
’Σ“ΣΘΚΜΖΕΰκΡ «ΝΫΗωΑ±ΜυΥαΆ®ΙΐκΡΦϋΥθΚœΕχ≥…ΒΡΜ·ΚœΈοΓΘΜΖΕΰκΡΨΏ”–ΕάΧΊΒΡΫαΙΙΗ≥”ηΤδΝΫΗω«βΙ©Χε”κ«β ήΧεΒΡΧΊ’ςΘ§Υϋ”κ≤ΜΆ§Α–±ξΈο÷ ΫαΚœΚσΡή≤ζ…ζ÷νΕύ–ß”ΠΘ§¥”Εχ±μœ÷≥ωΕύ÷÷…ζΈοΜν–‘ΓΘΜΖΕΰκΡά¥‘¥ΙψΖΚΘ§ΥφΉ≈œ÷¥ζΩΤΦΦΒΡΖΔ’ΙΘ§Ω…“‘¥”’φΨζΚΆœΗΨζ÷–Χα»ΓΖ÷άκ≥ωΜΖΕΰκΡΘ§“≤Ω…“‘Ά®Ιΐ…ζΈοΜρΜ·―ßΖΫΖ®Κœ≥…ΜΖΕΰκΡΓΘ≥ΐΝΥ‘ΎΩΙ…ζΥΊΓΔΩΙ÷ΉΝω“©Έο―–ΖΔΓΔΩΙ―θΜ·ΓΔΫΒ―ΣΧ«Β»ΖΫΟφΨΏ”–Ψό¥σΒΡ«±ΝΠΆβΘ§‘ΎΒςΫΎΡήΝΩ¥ζ–ΜΓΔΒςΩΊ ≥”ϊΓΔΗΡ…Τ…ώΨ≠œΒΆ≥ΚΆΫΒΒΆ–Ρ‘ύ¥ΪΒΦΥΌ¬ Β»ΖΫΟφ“≤”–ΚήΙψάΪΒΡ«ΑΨΑΓΘ“ρ¥ΥΘ§±ΨΈΡΕ‘Ϋϋ5Ρξά¥ΖΔœ÷ΒΡΜΖΕΰκΡΫχ––Ήέ ωΘ§÷ς“ΣΫι…ήΝΥΜΖΕΰκΡΒΡά¥‘¥ΓΔΫαΙΙ”κάμΜ·–‘÷ Θ§≤ΔΕ‘Τδ…ζΈοΜν–‘’ΙΩΣ÷ΊΒψ≤ϊ ωΘ§≤ΔΧα≥ωΝΥΫ®“ιΚΆ’ΙΆϊΓΘ
ΜΖΕΰκΡ «”…2ΗωΑ±ΜυΥα ΉΈ≤œύΝ§–Έ≥…ΒΡΓΔΨΏ”–”≈“λΗ’–‘ΫαΙΙΚΆ…ζΈοΜν–‘ΒΡΏΏύΚΕΰΆΣάύΜ·ΚœΈο(diketopiperazinesΘ§DKPs)ΓΘCurtiusΒ»[1]‘Ύ1888Ρξ Ή¥ΈΚœ≥…≥ωΒΎ“Μ÷÷ΜΖΕΰκΡcyclo(Gly-Gly)ΓΘΜΖΕΰκΡά¥‘¥ΙψΖΚΘ§ΥφΉ≈œ÷¥ζΩΤΦΦΒΡΖΔ’ΙΘ§Ω…“‘¥”’φΨζΚΆœΗΨζ÷–Χα»ΓΖ÷άκ≥ωΜΖΕΰκΡΘ§“≤Ω…“‘Ά®Ιΐ…ζΈοΜρΜ·―ßΖΫΖ®Κœ≥…ΜΖΕΰκΡΓΘΜΖΕΰκΡΒΡΫαΙΙ÷÷άύΖαΗΜΘ§ΫωΫω «≥ΘΙφΧλ»ΜΑ±ΜυΥαΫχ––ΜΖΜ·Υυ–Έ≥…ΒΡΜΖΕΰκΡ÷÷άύΨΆΕύ¥ο210÷÷Θ§ΤδΫαΙΙΒΡΕύ―υ–‘ ΙΒΟΜΖΕΰκΡΨΏ”–Εύ÷÷…ζΈοΜν–‘ΓΘ‘ΎΙΐ»ΞΒΡΦΗΡξ÷–Θ§‘Ϋά¥‘ΫΕύΒΡ―–ΨΩ±®ΒάΝΥΜΖΕΰκΡΒΡ…ζΈοΜν–‘Θ§Αϋά®≤Έ”κœΗΑϊΦδ–≈œΔ¥ΪΒίΓΔ“÷ΨζΚΆΩΙ÷ΉΝωΒ»Θ§Τδ÷–―–ΨΩΉνΈΣΙψΖΚΒΡ «ΤδΩΙΨζΚΆΩΙ÷ΉΝωΒΡΙΠ–ßΓΘάΐ»γcyclo(D-Ala-D-Trp)Ρή”––ß“÷÷Τ»ΥΙ§Ψ±Α©œΗΑϊΒΡ…ζ≥Λ[2]Θ§“‘ΦΑcyclo(Tyr-Gly)“÷÷Τ¥σ≥ΠΑΘœΘΨζΒΡΖ±÷≥[3]ΓΘΚσ”÷¬Ϋ–χ±®Βά÷Λ ΒΝΥΜΖΕΰκΡΒΡΩΙ―θΜ·ΚΆΫΒ―ΣΧ«Β»Ής”ΟΓΘ»γΫώ≤Γ‘≠ΨζΒΡΡΆ“©–‘‘Ϋά¥‘ΫΤ’±ιΘ§άΡ”ΟΩΙ…ζΥΊΒ»Έ Χβ“≤Έ¥ ήΒΫ÷Ί ”Θ§’β–©œ÷œσΕΦ¥Ό ΙΕ‘ΜΖΕΰκΡ…ζΈοΜν–‘Ϋχ––…ν»κ―–ΨΩΓΘ±ΨΈΡ÷ς“ΣΕ‘ΜΖΕΰκΡΒΡ…ζΈοΜν–‘Ϋχ––Ήέ ωΘ§Ά§ ±Ε‘Τδ“©Έο…ζ≤ζΦΑΝΌ¥≤”Π”ΟΒ»ΖΫΟφΉω≥ω’ΙΆϊΓΘ
1 ΜΖΕΰκΡΒΡά¥‘¥
1.1 Ή‘»Μά¥‘¥
ΜΖΕΰκΡά¥‘¥ΙψΖΚΘ§ΤυΫώΈΣ÷ΙΘ§Ψχ¥σΕύ ΐΜΖΕΰκΡά¥Ή‘ΗοάΦ“θ–‘Ψζ[4]Θ§…Ό≤ΩΖ÷ά¥Ή‘”Ύ÷≤ΈοΓΔΕ·ΈοΓΔΗοάΦ―τ–‘Ψζ“‘ΦΑ’φΨζ[5]ΓΘάΐ»γ2017ΡξΘ§GowrishankarΒ»[6]¥”ΫβΒμΖέ―ΩΑϊΗΥΨζ(MMS-50)Χα»ΓΒΡcyclo(L-LeuL-Pro)(CLP)Ρή“÷÷ΤœΗΨζ…ζΈοΡΛΘ§ΒΪ’β≤ΔΖ«…±ΨζΉς”ΟΓΘ2020ΡξΘ§Ιυ«λΖαΒ»[3]‘Ύ¥σΜΤ”ψΙ≤…ζΨζ(Bacillus coagulans LL1103)ΒΡ¥ζ–Μ≤ζΈο÷–ΖΔœ÷ΒΡcyclo(TyrGly)ΓΔcyclo(Pro-Gly)ΚΆcyclo(Pro-Val)Ε‘¥σ≥ΠΑΘœΘΨζΨΏ”–“÷÷ΤΉς”ΟΓΘ
1.2 Κœ≥…ΆΨΨΕ
ΡΩ«ΑΜΖΕΰκΡΒΡΚœ≥…ΖΫΖ®÷ς“ΣΖ÷ΈΣ“‘œ¬2÷÷ΓΘ
1.2.1 Μ·―ßΚœ≥…ΆΨΨΕ
”Π”ΟΉνΙψΖΚΒΡΖΫΖ® «ΙΧœύΚœ≥…Ζ®Θ§ΤδΚœ≥…Νς≥Χ»γΆΦ1Υυ ΨΓΘ¥ΥΖΫΖ®ΒΡ”≈Βψ «Α±ΜυΥαΆ®Ιΐ ς÷§’β“ΜΟΫΫιΜΖΜ·‘ΌΖ÷άκΘ§Ω…“‘Κή”––ßΒΊ±ήΟβΖ÷Ή”ΦδΒΡΤδΥϊΖ¥”ΠΘ§¥”ΕχΡήΙΜ…ζ≥…ΗϋΕύΒΡΜΖΕΰκΡ[7]ΓΘ
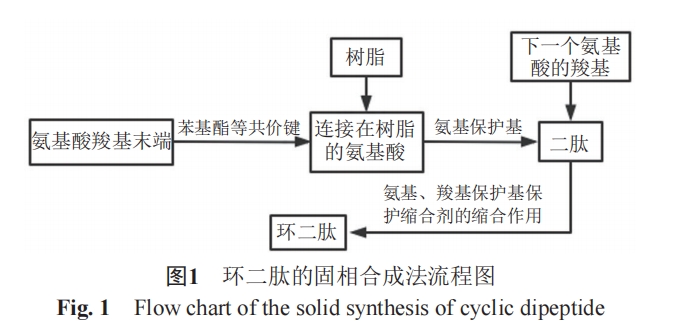
