3 ΓΣ8) ,Υϋ»‘±Θ≥÷ΝΥΫœΚΟΒΡ«ΉΚœΝΠ ; (3) ‘Ύ NPFF ΒΡΤ§ΕΈΚΆΑ±ΜυΥαΧφΜΜ―–ΨΩ÷–±μΟς ,Phe3 ΚΆ Gln4 ‘ΎΈ§≥÷ ÷Χε«ΉΚœΝΠΖΫΟφΕΦΨΏ”–÷Ί“ΣΒΡΉς”Ο ;ΒΪ‘ΎΕ®ΈΜΒψΑ±ΜυΥαΧφΜΜ–ό ΈΒΡ―–ΨΩ÷–ΖΔœ÷ ,Τδ≤ύΝ¥ΜυΆ≈ΗΡ±δΚσ ήΧε«ΉΚœ–‘»‘±Θ≥÷ΒΟΫœΚΟ ,¥”ΕχΑΒ ΨΒΎ 3 ΈΜΚΆΒΎ 4 ΈΜΑ±ΜυΥαΒΡ¥φ‘ΎΕ‘Έ§≥÷ΤδΥϋΈΜΒψ ,ΧΊ±π « C Ρ©ΕΥΥΡκΡΒΡΩ’ΦδΙΙœσ”–Ή≈÷Ί“ΣΒΡΉς”ΟΓΘ¥ΥΆβ ,Ά®ΙΐΑ±ΜυΥαΧφΜΜΖΔœ÷ΝΥΝΫ÷÷±» NPFF ΗϋΈ»Ε®ΒΡ NPFF ΦΛΕ·ΦΝ : 1DMe ( [D2Tyr1,(NMe) Phe3]2NPFF) ΚΆ 3D ([D2Tyr1, D2Leu2, D2Phe3]2NPFF) ΓΘ
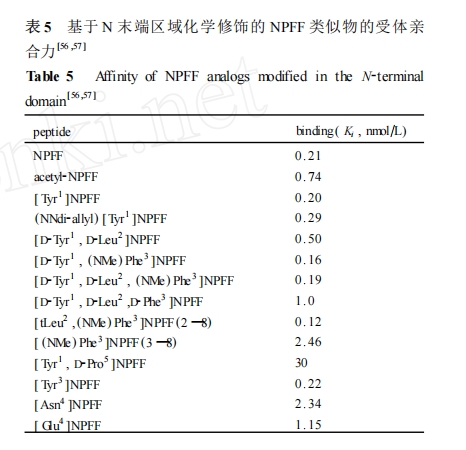
”…”Ύ‘γΤΎΒΡ«ΉΚœ ΐΨί÷ς“Σ «≤…”Ο‘»Ϋ§ΚσΒΡ¥σ σΦΙΥηœΗΑϊΡΛΈΣ≤ΡΝœά¥Φλ≤βΒΡ ,Εχ“―”–ΒΡ―–ΨΩ±μΟς¥σ σΦΙΥη÷ς“ΣΖ÷≤ΦΒΡ « NPFF2 ήΧε ,“ρ¥Υ’β–©«ΉΚœ ΐΨί÷ς“ΣΖ¥”≥ΒΡ «ΧφΜΜΚσΒΡάύΥΤΈο”κ NPFF2 ήΧεΒΡ«ΉΚœΝΠ ,¥”Εχ»±ΖΠ’β–©άύΥΤΈο”κ NPFF1 ήΧεΒΡΫαΚœ ΐΨίΓΘ’β–©Α±ΜυΥαΧφΜΜ–ό ΈΒΡ―–ΨΩΫαΙϊ”κ«Α“Μ≤ΩΖ÷ΙΙ–ßΙΊœΒΒΡΫα¬έΜυ±Ψ“Μ÷¬ΓΘΆ®ΙΐΖ÷Έω NPFF ΒΡ N Ρ©ΕΥΜ·―ß–ό ΈΒΡΙΙ–ßΙΊœΒ―–ΨΩΫαΙϊ≤ΜΡ―ΖΔœ÷ ,Υδ»ΜNPFF(5 ΓΣ8) »‘ΨΏ”–“ΜΕ®ΒΡΫαΚœΚΆΦΛΕ·Μν–‘ ,ΒΪ NPFFΒΡ N ΕΥ«χ”ρΒΡΙΙœσΗΡ±δ ,Ρή÷±Ϋ””ΑœλΥϋΕ‘ NPFF ήΧεΒΡ«ΉΚœ–‘ΚΆ―Γ‘ώ–‘ ,“ρ¥Υ ,N Ρ©ΕΥΒΡΑ±ΜυΥα≤–ΜυΈΜΒψ «ΨωΕ® NPFF άύΥΤΈοΕ‘ NPFF1 ΚΆ NPFF2 ήΧε«ΉΚœΝΠΚΆ―Γ‘ώ–‘ΒΡΙΊΦϋ«χ”ρΓΘ
2. 3 ΓΓ…ώΨ≠κΡ FF ΒΡ C Ρ©ΕΥ«χ”ρΒΡΜ·―ß–ό Έ
