
’Σ “Σ ”…”Ύ‘ΎΒΑΑΉ÷ Ζ÷άκΓΔ…ζΈο¥ΪΗ–ΤςΓΔΦ≤≤Γ’οΕœΚΆ÷ΈΝΤ“‘ΦΑΜυ¥ΓΒΡ…ζΈο“Ϋ“©―–ΨΩΒ»Νλ”ρΒΡ«±‘Ύ÷Ί“Σ”Π”Ο ,Κœ≥… ήΧε”κΕΧκΡ(3 ΓΣ20 Α±ΜυΥα≤–Μυ) œύΜΞΉς”ΟΒΡ―–ΨΩ“―“ΐΤπΝΥ»ΥΟ«ΒΡΙΊΉΔΓΘ±ΨΈΡΉέ ωΝΥΫϋΡξά¥Κœ≥… ήΧε”κΕΧκΡœύΜΞΉς”ΟΒΡ―–ΨΩΫχ’Ι ,Ϋι…ήΝΥ–ΓΖ÷Ή” ήΧε”κΗω±πΑ±ΜυΥα≤–ΜυΒΡœύΜΞΉς”Ο ,–ΓΖ÷Ή” ήΧεΕ‘ΕΧκΡΒΡ–ρΝ–―Γ‘ώ–‘ Ε±π ,–ΓΖ÷Ή” ήΧεΕ‘ΕΧκΡΕΰΦΕΫαΙΙΒΡ”’ΒΦ”κ Ε±π“‘ΦΑΗΏΖ÷Ή” ήΧε”κΕΧκΡΒΡœύΜΞΉς”Ο ,≤ΔΕ‘ΕΧκΡ ήΧεΒΡ…ηΦΤΫχ––ΝΥΫ®“ι”κ’ΙΆϊΓΘ
‘ΎΚή≥ΛΒΡ“ΜΕΈ ±Φδάο ,»ΥΟ«Ε‘”ΎΒΑΑΉ÷ -ΒΑΑΉ÷ œύΜΞΉς”Ο Φ÷’¥φ‘Ύ’β―υ“Μ÷÷»œ Ε ,Φ¥ΟΗ-ΒΉΈοΦΑΤδ“÷÷ΤΦΝΓΔΩΙ‘≠-ΩΙΧεΓΔΦΛΥΊ- ήΧεΒ»ΒΑΑΉ÷ Η¥ΚœΧεΒΡ–Έ≥… , «œύΜΞΉς”ΟΒΡΝΫΗωΖ÷Ή”Φδ’ϊΗωΜΞ≤ΙΫγΟφΒΡœύΜΞΉς”Ο , «’ϊΧεΩ’ΦδΙΙœσΒΡ Ε±π”κΤθΚœΓΘ“≤ΨΆ «ΥΒ ,ΝΫ’ΏΒΡΫαΚœ…φΦΑΫœ¥σΒΡ“ΜΗωΩ’Φδ ;≤Έ”κΫαΚœΒΡΑ±ΜυΥα≤–Μυ≤Μ «œΏ–‘κΡΕΈ ,Εχ «‘Ύ“ΜΦΕΫαΙΙ…œœύΨύΫœ‘Ε ,ΒΪ”÷Ι≤Ά§ΙΙ≥…“ΜΗω Κœ”ΎΫαΚœΒΡΝΔΧεΫαΙΙΓΘ’β÷÷ΙέΒψ‘ΎΚήΕύΒΡΒΑΑΉ÷ Η¥ΚœΧε÷–ΒΟΒΫ÷Λ Β[1 ΓΣ3 ] ΓΘΥφΉ≈Ε‘ΒΑΑΉ÷ -ΒΑΑΉ÷ ΫγΟφ―–ΨΩΒΡ…ν»κ ,ΚήΕύΈΡœΉ”÷±®ΒάΝΥΒΑΑΉ÷ œύΜΞΉς”Ο÷–ΒΡΫαΙΙ[4 ΓΣ8 ] ΓΔΡήΝΩ[6 , 9 ] ΓΔΒγ–‘ΜΞ≤Ι[10 ]ΦΑΕ·ΝΠ―ßΒ»ΖΫΟφΒΡ―–ΨΩΫαΙϊ ,ΈΣ…ν»κ―–ΨΩΒΑΑΉ÷ -ΒΑΑΉ÷ œύΜΞΉς”Ο¥ρœ¬ΝΥΜυ¥ΓΓΘ
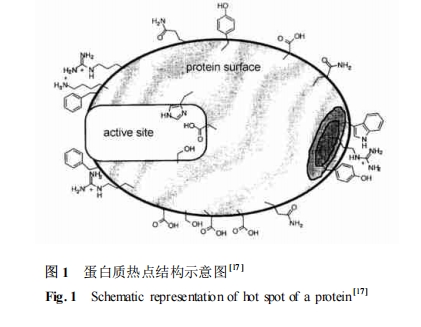
ΫϋΡξά¥ΒΑΑΉ÷ ΦδœύΜΞΉς”Ο―–ΨΩΒΡ“ΜΗω÷Ί“ΣΆΜΤΤ‘Ύ”ΎΒΑΑΉ÷ ±μΟφΓΑ»»Βψ ( hot spots) Γ±ΒΡ»ΖΕ® ,ΥυΈΫΒΡΓΑ»»ΒψΓ± «ΈΜ”ΎΜρΫ”ΫϋΒΑΑΉ÷ -ΒΑΑΉ÷ ΫγΟφΦΗΚΈ÷––ΡΒΡ“ΜΗω‘Φ 600A2 ΒΡ«χ”ρ ,»γΆΦ 1 Υυ ΨΓΘΙΙ≥…ΓΑ»»ΒψΓ±ΒΡ≤–ΜυΕ‘ΒΑΑΉ÷ -ΒΑΑΉ÷ Η¥ΚœΈοΒΡΈ»Ε®–‘Τπ÷Ί“ΣΒΡΉς”Ο[11 ΓΣ14 ] ΓΘWells Β»ΖΔœ÷ ,ΗΡ±δ»»Βψ«χ”ρΡΎΡ≥–©Α±ΜυΥα≤–ΜυΕ‘ΒΑΑΉ÷ ΒΡ«ΉΚΆ––ΈΣ”ΑœλΫœ¥σ ,Εχ»»Βψ÷ήΈßΒΡ≤–Μυ‘ρΕ‘ΒΑΑΉ÷ Η¥ΚœΈοΒΡΈ»Ε®–‘Ι±œΉΫœ–Γ[15 ] ΓΘ―–ΨΩΜΙΖΔœ÷‘Ύ»»Βψ«χ”ρΡΎΤπΉς”ΟΒΡ≤–Μυ÷– Trp (…ΪΑ±Υα) ΓΔPhe (±Ϋ±ϊΑ±Υα) “‘ΦΑ Arg(ΨΪΑ±Υα) 3 ΗωΑ±ΜυΥαΕ‘”ΎΫαΚœΡήΒΡΙ±œΉΉν¥σ ,’β”κ Geysen ΒΡ―–ΨΩΫαΙϊ[16 ] “Μ÷¬ΓΘGeysen »œΈΣ ,ΫαΚœΒΡΝΫΗωΖ÷Ή”Φδ…Ό ΐΦΗΗωΙΊΦϋΜυΆ≈ΒΡ»θœύΜΞΉς”ΟΧαΙ©ΝΥΨχ¥σ≤ΩΖ÷ΒΡΫαΚœΡή ;Κ§”–”κΫαΚœ”–ΙΊΒΡΦΗΗωΙΊΦϋΜυΆ≈ΒΡœΏ–‘ΕΧκΡΩ…ΡΘΡβΒΑΑΉ÷ ’έΒΰΒΡΩΙ‘≠ΨωΕ®¥ΊΓΘ
2 –ΓΖ÷Ή” ήΧε”κΕΧκΡœύΜΞΉς”Ο
ΡΩ«Α ,‘Ύ―–ΨΩΚœ≥… ήΧε”κΕΧκΡœύΜΞΉς”ΟΒΡΖ÷Ή”…ηΦΤ÷– ,÷ς“ΣΑϋά®ΦΗΗωΖΫΟφ : (1) ’κΕ‘ΧΊΕ®ΒΡΑ±ΜυΥα≤–Μυ ,»γ His(ΉιΑ±Υα) ΓΔArgΓΔTrp Β» ; (2) ’κΕ‘“Μάύ–‘÷ œύΫϋΒΡΑ±ΜυΥα ,»γΦν–‘Α±ΜυΥα :Lys(άΒΑ±Υα) ,Arg ;ΜρΥα–‘Α±ΜυΥα :Asp (ΧλΕ§Α±Υα) ΓΔGlu (Ι»Α±Υα) Β» ; (3) ’κΕ‘Α±ΜυΥαΒΡΕΥΜυ ,Φ¥ N ΕΥ―τάκΉ”Μρ C ΕΥ“θάκΉ” ; (4)’κΕ‘ΧΊΕ®ΒΡΕύκΡΗΡ–‘ΜυΆ≈ ,»γΝΉΥαΜ·ΜυΆ≈ ; (5) ’κΕ‘“ΜΗωΧΊΕ®ΒΡ–ρΝ– ,’β“≤ «ΡΩ«Α―–ΨΩΒΡ»»Βψ ,ΒΪ…φΦΑΒΡ–ρΝ–ΆυΆυΚήΕΧ ,ΚήΕύ―–ΨΩΕΦ…Ό”Ύ 3 ΗωΑ±ΜυΥα≤–ΜυΒΡΕΧκΡ ,ΉνΈΣΤ’±ιΒΡ «ΝΫΗωΑ±ΜυΥα≤–Μυ ,Μρ’ΏΕύκΡ“ΜΆΖ“ΜΈ≤ΒΡΑ±ΜυΥα≤–Μυ ; (6) ’κΕ‘ΧΊΕ®ΕύκΡΙΙœσΒΡ Ε±π ,Φ¥ΠΝ¬ί–ΐΚΆΠ¬’έΒΰΫαΙΙ ,’βάύ―–ΨΩΆυΆυΦ·÷–”ΎΚœ≥… ήΧε”κκΡΒΡ»θœύΜΞΉς”Ο”’ΒΦΕΰΦΕΫαΙΙΒΡ≤ζ…ζΓΘ