ΕχΚœ≥…ΜΖΕΰκΡΒΡΨ≠ΒδΖΫΖ® «»ή“ΚΚœ≥…Ζ®ΓΘΦ¥ΫΪœΏ–‘κΡΖ≈÷Ο”Ύ»ή“Κ÷–Θ§≤Δ Ι”ΟΥθΚœΦΝ ΙΤδΝ§Ϋ”≥…ΜΖΘ§“≤Ω…“‘ΫΪœΏ–‘κΡΒΡCΕΥΆ®ΙΐΝρθΞΖ®Β»ΜνΜ·»ΜΚσ‘ΌΫχ––≥…ΜΖΓΘ”≈Βψ‘Ύ”Ύ±ήΟβΑ±ΜυΥαΒΡ“λΙΙΜ·[8]ΓΘ
1.2.2 …ζΈοΚœ≥…ΆΨΨΕ
ΤυΫώΈΣ÷ΙΚœ≥… ΉΈ≤œύΝ§ΜΖκΡΒΡΆΨΨΕ“ΜΑψΑϋΚ§2÷÷ΓΘΒΎ“Μ÷÷ΈΣΖ«ΟΗΚœ≥…ΆΨΨΕΓΘάΐ»γΡΎ‘¥–‘cyclo(His-Pro)ΒΡΚœ≥…[9]ΓΘ Ήœ» «TRH-Gly(pGlu-His-Pro-Gly)±ΜΫΙΙ»Α±ΥαΑ±ΜυκΡΟΗΝ―Ϋβ…ζ≥…His-Pro-GlyΘ§Κσ’Ώ‘ΌΨ≠ΙΐΖ«ΟΗ¥ΌΜΖΜ·Κœ≥…cyclo(His-Pro)ΓΘΝμ“Μ÷÷ΈΣΟΗΚœ≥…ΆΨΨΕΘ§Φ¥κΡΝ¥±Ί–κ‘ΎΟΗΒΡ≤Έ”κœ¬≤≈ΡήΫχ––Ή‘…μΜΖΜ·ΓΘΤδΑϋΚ§NRPSs(non-ribosomal peptide synthetase, NRPS)“‘ΦΑCDPSs(cyclodipeptide synthase, CDPS)2÷÷Κœ≥…ΆΨΨΕΓΘNRPSsΚœ≥…ΆΨΨΕΦ¥Ζ«ΚΥΧ«ΧεκΡΚœ≥…ΆΨΨΕ[10]Θ§NRPSs”…œΌή’ΥαΜ·ΫαΙΙ”ρΓΔκΡθΘ‘ΊΧεΒΑΑΉΫαΙΙ”ρΚΆΥθΚœΫαΙΙ”ρ3÷÷ΫαΙΙ”ρΉι≥…ΓΘNRPSsΚœ≥…ΆΨΨΕ»γΆΦ2A ΨΓΘNRPSsΆΨΨΕ”≈Βψ «ΤδΨΏ”–Εύ―υ–‘Θ§¥”Εχ ΙΖ«ΚΥΧ«ΧεκΡΒΡΫαΙΙΚΆΙΠΡή“≤”–Εύ―υ–‘Θ§“ρ¥ΥΩ…“‘ Ι”Ο≤ΜΆ§ΒΡΒΉΈοΚœ≥…ΜΖΕΰκΡΓΘCDPSsΚœ≥…ΆΨΨΕΦ¥ΜΖΕΰκΡΚœΟΗΚœ≥…ΆΨΨΕ[11]Θ§CDPSsΚœ≥…ΆΨΨΕ»γΆΦ2BΥυ ΨΘ§ΗΟCDPSsΆΨΨΕ”≈Βψ «ΡήΙΜάϊ”Οœ÷”–ΒΡaa-tRNAΚœ≥…ΜΖΕΰκΡΘ§Υυ–ηΝς≥ΧΫœ…Ό[12]ΓΘ
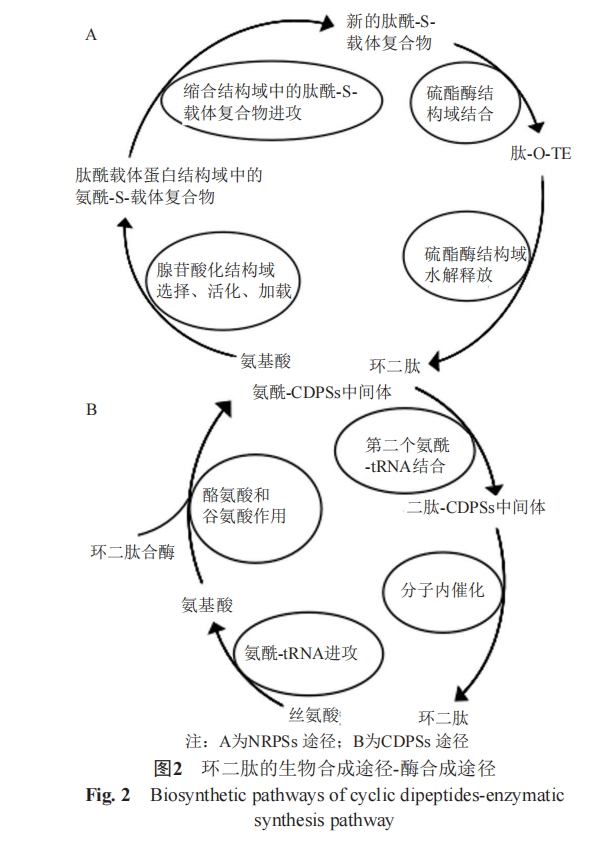
2 ΜΖΕΰκΡΒΡΫαΙΙ”κάμΜ·–‘÷
ΜΖΕΰκΡΨΏ”–ΕάΧΊΒΡΫαΙΙΗ≥”ηΤδ2Ηω«βΙ©Χε”κ«β ήΧεΒΡΧΊ’ςΓΘ”≈ ΤΖ÷Ή”÷ßΦήΆ®≥Θ±ΜΕ®“εΈΣ“Μ÷÷ΚΥ–ΡΜ·―ßΫαΙΙΓΘΤδΆ®ΙΐΫαΚœΗς÷÷ΙΌΡήΆ≈ά¥…ζ≥…ΫαΙΙΕύ―υΒΡ…ζΈοΜν–‘Ζ÷Ή”[13-16]ΓΘ±»»γΖΕΒ¬ΜΣΝΠ‘Ύ’β÷÷ΕάΧΊΒΡΡΘ Ϋ«ΐΕ·œ¬≤ζ…ζΒΡ«ΩΖ÷Ή”ΦδœύΜΞΉς”Ο ΙΜΖΕΰκΡΨΏ”–ΫœΗΏΒΡΖ÷Ή”Ή‘ΉιΉΑ«ψœρΘΜ―–ΨΩ±®ΒάKozlovskyΒ»[17]”ΟΏ≈ΏαΜυΆ≈–ό ΈDKPsΘ§Ζ÷άκΒΟΒΫΒΡpiscarine A(24)ΚΆpiscarine B(25)‘ΎDKPΫαΙΙ≤ύΝ¥…œΨΏ”–»ΐΜΖΜρΥΡΜΖΏ≈ΏαΜυ‘”ΜΖΘ§‘ΎΗ§Α±ΥαΜΖ…œΨΏ”–≤Μ±ΞΚΆΥΪΦϋΓΘ Β―ιΫαΙϊ≥θ≤Ϋ±μΟς’β2÷÷Μ·ΚœΈοΕ‘«ΑΝ–œΌΑ©œΗΑϊœΒΨΏ”–“ΜΕ®ΒΡœΗΑϊΕΨ–‘ΓΘ¥ΥΆβΘ§Μυ”ΎΜΖΕΰκΡ±Ψ…μΒΡΗ’–‘ΫαΙΙΚΆΖ÷Ή”ΦδœύΜΞΉς”ΟΒ»Θ§Ω…“‘Ά®Ιΐ…ζΈοΓΔΜ·―ßΆΨΨΕΚœ≥…ά¥ Βœ÷ΤδΫαΙΙΒΡΕύ―υ–‘ΓΘ“―÷Σ–μΕύΜΖΕΰκΡΒΡΧλ»Μ≤ζΈοΨΏ”–ΩΙ÷ΉΝωΓΔΩΙΨζΚΆΩΙ―θΜ·Β»Μν–‘Θ§ΕχΤδΙΧ”–ΒΡ…ζΈοΜν–‘Ηϋ «¥ΌΫχΝΥΤδ‘Ύ“©ΈοΚœ≥…ΚΆ“©Έο δΥΆΒ»Νλ”ρΒΡ―–ΨΩΖΔ’ΙΓΘΫϋΡξά¥Θ§CDP“――ΗΥΌ≥…ΈΣ…ηΦΤ…ζΈο≤ΡΝœΓΔœ»ΫχΚρ―Γ“©ΈοΚΆ“©Έο δΥΆΦΝΒ»ΒΡΉ®”Ο÷ßΦή[18]ΓΘΨΏ”–Εύ÷÷ΙΠΡήΒΡDKPsΒΡ¥ζ±μ–‘Μ·―ßΫαΙΙ»γΆΦ3Υυ ΨΘ§άΐ»γ≤Έ”κœΗΑϊΦδ–≈œΔ¥ΪΒί(1~4)ΓΔΩΙ÷ΉΝωΜν–‘(5~11)ΓΔΩΙΨζΜν–‘(12~21)ΓΔΩΙ―θΜ·”κΫΒ―ΣΧ«(22)ΓΔΉςΈΣΡΐΫΚ“ρΉ”Ήι≥…≥§Ζ÷Ή”Υ°Ρΐ(23)ΓΔΒςΩΊ ≥”ϊΦΑΡήΝΩ¥ζ–Μ(24)ΓΔ“÷÷ΤœΗΑϊ÷ήΤΎ(25~26)ΓΔΗΡ…Τ…ώΨ≠œΒΆ≥(13)ΓΘ
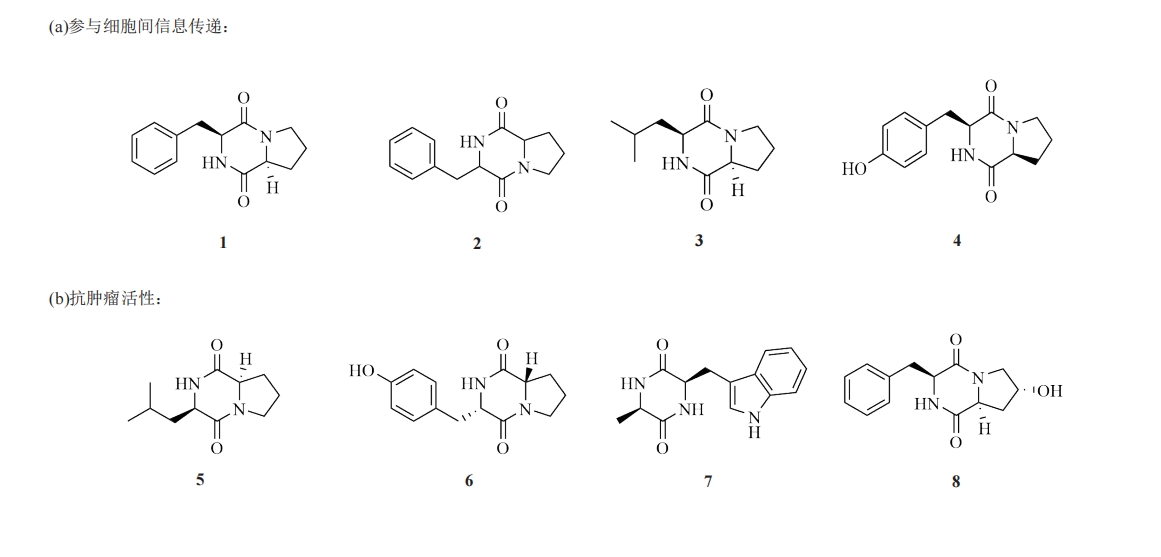
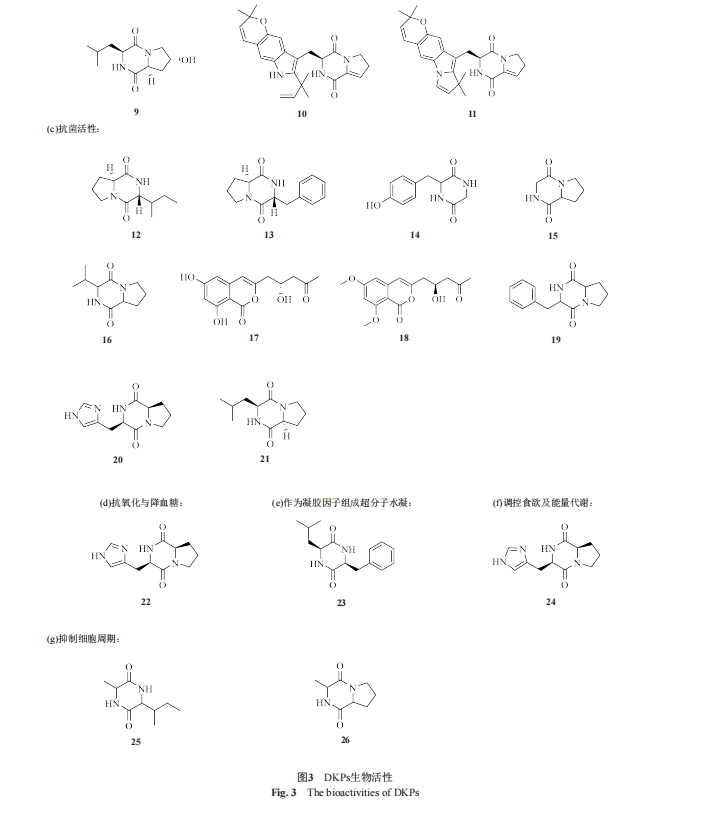