NPFF ΒΡ C Ρ©ΕΥ«χ”ρ «Υϋ”κ ήΧεΫαΚœΒΡ±Θ Ί–ρΝ–ΫαΙΙΓΘΕ‘ NPFF ΒΡ C Ρ©ΕΥ«χ”ρΫχ––ΙΙ–ßΙΊœΒ―–ΨΩΖΔœ÷(ΨΏΧεΒΡ«ΉΚœ ΐΨίΦϊ±μ 6)[57 ]: (1) NPFF ΒΡ C Ρ©ΕΥθΘΑΖΫαΙΙΕ‘”ΎΈ§≥÷«ΉΚœΝΠ «±Ί–ηΒΡ ,ΧφΜΜΈΣ¥ΦΜυΜρτ»ΜυΕΦΜαΒΦ÷¬Τδ ήΧεΫαΚœΡήΝΠΒΡœ¬ΫΒ…θ÷Ν…Ξ ß ;(2) ΧφΜΜΒΎ 8 ΈΜΒΡ Phe ΈΣ Tyr Μρ’ΏΗΡ±δ≤ύΝ¥ΖΦœψΜΖ”κCΠΝ ÷°ΦδΒΡΜ·―ßΦϋ≥ΛΕ» ,ΕΦ ΙΒΟ NPFF άύΥΤΈοΒΡ«ΉΚœΝΠ¥σ¥σΫΒΒΆ ; (3) ”Ο Lys ά¥ΧφΜΜ Arg7 “≤Μαœ‘÷χΫΒΒΆ«ΉΚœΝΠ ,¥”Εχ±μΟςΤδ≤ύΝ¥κ“ΜυΕ‘ ήΧεΫαΚœΤπΉ≈÷Ί“ΣΉς”Ο ,Εχ≤ΜΫωΫω÷Μ «Έ§≥÷Φν–‘ΒΡ≤ύΝ¥ΜυΆ≈ ; (4) ΒΎ 5 ΚΆ6 ΈΜΒΡΑ±ΜυΥα≤–ΜυΩ…ΡήΚΆ N Ρ©ΕΥ«χ”ρΒΡΒΎ 3 ΚΆ 4 ΈΜΒΡΑ±ΜυΥα≤–ΜυΒΡΉς”ΟœύΒ± ,ΕΦ «”Ο”ΎΈ§≥÷ C Ρ©ΕΥRF2NH2 ΒΡΩ’ΦδΙΙœσ ,“ΜΑψΒΡΑ±ΜυΥαΧφΜΜΕ‘Τδ ήΧε«ΉΚœΒΡ”Αœλ≤Μ¥σ ,ΒΪ «Ηω±πΒΡΧφΜΜΩ…Ρή”ΑœλΝΥ C Ρ©ΕΥΩ’ΦδΫαΙΙ ,»γ Pro5 ΧφΜΜΈΣ D –ΆΒΡΑ±ΜυΥαΚσ ήΧε«ΉΚœΝΠΟςœ‘œ¬ΫΒΓΘ
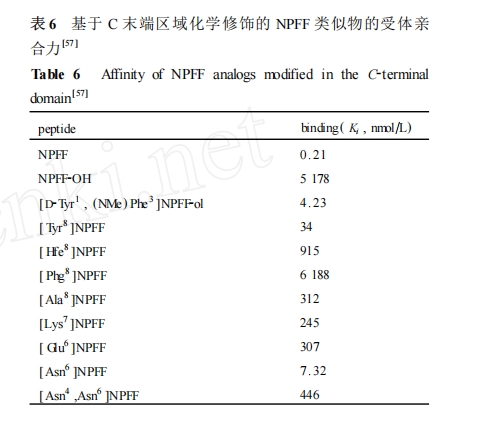
Ά§―υ ,±μ 6 ÷–ΥυΝ–≥ωΒΡ«ΉΚœ ΐΨί≤ΜΡήΖ¥”≥≥ω’β–©άύΥΤΈοΕ‘ΝΫ÷÷≤ΜΆ§ΒΡ NPFF ήΧεΒΡ―Γ‘ώ–‘ΓΘΒΪ «¥”’β–©ΙΙ–ßΙΊœΒ―–ΨΩΫαΙϊΩ…“‘ΆΤ≤β≥ω NPFF ΒΡ C Ρ©ΕΥΒΡ RF2NH2 ΫαΙΙΈΣΤδΓΑ–≈ Ι”ρΓ±, «ΨωΕ®ΥϋΫαΚœ≤ΔΦΛΜν NPFF ήΧεΒΡΙΊΦϋΈΜΒψ ,Φ¥”Αœλ NPFF άύΥΤΈο–ßΡήΒΡ÷Ί“Σ«χ”ρΓΘ“―”–ΒΡ NPFF ήΧεκΡάύόΉΩΙΦΝΒΡ…ηΦΤΚΆΜ·―ßΗΡ‘λ“≤÷ς“ΣΦ·÷–”Ύ¥ΥΫαΙΙ«χ”ρ ,ΨΏΧεΫΪ‘ΎΚσΟφ≤ΩΖ÷ΉςœξœΗΒΡΫι…ήΓΘ