2. 1 –ΓΖ÷Ή” ήΧε”κΗω±πΑ±ΜυΥα≤–ΜυœύΜΞΉς”Ο
’κΕ‘ΕύκΡΧΊΕ®ΒΡΑ±ΜυΥα≤–Μυ(ΕΥΜυΓΔ≤ύΝ¥ΚΆΝΉΥαΜ·≤–Μυ) …ηΦΤœύ”ΠΒΡ ήΧε , ΙΤδ”κΕύκΡΖΔ…ζ―Γ‘ώ–‘œύΜΞΉς”Ο «ΕύκΡ ήΧε…ηΦΤ÷–“ΜΗω÷Ί“ΣΖΫΖ®ΓΘΆ®ΙΐΫΪΡ≥–©Ρή”κΧΊΕ®Α±ΜυΥα≤–ΜυΖΔ…ζ»θœύΜΞΉς”ΟΒΡ–ΓΖ÷Ή”(ΜΖΚΐΨΪΓΔΑ±Μυ»ΐ““Υα(NTA) ΓΔΏ≤ΏχΒ») Ϋχ–––ό ΈΚΆΗΡ–‘ ,ΡΩ«Α“―Ψ≠ΖΔ’ΙΝΥΚήΕύ¥ΥάύΒΡ ήΧεΓΘ
Νθ”ΐΒ»[18 ] Ά®ΙΐΉœΆβΓΔ‘≤Εΰ…ΪΓΔ”ΪΙβΓΔΕΰΈ§ΚΥ¥≈ΖΫΖ® ,‘Ύ Tris-HCl ΜΚ≥ε“Κ÷–―–ΨΩΝΥ«≈Ν§ΥΪΜΖΚΐΨΪ (ΆΦ2a) ‘ΎΦ”»κΫπ τάκΉ”«ΑΚσ”κœΒΝ–÷§ΖΨΉεΕΧκΡ Leu-Gly ,Gly-Leu , Gly-Pro , Glu-Glu , Gly-Gly , Gly-Gly-Gly “‘ΦΑ Glu-Cys-Gly ΒΡΑϋΫα––ΈΣ ,¥”÷ς-ΩΆΧεΦδ”’ΒΦΤθΚœΚΆ Ε±πΜζάμΖΫΟφ―–ΨΩΝΥ―Γ‘ώ–‘ΦΑΦϋΚΆΡήΝΠΓΘΫαΙϊ±μΟς ,«≈ΝΣΥΪΜΖΚΐΨΪ ήΧεΕ‘ Glu-Cys-Gly ΨΏ”–Οςœ‘ΒΡ―Γ‘ώ–‘ ,«ΉΚΆ≥Θ ΐ « Glu-Glu ΒΡ 51 ±Ε ,ΥΒΟςΥυΚœ≥…ΒΡ«≈Ν§ΥΪΜΖΚΐΨΪ ήΧεΡή”––ß Ε±πΕΧκΡΒΡ≥Ώ¥γΚΆ ηΥ°–‘ΓΘΕχ‘Ύ¬γΚœ Ni2 + ÷°Κσ ,Ϋπ τ«≈ΝΣΥΪΜΖΚΐΨΪΕ‘ Glu-GluΨΏ”–ΉνΗΏΒΡΫαΚœ≥Θ ΐ ,¥οΒΫ 6182 ΓΝ104 M- 1,Ϋπ τάκΉ”ΒΡ≤Έ”κ”––ßΒΊΧαΗΏΝΥ«≈ΝΣΥΪΜΖΚΐΨΪΒΡΦϋΚΆΡήΝΠΚΆ―Γ‘ώ–‘ΓΘ
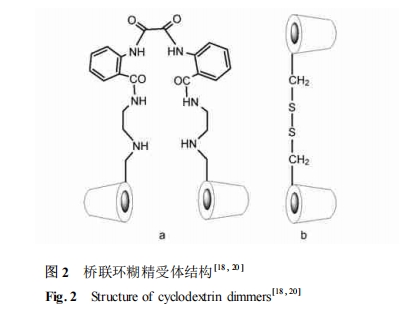
ΥΈά÷–¬Β»[19 ] …ηΦΤΝΥ”ΪΙβΥΊ±ξ ΕΒΡ (1 R ,3 R)-1-Α±Μυ-1 ,3-Εΰτ»ΜυΜΖΈλΆι―ή…ζΈο«≈ΝΣΒΡΥΪΜΖΚΐΨΪ ήΧε ,άϊ”Ο”ΪΙβΤΪ’ώΖΫΖ® ,―–ΨΩΤδ”κ“ΜœΒΝ–ΕΧκΡΒΡœύΜΞΉς”ΟΓΘ±»ΫœΝΥ÷ςΧεΖ÷Ή””κ“ΜΕ‘ ÷–‘“λΙΙΧεœύΜΞΉς”ΟΒΡΉ‘”…Ρή±δΜ·‘ωΝΩ“‘ΦΑΥϋΟ«÷°ΦδΒΡΫαΚœ≥Θ ΐ ,÷Λ ΒΜΖΚΐΨΪΕΰΨέΧεΕ‘ΕΧκΡΨΏ”– ÷–‘ Ε±πΡήΝΠ ,≤ΔΧΫΧ÷ΝΥΜΖΚΐΨΪΕΰΨέΧε”κΕΧκΡ÷°ΦδœύΜΞΉς”ΟΒΡλ -λΊ≤Ι≥Ξ––ΈΣΓΘ
Breslow Β»[20 ,21 ]Κœ≥…ΝΥΨΏ”–≤ΜΆ§ΦδΗτ±έΒΡ«≈ΝΣΜΖΚΐΨΪ ,≤ΔΖ÷±π―–ΨΩΝΥΥϋΟ«‘ΎΥ°»ή“Κ÷–”κΦΗ÷÷ΕΧκΡΒΡΫαΚœΡήΝΠΓΘΉœΆβΒΈΕ®ΫαΙϊœ‘ ΨΕΰΝρΦϋ«≈ΝΣΜΖΚΐΨΪ(ΆΦ 2b) Ε‘ Trp-Trp ΕΰκΡ±μœ÷≥ωΝΥΦΪ«ΩΒΡΫαΚœΡήΝΠ ,ΕχΕ‘ Phe-Phe ΕΰκΡ‘ρΟΜ”–ΧΊ“λ–‘ΫαΚœΓΘ
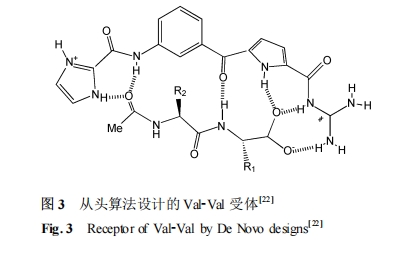
Υ°»ή“Κ÷–κΡΒΡ Ε±πΨΏ”–“ΜΕ®ΒΡΡ―Ε» ,“ΜΗω÷Ί“Σ‘≠“ρ ««βΦϋ ή»ήΦΝΗ…»≈ΚήΡ―ΖΔΜ”Ής”Ο ,Εχάϊ”ΟάκΉ”œύΜΞΉς”ΟΈ»Ε®«βΦϋ «“Μ÷÷…ηΦΤΥ°œύ ήΧεΒΡ”––ßΖΫΖ®ΓΘ2004 Ρξ Schmuck Β»[22 ] ≤…”Ο¥”ΆΖΥψΖ®…ηΦΤΝΥ“Μ÷÷–¬–ΆΒΡΕΰκΡ ήΧε(ΆΦ 3) ,Ω…‘ΎΥ°»ή“Κ÷–”––ß Ε±π CΕΥΈ¥±ΘΜΛΒΡΕΰκΡ ,ΫαΚœœΒ ΐ¥οΒΫ 10^4M- 1“‘…œΓΘΆ®Ιΐ‘Ύ ήΧε÷–“ΐ»κκ“τ Μυ ,”κΕΰκΡΒΡτ»Μυ–Έ≥…«βΦϋΚΆάκΉ”Ε‘ ,Ά§ ±ΕΰκΡ÷ςΝ¥Α±Μυ”κ ήΧε–Έ≥…ΒΡΤδΥϊ«βΦϋ“≤ΤπΒΫ‘ω«ΩΗ¥ΚœΈοΈ»Ε®–‘ΒΡΉς”ΟΓΘ‘Ύ¥ΥΜυ¥Γ…œ ,Ά®ΙΐΏδΏρΜΖΒΡ“ΐ»κ‘ωΦ”ΝΥΤ’Ά®κ“Μυ ήΧεΒΡ―Γ‘ώ–‘ΓΘΉœΆβ-Ω…ΦϊΒΈΕ®≤βΕ®Τδ‘ΎΥ°÷–”κ≤ΜΆ§ΒΉΈοΒΡΫαΚœ≥Θ ΐ( K) ,ΫαΙϊœ‘ Ψ K ÷Β :Val-Val > Ala-Ala > Gly-Gly ,’β“ΜΫαΙϊΫ“ ΨΑ±ΜυΥα≤ύΜυΒΡ ηΥ°ΜυΆ≈”κ ήΧεΦδΒΡ ηΥ°Ής”ΟΩ…“‘Ϋχ“Μ≤Ϋ‘ω«Ω«ΉΚΆ–‘ΓΘΗΟΖΫΖ®‘Ύάμ¬έΦΤΥψΒΡΜυ¥Γ…œΚœ≥…≥ω–¬ΒΡΗΏ–ßΒΡΕΰκΡ ήΧε ,”≈”ΎΒ± ±±®ΒάΒΡΤδΥϊΒΡΕΰκΡ ήΧεΓΘSchmuck ΩΈΧβΉι[23 ΓΣ26 ]ΥφΚσ”÷―–ΨΩΝΥΦΗ÷÷ΥΡκΡ”κΗΟάύ≈δΧεΒΡœύΜΞΉς”Ο ,“≤ΒΟΒΫΝΥ≤Μ¥μΒΡΫαΙϊΓΘ