¥σΕύ ΐΒΡΜΖΕΰκΡ≥ »θΦν–‘Θ§≥Θ≥ΘΈΔ»ή”ΎΥ°Θ§“Ή»ή”ΎΦΉ¥ΦΒ»”–Μζ»ήΦΝ÷–Θ§–ΈΧ§“ΜΑψΈΣΫαΨßΜρΖέΡ©Θ§»έΒψ“ΜΑψΗΏ”Ύ200 Γφ[19]ΓΘΆ®≥Θ≤…”Ο«Ω―θΜ·ΦΝ»ΞΦλ≤βΜΖΕΰκΡΒΡ¥φ‘ΎΘ§ΜΖΕΰκΡ”κ≤ΜΆ§ ‘ΦΝΖ¥”ΠΜα≥ œ÷≥ω≤ΜΆ§―’…ΪΒΡΑΏΒψ[20]Θ§άΐ»γ”ΪΙβ÷Η ΨΦΝΓΔΒβΜ·νιΦΊ ‘ΦΝΓΘ
3 ΜΖΕΰκΡΒΡ…ζΈοΜν–‘
3.1 ≤Έ”κœΗΑϊΦδ–≈œΔ¥ΪΒί
ΫϋΡξά¥ΒΡ―–ΨΩ±μΟςΘΚ»ΚΧεΗ–”Π(quorum sensingΘ§QS) «œΗΨζΆ®ΙΐΉ‘”’ΒΦΈο÷ (auto-inducerΘ§AI)Φύ≤βΤδ÷÷»ΚΟήΕ»Θ§”Ο”ΎΒςΫΎ…ζΈοΒΡ»ΚΧε––ΈΣΒΡ“Μ÷÷–≈œΔ¥ΪΒίΜζ÷ΤΓΘ»γ…ζΈοΡΛ–Έ≥…ΓΔ÷ζœΗΨζΗ–»ΨΓΔΖ÷ΟΎΕΨΝΠ“ρΉ”Β»[21]ΓΘΡΩ«ΑΘ§Ήν≥ΘΦϊΒΡ «ΗΏΥΩΑ±ΥαΡΎθΞ(acylhomoserine lactonesΘ§AHL)–≈Κ≈¥ΪΒίΆΨΨΕΘ§‘Ύ»ΚΧεΗ–”ΠΆ®¬Ζ÷–Θ§–≈Κ≈Ζ÷Ή”AHLs”κLuxR–ΆΒΑΑΉΫαΚœΘ§≤ζ…ζΖ¥”ΠΘ§ΒςΫΎœύΙΊΜυ“ρΒΡ±μ¥οΘ§¥”Εχ ΙœΗΨζΫχ––“ΜΕ®ΒΡ…ζάμΜνΕ·ΓΘ”…AHLsΫιΒΦΒΡ»ΚΧεΗ–”ΠœΒΆ≥ΒΡΈο÷ ±Μ»œΕ®ΈΣ»ΚΧεΗ–”Π“÷÷ΤΦΝΓΘΨί±®ΒάΘ§DKPsΡήΙΜΒςΫΎLuxR–Ά ήΧεΜν–‘ΓΘ»γΘ§¥¥…ΥΜΓΨζ(Vibrio vulnificus)≤ζ…ζΒΡcyclo(L-Phe-L-Pro)Ω…“‘”’ΒΦΖ― œΜΓΨζV. fischeriluxΜυ“ρΒΡ±μ¥ο[22]ΓΘDKPsΜΙΩ…“‘”––ßΧαΗΏœύΙΊΜυ“ρΒΡΉΣ¬ΦΥ°ΤΫΓΘάΐ»γΘ§‘ΎΤχΈΕ…≥άΉΨζ(Serratia odorifera)÷––¬ΖΔœ÷ΝΥ“Μ÷÷QSœΒΆ≥Θ§Τδ÷–cyclo(ProPhe)Β»AIΩ…“‘ΧαΗΏ’φΦßΙΫ(H. marmoreus)ΡΨ÷ ΥΊΫΒΫβΟΗΜυ“ρΒΡΉΣ¬ΦΥ°ΤΫ[23]ΓΘΨΏΧεΒΡœύΙΊ–≈Κ≈Ζ÷Ή”ΦΑΜζ÷ΤΜΙ¥ΐΫχ“Μ≤Ϋ―–ΨΩΓΘ
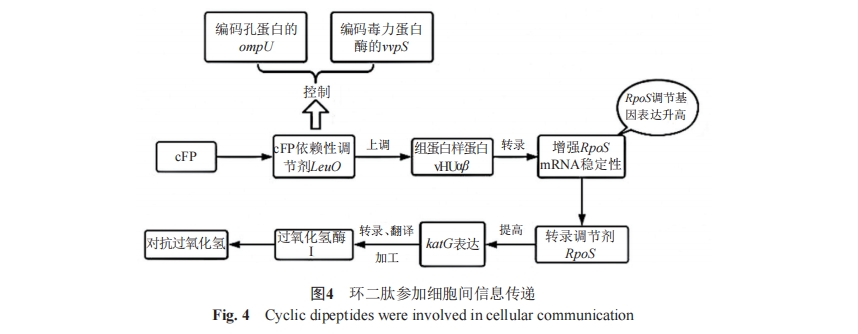
¥¥…ΥΜΓΨζ≤ζ…ζΒΡcyclo(L-phe-L-Pro)(CFP)Θ§ΉςΈΣ“Μ÷÷–≈Κ≈Ζ÷Ή”ΩΊ÷Τœ»ΧλœΗΨζΜυ“ρΒΡtoxR“άάΒ–‘±μ¥οΘ§Ά§ ±“≤ «“Μ÷÷ΕΨΝΠ“ρΉ”Θ§Ά®ΙΐΧαΗΏœΗΑϊΡΎΜν–‘―θΥ°ΤΫΕ‘»ΥάύœΗΑϊ≤ζ…ζ÷¬≤ΓΉς”ΟΓΘKimΒ»[24]‘Ύ2018ΡξΖΔœ÷CFP¥ΌΫχΝΥ¥¥…ΥΜΓΨζΕ‘Ιΐ―θΜ·«βΒΡ±ΘΜΛΓΘΥϋΡήΆ®ΙΐΧαΗΏRpoSΒΡΥ°ΤΫά¥”’ΒΦkatGΒΡ±μ¥οΘ§ ΙΒΟΙΐ―θΜ·«βΟΗIΒΡΚœ≥…‘ωΦ”Θ§¥οΒΫΫβΕΨH2O2“‘ΩΥΖΰ―θΜ·”ΠΦΛΒΡΡΩΒΡΓΘ≤Δ«“ΖΔœ÷CFP“άάΒ–‘ΒςΫΎΤςLeuOΡή÷±Ϋ””κvhuAΚΆvhuBΒΡ…œ”Έ«χ”ρΫαΚœ“‘‘ω«ΩΉΣ¬ΦΘ§ΫχΕχΖΔΜ”CLP…œΒςΉιΒΑΑΉ―υΒΑΑΉvHUΠΝΚΆvHUΠ¬ΒΡΉς”ΟΓΘCFPΫιΒΦΒΡToxR-LeuO-vHUΠΝΠ¬-RpoSΆ®¬Ζ“≤…œΒςΝΥ“―÷ΣΒΡ ήRpoSΒςΩΊΒΡΜυ“ρΘ§Χα ΨCFPΕ‘ ήRpoSΚΆLeuOΒςΩΊΒΡΜυ“ρΕΦΨΏ”–ΒςΫΎΉς”ΟΘ§CFPΉςΈΣΕΨΝΠ“ρΉ”ΚΆΆ§‘¥≤Γ‘≠ΧεΒΡ±ΘΜΛ–≈Κ≈ΖΔΜ”Ή≈÷Ί“ΣΙΠΡήΘ§»γΆΦ4Υυ ΨΓΘ
ΫϋΡξά¥Θ§ΈΣΝΥΫβΨω≤Γ‘≠ΨζΒΡΡΆ“©–‘»’“φ―œ÷ΊΒΡΈ ΧβΘ§»ΥΟ«‘Ϋά¥‘ΫΙΊΉΔΆ®Ιΐ“÷÷Τ…ζΈοΡΛΒΡ–Έ≥…ΚΆΕΨΝΠ“ρΉ”ΒΡΚœ≥…Β»ΆΨΨΕΉηΑ≠ΤδΫχΜ·ΈΣΡΆ“©Ψζ÷ξΒΡΖΫΖ®ΓΘ2019ΡξΘ§ShanmugarajΒ»[25]―–ΨΩΝΥcyclo(LLeu-L-Pro)(CLP)Ε‘πΛ÷ …≥άΉΨζ(Serratia marcescens)ΒΡΩΙQS–ßΙϊΓΘΥϊΟ«Ϋχ––ΝΥΧεΡΎΓΔΧεΆβΒΡΕύ÷÷ Β―ι―–ΨΩΘ§Τδ÷–Αϋά®Ήν–Γ“÷Ψζ≈®Ε»(minimum inhibitory concentrationΘ§MIC) ‘―ιΓΔΒΑΑΉΟΗ ‘―ιΓΔ”ΈΕ·ΚΆ»ΚΦ·‘ΥΕ· ‘―ιΘ§–ψάω“ΰΗΥœΏ≥φ(Caenorhabditis elegans)ΡΘ–ΆΒ»ΓΘ≤ΔΆ®ΙΐqPCRΦΦ θ÷Λ ΒΝΥCLP»Ζ ΒΡή”––ß“÷÷ΤπΛ÷ …≥άΉΨζ÷–QSΩΊ÷ΤΒΡΝιΗΥΨζΥΊΒΡ≤ζ…ζΓΘ‘ΎΖ÷Ή”Υ°ΤΫ…œΘ§Ά®Ιΐœ¬ΒςfimAΚΆfimCΒΡ±μ¥οΘ§Φθ…ΌΨζΟΪΒΑΑΉΒΡΚœ≥…Θ§ΫχΕχ“÷÷ΤπΛ÷ …≥άΉΨζΒΡΗΫΉ≈Θ§ΉηΑ≠…ζΈοΡΛΒΡΉιΉΑΓΘ¥ΥΆβΘ§CLPΜΙœ¬ΒςΝΥFlhDΓΔFlhCΚΆbsmBΜυ“ρΒΡ±μ¥οΘ§ΖΔΜ”“÷÷ΤπΛ÷ …≥άΉΨζΒΡ”Έ”ΨΚΆ»ΚΦ·‘ΥΕ·“‘ΦΑΖΝΑ≠πΛ÷ …≥άΉΨζΒΡΧεΡΎΗ¥÷ΤΚΆ“Τ÷≤ΒΡΙΠ–ßΓΘΤδΫαΙϊ±μΟςCLPΕ‘…ζΈοΡΛΒΡ“÷÷ΤΉς”Ο «ΩΣΖΔ…ζΈοΡΛΦΝΒΡΜυ¥ΓΓΘ2020ΡξΘ§¥”ΟΉ«ζΟΙ(Aspergillus oryzae)ΒΡ¥ζ–Μ≤ζΈο÷–Ζ÷άκΒΟΒΫΒΡΜΖΕΰκΡΕ‘”ΪΙβΦΌΒΞφΏΨζ(Pseudomonas fluorescens)ΒΡ»ΚΧεΗ–”Π±μ–ΆΨΏ”–ΚήΚΟΒΡ“÷÷ΤΉς”ΟΘ§”…¥ΥΆΤ≤βΜΖΕΰκΡΩ…Ρή «“Μ÷÷–¬»ΚΧεΗ–”Π“÷÷ΤΦΝ[26]ΓΘ2021ΡξΘ§¥”ΚΘ―σ’φΨζDXY-1¥÷ΧαΈο÷–Ζ÷άκΦχΕ®≥ωΒΡcyclo(L-Tyr-L-Pro)Ε‘Ήœ…Ϊ…ΪΗΥΨζ(Chromobacterium violaceum CV026)ΚΆΆ≠¬ΧΦΌΒΞΑϊΨζ(P. Aeruginosa PA01)±μœ÷≥ωœ‘÷χΒΡQSΒςΩΊΒΡΕΨΝΠ“÷÷ΤΉς”Ο[27]ΓΘΜΖΕΰκΡ”–Άϊ≥…ΈΣ“ΜΗωΩΙΨζ“©ΈοΩΣΖΔΒΡ–¬ΒΡ«±‘ΎΑ–ΒψΓΘ
3.2 ΩΙ÷ΉΝωΜν–‘
ΨίΆ≥ΦΤΘ§‘ΎΈ“ΙζΨΓΙή ≥ΙήΑ©ΓΔΗΈΑ©Β»Α©÷ΔΒΡ÷ΉΝωΥάΆω¬ ÷πΡξΫΒΒΆΘ§ΖΈΑ©ΓΔ»ιœΌΑ©ΓΔΫα÷±≥ΠΑ©ΓΔΙ§Ψ±Α©ΚΆ«ΑΝ–œΌΑ©ΒΡΖΔ≤Γ¬ »‘»Μ≥ œ÷…œ…ΐΒΡ«ς ΤΓΘ―–ΖΔ’κΕ‘–‘ΒΡΑ–œρ“©ΈοΒΡΫ≈≤ΫΩΧ≤Μ»ίΜΚ[28]ΓΘ
2020ΡξΘ§LinΒ»[29]‘Ύ―–ΨΩ ±ΖΔœ÷cyclo(L-Leu-LPro)‘Ύ20 ΠΧmol/L…œ±μœ÷≥ωΕ‘ECA-109ΓΔHeLa-S3ΚΆPANC-1Β»3÷÷Α©œΗΑϊœΒ÷–Β»ΒΡ“÷÷ΤΉς”ΟΘ§ΖΕΈß¥”14%(pan-1)ΒΫ55%(ECA-109)ΓΘKaranamΒ»[30]ΒΡ―–ΨΩ±®Βά÷–ΧαΒΫcyclo(Pro-Tyr)(DP)ΨΏ”–”’ΒΦœΗΑϊΒρΆωΒΡΜν–‘Θ§ΤδΕ‘HepG2œΗΑϊ”–ΦΝΝΩ“άάΒ–‘ΒΡœΗΑϊΕΨ–‘Ής”ΟΘ§ΕχΕ‘–Γ σ≥…œΥΈ§McCoyœΗΑϊΈόœ‘÷χΕΨ–‘Ής”ΟΘ§ΤδΉς”ΟΜζ÷Τ»γΆΦ5Υυ ΨΓΘΜΖΕΰκΡΒςΩΊœΗΑϊ…ζ≥ΛΚΆΒρΆωΈΣΩΙΑ©ΧαΙ©ΝΥ“ΜΧθ«εΈζΒΡΥΦ¬ΖΓΘ”…¥ΥΩ…÷ΣΘ§DPΩ…Ρή≥…ΈΣ“Μ÷÷«±‘ΎΒΡ÷ΈΝΤΗΈœΗΑϊΑ©ΒΡΧφ¥ζ“©ΈοΓΘJinendiranΒ»[31]‘ΎΧεΆβΚΆΧεΡΎΑΏ¬μ”ψ“Τ÷≤¥σ≥ΠΑ©HT-29œΗΑϊΡΘ–Ά÷–Θ§ΤάΦέΝΥ4÷÷ά¥Ή‘”Ύ““θΘΕΧΗΥΨζ(E. acetylicum S01)Χα»ΓΈοΒΡDKPsΒΡΩΙΑ©«±ΝΠΓΘ4÷÷DKPsΕ‘HT-29œΗΑϊΜν–‘“÷÷ΤΉς”Ο≥ ΦΝΝΩ“άάΒ–‘Θ§‘ΎΖ«÷¬ΥάΦΝΝΩΖΕΈßΡΎΘ§’β–©DKPsœ‘÷χ“÷÷ΤΑΏ¬μ”ψ“λ÷÷“Τ÷≤ΝωΡΘ–ΆΒΡ÷ΉΝωΫχ’ΙΘ§≤Δ«“Ε‘–Γ σ’ΐ≥Θ≥…œΥΈ§3T3œΗΑϊΈόœΗΑϊΕΨ–‘Ής”ΟΓΘΗΟ―–ΨΩΫαΙϊΧα ΨΘΚ’β4÷÷ΜΖΕΰκΡ‘ΎΩΙΑ©“©Έο―–ΖΔΖΫΟφΚή”–«ΑΨΑΓΘ2021ΡξΘ§¥”ΦΪΒΊ’φΨζ(Geomyces pannorum SA3-2-YM)ΒΡΖΔΫΆ“ΚΧα»ΓΈο÷–ΖΔœ÷ΒΡcyclo(D-Ala-D-Trp)Ψ≠ Β―ι≤βΒΟΕ‘HeLaœΗΑϊΒΡ“÷÷Τ¬ Ω…¥ο62.28%[2]ΓΘ2022ΡξΘ§YoussefΒ»[32]ΖΔœ÷cyclo(L-NMe-Leu-L-NAc-Phe)ΚΆcyclo(D-6-OH-Pro-DPhe)Ε‘HCT 116ΒΡ―Γ‘ώ–‘”≈”ΎMDA-MB-231(»Υ»ιœΌΑ©œΗΑϊ)Θ§ΕχΕ‘HeLaœΗΑϊΒΡΉς”ΟΫœ»θΘΜΤδ÷–cyclo(LNMe-Leu-L-NAc-Phe)Ε‘HCT 116Μν–‘Ήν«ΩΓΘ
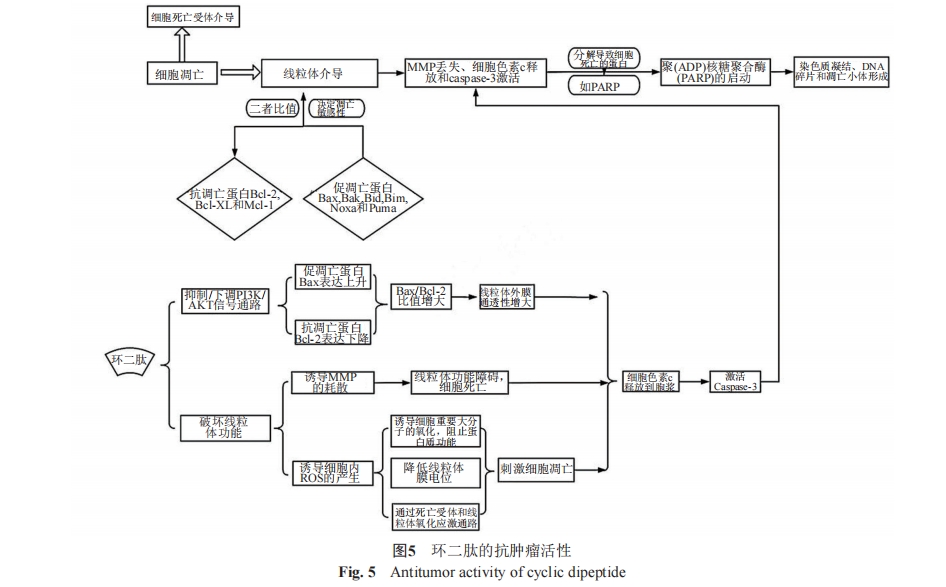
Ά®Ιΐ Β―ι ΐΨίΩ…“‘ΖΔœ÷ΝΫΗω”ΑœλΕΨ–‘Ής”ΟΒΡ“ρΥΊΓΘ Ήœ» «Α©œΗΑϊΒΡάύ–ΆΘ§YeΒ»[33]¥”Κλ ςΝ÷Ν¥ΟΙΨζ(Mangrove Streptomyces sp. Q24)÷–Ζ÷άκ≥ωά¥ΒΡcyclo(L-Phel-L-Hyp)‘Ύ»ΥΫΚ÷ ΝωU87-MGΚΆU251œΗΑϊΒΡ‘ω÷≥“÷÷Τ Β―ι÷–Θ§Ζ÷±πΒ±IC50ΈΣ5.8 ΚΆ18.6 ΠΧmol/L ±±μœ÷≥ω“ΜΕ®–ßΝΠΘ§Εχ‘ΎHCT-116ΓΔOVCAR-8ΚΆSF-295œΗΑϊœΒ…œ≤β ‘ ±ΖΔœ÷‘Ύ25 µg/mL[34]…œ“≤ΈόΖ®œ‘ Ψ≥ωΟς»ΖΒΡœΗΑϊΕΨ–‘Ής”ΟΓΘ≥ΐΝΥΑ–œρΑ©œΗΑϊΒΡάύ±π‘λ≥…ΒΡ≤ν“λΆβΘ§ΜΖΕΰκΡ±Ψ…μΒΡΫαΙΙΕ‘ΕΨ–‘Ής”ΟΒΡ≤ζ…ζ“≤”–”ΑœλΘ§άΐ»γcyclo(L-Leu-L-Hyp)ΦΑΤδΫαΙΙάύΥΤΈοcyclo(L-Leu-L-Pro)Ε‘U251ΚΆU87-MGœΗΑϊœΒΒΡΕΨ–‘ Β―ι÷–Θ§Εΰ’ΏΖ÷±π‘ΎIC50ΈΣ29.4”κ14.5 ΠΧmol/L“‘ΦΑIC50ΈΣ19.8”κ1.3 ΠΧmol/L[33] ±Ε‘ΗΟœΗΑϊœΒ±μœ÷≥ωΕΨ–‘Ής”ΟΓΘ“‘…œ±μΟςΘ§ΜΖΕΰκΡΥδ»ΜΨΏ”–ΩΙ÷ΉΝωΒΡ…ζάμΧΊ–‘Θ§ΒΪ «ΜΖΕΰκΡάύ(CDPs)Μ·ΚœΈο≤ΜΡήΕ‘Υυ”–ΒΡ÷ΉΝωœΗΑϊΕΦΨΏ”–Ϋœ«ΩΒΡ“÷÷ΤΉς”ΟΘ§ΤδΗϋ«ΩΒΡΕΨ–‘Ής”ΟΧεœ÷‘ΎœΗΑϊΧΊ“λ–‘ΖΫΟφ[35]ΓΘ
ΫϋΡξά¥Θ§ΙΊ”ΎΚ§ΝρDKPsΒΡ―–ΨΩ±®ΒάΝΥ“Μ–©ΜΖΕΰκΡΕ‘Α©œΗΑϊΒΡΕΨ–‘Ής”ΟΘ§’β–©DKPs¥σΕύά¥Ή‘”Ύ’φΨζ[36]ΓΘΙΎΟΪΗΥΨζ(C. cristatum)÷–ΖΔœ÷ΒΡCristazineΆ®ΙΐΥάΆω ήΧεΆΨΨΕ”’ΒΦœΗΑϊΒρΆωΘ§Ε‘HeLaΚΆA451ΨΏ”–œΗΑϊΕΨΉς”Ο[32-37]ΓΘ“Ε¥Ώ≤ζΥΊ «“Μ÷÷Κ§ΕΰΨέΝρΒΡDKPsΘ§Ε‘ ΒΧε÷ΉΝωΚΆ―Σ“Κ÷ΉΝωΕΦ±μœ÷≥ωΙψΖΚΒΡœΗΑϊΕΨ–‘Ής”Ο[38-39]Θ§‘ΎLaiΒ»[40]ΒΡ―–ΨΩ÷–Θ§Τδ‘ΎΦ±–‘ΥηœΒΑΉ―Σ≤ΓœΗΑϊHL-60ΓΔU937ΚΆKG-1a÷–”’ΒΦΖ÷Μ·Θ§≤Δ”κΤδΥϊ±μΙέ“≈¥Ϊ“©Έοœ‘ Ψ–≠Ά§œΗΑϊΕΨ–‘ΓΘ≥ΐΝΥœΗΑϊΕΨ–‘Ής”ΟΘ§≤ΩΖ÷DKPsΜΙΡή“÷÷Τ“Μ–©÷ΉΝωœύΙΊΈο÷ ΒΡΚœ≥…[36]ΓΘBRD4ΒΑΑΉΨ≠≥Θ‘ΎΗς÷÷άύ–ΆΒΡ÷ΉΝω÷–±μ¥οΘ§‘ΎYuΒ»[41]ΒΡ±®Βά÷–Θ§Μυ”ΎΗ§Α±ΥαΒΡΦΗ÷÷ΈεΜΖΓΔΝυΜΖΜρΕΰΨέΧεDKPsΩ…Ρή «BRD4ΒΑΑΉ(Κ§δε”ρΒΑΑΉ4)ΒΡ”––ß“÷÷ΤΦΝΓΘ
3.3 ΩΙΨζ”κ“÷ΨζΜν–‘
SongΒ»[42]‘Ύ2018ΡξΖ÷άκ≤ΔΦχΕ®ΝΥ3÷ξΕ‘«―ΩΤάΉΕϊ œΨζ(Ralstonia Solanacearum GMI1000)ΨΏ”–ΗΏΩΙΨζΜν–‘ΒΡ–¬Ψζ÷ξΓΘ‘Ύ«ΑΤΎΙΛΉς÷–ΖΔœ÷¥σ≥ΠΑΘœΘΨζ(Escherichia coli GZ-34)Ε‘«―ΩΤάΉΕϊΨζœύΕ‘Ζά÷Έ–ßΙϊΉνΦ―Θ§¥οΒΫ82.27%Θ§«“Φ≤≤Γ÷Η ΐΫΒΒΆ÷ΝΉν–ΓΘ§¥οΒΫ16.67ΓΘ―–ΨΩ»Υ‘±Ε‘¥σ≥ΠΑΘœΘΨζGZ-34ΒΡΩΙ«ύΩί≤ΓΜν–‘ΉωΫχ“Μ≤Ϋ―–ΨΩΓΘ‘ΎΜν–‘―–ΨΩ÷–ΖΔœ÷cyclo(L-Pro-DIle)ΚΆcyclo(L-Pro-L-Phe) «¥σ≥ΠΑΘœΘΨζGZ-34≤ζ…ζΒΡ÷ς“ΣΩΙ«ύΩί≤ΓΜν–‘Έο÷ ΓΘ«“Κσ’Ώ «ΗΟΨζ÷ξΕ‘ΒΨΈΝ≤ΓΨζ(Magnaporthe grisea)φΏΉ”–Έ≥…ΒΡ“÷÷ΤΜν–‘ΒΡΙΊΦϋ≥…Ζ÷ΓΘ’β–©ΖΔœ÷Χα ΨΘ§¥σ≥ΠΑΘœΘΨζ÷–ΒΡΜΖΕΰκΡΩ…Ρή±ΜΩΣΖΔΈΣ–¬ΒΡ«±‘ΎΩΙΨζΦΝΘ§”Ο”Ύ≈©“ΒΖά÷ΈΓΘ