ΈΣΝΥΫχ“Μ≤ΫΤάΦέ NPFF ΒΡ C Ρ©ΕΥΒΡ RF2NH2 ΫαΙΙΕ‘Τδ“©άμ―ßΜν–‘ΒΡ”ΑœλΓΘΜυ”Ύ NPFF ΒΡ C Ρ©ΕΥΒΡΥΡκΡΫαΙΙ ,≤Δ‘Ύ N Ρ©ΕΥ“ΐ»κΒΛθΘΜ·–ό Έά¥‘ω«ΩΤδ ήΧε«ΉΚœΡήΝΠ ,Έ“Ο« Β―ι–ΓΉι…ηΦΤΚœ≥…ΝΥ 4 ΗωΨΏ”–”ΪΙβΜν–‘ΒΡ NPFF άύΥΤΈο :C Ρ©ΕΥ÷–Κ§”–ΓΑ2RF2NH2Γ±ΫαΙΙΒΡ dansyl2PQRF2NH2 ΚΆ dansyl2GSRF2NH2 ; “‘ΦΑ CΡ©ΕΥ “‘ΓΑ2R2NH2 Γ±Ϋα ΙΙ Ϋα Έ≤ ΒΡ dansyl2PQR2NH2 ΚΆdansyl2GSR2NH2 ΓΘΆ®Ιΐ“ΜœΒΝ–ΧεΆβΚΆΧεΡΎ“©άμ―ß Β―ιœΒΆ≥ΒΊΧΫΧ÷ΝΥΤδΜ·―ßΫαΙΙΒΡ±δΜ·Ε‘Τδ“©άμ―ßΙΠΡήΒΡ–ßΡήΚΆ–ßΦέΒΡ”Αœλ ,―–ΨΩΫχ“Μ≤Ϋ―ι÷ΛΝΥ NPFF ΒΡ CΡ©ΕΥ2RF2NH2 ΫαΙΙΨωΕ®ΝΥΤδάύΥΤΈο–ßΡή ,ΜΙΖΔœ÷2RF2NH2ΠΑ2R2NH2 ΒΡ N Ρ©ΕΥΒΡΑ±ΜυΥα≤–ΜυΕ‘Τδ–ßΦέΒΡ¥σ–Γ“≤ΨΏ”–“ΜΕ®ΒΡ”ΑœλΓΘ¥ΥΆβ ,ΜΙΖΔœ÷–¬…ηΦΤΒΡdansyl2GSRF2 NH2 ΈΣ“Μ÷÷ΗΏ–ßΒΡΓΔΨΏ”–”ΪΙβΜν–‘ΒΡNPFF ήΧεΦΛΕ·ΦΝ[59 ,60 ] ΓΘ
2. 4 ΓΓΤδΥϋκΡάύΖ÷Ή”ΦΑΤδœύΙΊΒΡΙΙ–ßΙΊœΒ―–ΨΩ
”…”Ύ NPFF1 ΚΆ NPFF2 ήΧε”κΤδΥϋΦΙΉΒΕ·ΈοΒΡRF2NH2 κΡΨΏ”–ΫœΗΏΒΡΆ§‘¥–‘ ,“Μ–©ΤδΥϋΒΡ RF2NH2κΡΡή”κ NPFF ήΧεΫαΚœ , “―”–―–ΨΩ±μΟς FMRFaΓΔ26RFaΓΔ43RFaΓΔhPrRP231 ΚΆ fPP36 Β»…ώΨ≠κΡ“≤Ρή”––ßΒΊΫαΚœΚΆΦΛΜν NPFF1 ΚΆ NPFF2 ήΧε ,ΤδΨΏΧεΒΡ«ΉΚœ ΐΨί»γ±μ 7 Υυ Ψ[45 ,58 ] ΓΘΫχ“Μ≤Ϋ―–ΨΩ’β–©…ώΨ≠κΡ”κNPFF ήΧεΒΡΙΙ–ßΙΊœΒ“≤Ω…ΡήΈΣ―Α’“ NPFF1 ΚΆNPFF2 ήΧεΒΡ―Γ‘ώ–‘≈δΧεΧαΙ©–¬ΒΡΥΦ¬ΖΚΆΜ·―ß…ηΦΤΡΘΑεΓΘ

NPFF ΒΡΖΔœ÷”κ FMRFa ¥φ‘ΎΉ≈Οή«–ΒΡΝΣœΒ ,Ϋœ‘γ”Ο”Ύ NPFF ΙΙ–ßΙΊœΒ―–ΨΩΒΡ RF2NH2 κΡΈΣ FMRFaΓΘ1993 Ρξ ,Payza Β»[61 ]Ά®Ιΐ±»Ϋœ NPFF ΚΆ FMRFa Ζ÷±π”κNPFF ΫαΚœΈΜΒψΒΡ«ΉΚœΉς”Ο ,ΖΔœ÷ NPFF ΒΡ C Ρ©ΕΥΥΡκΡ PQRF2NH2( Ki = 12 nmolΠΑL) ΒΡ«ΉΚœ–‘ΒΆ”Ύ FMRF2NH2( Ki = 