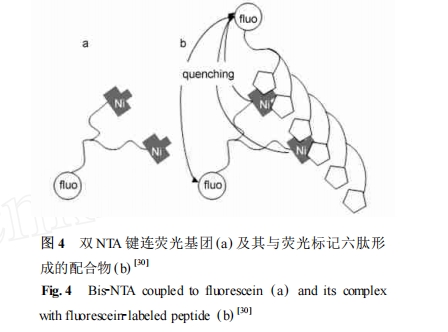
‘Ύ ήΧε…œ“ΐ»κΩ…–Έ≥…≈δΈΜΦϋΒΡΙΐΕ…Ϋπ τ‘ΣΥΊ ,”κΕύκΡ–ρΝ–÷–ΧΊΕ®≤–ΜυΜρΝΉΥαΜ·ΜυΆ≈–Έ≥…«ΩΒΡ≈δΈΜάκΉ”œύΜΞΉς”Ο ,“≤ « Βœ÷ ήΧε‘ΎΥ°œύ÷–Ε‘ΕύκΡ Ε±π”––ßΒΡΖΫΖ®[27 ΓΣ29 ] ΓΘPiehler Β»[30 ] ―–ΨΩΝΥΚ§ 2 ΓΣ4 ΗωNTA ΒΡ”ΪΙβΧΫ’κΜ·ΚœΈο”κΚ§ΕύΗω His ≤–ΜυΒΡ”ΪΙβΝυκΡΦδΒΡœύΜΞΉς”ΟΕχ“ΐΤπ”ΪΙββßΟπΒΡœ÷œσ (ΆΦ 4) ,÷ΛΟςΗΟΧΫ’κΜ·ΚœΈοΕ‘ His ±ξΦ«ΒΡκΡ”–ΫœΗΏΒΡ«ΉΚΆ–‘ΓΘ‘ΎΫπ τάκΉ”≈δΈΜœύΜΞΉς”Ο Ε±πΕύκΡΖΫΟφ ,Ώ≤ΏχΦΑΤδ―ή…ζΈοΉςΈΣ ήΧεΒΡ±®ΒάΚήΕύ[31 ΓΣ34 ],’βάο≤Μ‘Όœξ ωΓΘ
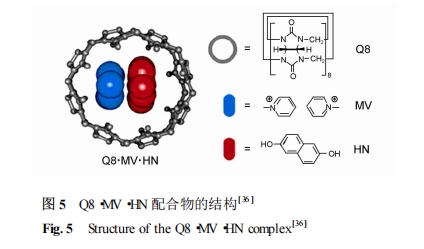
Ψ≠ΒδΒΡ“―ΈΣ»ΥΟ«Υυ λ÷ΣΒΡ»ΥΙΛ ήΧε»γΙΎΟ―ΓΔΜΖΖ§Β»»γΦ”“‘«…Ονάϊ”Ο“≤Ω…“‘…ηΦΤ≥ω–‘Ρή”≈ΝΦΒΡ ήΧε[35 ],ΆΦ 5 ΈΣ Urbach Β»[36 ] ±®ΒάΒΡΚœ≥… ήΧεΚυ¬Ϊκε(Q8) Ε‘ N ΕΥ…ΪΑ±ΥαΒΡ Ε±π ,Q8 Ω…“‘”κΦΉΜυΉœ¬όΦνMV ΦΑ“ΜΗωΖΦΜΖΩΆΧε–Έ≥…1ΓΟ1ΓΟ1–Ά≈δΚœΈοΓΘΥϊΟ«”ΟΒ»Έ¬ΒΈΕ®ΈΔΝΩ»»ΓΔ÷ ΤΉΓΔΉœΆβ-Ω…ΦϊΓΔ”ΪΙβΓΔ1H NMR ―–ΨΩΝΥ Q8ΓΛMV ”κ 4 ÷÷ΑϋΚ§…ΪΑ±ΥαΒΡ»ΐκΡΒΡœύΜΞΉς”Ο ,ΖΔœ÷ΤδΕ‘ Trp-Gly-Gly ΒΡΫαΚœ≥Θ ΐ (113 ΓΝ10^5) , « Gly-Trp-Gly ΒΡ 6 ±Ε , « Gly-Gly-Trp ΒΡ 40 ±ΕΓΘΖ÷Έω±μΟς ,’β÷÷≤ν“λ «”…Ώ≈ΏαΒΡΒγΚ…œύΜΞΉς”Ο≤ζ…ζ ,Μ·ΚœΈο–Έ≥…ΑιΥφΉ≈ΒγΚ…ΉΣ“ΤΚΆΏ≈Ώα”ΪΙβΒΡ±δΜ· ,’β–©Ιβ―ß–‘÷ “‘ΦΑΧεœΒΒΡΈ»Ε®–‘ΚΆ―Γ‘ώ–‘‘ΎΧΫ≤βΚΆΖ÷άκΧΊΕ®ΒΡκΡΖΫΟφΨΏ”–“ΜΕ®ΒΡ”Π”Ο«ΑΨΑΓΘ
ΒΑΑΉ÷ ΝΉΥαΜ· «…ζΈοΫγΉνΤ’±ι ,“≤ «Ήν÷Ί“ΣΒΡ“Μ÷÷ΒΑΑΉ÷ Ζ≠“κΚσ–ό Έ[37 ] ΓΘΩ…ΡφΒΡΝΉΥαΜ·Ιΐ≥ΧΦΗΚθ…φΦΑΥυ”–ΒΡ…ζάμΦΑ≤ΓάμΙΐ≥Χ ,»γœΗΑϊ–≈Κ≈¥ΪΒΦΓΔ–¬≥¬¥ζ–Μ“‘ΦΑœΗΑϊΒΡ‘ω÷≥ΓΔΖΔ”ΐΚΆΖ÷Μ·Β»ΓΘάΐ»γ ,ά“ΒΑΑΉΝΉΥακΡ (CPP) «“ΜάύΗΜΚ§ΝΉΥαΥΩΑ±Υα…ζΈοΜν–‘κΡ ,Ζ÷≤Φ”Ύ≈Θ»ιά“ΒΑΑΉΒΡ≤ΜΆ§«χ”ρ ,ΤδΜν–‘÷––Ρ «Ν§–χΒΡΝΉΥαΜ·ΥΩΑ±Υα≤–ΜυΚΆΙ»Α±Υα≤–Μυ ,Μυ±ΨΫαΙΙΩ…±μ ΨΈΣ :pSer-pSer-pSer-Glu-GluΓΘCPP ‘Ύ÷––‘ΚΆΦν–‘Ή¥Χ§œ¬Ω…ΉςΈΣΈόΜζάκΉ”‘ΊΧε ,Ά®ΙΐΝΉΥαΜ·ΥΩΑ±Υα”κΗΤΓΔ–ΩΓΔΧζΒ»άκΉ”ΫαΚœ ,–Έ≥…Ω…»ή–‘Η¥ΚœΈο ,”…–Γ≥Π±ΎœΗΑϊΈϋ ’Κσ‘Ό ΆΖ≈≥ωά¥ ,¥”Εχ”––ß±ήΟβΝΥΤδ‘Ύ–Γ≥Π÷–±Μ≥ΝΒμ ,¥ΌΫχΝΥ’β–©άκΉ”ΒΡΈϋ ’[38 ] ΓΘ
”…”Ύ’κΕ‘ΝΉΥαΜ·ΒΑΑΉΒΡΖ÷άκΚΆΖ÷Έω»±…ΌΝ°ΦέΗΏ–ßΒΡ Ε±π≤ΡΝœ , “ρ¥Υœό÷ΤΝΥΗΟΝλ”ρΒΡΖΔ’ΙΓΘΆ®ΙΐΕ‘ΝΉΥαΜ·ΕύκΡΒΡ Ε±π―–ΨΩ ,Ω…ΜΐάέΕ‘Κ§≤ΜΆ§ΒγΚ…≤–Μυ Ε±πΒΡΒςΩΊΨ≠―ι ,”–άϊ”Ύ…ν»κΝΥΫβάκΉ”Μ·ΜυΆ≈Ε‘ΕύκΡ Ε±πΒΡ”ΑœλΓΘΕ‘ΝΌ¥≤ΩλΥΌΦλ≤βΝΉΥαΜ·ΒΑΑΉΨΏ”–÷ΗΒΦ“β“εΓΘ‘ΎΑ±ΜυΥα÷– ,Ser (ΥΩΑ±Υα) ΓΔThr (Υ’Α±Υα) ΚΆTyr(ά“Α±Υα) …œΕΦΚ§”–τ«Μυ , «ΝΉΥαΜ·ΒΡΈΜΒψΓΘ“ρ¥Υ ,Ε‘”ΎΝΉΥαΜ·κΡΒΡ Ε±πΆυΆυ «Ά®ΙΐΚœ≥…Μρ’Ώ…ζΈο ήΧε”κΝΉΥαΫαΙΙΒΞ‘Σ–Έ≥…ΒΡάκΉ”ΦϋΓΔ¬γΚœ≈δΈΜΒ»–ßΙϊ Βœ÷ΓΘ
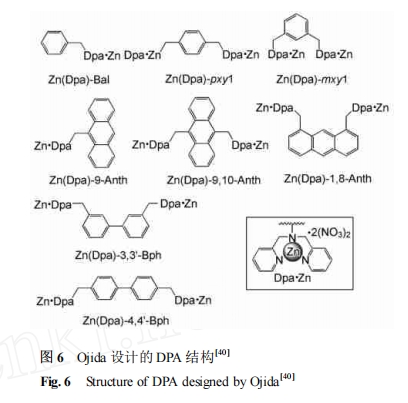
ΨΆΡΩ«ΑΕ‘ΝΉΥαΜ·κΡΒΡ Ε±πά¥ΥΒ ,Κœ≥…ΒΡ ήΧε÷ς“Σ «άϊ”ΟΝΉΥαΫαΙΙΒΡΫπ τάκΉ”¬γΚœ≈δΈΜΉς”ΟΓΘOjidaΒ»Κœ≥…ΝΥΫπ τάκΉ”–Ω≈δΈΜΒΡ DPA ΫαΙΙ ,Ω…“‘Ε‘ΝΉΥαΜ·κΡΫχ––”––ßΒΡ Ε±πΓΘΆΦ 6 ΈΣΥϊΟ«…ηΦΤΒΡ“ΜœΒΝ–ΒΡDPA ΫαΙΙΓΘ‘ΎΝΫΗω DPA ΫαΚœ±έ÷°Φδ”…ΨΏ”–”ΪΙβ–ßΙϊΒΡΫαΙΙœύΝ§Ϋ” ,Β±ΥϋΟ«”κΝΉΥαΜ·κΡΫχ––ΫαΚœΒΡ ±Κρ ,”ΪΙβ–ßΙϊΜαΥφΉ≈ΫαΚœ≥ΧΕ»ΒΡ≤ΜΆ§Εχ≤ΜΆ§≥ΧΕ»ΒΊ‘ω«Ω ,Ψί¥ΥΜώΒΟ÷ςΧεΩΆΧε÷°ΦδœύΜΞΉς”ΟΒΡΦΤΝΩ±»“‘ΦΑΫαΚœ≥Θ ΐ ,≤ΔΕ‘ Ε±πΙΐ≥ΧΫχ––ΝΥΖ÷ΈωΓΘ
Yamaguchi Β»[41 ] ΚΆ Yoshimura Β»[42 ] “≤Κœ≥…ΝΥάύΥΤΒΡΫαΙΙ ,DPA ΫαΙΙΒΞ‘ΣΆ®ΙΐΠ–2Π–Ε―ΜΐΙΧΕ®ΒΫΡΐΫΚΫαΙΙ…œ ,Ε‘ΝΉΥαάύΫαΙΙ“‘ΦΑΚ§”–ΝΉΥαΫαΙΙΒΡ―ή…ζΈοΫχ–– Ε±πΓΘΥϊΟ«ΜΙΖ÷ΈωΝΥ’β÷÷Υ°ΡΐΫΚΕ‘≤ΜΆ§ΝΉΥα―ή…ζΈο ,»γ ATPΓΔΝΉΥαΜ·ΒΡΥΩΑ±ΥαΓΔΝΉΥαΗυάκΉ”Β»ΒΡ―Γ‘ώ–‘ΓΘ
Υδ»Μ’κΕ‘ΧΊΕ®≤–Μυ…ηΦΤΒΡ ήΧεΕ‘ΕύκΡ”–“ΜΕ®ΒΡ―Γ‘ώ–‘ ,ΒΪ”…”ΎΉ‘»ΜΫγ÷–κΡΒΡ÷÷άύΈόΖ®ΙάΝΩ ,Κ§”–œύΆ§≤–ΜυΒΡκΡΒΡ ΐΝΩ“≤ΚήΨό¥σ ,ΒΦ÷¬Ε‘ΕύκΡΒΡ―Γ‘ώ–‘Ϋœ≤νΓΘκΡ–ρΝ–―Γ‘ώ–‘ ήΧεΒΡ…ηΦΤΚΆΉιΚœΜ·―ßΒΡΖΔ’ΙΈΣκΡΒΡΉ®“Μ–‘ Ε±πΧαΙ©ΝΥ“ΜΕ®ΒΡΑο÷ζΓΘ
2. 2 –ΓΖ÷Ή” ήΧεΕ‘ΕΧκΡΒΡ–ρΝ–―Γ‘ώ–‘ Ε±π
Ε±π“ΜΗωΒΑΑΉΒΡ±μΟφ–η“Σ‘ΎΙΙ–Ά»ΖΕ®ΒΡΜΖΨ≥ΡΎ≈≈Ν–¥σΝΩΒΡΙΠΡήΜυΆ≈[17 ] ΓΘΕύκΡΒΡ–ρΝ–―Γ‘ώ–‘ Ε±π «ΒΑΑΉ±μΟφΈΜΒψΉ®“Μ–‘ Ε±πΒΡΜυ¥Γ ,Ε‘ΒΑΑΉΙΠΡήΜ·ΩΊ÷ΤΓΔΒΑΑΉ-ΒΑΑΉΦΑΒΑΑΉ-Ι―κΡœύΜΞΉς”ΟΒΡάμΫβ”–÷Ί“ΣΑο÷ζΓΘStill Ήι «Ήν‘γΩΣ Φ“‘ΕύκΡΈΣΡΩ±ξΈοΫχ–––ρΝ–―Γ‘ώ–‘ Ε±πΒΡ―–ΨΩΉι÷°“Μ ,ΥϊΟ«ΒΡ÷ς“ΣΡΩ±ξ «ΜΐάέκΡ Ε±πΒΡάμ¬έ÷Σ Ε ,≤ΔΫΪ’β–©άμ¬έ÷Σ Ε”Ο”Ύ«ΉΚΆ–‘ΒΡ»ΖΕ®ΚΆΦΤΥψΜζ‘Λ≤βΓΘStill Β»[43 ΓΣ46 ] “―Ψ≠Ηχ≥ω“ΜΗωΑϋΚ§ΚήΕύ Ε±π‘ΣΥΊΒΡΤ·ΝΝΉιΚœ ,’β–©–≈œΔ‘ΎκΡ Ε±πΚΆκΡΩβΙΙΫ®ΖΫΟφΨΏ”–÷Ί“ΣΒΡ“β“εΓΘ¥σ≤ΩΖ÷–ρΝ–―Γ‘ώ–‘ Ε±πΒΡ±®ΒάΕΦ «Ά®ΙΐΒΞ“ΜΑ±ΜυΥα≤–Μυ Ε±πΈΜΒψΒΡΦρΒΞΝ§Ϋ”ΙΙ÷ΰ ,»γΜΖΚΐΨΪΓΔΙΎΟ―ΜρθΘΑΖΜυΆ≈ΓΘ
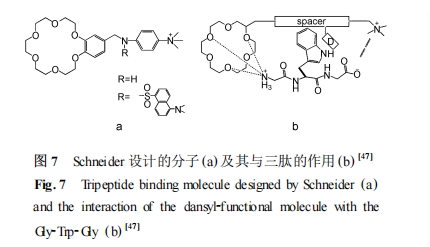
Schneider Β»’κΕ‘Ζ«±ΘΜΛκΡΕΥΜυΚΆ≤ύΝ¥“ΐ»κΕύ÷÷ Ε±πΜυΆ≈ ,άϊ”Ο ήΧεΕ‘ΥΪΕΥΜυΚΆ≤ύΝ¥ΒΡΙ≤Ά§Ής”Ο Ε±πΕύκΡ–ρΝ–ΓΘ»γΆΦ 7 Υυ Ψ ,Υ°÷–ΚΆΦΉ¥Φ÷–ΒΡΚΥ¥≈ΒΈΕ®÷ΛΟς ,18-ΙΎ-6 ΜυΆ≈ΚΆΕύκΡ N ΕΥ―τάκΉ”œύΜΞΉς”Ο , ήΧεΑ±Μυ―τάκΉ””κΕύκΡ C ΕΥœύΜΞ“θάκΉ”Ής”ΟΓΘΒΞΜ«ΜυΆ≈ΒΡ“ΐ»κ”–ΝΫΗωΡΩΒΡ ,“ΜΗω «‘ωΦ”ΝΥ ήΧεΖ÷Ή””κκΡΖΦΜΖ≤ύΝ¥ΒΡœύΜΞΉς”Ο ,Νμ“ΜΗω‘ρ «Ω…“‘”Ο”ΪΙβΙβΤΉ≤βΕ®œύΜΞΉς”ΟΓΘ’βΗωΙΛΉςΉν¥σΒΡ“β“ε‘Ύ”Ύ‘ΎΆ§“ΜΧεœΒ÷–“ΐ»κΕύ÷÷ΙΠΡήΜυΆ≈Ι≤Ά§Ής”Ο ,ΨΏ”–Κή¥σΒΡ÷ΗΒΦ“β“εΓΘΚσά¥ΥϊΟ«”÷Ά®ΙΐάύΥΤΒΡΖΫΖ®“ΐ»κΏ≤ΏχΜυΆ≈ΉςΈΣΑ±ΜυΥαΚΆ»ΐκΡΒΡΉœΆβ2Ω…Φϊ¥ΪΗ–Τς[48 ] ΓΘ