2020ΡξΙυ«λΖαΒ»[3]‘Ύ―–ΨΩ“÷ΨζΜν–‘÷–ΖΔœ÷ΘΚΜ·ΚœΈοcyclo(Tyr-Gly)ΓΔcyclo(Pro-Gly)ΚΆcyclo(Pro-Val)Ε‘¥σ≥ΠΑΘœΘΨζ”–Ϋœ«ΩΒΡ“÷÷ΤΉς”ΟΘ§MICΖ÷±πΈΣ16.0ΓΔ4.0ΚΆ8.0 µg/mLΓΘ2022ΡξΘ§Youssef Β»[43]‘ΎΕ‘«ζΨζ τ÷––¬ΖΔœ÷ΒΡΝΫ÷÷Μ·ΚœΈοΫχ––ΒΡ‘≤≈Χά©…Δ Β―ιΫαΙϊ±μΟςΘΚcyclo(L-NMe-Leu-L-NAc-Phe)ΚΆcyclo(D-6-OH-Pro-D-Phe)Ε‘¥σ≥ΠΑΘœΘΨζΚΆΫπΜΤ…ΪΤœΧ―«ρΨζ(Staphylococcus aureus)ΒΡ“÷ΨζΉς”ΟΗΏ”ΎΑΉΡν÷ιΨζ(Candida albicans)ΓΘΡΩ«Α“―÷ΣΒΡ±»Ϋœ”––ßΒΡ «cyclo(Phe-Pro)(CFP)ΚΆΈΣcyclo(His-Pro)(CHP)ΓΘΤδ÷–CFP÷ς“Σ «Ά®Ιΐ¥ΧΦΛΆβΡΛΩΉΒΑΑΉOmpUΒΡ±μ¥οΫχΕχΖΔΜ”Ής”ΟΘΜCHP÷ς“ΣΆ®Ιΐ‘ωΦ”ΩΙ¥ΌΒρΆωΒΑΑΉBcl-2ΒΡ±μ¥οΘ§ΫΒΒΆ”…―θΜ·”ΠΦΛΥυ“ΐΤπΒΡœΗΑϊΒρΆωΫχΕχΖΔΜ”Ής”Ο[44]ΓΘ
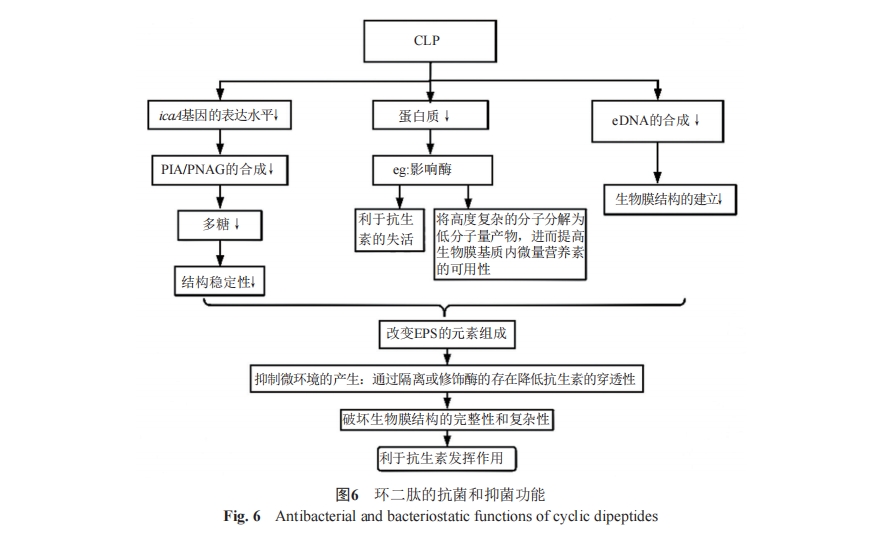
¥ΥΆβΘ§ΜΖΕΰκΡΖΔΜ”ΩΙΨζΓΔ“÷ΨζΧΊ–‘ΒΡΜζ÷Τ÷°“Μ «Ά®Ιΐ“÷÷Τ…ζΈοΡΛΒΡ–Έ≥…ΓΘ2017ΡξΘ§GowrishankarΒ»[6]ΖΔœ÷ΝΥcyclo(L-Leu-L-Pro)(CLP)Ρή“÷÷ΤœΗΨζ…ζΈοΡΛΘ§ΒΪ’β≤ΔΖ«…±ΨζΉς”ΟΓΘΥΐΟ«άϊ”Ο±μΤΛΤœΧ―«ρΨζ(Staphylococcus epidermidis RP62A)“‘ΦΑΗς÷÷Ε®–‘ΚΆΕ®ΝΩΖ÷Έωά¥Ϋβ ΆCLPΒΡΩΙΨζΡΛΙΠ–ßΒΡ…ζΈοΜ·―ßΜυ¥ΓΓΘ“―÷ΣΑϊΆβΨέΚœΈο(extracellular polymeric substances, EPS)ΦΑΙΧΉ≈œΗΑϊ»ΚΩ…“‘Β÷ΩΙΩΙ…ζΥΊΒΡΉς”Ο“‘ΦΑΥό÷ςΒΡΟβ“ΏΖ¥”ΠΓΘCLPΆ®ΙΐΒςΫΎEPSά¥“÷÷Τ±μΤΛΤœΧ―«ρΨζΒΡ…ζΈοΡΛΜζ÷Τ»γΆΦ6Υυ ΨΓΘCLP÷ς“ΣΆ®Ιΐ3ΧθΆΨΨΕ”ΑœλEPSΒΡ‘ΣΥΊΉι≥…ΓΘΒΎ“ΜΘ§CLPœ¬ΒςicaAΜυ“ρΒΡΥ°ΤΫά¥“÷÷ΤPIA/PNAGΒΡΚœ≥…Θ§Φθ…ΌΕύΧ«ΒΡΚœ≥…“‘œς»θΨζΡΛΒΡΫαΙΙΈ»Ε®–‘ΓΘΒΎΕΰΘ§“―÷ΣΨχ¥σΕύ ΐΟΗΒΡ±Ψ÷ «ΒΑΑΉ÷ ΓΘEPSΒΡΦΗ÷÷ΑϊΆβΟΗΒΡ¥φ‘Ύάϊ”ΎΩΙ…ζΥΊΒΡ ßΜνΘ§«“ΤδΡήΫΪΗΏΖ÷Ή”ΨέΚœΈοΖ÷ΫβΈΣΒΆΖ÷Ή”ΝΩ≤ζΈοΘ§ΫχΕχΧαΗΏ…ζΈοΡΛΜυ÷ ΡΎΈΔΝΩ”Σ―χΥΊΒΡάϊ”Ο¬ ΓΘCLPΆ®Ιΐ“÷÷ΤΒΑΑΉ÷ ΒΡΚœ≥…ά¥Ήη÷ΙœύΙΊΟΗΖΔΜ”Ής”ΟΓΘΒΎ»ΐΘ§CLP“÷÷ΤeDNAΒΡΚœ≥…“‘ΉηΑ≠…ζΈοΡΛΫαΙΙΒΡΫ®ΝΔΓΘ‘Ύ Β―ιΙΐ≥Χ÷–Θ§ΥϊΟ« Ήœ»Ϋχ––MIC≤βΕ®Θ§ΫαΙϊœ‘ ΨCLP‘Ύ512 µg/mL≈®Ε» ±Ε‘±μΤΛΤœΧ―«ρΨζRP62AΨΏ”–Ϋœ«ΩΒΡΧεΆβΩΙΨζΉς”ΟΓΘΤδ¥ΈΆ®ΙΐΕ®ΝΩ(XTTΖ÷Έω)ΚΆΕ®–‘(Ι≤ΨέΫΙœ‘ΈΔΨΒΚΆ…®ΟηΒγΉ”œ‘ΈΔΨΒ)Ζ÷ΈωΘ§÷Λ ΒΝΥCLP «Ά®ΙΐΗΡ±δEPSΒΡΙΧ”–ΙΌΡήΆ≈ΚΆΒγΚ…ά¥ΖΔΜ”ΩΙΨζΡΛΉς”ΟΒΡΓΘEPS «±μΤΛΤœΧ―«ρΨζ…ζΈοΡΛ–Έ≥…ΒΡΙΊΦϋ“ρΥΊΘ§Εχ…ζΈοΡΛΒΡ–Έ≥… «±μΤΛΤœΧ―«ρΨζΡΆ“©ΒΡ÷ς“Σ‘≠“ρΓΘ“―÷Σ‘γΤΎΕυΆ·»Θ≤Γ(early-childhood cariesΘ§ECC)ΒΡΜΦ≤Γ¬ ΦΑ―œ÷Ί≥ΧΕ»”κΑΉ…ΪΡν÷ιΨζΚΆ±δ–ΈΝ¥«ρΨζ(Streptococcus mutans)ΒΡΨζΑΏΚΆΆΌ“Κ–·¥χ”–ΙΊΓΘSimonΒ»[45]ΒΡ―–ΨΩ÷Λ ΒΝΥcyclo(L-Phe-L-Tyr)ΓΔcyclo(D-Phe-D-Tyr)Β»Ε‘±δ–ΈΝ¥«ρΨζΚΆΑΉ…ΪΡν÷ιΨζ…ζΈοΡΛ–Έ≥…ΒΡ“÷÷Τ–ßΙϊΘ§ΒΪ≤Μ”Αœλ’φΨζ…ζ≥ΛΓΘ’βΗωΫα¬έΫΪ”–÷ζ”ΎΕυΆ·»Θ≤ΓΒΡΫχ“Μ≤Ϋ÷ΈΝΤΓΘCLPΒΡ…ζΈοΡΛ“÷÷Τ–ßΙϊΩ…ΡήΜαœό÷Τ≤Γ‘≠ΧεΕ‘ΩΙΨζΥΊ/ΩΙ…ζΥΊΒΡΡΆ“©–‘ΓΘ“ρ¥ΥΩΣΖΔ’κΕ‘’β÷÷≤Γ‘≠ΧεΒΡEPSΒΡ÷ΈΝΤ≤Ώ¬‘Ω…Ρή «“Μ÷÷±»ΩΙ…ζΥΊ÷ΈΝΤΗϋ”–«ΑΨΑΒΡΧφ¥ζΖΫΑΗΓΘ
3.4 ΩΙ―θΜ·ΓΔΫΒ―ΣΧ«ΒΡΜν–‘