118 nmolΠΑL) ,ΒΪΥϋΟ«ΒΡΫαΚœΡήΝΠΕΦ‘ΕΒΆ”ΎNPFF( Ki = 216 nmolΠΑL) ΓΘΫχ“Μ≤Ϋ ,“‘ PQRF2NH2 ΈΣΡΘΑε ,Ά®ΙΐΕ®ΈΜΒψΑ±ΜυΥα≤–ΜυΒΡΧφΜΜΕχ…ηΦΤΚœ≥…ΝΥ“ΜœΒΝ–ΒΡάύΥΤΈο ,―–ΨΩΫαΙϊœ‘ ΨΆ®ΙΐΗΡ±δ FMRFa ΒΡPhe1 ΚΆ Met2 Ω…“‘ΧαΗΏΤδ ήΧε«ΉΚœΝΠΓΘ≤Δ«“ ,…Η―ΓΒΟΒΫ 3 Ηω«ΉΚœ–‘ΫœΗΏΒΡάύΥΤΈο : PWRF2NH2( Ki = 0154nmolΠΑL) ΓΔFFRF2NH2( Ki = 0125 nmolΠΑL)ΚΆ FWRF2NH2( Ki= 0142 nmolΠΑL) ΓΘΆ§ ± ,ΜΙΒΟΒΫΝΥ“ΜΗω«ΉΚœ–‘«Ω«“¥χ”–”ΪΙβΜν–‘ΒΡάύΥΤΓΣΓΣΓΣD2YpbFnLRFa(D2Tyr2(p2benzoyl2Phe)2norLeu2Arg2Phe2NH2 , Ki = 0123 nmolΠΑL)[61] ΓΘ
‘Ύ¥ΥΙΙ–ß―–ΨΩΒΡΙΛΉςΜυ¥Γ…œ ,Ά§ ±”Ο Pro1 ΚΆPhe2 ά¥ΧφΜΜ FMRFa ΕχΜώΒΟΒΡΥΡκΡΫαΙΙ PFRFa ΈΣNPFF ήΧεΒΡΦΌΕ®ΦΛΕ·ΦΝ ,άϊ”ΟΫαΙΙœό÷Τ–‘ΒΡ Tic ά¥ΧφΜΜ’βΗωΦΛΕ·ΦΝ 4 ΈΜΒΡ Phe ΒΟΒΫΝΥ“ΜΗω–¬ΒΡ NPFFάύΥΤΈο PFR(Tic)2NH2 ,≤Δ«“¥ΥάύΥΤΈο‘Ύ¥σ σ’ϊΧε―Σ―ΙΒΡ≤βΕ®÷–Ρή Ι¥σ σΤΫΨυΕ·¬ω―Ιœ¬ΫΒ ,Υϋ‘ΎΑΔΤ§≥…ώΪ Β―ι÷–±μœ÷≥ω»θόΉΩΙΦΝΒΡΜν–‘[62 ,63 ] ΓΘΒΪ «ΉνΫϋΒΡάκΧεΙΠΡή≤βΕ® Β―ι÷–ΖΔœ÷ , PFR ( Tic)-NH2 ±μœ÷ΈΣNPFF1 ΚΆ NPFF2 ήΧεΒΡ“ΜΗωΓΑ≥§ΦΛΕ·ΦΝΓ±[64 ,65 ] ΓΘΈΣΝΥΫχ“Μ≤ΫΤάΦέ PFRFa ΒΡΫαΙΙΗΡ±δΕ‘Τδ–Ρ―ΣΙήΜν–‘ΒΡ”Αœλ ,Έ“Ο«”Ο“ΜœΒΝ– Phe ΒΡάύΥΤΈοά¥ΧφΜΜΤδ Phe4,“©άμ―ßΦλ≤βΖΔœ÷ C Ρ©ΕΥ Phe ≤ύΝ¥ΒΡΩ’Φδ…λ’ΙΖΫœρΚΆΖΦœψΜΖ…œΒΡΒγΉ”‘ΤΖ÷≤ΦΒΡΗΡ±δΕΦΕ‘Τδ–Ρ―ΣΙήΒςΫΎΉς”Ο≤ζ…ζ“ΜΕ®ΒΡ”Αœλ(¥ΐΖΔ±μΒΡ Β―ιΫαΙϊ) ΓΘ
3 ΓΓ…ώΨ≠κΡ FF ήΧεόΉΩΙΦΝΒΡ―–ΨΩΫχ’Ι
Ά®ΙΐΙΙ–ßΙΊœΒ―–ΨΩœ»ΚσΖΔœ÷ΝΥ“Μ–© NPFF ήΧεΒΡΦΛΕ·ΦΝ ,Εχ NPFF ήΧεόΉΩΙΦΝΒΡ―–ΨΩΫχ’ΙœύΕ‘ΜΚ¬ΐΓΘΉνΫϋ NPFF ήΧεΒΡ―Γ‘ώ–‘όΉΩΙΦΝ RF9 ΒΡΖΔœ÷ Ι¥ΥΖΫΟφΒΡ―–ΨΩΙΛΉς»ΓΒΟΝΥΆΜΤΤ[66 ], ΒΪ « RF9 Ε‘NPFF1 ΚΆ NPFF2 ήΧεΒΡ―Γ‘ώ–‘≤Μ«Ω ,“ρ¥Υ ,ΤυΫώΈΣ÷Ι»‘»±ΖΠ NPFF1 ΚΆ NPFF2 ήΧεΒΡΉ®“Μ–‘ΒΡόΉΩΙΦΝΓΘ‘Ύ¥ΥΫΪ NPFF ήΧεόΉΩΙΦΝΒΡΖΔ’ΙΉς“ΜΦρ ωΓΘ“―±®ΒάΒΡ NPFF ήΧεΒΡόΉΩΙΦΝΜρΦΌΕ®όΉΩΙΦΝ÷ς“ΣΖ÷ΈΣκΡάύΚΆΖ«κΡάύΝΫ¥σάύΓΘ
NPFF ήΧεΒΡκΡάύόΉΩΙΦΝΕΦ «Μυ”ΎΤδΡΗΧεΜρΡΗΧεΒΡ C Ρ©ΕΥΥΡκΡΒΡ–ρΝ–ΫαΙΙΕχ―ή…ζ≥ωά¥ΒΡ ,ΒΪ «“―±®ΒάΒΡ’βάύόΉΩΙΦΝΕΦΕ‘ NPFF ΒΡΓΑ–≈ Ι”ρΓ±ΉωΝΥ“ΜΕ®ΒΡΜ·―ß–ό Έ ,ΥϋΟ«”κ NPFF ήΧεΒΡ«ΉΚœΝΠ“ΜΑψΕΦΫœΒΆ ,≤Δ«“’β–©κΡάύόΉΩΙΦΝΕΦ «Ά®Ιΐ“©άμ―ßΜν–‘ Β―ι…Η―Γ≥ωΒΡ ,ΕΦ»±ΖΠ…ζΜ·ΦχΕ®ΒΡ ΐΨί÷ß≥÷ΓΘ
‘γΤΎΒΡ―–ΨΩ±μΟς ,NPFF ΒΡ C Ρ©ΕΥΒΡ RF2NH2 «ΨωΕ® NPFF ΦΛΕ·Μν–‘ΒΡΙΊΦϋΈΜΒψ ,“ρ¥Υ―–ΨΩ»Υ‘±…ηΦΤκΡάύόΉΩΙΦΝΒΡΥΦ¬Ζ÷ς“Σ «Ά®Ιΐ…ΨΒτ NPFF ΦΑΤδΦΛΕ·ΦΝΒΡ C Ρ©ΕΥΒΡ Phe Εχ ΙΥϋΟ«¥”ΦΛΕ·ΦΝΉΣΜ·≥…όΉΩΙΦΝΓΘΉν‘γ±®ΒάΒΡ NPFF ΒΡΦΌΕ®όΉΩΙΦΝΈΣ daY8Ra(desaminotyrosyl2Phe2Leu2Phe2Gln2Pro2Gln2Arg2NH2) ,ΤδΕ‘”ΠΒΡΦΛΕ·ΦΝΈΣ daY9Fa ,Ϋχ“Μ≤ΫΒΡ“©άμ―ßΜν–‘Φλ≤βΖΔœ÷Υϋ‘Ύ¥σ σΦΙΥηœΗΑϊΡΛ…œΒΡ«ΉΚœΝΠ Ki =840 nmolΠΑL ,≤Δ«“ΥϋΡήΦθ»θ¬πΖ»ΒΡΡΆ ή ,Φθ«αΗΏΦΝΝΩΒΡNPFF ‘Ύ’ΐ≥Θ¥σ σ…œ“ΐΤπΑΔΤ§≥…ώΪΫδΕœ―υΖ¥”Π[67 ΓΣ69 ] ΓΘΉνΫϋΒΡ―–ΨΩΜΙΖΔœ÷Ψ≤¬ωΉΔ…δ daY8Ra Ε‘¥σ σΒΡΤΫΨυΕ·¬ω―ΙΈό”Αœλ ,ΒΪΥϋΡή”––ßΒΊΦθ»θ NPFFΥυ“ΐΤπΒΡ…ΐ―ΙΉς”Ο[70 ] ΓΘ‘ΎΥφΚσΒΡΙΙ–ß―–ΨΩΙΛΉς÷–”÷ΖΔœ÷÷§»ή–‘ΒΡάύΥΤΈο dansyl2PQRF2NH2 ΨΏ”– NPFF―υΒΡΦΛΕ·Μν–‘ΓΘ‘Ύ daY8Ra ΒΡ…ηΦΤΥΦ¬ΖΒΡΤτ Ψœ¬ ,…ηΦΤ≤ΔΚœ≥…ΝΥ NPFF ήΧεΒΡΩ…ΡήΒΡ÷§»ή–‘ΦΌΕ®όΉΩΙΦΝdansyl2PQR2NH2[71 ] ΓΘ‘ΎΥφΚσΒΡΜν–‘Φλ≤β÷–ΖΔœ÷ΥϋΡή”κ NPFF ήΧε”–“ΜΕ®ΒΡΫαΚœ ( Ki = 1316ΠΧmolΠΑL) ,≤Δ«“Υϋ‘ΎΑΔΤ§≥…ώΪΚΆΜΖΨ≥ΈΜ÷ΟΤΪΑ°(CPP) Β―ι÷–±μœ÷≥ωόΉΩΙΦΝΒΡΜν–‘[63 ,72 ] ΓΘΉν–¬ΒΡ±®ΒάΜΙΖΔœ÷Ψ≤¬ωΉΔ…δdansyl2PQR2NH2 Ρή“ΐΤπ“ΜΟςœ‘ΒΡΫΒ―ΙΉς”Ο ,≤Δ«“ΥϋΡήΦθ»θ NPFF Υυ“ΐΤπΒΡ…ΐ―ΙΉς”Ο[59 ,70 ] ΓΘ“‘ dansyl2PQR2NH2 ΈΣΜ·―ßΡΘΑε ,άϊ”ΟΉιΚœΜ·―ßΒΡΖΫΖ®Ά®Ιΐ±ΘΝτ Arg ≤–ΜυΕχΖ÷±πΧφΜΜ Pro ΚΆ Glu ≤–Μυ ,¥”Μ·ΚœΈοΩβ…Η―Γ”≈Μ·ΒΟΒΫ“ΜΗω«ΉΚœ–‘ΗϋΗΏΒΡάύΥΤΈο ΓΣΓΣΓΣdansyl2GSR2NH2 ( Ki = 114ΠΧmolΠΑL)[68 ] ΓΘ
NPFF ήΧεΖ«κΡάύόΉΩΙΦΝΕΦ «‘Ύ NPFF ΒΡ C Ρ©ΕΥ RF2NH2 ΒΡΜυ¥Γ…œΆ®ΙΐΜ·―ß–ό ΈΕχΒΟΒΫΒΡ ,ΡΩ«Α“―±®ΒάΒΡ÷ς“Σ”–“‘œ¬œΒΝ– : (1) BIBP3226 ΦΑΤδ―ή…ζΈο ; (2) RF œΒΝ–Μ·ΚœΈο(ΆΦ 1) ΓΘ
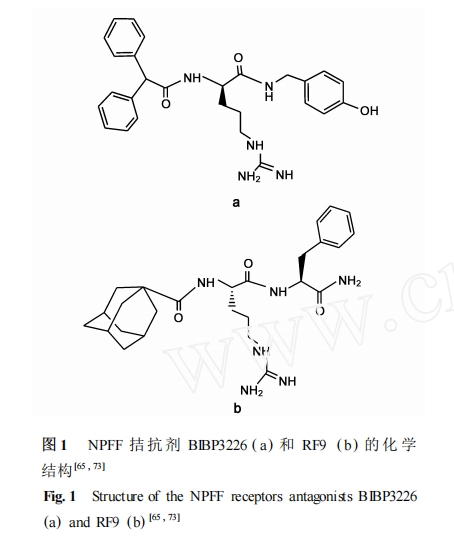
BIBP3226 Ήν≥θ «ΉςΈΣ NPY Y1 ήΧεΒΡ―Γ‘ώ–‘όΉΩΙΦΝΕχ±ΜΖΔœ÷[73 ,74 ],ΉνΫϋΒΡ―–ΨΩ±μΟςΥϋ“≤ΡήΫαΚœNPFF ήΧε ,ΧΊ±π « NPFF1 ήΧε ,≤Δ«“‘ΎΧεΆβΒΡΦλ≤β÷–±μœ÷≥ωόΉΩΙΜν–‘[45 ,49 ,64 ,75 ] ΓΘΈ“Ο«Ϋχ“Μ≤ΫΒΡ―–ΨΩΜΙΖΔœ÷ BIBP3226 ‘Ύ“ΜœΒΝ–ΒΡΧεΆβΚΆΧεΡΎ“©άμ―ßΜν–‘ Β―ι÷–ΕΦΡήόΉΩΙ NPFF ΚΆ NPVF Υυ“ΐΤπΒΡΦΛΕ·Μν–‘[76 ,77 ] ΓΘ”–»ΛΒΡ « ,NPY ήΧεΒΡ“Μ–©ΤδΥϋΖ«κΡάύ≈δΧε“≤Ρή”––ßΒΊΫαΚœ NPFF ήΧε ,»γ NPY Y1 ήΧεΒΡ―Γ‘ώ–‘όΉΩΙΦΝ BIBO3304 ΚΆ NPY Y1 ήΧεόΉΩΙΦΝ/Y4 ήΧεΦΛΕ·ΦΝ GR231118 , ≤Δ”–±®Βά±μΟς BIBO3304 ΚΆGR231118 ‘ΎΧεΆβΒΡΙΠΡή Β―ι÷–Ζ÷±π±μœ÷ΈΣ NPFF ήΧεΒΡόΉΩΙΦΝΚΆΦΛΕ·ΦΝ[45 ,75 ] ΓΘ¥ΥΆβ ,ΈΣΝΥΫχ“Μ≤Ϋ”≈Μ·BIBP3226 ά¥―Α’“ NPFF ήΧεΒΡ―Γ‘ώ–‘όΉΩΙΦΝ ,…ηΦΤΚœ≥…ΝΥ“ΜœΒΝ– BIBP3226 άύΥΤΈο ,Υδ»Μ≤ΩΖ÷άύΥΤΈοΕ‘NPFF1 ΚΆ NPFF2 ήΧεΨΏ”–“ΜΕ®ΒΡ―Γ‘ώ–‘ ,ΒΪ“≈ΚΕΒΡ « ,ΥϋΟ«Ά§ ±“≤”κ NPY Y1 ήΧεΫαΚœ¥”Εχœό÷ΤΝΥΥϋΟ«‘Ύ“©άμ―ßΙΠΡήΦχΕ®÷–ΒΡΙψΖΚ Ι”Ο[45 ] ΓΘ
ΉνΫϋΒΡ±®Βά±μΟς ,ΗυΨί NPFF ΒΡ C Ρ©ΕΥΒΡΙΊΦϋΈΜΒψ RF2NH2 ΫαΙΙ ,“‘ RF2NH2 ΈΣΡΘΑε ,Ά®ΙΐΤδ N ΕΥΝ§Ϋ”≤ΜΆ§ΒΡ―ή…ζΜυΆ≈ΕχΜώΒΟΝΥΫϋΑΌΗωΖ«κΡάύΒΡΜ·ΚœΈο ,≤ΔΆ®ΙΐΜν–‘Φλ≤βΖΔœ÷’β–©άύΥΤΈο÷–ΒΡ RF2ΓΔRF9ΚΆ RF48 Ε‘”Ύ NPFF ήΧε”–ΫœΚΟΒΡ«ΉΚœΝΠ[65 ] ΓΘRF9Ε‘”Ύ NPFF1 ΚΆ NPFF2 ήΧεΒΡ«ΉΚœΝΠΉνΗΏ ,Εχ«“ΤδΕ‘”ΎΑΔΤ§ ήΧεΚΆΤδΥϊΒΡ RF Φ“ΉεœύΙΊ ήΧεΕΦΟΜ”–ΫαΚœ ,“ρ¥Υ RF9 «“ΜΗωΕ‘”Ύ NPFF ήΧεΨΏ”–―Γ‘ώ–‘ΒΡΗΏ–ßόΉΩΙΦΝΓΘ‘ΎΧεΒΡ―–ΨΩ±μΟς ,RF9 ±Ψ…μ≤Μ”Αœλ¥σ σΒΡ―Σ―ΙΚΆ–Ρ¬ ,»¥Ω…“‘“÷÷Τ NPFF “ΐΤπΒΡ¥σ σΤΫΨυΕ·¬ω―ΙΚΆ–Ρ¬ …ΐΗΏ ,Υυ“‘ NPFF Ε‘”Ύ―Σ―ΙΚΆ–Ρ¬ ΒΡΒςΫΎ «÷±Ϋ”Ά®ΙΐΤδ ήΧεΉς”ΟΒΡΓΘ Ι”Ο RF9 ±Ψ…μ≤Μ”Αœλ¥σ σΒΡΜυ¥ΓΆ¥ψ– ,Εχ RF9 Ω…“‘Ά®ΙΐΉηΕœ NPFFΒΡ ήΧε”––ßΒΊ“÷÷Τ≥ΛΤΎΉΔ…δΚΘ¬ε“ρ“ΐΤπΒΡΆ¥ΟτΚΆΡΆ ή[65 ] ΓΘ“ρ¥Υ ,NPFF ήΧεΗΏ–ßόΉΩΙΦΝΒΡΖΔœ÷Ε‘”ΎΧαΗΏ¬πΖ»÷ΈΝΤ¬ΐ–‘Ά¥ΒΡ–ßΙϊ”–÷Ί“ΣΒΡ“©”ΟΦέ÷ΒΓΘΈ“Ο«ΜΙΖΔœ÷ RF9 Ρή”––ßΒΊόΉΩΙ NPFF ‘ΎΧεΈ¬ΚΆΆ¥ΨθΒςΫΎ÷–ΒΡΦΛΕ·Μν–‘(¥ΐΖΔ±μΒΡ Β―ιΫαΙϊ) ΓΘ
¥ΥΆβ ,“©ΈοΙΪΥΨάϊ”Ο“Μ–©”–Μζ–ΓΖ÷Ή”ά¥ΡΘΡβNPFF ΒΡ C Ρ©ΕΥΜ·―ßΫαΙΙ ,¥”ΕχΖΔœ÷ΝΥ≤ΩΖ÷”–Μζ–ΓΖ÷Ή”“≤ΨΏ”–ΫœΚΟΒΡ NPFF όΉΩΙΜν–‘[55 ] ΓΘ
4 ΓΓΫα”ο
ΥφΉ≈ΫϋΡξά¥ΡΎ‘¥–‘ΑΔΤ§ΒςΫΎκΡ‘ΎΑΔΤ§ΒΡΡΆ ήΚΆ“άάΒ÷–ΒΡΉς”Ο‘Ϋά¥‘Ϋ ήΒΫ»ΥΟ«ΒΡΙΊΉΔ ,NPFF “©άμ―ßΙΠΡήΒΡ―–ΨΩ“≤±Η ή÷Ί ”ΓΘ”κ¥ΥΆ§ ± ,ΉνΫϋΦΗΡξά¥RFa κΡΦ“Ήε–¬≥…‘±ΒΡ≥ωœ÷ ,RFa κΡœύΙΊΒΡ¬έΈΡ“≤≤ΜΕœ‘ωΕύ , ΧΊ±π «»ΞΡξΙζΦ ΕύκΡΝλ”ρΒΡ÷ΣΟϊΩ·ΈοPeptides Ή®Ο≈≥ωΑφΝΥ“ΜΤΎ“‘ RFa κΡΈΣ÷ςΧβΒΡΧΊΩ· ,’β“≤Ηχ NPFF ΒΡ―–ΨΩ¥χΝΥ–¬ΒΡΖΔ’ΙΜζ”ωΓΘΫ“ Ψ NPFF1ΚΆ NPFF2 ήΧεΠΑNPFF œΒΆ≥ΒΡ“©άμ―ßΙΠΡήΦΑΤδΉς”ΟΜζ÷Τ±ΊΫΪ≥…ΈΣ NPFF ΦΑ RFa κΡ―–ΨΩΒΡ»»ΒψΈ Χβ÷°“ΜΓΘΖΔ’Ι NPFF1 ΚΆ NPFF2 ήΧεΒΡ―Γ‘ώ–‘όΉΩΙΦΝΚΆΦΛΕ·ΦΝ ,Ε‘”Ύ¥ΌΫχ NPFF ΒΡ―–ΨΩΨΏ”– °Ζ÷÷Ί“ΣΒΡ“β“εΓΘ
άϊ”ΟΕύκΡΜ·―ß…ζΈο―ßΒ»―–ΨΩ ÷ΕΈΫχ“Μ≤ΫœΒΆ≥ΒΊ―–ΨΩ NPFF ΒΡΙΙ–ßΙΊœΒΕ‘”ΎΧΫΧ÷ NPFF ΒΡΫαΙΙ”κΙΠΡή÷°ΦδΒΡΙΊœΒΨΏ”–÷Ί“ΣΒΡ¥ΌΫχΉς”Ο ,’βΫΪΈΣ NPFF ήΧεΒΡ―Γ‘ώ–‘ΦΛΕ·ΦΝΚΆόΉΩΙΦΝΒΡ…ηΦΤΧαΙ©άμ¬έ“άΨί ,≤ΔΫχ“Μ≤ΫΈΣ NPFF œύΙΊκΡΦΑΤδ ήΧεΒΡ―–ΨΩΧαΙ©”––ßΒΡ“©άμ―ßΙΛΨΏΓΘ¥ΥΆβ ,…ηΦΤΓΔ”≈Μ·ΚΆ…Η―Γ NPFF1ΚΆ NPFF2 ήΧεΉ®“Μ–‘ΒΡ”–Μζ–ΓΖ÷Ή”≈δΧε“≤ «―Α’“”––ßΒΡ NPFF “©άμ―ß―–ΨΩΙΛΨΏΒΡΫίΨΕ ,≤Δ«“’β–©ΗΏ–ßΒΡ≈δΧεΕΦΨΏ”–«±‘ΎΒΡΝΌ¥≤”Π”ΟΦέ÷ΒΓΘ
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷ΨΓΘ