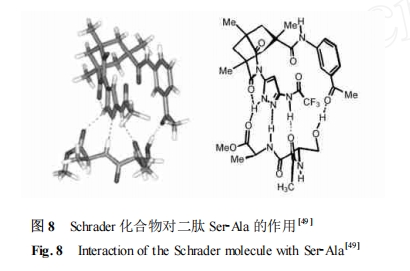
ΡΘΡβΉ‘»ΜΫγΒΡκΡ ήΧε «…ηΦΤκΡ–ρΝ–―Γ‘ώ–‘ ήΧεΒΡ“ΜΗω÷Ί“ΣΖΫΖ®ΓΘSchrader Β»[49 , 50 ] ΖΔ’ΙΝΥ–ρΝ–―Γ‘ώ–‘κΡ ήΧεΒΡΚœάμ…ηΦΤάμ¬έΓΘΥϊΒΡάμ¬έ»œΈΣΚήΕύ…ζΈοΧεœΒΕΦΈΣκΡ–ρΝ–ΒΡ”––ß Ε±πΧαΙ©“ΜΗω«≥ΙΒ ,≤ΔΗ®“‘“ΜΗωΕ‘ΡΩ±ξΑ±ΜυΥα≤–ΜυΒΡΉ®“Μ–‘ΦϋΚœΒΡΩΎ¥ϋΓΘSchrader ΩΈΧβΉι―Γ”Ο kempΓδs ΥαΉςΈΣ ήΧεΙ«Φή ,ΈΣκΡ Ε±πΧαΙ©Ω’ΦδΓΘKempΓδs ΥαΝΫΗωτ»Μυ“ΜΗωΝ§Ϋ”ΒΞθΘΜ·ΒΡ 3 ,5-ΕΰΑ±ΜυΏΝΏρ ,Νμ“ΜΗωΫ”ΦδΈΜ»Γ¥ζΒΡ±ΫΑΖ―ή…ζΈοΉςΈΣ Ε±πΈΜΒψ ,ΆΦ 8 Ηχ≥ωΝΥΗΟΜ·ΚœΈοΕ‘ΕΰκΡSer-Ala ΒΡ Ε±π«ιΩω ,Monte Carlo ΫαΙϊœ‘ ΨΤδ”κ Ser-Ala ΕΰκΡΒΡ Ε±πΈΣ»ΐΒψ–≠Ά§Ής”ΟΓΘ
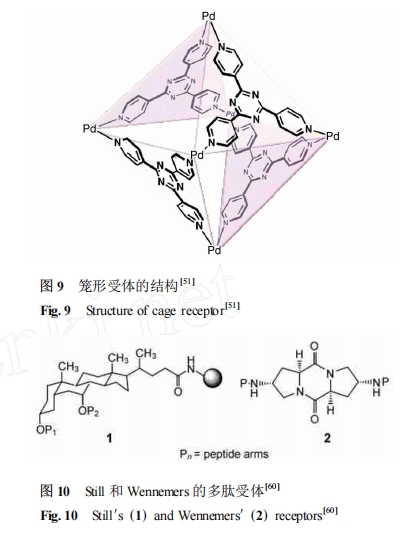
»ΜΕχ Fujita Β»[51 ] ”ΟΙΐΕ…Ϋπ τ Pd ÷Τ±ΗΝΥ“Μ÷÷–¬–ΆΒΡ Pd ≈δΈΜΒΡ»ΐύΚΝΐ–Έ ήΧε (ΆΦ 9) ΓΘ’β÷÷Ή‘ΉιΉΑ≈δΈΜΝΐ ήΧεΆ®ΙΐΠ–-Π–œύΜΞΉς”ΟΚΆ CH-Π–œύΜΞΉς”ΟΒΡ–≠Ά§Ής”ΟΕ‘Κ§”– Trp-Trp-Ala Ν¥ΕΈΒΡκΡΨΏ”– 10^6M- 1“‘…œΒΡΉς”Ο≥Θ ΐ ,ΕχΕ‘”ΎΚ§―τάκΉ”–Ά His ≤–ΜυΒΡ Ac-Trp-His-Ala-NH2 ≤Μ¥φ‘ΎΫαΚœΡήΝΠ ,œ‘ ΨΝΥΤδΗΏΕ»ΒΡ–ρΝ–―Γ‘ώ–‘ΓΘ ήΧε”κΕύκΡΒΡΒΞΨßΫαΙϊ÷Λ ΒΝΥΥϋΟ«÷°ΦδΒΡΉς”ΟΡΘ Ϋ «“Μ÷÷ ηΥ°Ής”ΟΓΘ
ΉιΚœΜ·―ß‘ΎΕύκΡΒΡ–ρΝ–―Γ‘ώ–‘ ήΧε―–ΨΩΖΫΟφΨΏ”–÷Ί“Σ”Π”Ο ,Ά®ΙΐκΡΩβΩ…“‘…Η―Γ≥ωΚœ ΒΡ–ρΝ–―Γ‘ώ–‘ ήΧε[52 , 53 ],ΡΩ«Α’βΖΫΟφΒΡ―–ΨΩ±®ΒάΚήΕύ ,Εύ ΐ≤Έ”κΕύκΡ–ρΝ–―Γ‘ώ–‘ Ε±π―–ΨΩΒΡΩΈΧβΉι‘ΎΉιΚœΜ·―ßΖΫΟφΕΦ”–≤ΜΆ§≥ΧΕ»ΒΡΫι»κΓΘΆ®ΙΐΉιΚœΜ·―ß”≈Μ· ,“―Ψ≠»ΓΒΟ“Μ–©Ω…ΙέΒΡ≥…Ιϊ[54 ΓΣ57 ] ΓΘΉιΚœΜ·―ßΦΦ θ“≤ Ι¥σΝΩ”––ß ήΧεΒΡ…ηΦΤΚΆΚœ≥…≥…ΈΣΩ…ΡήΓΘStill ΒΡ ήΧε1[58 ]ΚΆ Wennemers ΒΡΗ’–‘Ϋœ«ΩΒΡ ήΧε 2[59 ] ΨΆ «ΫœΚΟΒΡάΐΉ”(ΆΦ 10) ΓΘ
Anslyn Β»[60 ]άϊ”Ο Still ΚΆ Wennemers ΒΡΥΦ¬Ζ…ηΦΤΝΥΑϋΚ§Η’–‘Ι«ΦήΚΆΩ…±δ»ΐκΡ±έΒΡ ήΧε Ε±π»ΐκΡ ,―–ΨΩ±μΟςΗΟ ήΧεΕ‘ Xxx-Lys-Lys ( Xxx ΈΣ HisΓΔCys ΜρMet) ΒΡ»ΐκΡΨΏ”––ρΝ–―Γ‘ώ–‘ ,ΫαΚœ≥Θ ΐΫ”Ϋϋ 10^6M- 1 ΓΘΥϊΟ«Ά§ ±ΜΙ÷Λ Β»ΐκΡ ÷±έΒΡ–≠Ά§Ής”Ο (άκΉ”Ε‘) Ρή‘ωΦ”ΫαΚœΡήΝΠ ,»γ His-Lys-Lys(110 ΓΝ10^6 M- 1)> His-Gly-Gly(115 ΓΝ10^4 M- 1) > Lys-Lys-His(0) , Gly-Gly-Gly(0) ΓΘ
2. 3 –ΓΖ÷Ή” ήΧεΕ‘ΕύκΡΕΰΦΕΫαΙΙΒΡ”’ΒΦ”κ Ε±π
Εύ ΐ…Ό”ΎΜρΒ»”Ύ 30 ΗωΑ±ΜυΥα≤–ΜυΒΡΕύκΡ‘ΎΥ°»ή“Κ÷–≤Δ≤Μ“‘Ιφ’ϊΒΡΙΙ–Ά¥φ‘Ύ ,Εχ «“‘–μΕύΉ‘”…ΡήΫϋΥΤΒΡΙΙ–Ά»Κ–Έ Ϋ¥φ‘Ύ[61 ] ΓΘ“ρ¥Υ ,Ω…“‘…ηΦΤ“ΜΕ®ΒΡ ήΧε ,Ά®ΙΐœύΜΞΉς”Ο”’ΒΦ≤ΔΈ»Ε®ΕύκΡΒΡΡ≥“ΜΙΙ–Ά , ΙΤδΉ‘”…ΡήΫΒΒΆ≤Δ‘ΎΙΙ–Ά»Κ÷–’Φ”–Εύ ΐ ,¥”Εχ¥οΒΫ Ε±πΕύκΡΒΡΉν÷’ΡΩΒΡΓΘΒ±«Α―–ΨΩ÷–÷ς“ΣΦ·÷–”ΎΠΝ¬ί–ΐΚΆΠ¬’έΒΰΒΡ”’ΒΦ Ε±πΓΘ
ΠΝ¬ί–ΐ «“Μ÷÷÷Ί“ΣΒΡΕΰΦΕΫαΙΙ ,‘ΎΕύ÷÷ΒΑΑΉ÷ -ΒΑΑΉ÷ ±μΟφœύΜΞΉς”Ο÷–Αγ―ίΉ≈÷Ί“ΣΒΡΫ«…ΪΓΘΚœάμ…ηΦΤΕ‘ΠΝ¬ί–ΐΨΏ”–ΧΊΕ®―Γ‘ώ–‘ Ε±πΒΡ ήΧε «“ΜΗωΧτ’Ϋ ,’β÷÷Χτ’Ϋ‘¥”ΎΠΝ¬ί–ΐΫœ¥σΒΡ±μΟφ«χ”ρ ,“‘ΦΑ ηΥ°ΚΆΨ≤ΒγΜΞ≤Ι‘Ύ±μΟφΆΊΤΥ―ß…œΒΡΤΞ≈δΓΘ