“―÷ΣCHP τ”ΎΡΎ‘¥–‘ΜΖΕΰκΡΘ§‘ΎΩΙ―θΜ·ΓΔΫΒ―ΣΧ«Β»ΖΫΟφ”–Οςœ‘–ßΙϊΓΘ2016ΡξΘ§Ζ°Κλ–ψΒ»[46]Ε‘Ψ≠ΗΏΈ¬ΗΏ―ΙΜΖΜ·ΒΡ”ώΟΉΒΑΑΉΥ°ΫβΈοΫχ––Ζ÷άκ¥ΩΜ·ΓΘ‘Ύ≤βΕ®ΤδΧεΆβΫΒ―ΣΧ«Μν–‘ΒΡ Β―ι÷–ΖΔœ÷Υ°ΫβΈο÷–ΡήΙΜΚήΚΟΒΊ“÷÷ΤΠΝ-ΤœΧ―Χ«ή’ΟΗΚΆΠΝ-ΒμΖέΟΗΒΡΜν–‘Ήν÷’¥οΒΫΫΒ―ΣΧ«ΡΩΒΡ÷ς“ΣΖ÷Ή” «CHPΘΜ¥ΥΆβΥϊΟ«ΜΙΆ®Ιΐ≤ΔΆ®ΙΐDPPHΉ‘”…Μυ«ε≥ΐΜν–‘ Β―ιΚΆΜΙ‘≠ΝΠ Β―ιΖΔœ÷CHP «‘ΎΤδ÷–Ε‘ΩΙ―θΜ·Τπ÷ς“ΣΉς”ΟΒΡΖ÷Ή”ΓΘ
‘ΎMinelliΒ»[47]ΒΡ―–ΨΩ÷–Ω…“‘ΖΔœ÷Θ§PC12œΗΑϊΆ®Ιΐ±μ¥ο±ΘΜΛΉ‘…μΟβ ή―θΜ·ΦΝΚΆ«ΉΒγΜ·ΚœΈοΫιΒΦΒΡ”ΠΦΛΒΡΜυ“ρά¥Ε‘ΜΖΨ≥Υπ…ΥΉω≥ωΖ¥”ΠΓΘ’β–©Μυ“ρΒΡ”’ΒΦ÷ς“Σ”…Nrf2ΫιΒΦΘ§‘ΎœΗΑϊΚΥ÷–Θ§Nrf2”κ–ΓMafΦ“ΉεΒΡ≥…‘±–Έ≥…“λ‘¥ΕΰΨέΧεΘ§≤Δ Ε±π≤ΔΫαΚœΩΙ―θΜ·Ζ¥”Π‘ΣΦΰ(ARE)ΤτΕ·Ή”–ρΝ–Θ§¥”Εχ¥ΌΫχΩΙ―θΜ·Μυ“ρΒΡΉΣ¬ΦΦΛΜνΘ§»γΆΦ7Υυ ΨΓΘ
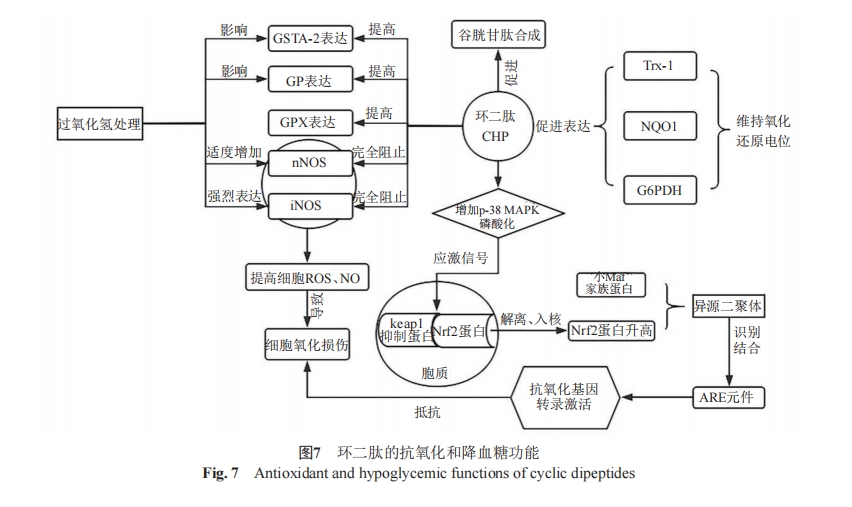
ΤυΫώΈΣ÷ΙΘ§“―”–3÷÷Ω…ΡήΒΡΜζ÷Τά¥Ϋβ ΆCHPΒΡΫΒ―ΣΧ«Ής”Ο[48]ΓΘΔΌ≥Π¥Ό“»ΒΚΥΊ―υΜζ÷ΤΘ§¥ΌΫχ“»ΒΚΥΊΖ÷ΟΎΓΘMoriΒ»[49]‘Ύ≤βΕ®¥σ σ“»œΌΧα»ΓΈο÷–ΒΡCHP ±ΆΤ¬έcyclo(His-Pro)Ω…ΡήΆ®Ιΐ≤Έ”κΕυ≤ηΖ”ΑΖ¥ζ–ΜΓΔ¥ΌΫχ“»ΒΚΥΊΦΑΈΗ≥ΠΦΛΥΊΒΡΖ÷ΟΎά¥≤Έ”κΧ«¥ζ–ΜΓΘΔΎ¥ΌΫχ≥ΠΒάΕ‘–ΩΒΡΈϋ ’“‘ΦΑ¥ΧΦΛΦΓ»βœΗΑϊΕ‘ΤœΧ―Χ«ΒΡΈϋ ’ΒΡΜζ÷ΤΓΘ“―÷ΣGoto-Kakizaki¥σ σΡΘ–Ά «―–ΨΩ”κΖ ≈÷ΈόΙΊΒΡΧ«Ρρ≤ΓΒΡΉνΦ―ΡΘ–ΆΓΘSongΒ»[50]άϊ”Ο¥ΥΡΘ–Ά―–ΨΩ–ΩΦ”CHP‘Ύ“≈¥Ϊ–‘Χ«Ρρ≤Γ÷–ΒΡΩΙΗΏ―ΣΧ«Μν–‘ ±ΖΔœ÷Θ§CHPΡήΆ®Ιΐ¥ΧΦΛΤœΧ―Χ«άϊ”Οά¥ΗΡ…Τ―ΣΧ«¥ζ–ΜΓΘΤδΙΐ≥ΧΩ…Ρή «Ά®Ιΐ”Αœλ“»ΒΚΥΊ ήΧε―«ΜυΒΡΉ‘ΝΉΥαΜ·ά¥¥ΧΦΛ¥σ σΦΓ»βΉι÷·÷–ΒΡΤœΧ―Χ«…ψ»ΓΓΘΔέΜυ”ΎΕ‘Μν–‘τ ΜυΈο÷÷(reactive carbonyl speciesΘ§RCS)ΨΏ”–œ‘÷χΒΡΫβΕΨΉς”ΟΒΡΧΊ–‘Θ§¥ΌΫχΗΏΦΕ÷§÷ ―θΜ·÷’≤ζΈο(advanced lipoxi-dation end-productsΘ§ALEs)ΒΡ…ζ≥…Φθ…ΌΓΘ“―÷Σ≥÷–χΒΡΗΏ―ΣΧ«Υ°ΤΫΜα¥ΌΫχΉ‘”…ΜυΒΡ…ζ≥…Θ§ΕχΉ‘”…ΜυΜα¥ΏΜ·RCSΒΡ…ζ≥…Θ§ΉνΚσ‘λ≥…Π¬œΗΑϊΒΡΙΠΡή’œΑ≠[51]ΓΘ’β–©…ζΈο–ß”ΠΕ‘ΩΙ―θΜ·ΓΔΫΒ―ΣΧ«Β»κΡάύ“©ΈοΒΡ―–ΖΔΨΏ”–Χα Ψ“β“εΓΘ
3.5 ΗΡ…Τ…ώΨ≠œΒΆ≥
ΜΖΕΰκΡΩ…“‘Ά®Ιΐ“÷÷ΤΟΗΒΡΜν–‘ά¥ΗΡ…Τ÷– ύ…ώΨ≠œΒΆ≥ΓΘ…ξάωΒ»[52]‘Ύ―–ΨΩ»Ι¥χ≤ΥΡΎ…ζΨζ(Bacillus sp.QD4)ΖΔΫΆ“ΚΒΡΜ·―ß≥…Ζ÷ ±ΖΔœ÷Θ§‘Ύ400 µg/mL≈®Ε» ±Θ§cyclo(L-Pro-D-Tyr)ΚΆcyclo(L-Pro-L-Tyr)Ε‘AChEΒΡ“÷÷Τ¬ Ζ÷±πΈΣ5.62ΘΞΓά1.63ΘΞΚΆ11.62ΘΞΓά2.29ΘΞΓΘΨί―–ΨΩ±μΟςΘ§≤ΩΖ÷ΑΔΕϊ¥ΡΚΘΡ§≤Γ (alzheimerdiseaseΘ§AD)ΜΦ’ΏΒΡΚΘ¬μΚΆ–¬ΤΛ÷ ΒΡ““θΘΒ®Φν(acetylcholineΘ§ACh)Μαœ‘÷χΦθ…ΌΓΘ…œ ω2÷÷ΜΖΕΰκΡ“÷÷ΤAChEΜν–‘Θ§ΒΦ÷¬AChEΒΡΦθ…ΌΘ§¥οΒΫΟςœ‘ΗΡ…ΤΚΆΜΚΫβADΜΦ’ΏΒΡ÷ΔΉ¥ΒΡ≥…ΙϊΓΘ’βΈΣ―–ΖΔAChE“÷÷ΤΦΝΧαΙ©ΝΥ–¬ΖΫœρΓΘ
¥ΥΆβΘ§ΜΖΕΰκΡΜΙΩ…“‘ΖΔΜ”…ώΨ≠±ΘΜΛΚΆ…ώΨ≠ΒςΫΎΙΠΡήΓΘ“―÷Σcyclo(Gly-Pro)(CGP)Ε‘÷– ύ…ώΨ≠œΒΆ≥”–ΝΦΚΟΉς”ΟΘ§Αϋά®ΩΙΫΓΆϋΓΔ¥ΌΫχΦ«“δΉς”Ο[53]Θ§…ώΨ≠±ΘΜΛΉς”Ο[54]ΓΘKanekoΒ»[55]Ε‘ΩΎΖΰCGP¥ΌΫχ¥σ σ÷–ΖγΡΘ–ΆΒΡ»ΈΈώ―ßœΑ’ΙΩΣ―–ΨΩΓΘ“―÷Σ¥οΒΫ―ßœΑ±ξΉΦΥυ–ηΧλ ΐΒΡ÷ς“Σ”Αœλ“ρΥΊΩ…Ρή «≤Γ±δ«χΒΡ¥σ–ΓΓΘΕχ«“Ρ‘ “œ¬«χ(subventricular zoneΘ§SVZ)…ώΨ≠Η…œΗΑϊ ΐΝΩΒΡ‘ωΦ”±Μ»œΈΣ «ΤΛ÷ Υπ…ΥΚσ–¬…ώΨ≠‘ΣΒΡά¥‘¥ΓΘ‘ΎΗΟ Β―ι÷–Θ§ΥϊΟ«Φύ≤βΒΫ¥σ σ‘ΎΩΎΖΰCGPΚσΥπ…ΥΧεΜΐΦθ–ΓΘ§Η…œΗΑϊ ΐΝΩ‘ωΦ”ΓΘΒΆCGPΉι¥σ σ―ßΜα»ΈΈώΒΡΥυ–η ±ΦδΗϋΕΧΓΘΨί―–ΨΩ±μΟς[56]ΘΚ¥σΕύ ΐΡΎ‘¥–‘IGF-1”κIGFBP-3ΫαΚœΘ§…Ό≤ΩΖ÷”ΈάκΒΡIGF-1Ά®Ιΐ”κIGF-1RΫαΚœΘ§”Αœλ…ώΨ≠‘Σ…ζ≥ΛΓΔ¥φΜνΚΆ…ζ≥ΛΓΘCGPΩ…“‘Ά®Ιΐ”κΡΎ‘¥–‘IGF-1ΨΚ’υΫαΚœIGFBP-3ά¥ΒςΫΎ”ΈάκΒΡIGF-1ΓΘΥϊΟ«»œΈΣ «“ρΈΣΆβ‘¥–‘CGPΒΡΧμΦ”ΒςΫΎΝΥIGF-1ΒΡΝΩΓΘΥπ…Υ«χ”ρΗΫΫϋΒΡIGF-1≈®Ε»‘ωΦ”Θ§¥ΌΫχΝΥ―ßœΑΓΘΗΟ±®Βά Ή¥Έ≤ϊ ωΝΥΩΎΖΰΉνΦ―ΦΝΝΩΒΡCGPΩ…Ά®Ιΐ…ώΨ≠Ω…Υή–‘ΚΆΆχ¬γ÷ΊΙΙΒΡΖΫ ΫΘ§¥ΌΫχΨ÷‘ν–‘»±―Σ–‘ΙΘΥά¥σ σΒΡΗ–Ψθ-‘ΥΕ·ΝΣœκ―ßœΑΘ§ΕχΖ«Ά®Ιΐ…ώΨ≠±ΘΜΛΉς”ΟΓΘΗΟ―–ΨΩΫαΙϊΧα ΨCGPΗχ“©”–Άϊ≥…ΈΣΦ”ΥΌΡ‘Υπ…Υ–όΗ¥ΒΡ≤Ώ¬‘[55]ΓΘ
3.6 ΤδΥϊ
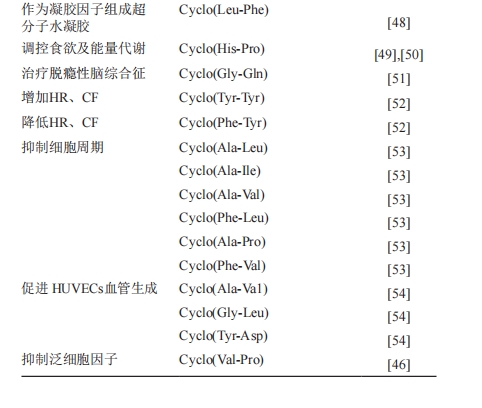
≥ΐΝΥ…œ ωΒΡ…ζΈοΜν–‘“‘ΆβΘ§“―”–―–ΨΩΜΙΖΔœ÷ΘΚcyclo(Leu-Phe)Ω…ΉςΈΣΡΐΫΚ“ρΉ”Ήι≥…≥§Ζ÷Ή”Υ°ΡΐΫΚ[57]ΘΜcyclo(His-Pro)ΡήΒςΩΊ ≥”ϊΦΑΡήΝΩ¥ζ–Μ[58]ΓΔΩ…“÷÷Τ»Υάύ–οΨΤ[59]ΘΜcyclo(Gly-Gln)Ε‘Ά―ώΪ–‘Ρ‘ΉέΚœ’ς”–÷ΈΝΤ“β“ε[60]ΘΜcyclo(Tyr-Tyr)Ε‘‘ωΦ”–Ρ¬ (HR)ΚΆΙΎ¬ω―ΣΝςΝΩ(CF)”–Αο÷ζΘΜcyclo(Phe-Tyr)Ω…“‘ΫΒΒΆHRΓΔCFΓΔΉσ–Ρ “ ’Υθ―Ι(LVSP) ΚΆ–Ρ‘ύ¥ΪΒΦΥΌ¬ [61]ΘΜcyclo(AlaLeu)ΓΔcyclo(Ala-Ile)ΓΔcyclo(Ala-Val)ΓΔcyclo(PheLeu)ΓΔcyclo(Ala-Pro)ΓΔcyclo(Phe-Val)ΨΏ”–œΗΑϊ÷ήΤΎ“÷÷ΤΜν–‘[62]ΘΜcyclo(Ala-Va1)ΓΔ cyclo(Gly-Leu)ΓΔcyclo(Tyr-Asp)Ω…“‘ΡήΦΪœ‘÷χΒΊ¥ΌΫχ HUVECs―ΣΙή…ζ≥…(P<0.01Μρ0.001)[63]ΘΜcyclo(Val-Pro)“÷÷ΤΖΚœΗΑϊ“ρΉ”Θ§”–Άϊ≥…ΈΣ–¬ΒΡΩΙ―Ή“©Β»[54]ΓΘ
4 ΉήΫαΚΆ’ΙΆϊ
Ήέ…œΥυ ωΘ§ΜΖΕΰκΡΒΡ…ζΈοΜν–‘≤ΜΫωœό”ΎΩΙΨζΓΔΩΙ÷ΉΝωΘ§ΜΙΑϋά®”ΑœλΩΙ―θΜ·ΓΔΫΒ―ΣΧ«ΓΔΦ”ΥΌΡ‘Υπ…Υ–όΗ¥Β»Ής”ΟΓΘœ÷‘ΎΒΡΩΤ―ßΦΦ θ‘Ϋά¥‘ΫΖΔ¥οΘ§Έ“Ο«ΡήΙΜΗϋ»ί“ΉΒΊΧα»Γ≥ωΜΖΕΰκΡάύΜ·ΚœΈοΘ§“≤ΡήΗϋ»ί“ΉΚœ≥…ΜΖΕΰκΡάύΜ·ΚœΈοΘ§ΜΖΕΰκΡ «ΜΖκΡάύΜ·ΚœΈοάοΉν–ΓΘ§ΉνΦρΒΞΒΡΜΖκΡΜ·ΚœΈοΘ§“≤ «Ήν”–ΧΊΒψΒΡΜ·ΚœΈοΓΘ±ΨΩΈΧβΉι‘ΎΕ‘20”ύ÷÷Μ·―ßΚœ≥…ΒΡΜΖΕΰκΡ―υΤΖΫχ––œΗΑϊΕΨ–‘ Β―ι ±Θ§ΖΔœ÷ΦΗ÷÷Ε‘NIH-3T3œΗΑϊ…ζ≥ΛΨΏ”–¥ΌΫχΉς”ΟΒΡΜν–‘ΫαΙΙΓΘ”…¥ΥΩ…ΦϊΘ§Ά®ΙΐΚœάμ…ηΦΤ”κΜ·―ßΚœ≥…”–ΆϊΩΣΖΔ≥ω–¬–ΆΙΠΡήΜΖΕΰκΡΓΘ

ΗυΨίΉν–¬ΒΡΈΡœΉΦΑ ΐΨίΉήΫαΝΥΕύ÷÷ΜΖΕΰκΡΒΡ…ζάμΜν–‘Θ§ΥϋΟ«Ω…“‘ΉςΈΣ–≈œΔΖ÷Ή”≤Έ”κœΗΑϊΦδ–≈œΔ¥ΪΒίΘ§“≤Ω…“‘ΩΙ÷ΉΝωΓΔ“÷ΨζΓΔΩΙ―θΜ·ΓΔΫΒ―ΣΧ«ΚΆΧαΗΏ–Ρ―ΣΙήœΒΆ≥ΡήΝΠΒ»Θ§»γ±μ1Υυ ΨΒ»ΓΘΉέ…œΥυ ωΘ§œΘΆϊΦ”«ΩΜΖΕΰκΡΒΡ…ζΈο―ßΜν–‘ΚΆ“©άμΦέ÷ΒΖΫΟφΒΡ―–ΨΩΘ§ΗϋΦ”≥δΖ÷ΒΊΝΥΫβΥϋΒΡΧΊ–‘Θ§ΖΔΨρΥϋ‘ΎΩΙΨζΓΔΩΙ÷ΉΝω“©Έο―–ΖΔΓΔ≈©“Β”Π”ΟΒ»ΖΫΟφΗϋ¥σΒΡ«±ΝΠΘ§¥¥‘λ≥ωΗϋ¥σΒΡ”Π”ΟΦέ÷ΒΓΘ
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷ΨΓΘ