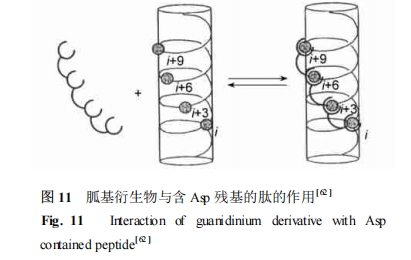
Hamilton ΩΈΧβΉι[17 , 62 ] άϊ”Οκ“Μυ”κκΡ…œ Asp ≤–ΜυΒΡΜΞ≤ΙΒΡΧΊ–‘ ,…ηΦΤΝΥ“ΜœΒΝ–Κ§”–ΕύΗωκ“ΜυΫαΙΙΒΡ ήΧε ,’β–© ήΧεΩ…“‘‘ΎΨΚ’υ–‘»ήΦΝ÷–Ε‘ΡΘ–ΆΠΝ¬ί–ΐκΡΫχ–– Ε±πΓΘ”…”ΎΫΒΒΆΝΥ»Ξ»ήΦΝΜ·ΡήΝΩΚΆ»ήΦΝΫœΒΆΒΡΫιΒγ≥Θ ΐ ,’β÷÷œύΜΞΉς”Ο‘Ύ”–Μζ»ήΦΝάοΗϋΈΣΟςœ‘ΓΘ»γΆΦ 11 Υυ Ψ ,κ“ΜυΖ÷Ή””κ Asp ≤ύΝ¥œύΜΞΉς”Ο ,κ“ΜυΦδΨύάκ“άάΒ”Ύ Asp ≤–ΜυΦδΨύάκ( i + 3 n) ΓΘ
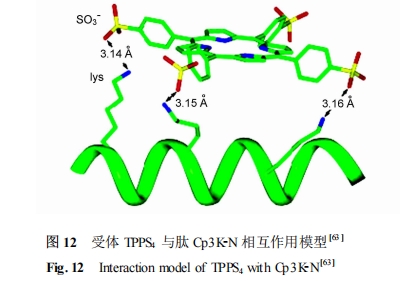
Fairman Β»[63 ]άϊ”Ο Lys ≤ύΝ¥Α±Μυ’ΐάκΉ”Ρή”κ“θάκΉ”œύΜΞΉς”ΟΒΡΧΊΒψ ,…ηΦΤΝΥΑϋΚ§ 3 Ηω≤ΜΝ§–χ Lys≤– Μυ ΒΡ κΡ : Ile-Gln-Gln-Leu-Lys-Asn-Gln-Ile-Lys-Gln-Leu-Leu-Lys-Gln (Cp3K2N) ΦΑΤδœύ”ΠΒΡ“θάκΉ”Ώ≤Ώχ ήΧε(ΡΎœϊ–ΐ-ΥΡ-(4-Μ«Μυ±Ϋ)-Ώ≤Ώχ , TPPS4) ΓΘΉœΆβΓΔ‘≤Εΰ…ΪΦΑΖ÷Έω≥§ΥΌάκ–ΡΫαΙϊ±μΟς , ήΧε TPPS4 ΡήΆ®ΙΐΕύ÷ΊΨ≤ΒγœύΜΞΉς”Ο”κκΡ Cp3K-N ΖΔ…ζΧΊ“λ–‘œύΜΞΉς”Ο ,≤Δ”’ΒΦΤδ–Έ≥…ΠΝ¬ί–ΐΫαΙΙ (ΆΦ 12) ΓΘTPPS4 ΒΡ’β÷÷ΧΊ“λ–‘ΚΆ”’ΒΦΠΝ¬ί–ΐΒΡΡήΝΠ‘ΎΒγΉ”ΚΆΙβ―ßΜν–‘ΒΡΗ–”Π≤ΡΝœΖΫΟφΨΏ”–«±‘ΎΒΡ”Π”ΟΓΘ
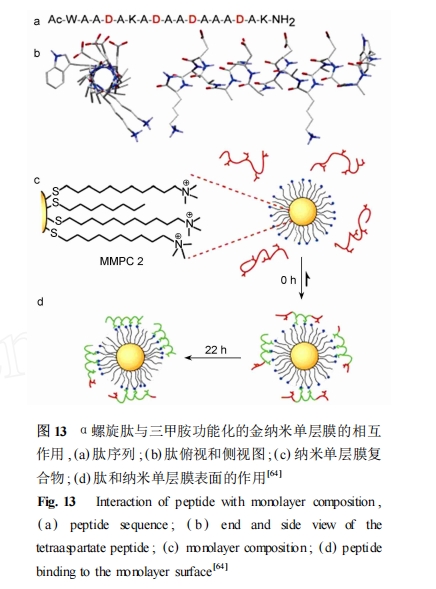
ΧαΗΏ ήΧεΕ‘…ζΈοΖ÷Ή”«ΉΚΆ–‘ΒΡ“ΜΗω”––ßΖΫΖ®ΨΆ «άϊ”ΟΡΩ±ξΒΑΑΉ±μΟφΒΡΡΘΑε ήΧεΫχ–– Ε±π ,’β“―Ψ≠Ά®ΙΐΖ÷Ή”±μΟφ”ΓΦΘΓΔ ς÷§”ΓΦΘ“‘ΦΑΫπ τ”ΓΦΘΒΡΡΘΑεΨέΚœ Βœ÷ΓΘΡ…ΟΉΒΞ≤ψΡΛΉςΈΣΡΘΑεΡήΈΣΡΩ±ξΖ÷Ή”ΧαΙ©Ϋœ¥σΒΡ±μΟφ ,’β ΙΥϋΟ«≥…ΈΣ…ζΈο±μΟφ Ε±πΒΡΙΛΨΏ ,“ρ¥Υ“≤≥…ΈΣΡΩ±ξΠΝ¬ί–ΐΒΡ«±‘Ύ ήΧεΓΘRotello Β»[64 ]±®ΒάΝΥ‘ΎΥ°»ή“Κ÷–»ΐΦΉΑΖΙΠΡήΜ·ΒΡΫπΡ…ΟΉΒΞ≤ψΡΛΕ‘Κ§ 4ΗωΧλΕ§Α±Υα≤–ΜυΒΡΠΝ¬ί–ΐκΡ”–œ‘÷χ Ε±πΚΆΈ»Ε®Ής”Ο ,»γΆΦ 13 Υυ ΨΓΘ
Π¬’έΒΰΒΡ”’ΒΦ Ε±π÷ς“Σ”–ΝΫ÷÷ΖΫ Ϋ ,Φ¥ΡΘΡβ≥…ΚΥΝ¥ ( nucleating strand mimics) ΚΆ ΡΘ Ρβ ≥… ΚΥ ΉΣ Ϋ«( nucleating turn mimics) , Nowick Β»[65 ,66 ] ΚΆ KellyΒ»[67 ,68 ]ΒΡΖ÷±πΫ®ΝΔΙΐ≥…ΙΠΒΡΡΘ–ΆΧεœΒΓΘ
3 ΗΏΖ÷Ή”ΚΆΕΧκΡœύΜΞΉς”Ο
3.1 œΏ–‘ΗΏΖ÷Ή””κΕΧκΡœύΜΞΉς”Ο
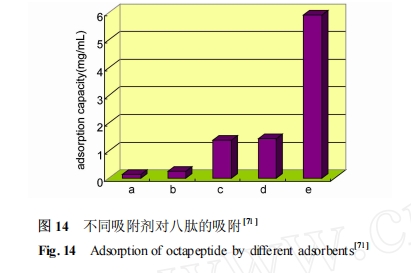
œΏ–‘ΗΏΖ÷Ή””κκΡœύΜΞΉς”ΟΒΡ±®Βά‘ΎΈΡœΉ÷–≥ωœ÷Ϋœ…Ό[69 , 70 ],±Ψ―–ΨΩΉιΆ®Ιΐ”κ«ΉΥ°ΒΞΧεΙ≤ΨέΒΡΖΫΖ®ΫΪΙΠΡήΒΞΧε“ΐ»κΗΏΖ÷Ή”ΧεœΒ ,÷Τ±ΗΝΥœΒΝ–ΑϋΚ§ΙΠΡήΒΞΧεΒΡΥ°»ή–‘Ι≤ΨέΈο , ≤ΔΫΪΤδ”ΟΉςΈϋΗΫ≤ΡΝœΒΡΡΘ–Ά[71 ΓΣ75 ] ΓΘΆ®ΙΐΒ»Έ¬ΒΈΕ®ΈΔΝΩ»»ΓΔΚΥ¥≈ΓΔ”ΪΙβΚΆ±μΟφΒ»άκΉ”ΧεΙ≤’ώΒ»œ÷¥ζΖ÷Έω≤β ‘ ÷ΕΈ―–ΨΩΡΘ–ΆΨέΚœΈο”κΕύκΡ‘ΎΥ°÷–ΒΡΉς”ΟΜζάμ ,¥”ΕχΫχ“Μ≤Ϋ÷ΗΒΦΈϋΗΫΦΝΒΡΫαΙΙ…ηΦΤ ,ΈΣΗΏ―Γ‘ώ–‘ΕύκΡΈϋΗΫ≤ΡΝœΒΡ…ηΦΤΧαΙ©ΝΥ“ΜΕ®ΒΡάμ¬έ÷ΗΒΦΓΘΆΦ 14 Ηχ≥ωΝΥΦΗ÷÷ΈϋΗΫΦΝΕ‘ΡρΕΨ÷Δ÷–Ζ÷Ή”ΑΥκΡ(Val-Val-Arg-Gly-Cys-Thr-Trp-Trp) ΒΡΈϋΗΫΡήΝΠ ,¥”ΆΦ÷–Ω…“‘Ω¥≥ω ,ΕΓΜυ–ό ΈΒΡΨέ““œ©ΑΖ(E) ΈϋΗΫΦΝΕ‘ΑΥκΡΒΡΈϋΗΫΡήΝΠΟςœ‘«Ω”ΎΤδΥϊΦΗ÷÷ΈϋΗΫΦΝΓΘΕΰΈ§ΚΥ¥≈Ε‘Κ§”–œύΆ§ΙΠΡήΒΞΧεΒΡΥ°»ή–‘ΨέΚœΈοΡΘ–Ά”κΑΥκΡœύΜΞΉς”ΟΒΡ―–ΨΩ±μΟς ,ΡΘ–ΆΨέΚœΈο”κΑΥκΡΒΡœύΜΞΉς”Ο«Ω»θ«ς Τ”κΈϋΗΫΦΝΒΡΈϋΗΫΫαΙϊΜυ±Ψ“Μ÷¬ ,÷ΛΟςΝΥœΏ–‘ΨέΚœΈοΉςΈΣΈϋΗΫΦΝΡΘ–Ά…Η―Γ”––ßΙΠΡήΒΞΧεΒΡ”––ß–‘ΓΘΆ§ ± ,Ά®ΙΐΕ‘ΈϋΗΫΦΝΡΘ–Ά”κΑΥκΡœύΜΞΉς”ΟΒΡ―–ΨΩ ,÷ΛΟςΝΥΈϋΗΫΦΝΕ‘ΑΥκΡΒΡΈϋΗΫΜζάμ « ηΥ°Ής”ΟΚΆΨ≤ΒγΉς”ΟΒΡ–≠Ά§Ής”ΟΓΘ
3. 2 κΡΈϋΗΫΖ÷άκ≤ΡΝœ
ΥφΉ≈…ζΈοΦΦ θΒΡ≤ΜΕœΖΔ’Ι ,‘Ύ―–ΨΩΕύκΡΚΆΒΑΑΉ÷ ΒΡΫαΙΙ”κΙΠΡή ± ,“ΜΗω±Ί≤ΜΩ……ΌΒΡΜΖΫΎ «“ΣΫΪΤδ¥”≥…«ß…œΆρ÷÷Η¥‘”ΒΡΜλΚœΈο÷–Ζ÷άκ≥ωά¥ ,≤ΔΒΟΒΫΗΏ¥ΩΕ»ΒΡ―υΤΖΓΘ“ρ¥Υ ,ΫϋΡξά¥Ε‘”ΎΕύκΡΒΡΖ÷άκ¥ΩΜ·ΦΦ θ“―Ψ≠ΖΔ’Ι≥…ΈΣ…ζΈοΦΦ θΒΡ“ΜΗω÷Ί“ΣΖ÷÷ßΓΘ‘ΎΖ÷Ή” Ε±πΜυ¥Γ…œΫχ––ΕύκΡΖ÷άκ≤ΡΝœΒΡ…ηΦΤΫΪΦΪ¥σΒΊΧαΗΏΖ÷άκ–ßΙϊΓΘΡΩ«ΑΜυ”ΎΖ÷Ή” Ε±π‘≠άμ…ηΦΤΒΡΕύκΡΖ÷άκ¥ΩΜ·≤ΡΝœ÷ς“ΣΖ÷ΈΣ«ΉΚΆΈϋΗΫΦΝΓΔΖ÷Ή””ΓΦΘΨέΚœΈοΒ»ΦΗάύΓΘ
3. 2. 1 «ΉΚΆΈϋΗΫΦΝ
«ΉΚΆΈϋΗΫΦΝΩ…“‘”Π”Ο”ΎΕύ÷÷Έο÷ ΒΡΖ÷άκ ,ΥϋΒΡΖ÷άκ‘≠άμ «Μυ”ΎΖ÷Ή”ΦδΨ≤ΒγΓΔ«βΦϋΓΔ ηΥ°œύΜΞΉς”Ο“‘ΦΑΫπ τ≈δΈΜΉς”ΟΒΡΖ÷Ή” Ε±πΓΘ«ΉΚΆ≈δΧεΑϋά®…ζΈοΧΊ“λ–‘≈δΧεΚΆΖ¬…ζΈοΧΊ“λ≈δΧεΓΘ”…”ΎΩΙΧε °Ζ÷ΑΚΙσ«“»ί“Ή±δ–‘ ,“ρ¥Υ…ζΈοΧΊ“λ–‘≈δΧεΈϋΗΫΦΝΒΡ”Π”Ο”–Κή¥σΒΡΨ÷œό–‘ΓΘΡΩ«Α ,”Π”ΟΗΟΖΫΖ®…ηΦΤΕύκΡ«ΉΚΆΈϋΗΫΦΝ÷ς“Σ «ΙΧΕ®Μ·Ϋπ τάκΉ”«ΉΚΆ…ΪΤΉ (immobilizedmetal ion affinity chromatography , IMAC) ΒΡ÷υΧν≥δ≤ΡΝœ[76 ΓΣ81 ]
IMAC ΉςΈΣ“Μ÷÷–¬–ΆΗΏ–ß…ΪΤΉΖ÷άκΡΘ Ϋ ,άϊ”Ο«ΉΚΆΈϋΗΫΦΝΉςΈΣ÷υΧν≥δ≤ΡΝœ ,“―Ψ≠±ΜΙψΖΚ”Π”Ο”ΎκΡάύΈο÷ ΒΡΖ÷άκΖ÷ΈωΓΘIMAC ≥ΘΉςΈΣ≤ΕΜώΠΑ ΆΖ≈÷υ ,ΗΜΦ·Ρ≥“ΜάύκΡΕΈ(»γ±μΟφΚ§”–¬ψ¬ΕΒΡΉιΑ±Υα≤–ΜυΜρΝΉΥαΜ·ΒΡκΡΕΈ) ,“‘ΦΑΚ§”–ΉιΑ±Υα±ξ«©ΒΡ÷ΊΉιΕύκΡΒΡΖ÷άκ¥ΩΜ·ΓΘΉόΚΚΖ®Β»[76 ] Ά®Ιΐ÷±Ϋ”ΨέΚœΖΫΖ®ΒΟΒΫΝΥΚ§”–ΝΉΥαθΞΜυΆ≈ΒΡ’ϊΧε÷υ≤ΡΝœ ,άϊ”ΟΝΉΥαθΞΜυΆ≈”κ Ti4 +ΒΡρϋΚœ≈δΈΜΉς”ΟΙΧΕ®Μ· Ti4 +,÷Τ±ΗΝΥ Ti4 +-IMAC(ΆΦ15) ΓΘ‘ΌΆ®Ιΐ Ti4 + ”κΝΉΥαΜ·κΡ…œΝΉΥαΜυΆ≈ΒΡ―Γ‘ώ–‘Ής”Ο¥οΒΫΗΜΦ·ΒΡΡΩ±ξΓΘ Β―ιΫαΙϊ±μΟς Ti4 + ΙΧΕ®Μ·«ΉΚΆ…ΪΤΉΙΧΕ®œύΕ‘ΝΉΥαΜ·κΡΕΈΨΏ”–ΚήΗΏΒΡ―Γ‘ώ–‘ΚΆΟςœ‘ΒΡΗΜΦ·–ßΙϊΓΘΖ÷άκΚΆΗΜΦ·ΝΉΥαΜ·κΡΒΡ―Γ‘ώ–‘ΚΆ–߬ ”≈”ΎΤδΥϊΗΜΦ·ΖΫΖ®»γ :Fe3 +-IMAC ,Zr4 +-IMAC[82 ],TiO2 “‘ΦΑ ZrO2 Β»ΓΘ

3. 2. 2 Ζ÷Ή””ΓΦΘΨέΚœΈο
ΫΪ“ΣΖ÷άκΒΡΡΩ±ξΖ÷Ή””κΫΜΝΣΦΝ‘ΎΨέΚœΈοΒΞΧε»ή“Κ÷–Ϋχ––Ι≤Ψέ÷Τ±ΗΒΟΒΫΩ≈ΝΘΫι÷ ,»ΜΚσάϊ”Οœ¥Ά―ΜρίΆ»ΓΒΡΖΫ Ϋ≥ΐ»ΞΑϋ¬ώ‘ΎΫι÷ ÷–ΒΡΡΩ±ξΖ÷Ή” ,±ψΒΟΒΫΖ÷Ή””ΓΦΘΨέΚœΈοΓΘ¥Υ ±ΒΡΗΏΖ÷Ή”Ϋι÷ ÷––Έ≥…ΝΥ‘Ύ»ΐΈ§Ω’Φδ¥σ–ΓΓΔ–ΈΉ¥“‘ΦΑΙΠΡή≈δΧεΕΦ”κΡΘΑεΖ÷Ή”ΜΞ≤ΙΒΡΖ÷Ή””ΓΦΘΈΔ«Μ ,Ω…“‘ΫΪΡΩ±ξΖ÷Ή”ΒΡΩ’ΦδΫαΙΙ±Θ≥÷ΓΑ”ΓΦΘΓ±ΜρΝτœ¬ΓΑΦ«“δΓ±ΓΘ’β÷÷Ζ÷Ή””ΓΦΘΨέΚœΈο”÷±Μ–ΈœσΒΊ≥ΤΈΣΓΑΥήΝœΩΙΧεΓ±ΜρΓΑ»ΥΙΛΩΙΧεΓ±,Ω… Βœ÷Ε‘ΡΘΑεΖ÷Ή”ΒΡΧΊ“λ–‘ Ε±π ,’β÷÷ΧΊ“λ–‘Ω…“‘ΚΆΕύΩΥ¬ΓΩΙΧεœύφ«Οά[83 , 84 ] ΓΘ
»ΜΕχ ,”…”ΎΕύκΡ‘Ύ»ή“Κ÷–ΙΙ–ΆΉ‘”…Ε»±»Ϋœ¥σ ,≤Δ≤Μ Κœ”ΎΖ÷Ή””ΓΦΘΓΘ≥ΐ…Ό ΐ‘ΎΥ°÷–ΫαΙΙΚή»ΖΕ®ΒΡΕύκΡ[85 ]Άβ ,’κΕ‘ΕύκΡΖ÷Ή””ΓΦΘΒΡ±®ΒάœύΕ‘Ϋœ…ΌΓΘ“ΜΑψΕ‘ΕύκΡΒΡ”ΓΦΘΕΦ «Μυ”Ύ»ΐΗωΖΫΟφ : (1) ΒΑΑΉ÷ ±μΟφΫαΙΙΒΡ”ΓΦΘ ; (2) ΒΑΑΉ÷ ±©¬Ε”ρΕΧκΡ–ρΝ–ΒΡ”ΓΦΘ ; (3) “‘ Ε±πΒΑΑΉ÷ ΒΡ…ζΈοΖ÷Ή”ΉςΈΣΙΠΡήΒΞΧε÷Τ±Η”ΓΦΘΨέΚœΈοΓΘ
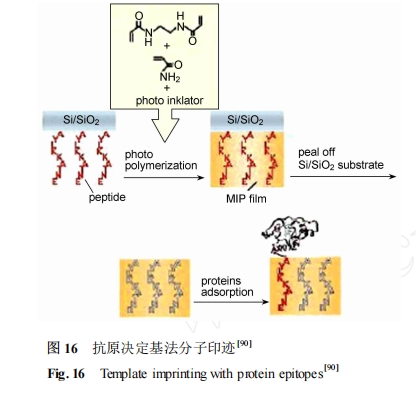
ΡΩ«ΑΖΔ’ΙΫœΈΣ≥… λΒΡ±μΟφ”ΓΦΘΦΦ θ «άϊ”ΟΫπ τάκΉ””κΉιΑ±Υα÷°ΦδΒΡρϋΚœΉς”ΟΖ÷άκ¥χ”–ΉιΑ±ΥαΤ§ΕΈΒΡΕύκΡΓΘShea Β»[86 , 87 ]άϊ”Ο±ϊœ©θΘΑΖΈΣΙΠΡήΒΞΧε ,”ΎΗΜΥ°œύ÷–÷Τ±ΗΖ÷Ή””ΓΦΘΨέΚœΈοΖ÷άκΚ§”– N ΕΥΉιΑ±ΥαΒΡΕύκΡ ,≤ΔΕ‘±»ΝΥ Ni2 + ΓΔCu2 + ΓΔZn2 + ≤ΜΆ§Ϋπ τάκΉ”Ε‘”ΓΦΘ–ßΙϊΒΡ”ΑœλΓΘ Β―ιΫαΙϊ±μΟς Ni2 + Ε‘κΡΒΡΧΊ“λ–‘ Ε±πΉς”ΟΉν«ΩΓΘΒΪ“≈ΚΕΒΡ « ,άϊ”ΟΫπ τρϋΚœΉς”ΟΒΡ±μΟφ”ΓΦΘΦΦ θ÷ΜΡή”Π”Ο”ΎΚ§”–¬ψ¬ΕΉιΑ±Υα≤–ΜυΒΡΒΑΑΉ÷ ,≤Δ«“Ϋπ τρϋΚœΉς”ΟΒΡΧΊ“λ–‘≤Μ«ΩΓΘΫϋΡξά¥ ,–¬ΖΔ’ΙΒΡΩΙ‘≠ΨωΕ®ΜυΖ®[88 , 89 ] «Μυ”Ύ Ι”ΟΕύκΡΜρΒΑΑΉ÷ –ρΝ–÷–¬ψ¬ΕΒΡ“ΜΗωΧΊ“λΕΧκΡ(ΩΙ‘≠ΨωΕ®≤ΩΈΜ)Τ§ΕΈΉςΈΣΡΘΑεΖ÷Ή”(ΆΦ 16) ,άϊ”ΟΥϋΥυΚœ≥…ΒΡΖ÷Ή””ΓΦΘΨέΚœΈο“≤Ρή”––ß Ε±πΕύκΡΚΆ’ϊΗωΒΑΑΉ[90 ] ΓΘ
Ζ÷Ή””ΓΦΘΨέΚœΈοΩ…“‘ΗΏ―Γ‘ώ–‘ΒΊΈϋΗΫΜλΚœΈο÷–ΒΡ”ΓΦΘΖ÷Ή” ,ΒΪ÷Τ±ΗΖ÷άκ≤ΡΝœ÷°«Α±Ί–κΜώΒΟ¥ΐΖ÷άκΈο÷ ΒΡ¥ΩΤΖΓΘ»γΙϊ¥ΩΤΖΡ―”ΎΒΟΒΫΜρΦέΗώΑΚΙσ ,Ρ«Ο¥”ΓΦΘΖΫΖ®ΫΪΡ―”Ύ Βœ÷ΓΘΝμΆβ ,”…”ΎκΡΚΆΒΑΑΉ”…”ΎΤδΫαΙΙΒΡΗ¥‘”–‘ΦΑΤδ”κ”ΓΦΘ≥Θ”ΟΒΡ”–Μζ»ήΦΝΒΡ≤ΜΤΞ≈δ–‘ ,ΒΦ÷¬ΒΑΑΉ÷ …ζΈοΜν–‘ΒΡ…Ξ ß ,»γΚΈΡή‘ΎΥ°œύΫχ––Ζ÷Ή””ΓΦΘ»‘ «“Μ÷÷Χτ’Ϋ[91 ] ΓΘ
4 ΉήΫαΚΆ’ΙΆϊ
÷ΎΕύΒΡΈΡœΉ±®ΒάΕ‘ΕΧκΡ Ε±πΒΡ―–ΨΩΜΙ÷ς“Σœό”Ύ–ΓΖ÷Ή” ήΧεΒΡ…ηΦΤΦΑΚœ≥… ,ΕΧκΡΫαΙΙΒΡ»αΥ≥–‘ΚΆΕύ―υ–‘‘ωΦ”ΝΥΤδ±Μ Ε±πΒΡΡ―Ε»ΓΘΧΊ±π «‘ΎΦΪ–‘»ήΦΝ(…ζΈοΧεœΒ) ÷– ,“Μ–©»θœύΜΞΉς”ΟΆυΆυ“ρ ήΒΫ»ήΦΝΖ÷Ή”ΒΡΨΚ’υ ,Ής”Ο«ΩΕ»±Μ¥σ¥σœς»θ ,ΒΦ÷¬ Ε±πΒΡΡ―Ε»‘ω¥σΓΘΕχ ηΥ°–‘–ΓΖ÷Ή” ήΧεΒΡ…ηΦΤ”÷ ήΒΫΥ°œύ»ήΫβ–‘ΒΡœό÷Τ ,ΖΔ’Ι≤Δ≤ΜΥ≥άϊΓΘΕ‘”ΎΗΏΖ÷Ή”ΧεœΒΕχ―‘ ,ΨΓΙήΖ÷Ή””ΓΦΘΦΦ θ”Ο”ΎΕύκΡΒΡ Ε±πΒΟΒΫΝΥ“ΜΕ®≥ΧΕ»ΒΡ―–ΨΩ ,ΒΪΖ÷Ή””ΓΦΘ”…”ΎΧεœΒΒΡœό÷ΤΡ―”Ύ”–ΚΟΒΡ±μœ÷ΓΘ
¥”ΡΩ«ΑΖΔ’ΙΒΡ«ς Τά¥Ω¥ ,–ΓΖ÷Ή” ήΧε”ΠΗΟ“‘Ης÷÷ Ε±πΜυΆ≈ΒΡ”≈Μ·ΚΆΉιΚœΈΣΜυ¥Γ ,÷Τ±Η‘ΎΥ°ΧεœΒΡΎΨΏ”––ρΝ–―Γ‘ώ–‘ Ε±πΙΠΡήΒΡ ήΧεΓΘΕχ Βœ÷κΡΈϋΗΫΖ÷άκ≤ΡΝœΒΡΗΏ―Γ‘ώ–‘ Ήœ»“Σάϊ”ΟΖ÷Ή” Ε±πάμ¬έΫχ––ΚœάμΒΡ…ηΦΤ≤ΔΆ®ΙΐΦΤΥψΜζΡΘΡβΗ®÷ζ…Η―Γ”––ßΒΡ≈δΧε ,Ά®ΙΐΫχ“Μ≤Ϋ”≈Μ·≈δΧεΚσΫΪ”≈Μ·ΒΟΒΫΒΡ≈δΧε≤ΦΨ÷‘ΎΈϋΗΫΖ÷άκ≤ΡΝœΒΡ±μΟφ ,ΡΘΖ¬ΒΑΑΉ÷ ΓΑάϊ”Ο»θœύΜΞΉς”ΟΫχ––ΉιΉΑΚΆ Ε±πΓ±ΒΡΧΊΒψ , Βœ÷≈δΧεΒΡ–≠Ά§Ής”Ο[92 ],”–Ω…Ρή≤ζ…ζάύΥΤΉ‘»ΜΧεœΒΒΡœλ”Π–‘ΚΆΚΆ–≠Ά§–‘ ,¥οΒΫΗΏ―Γ‘ώ–‘ΒΡ Ε±π–ßΙϊΓΘ
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷ΨΓΘ
