
ժ Ҫ�� ���ĸ߷��Ӳ�����һ���ɶ��ĺ�������ﹹ�ɵĸ߷��Ӳ��ϡ����ĸ߷��Ӳ��Ͼ�������Ȼ����/�ǵ������ƵĻ�ѧ��ɣ��ܹ���һ���̶���ģ����Ȼ����/�ǵ��Ľṹ���ܡ������ܽ������ĸ߷��Ӳ��ϵĺϳɷ�����������Ƽ���������ҽѧ�����Ӧ�ã��ص����������ĸ߷��Ӳ������������η��ӡ�����װ���ĺ����ľۺ��﷽��IJ�����ƣ��Լ����ĸ߷��Ӳ����ڿ��������������硢����֧�ܡ���֯�������������Ӧ�á�������ĸ߷��Ӳ��ϵķ�չ��ǰ��������չ����
�������ɸ��ְ�����ͨ���ļ�����һ��˳�����Ӷ��ɵģ�����������ڰ������뵰����֮�䡣���ڶ���Դ�����������ڣ���˶��IJ��Ͼ������õ����������ԺͿɽ�����[1, 2]��ͨ�����ض��ķ����а���������ࡢ������˳���ڰ����������θ��ֲ�ͬ�Ĺ����ţ�����ʵ�ֶ��ĵ�����װ����������װ���ϱȶ��ı������и��ߵ��ȶ��Ժ��õ����ơ���������ܣ��㷺Ӧ���ڸ�������ҽѧ����[3-5]���������鷢չ��һϵ�о�������װ���ܵĶ��IJ��ϣ��ڰ�֢��ϼ����ơ�ϸ����Ⱦ��ϼ����Ƶȷ�����й㷺��Ӧ��[6-8]��
�����Ǵ�������Ȼ���һ����Ҫ������ӣ��ṹ��Ϊ�������ӣ����д���Դ������ṹ���Ĺ���[9, 10]���ǶԵ����ʵ������ǵ����ʷ�������Ҫ����֮һ���Զ��ֹؼ�������̣������������۵������䡢ϸ��𤸽��ϸ��������ϸ���ֻ��ȣ�������Ҫ[11-13]�������ʵ��쳣�ǻ���������ϸ����ʶ���������֢�����������Լ������ڵ��������ؼ����йء�����ϵͳ��Ҫ����ϸ����������������������ϸ����ϸ���ڸ�Ⱦ�����Ǵ���ϸ�����������쳣�ǻ����ṹ�ӱ�����ϵͳ�ļ��Ӻ�ʶ��[14, 15]������Ҳ�����������ǻ���������װ�������İ�Ĥ�ǵ��ף��Ӷ������ڱ��������[16]�������������Լ������ʪ�ؽ���ϵͳ�Ժ���Ǵ��У�������G��IgG�����ͷ��������仯�������ʪ�ؽ����У�IgG�ۼ��İ����ǻ����ͱ���¶�ǽ��������������ʶ�𣬴Ӷ����ܲ��ʵ�������������ϵͳ[17]�����⣬�������ķ�չ�����У�ϸ����������ķֲ��������仯���ƺ���ת�Ƶ�״̬�йء�����ḻ������ѧ��������Ӧ��������ҽѧ�о������Ʒ�չ����������Խ��Խ��Ĺ�ע[18-21]��Ȼ������Ȼ����ĸ����Ժ������Ը��������ͱ��������˾����ѣ��Խ�һ���Ľṹ���Ρ������о���ҩ��Ӧ������˼������ơ�Ϊ�˳��������������ƣ�ͬʱ�����Ȼ���ϵ�ȱ�㣬�������Ʊ����ʵ;��ǺͶ��Ƿ�����������Ŭ��[22-24]��
���ĸ߷��Ӳ������ǻ�����ͨ�����ۼ�������������γɵ�һ����ӡ��������η�����һ�ֵ��͵�֧״���ĸ߷��Ӳ��ϣ�һ��Ϊ���׳߶ȣ�ͨ����3��������ɣ����η��ӵĺˡ���֧��Ԫ��ĩ�˻��š��������η��ӵĺ���λ�ڷ�������㣻���ž�ȷ��Ƶİ��������кͽṹ�ķ�֧��Ԫ�ں˺ͷ��ӱ���֮�䣻һ���ɰ�������ɵ�ĩ�˻���λ�ڷ��ӱ��棬�������β�ͬ�ǻ����Ŷ�������ͬ����ѧӦ�á�ĩ�˾��еĴ������ſ��Խ��ж�λ���ǻ������Σ����ǻ�������������з��Ӷ��ЧӦ������Ǽ����������ԡ���ѧ�����Ժ����õ����������Ե����ƶ��ܵ��˹㷺�Ĺ�ע������Ҫ���ǣ�ͨ������װ�ķ�ʽ�������Ծۼ���ʽ���ָ�������壬��ʵ�ֶ�������[25-28]����ˣ�����װ���ĸ߷��Ӳ���Ҳ��һ����Ҫ���о������ķ��Ӽ����ͨ���������������á���-�� �ѻ�����ˮ���õȷ��Ӽ�����������װ�γɲ�ͬ�ij����ӽṹ[29-31]���ǶԶ��ĵ����β��������˶����������﹦�ܣ�Ҳ��ı���ĵĽṹ������װ���ĸ߷��Ӳ����и����ǻ����ŵļ����ؿ������ķ���������װʱ��������ã�����������װ���ĸ߷��Ӳ��ϵĽṹ�ȶ��ԣ��ؿ��˶�������װ������ҽѧӦ�á�
�������������IJ��ϵĻ�ѧ�ϳɡ���Ʋ��Լ���������ҽѧ�����Ӧ�ã�ͼ1�����ص�������������ײ��ϵĹ������ԣ�������������ķ�չǰ�������ٵ���ս���������ۡ�
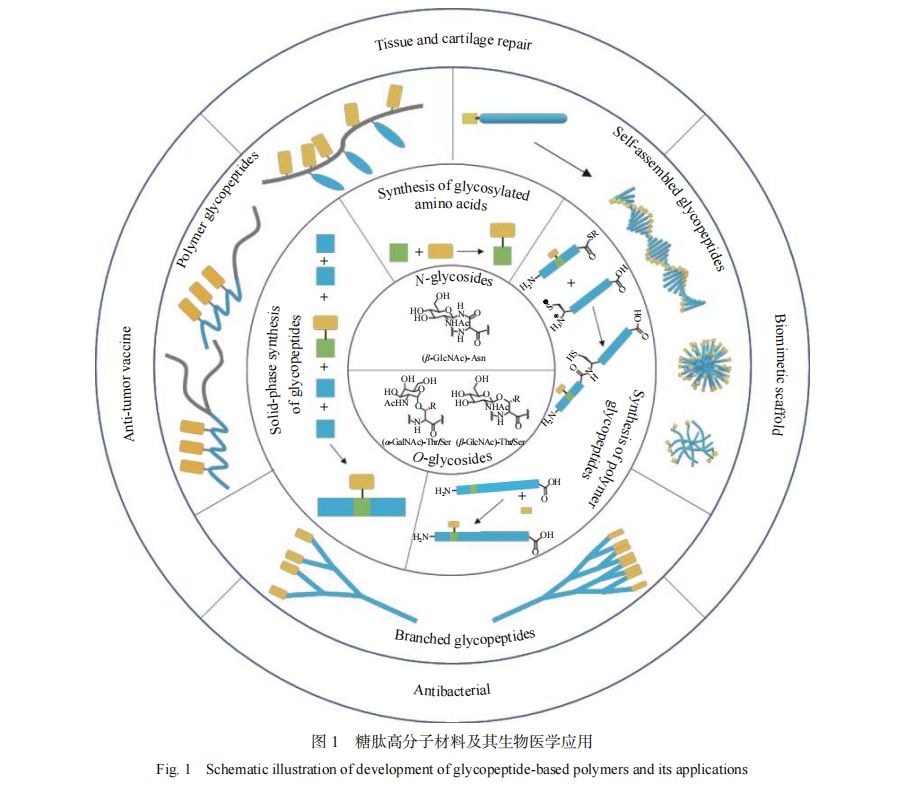
1 ���IJ��ϵĻ�ѧ�ϳ�
1.1 �ǻ���������ĺϳ�
����������Ȼ�γɵ����ռ����ɷ�Ϊ2���ࣺN-�����ࣨ���춬����������������O-�����ࣨ��˿���ᣨSer�����հ��ᣨThr�����Ұ��ᣨTyr���IJ���������Ҳ���ټ������������ᣨHyl�����Ǹ������������������Ŀǰ����۲쵽�Ķ���/�����ǻ���λ�����춬������Asn�����������������������ض�������Asn-X-Thr/Ser����ɵģ�����X�dz�������������κΰ����ᡣ��һ��λ�õ�Ser��Thr�л����ǻ����Ǿ��Ա�Ҫ�ġ�����Ȼ������O-�ǵ����У���ḻ��̼ˮ��������N-����-D-�����ǰ���N-Acetyl-D-galactosamine��GalNAc���л���˿������հ����ϵĦ�-O-���գ���ˣ������Ĺ��������ڽ����ʵ������Ħ�-GalNAc-Thr/Ser�����顣���������ձ���Ϊ��-O-N-�������������ǣ�N-Acetyl-D-Glucosamine��GlcNAc��-Ser/Thr�����ڵ����ʵ�˲ʱ�����о��е������ã��������Ҳ�Ԧ�-O-GlcNAc-Ser/Thr���ֵĺϳɸ����˺ܶ��ע�����⣬��-��¶�ǣ�Mannose��Man��-Thr/Ser����-�����ǣ�galactose��Gal��-Hyl�Լ�����O-���ջ�������Ҳ���������ǵĹ�ע��
1.2 ���ĵĺϳ�
���Ļ�ѧ�ϳɵ�һ���ؼ������ǽ��ǻ�����Ĺ���ż����Ϊ�˴ﵽ��һĿ�ģ����Կ������ַ�����һ���Ƕ��ʵ�������ȫ�����Ľ���ֱ���ǻ�������һ����ʹ��Ԥ���γɵ��ǻ��������ṹ�����ϳɶ���������ֱ���ǻ������ŵ��ǿ��Կ��ٻ�ò�ͬ���Ľṹ��Ȼ�����ڻ�ѧ�ǻ������õ������£������ǻ��ķ�Ӧ���Ե����ĵ��ܽ�ȵͣ�ֱ��O-�ǻ��������ܵ����ʵ͵����š����ǰ��ͺ����춬�������ֱ�����ϻ��N-�ǻ����ĵķ����������صĸ���Ӧ[32]��
Ŀǰ����õ�������ϳɷ����Dz���Ԥ���γɵ��ǻ���������ģ��������Ĺ���ϳɣ�Solid Phase Peptide Synthesis��SPPS����Ȼ�����ǻ����������ĺϳɴ����˸����ԺͲ�ȷ���ԣ�������SPPS�г��õķ�Ӧ�����������������������ĵ��Ʊ�[33, 34]����ˣ���ѡ������ʱ�����ر�С�ġ���O-���ռ�����ȶ��ģ�����ǿ���������ֻᷢ����-������ǿ��Ҳ��ʹ����������Ŀǰ��ʹ����������Ϊ�����ǻ��ı�����9-�̻��������ʻ���Fmoc����Ϊ�������-�����ı�����N,N'-�������̼���ǰ���DIC��/ 1-�ǻ���������HOBt����1H-��������-1-����������������������Σ�PyBOP��/HOBt������������-N,N,N',N'-�ļ������������Σ�HBTU��/HOBt��O-��7-���ӱ�������-1-����-N,N,N��,N��-�ļ���������������HATU��/ N-�ǻ�-7-���ӱ���������HOAt���µ���Ϊż�����ѳ�Ϊ�������ĺϳɵı�����[35-38]����Fmoc�ϳɷ����У����º͵ļ�������������ऺ��������¿�����ȥ��-�����ı�����������Ӱ�������е�O-���ռ�[39, 40]��Ȼ�������嶡���ʻ���Boc��/�л���benzyl�������У���-��������������ȥ����ǿ���������ᣨTFA����������ɵģ�����ǿ�����������������ռ��IJ��ȶ���O-��������O-���������������ӻ��ţ���TFA�ѽ���ĵĹ����п����ȶ����ռ����״��Ƶļ״���ˮ������Һ����ȥ��������[41]���ڱ������η�����Ҫ�Ŀ�������ʹ�����Dz�̫�ʺ��������ĵĺϳ�[42]��
1.3 �߷������ĵĺϳ�
�����SPPS�ϳɽ�����50�����������ڣ������Ȼ��ѧ���ӣ�Native Chemical Ligation��NCL���Ǻϳɸ�����������Ч����[43]��NCL��һ�ֻ�ѧѡ���Է�Ӧ��������pH�£�1��N�˰��װ���л���1��C����������ѡ���Է�Ӧ��ͼ2�����÷�����Kent������[44]��20����90�����չ�����ģ��˺ϸĽ��������ʵ���ԡ���NCL�ĵ�һ����C��������N�˰��װ���л����ϻ�֮�䷢�����淴ʽ���������Ӧ����Ϻ�ĵ����������Է��ط��������Ҳ�����ķ�����S��N�ƶ����Ӷ������Ӵ���������ѧ�ȶ�����Ȼ��������NCL�ض��ط�����N�˰��װ���л��ϣ������������ڲ����װ���л��ء������������ӷ������ǺͶ��ļ��ݣ���˿��������ǵ��ṹ�������������еļ������ԣ������������Ʊ����������Boc����[45, 46]��Fmoc�������ĺϳ��еĹ㷺Ӧ���Լ����ĵ��������Դٽ��˶��������Ʊ��·����ķ�չ[47]��
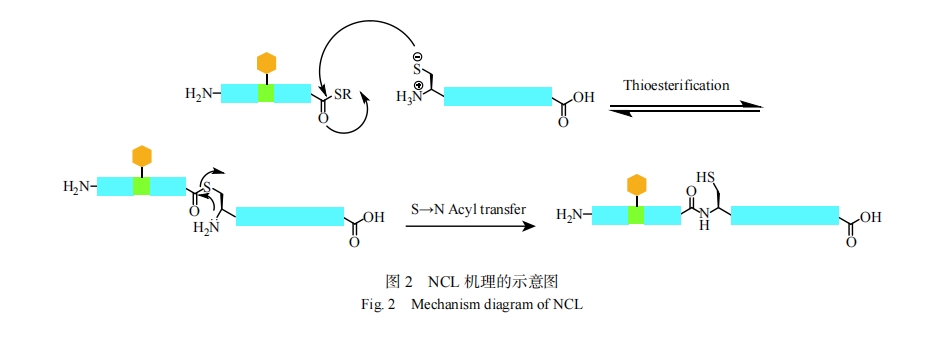
2 ���ĸ߷��Ӳ���
2.1 ���������
�����Ǻ͵�����֮��������ͨ��������������������������ܹ���ǿ��������ã����������Ϊ����ЧӦ�����ЧӦ���������������η������ڵ�֧״����ͨ��ż�����γɵ�֧״���Ŀ������ö��̬��ЧӦ��ǿ���뵰������á����η�����һ�ָ߷�֧�ۺ��������ȷ�Ҷ��ɢ�Եĺ�-�����ṹ����֧��ԪΧ�ƺ��ĵ�Ԫ����ϳɣ��㷺Ӧ���ڶ�������ҽѧӦ��[48-52]���������η��ӵĸ߶ȷ�֧��������ʺͷ��ӽṹ�����������η�����������Դ�Ⱦ���Ͱ�֢�����߷�Ӧ���ڼ������Լ��������η��ӵĿ���Ⱦ�Ϳ���ҩ��������ϡ�
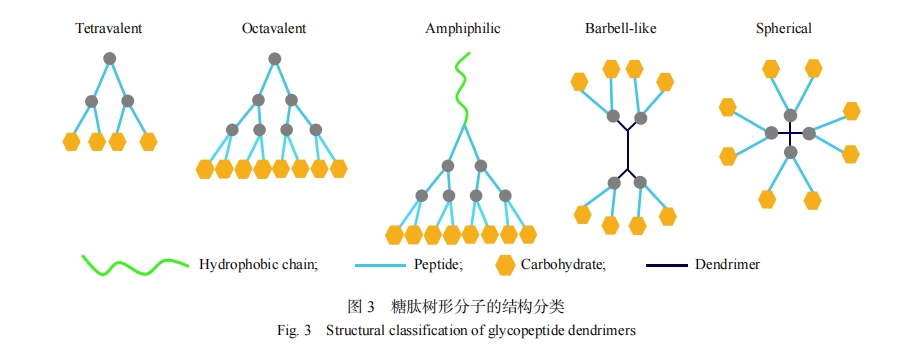
�������η��ӿ��Ը��ݾ���Ҫ����ж�����ƣ���ṹ����ʾ��ͼ3���������η��ӵĺ��IJ��ֿ����ǰ��������У�Ҳ�������ӳ�������ˮ��������γ����������η��ӡ����⣬����֧��Ҳ����ʹ���������η��ӣ��羭��ľ�����-���ͣ�Polyamindoamine��PAMAM�����η��ӡ��ظ��ķ�֧��Ԫͨ����һЩ�Ż��İ��������У�һ������������з�֧�����ŷ�֧�����ӣ������������ӡ�ĩ�˻��ŵ���Ŀ���Ŵ��������Ӷ�ָ�����ӣ��������η��ӵ����ܶ�Ҳ���ӣ��������η��ӵ����Ժ����й����ŵĿɼ��Խ��͡��ḻ��ĩ�˻��ſ������δ������ǻ��ţ���Щ�ǻ��ſ�������ͬ���ǣ�Ҳ�����Dz�ͬ���ǡ���һ���̶��ϣ�����������ʹ�ǻ��ŵ���Ŀ���ӣ���ǿ���Ǵ��뵰������ã��������ڹ����ſɼ�������������Ӷ����ͣ����η��ӵĴ��������������ӡ������������η��Ӿ���Ľṹ���ֿ��Է�Ϊ�����ԡ������κ����εȡ�
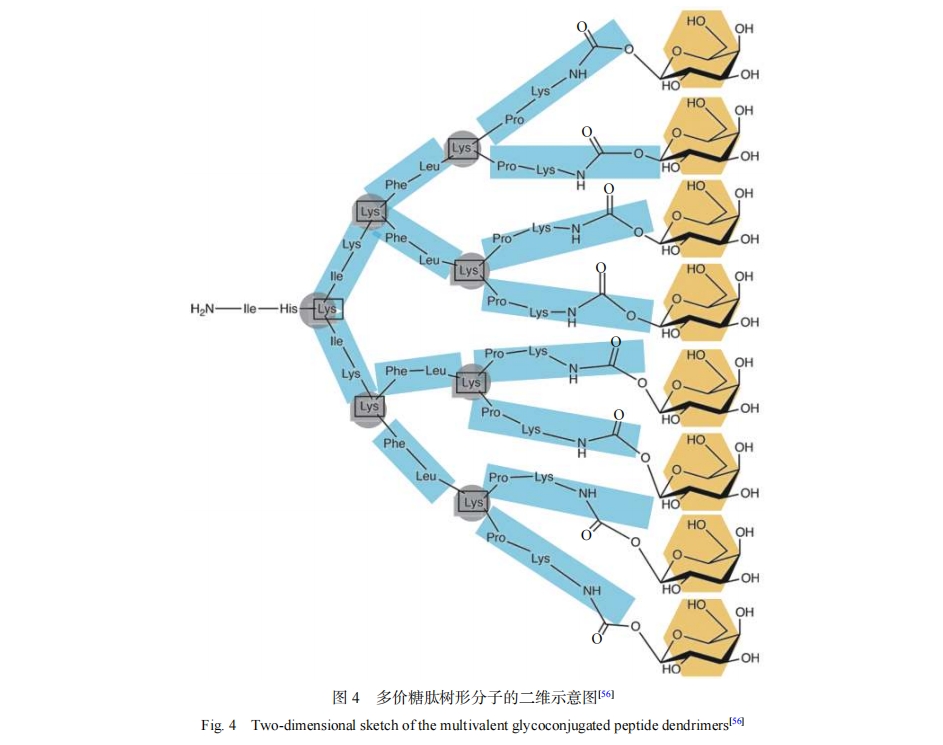
Reymond������[53-55]����˾���4��ĩ�˵�G2�������η��ӣ�ĩ�����Ӱ����ǻ������ǣ��ֱ����ڰ���ͭ�̼ٵ�������LecA��LecB�����ء�����4��ĩ�˵��������η���FD2��D-FD2��GalAG2��GalBG2����Ч���ͭ�̼ٵ���������Ĥ���γɲ��յ�����Ĥ������ķ�ɢ��2��ĩ�˵��������η��ӻ�ĩ�˵����ķ��Ӷ������صĽ����Ҫ���ö࣬��8��ĩ�˵��������η����������صĽ�������ӣ�����������16��ĩ��ʱ��ƽ����ÿ�������ǻ��Ľ��Ч���������͡�����������������LecA��LecBΪ�ľ���ṹ��ÿ��������1�����λ�㣬1��LecA��LecB��4�����λ�㣬����������1�����ε�4�����㣬����Ƶ��������η����ǻ�֮��ļ�����ľ����ڽ��λ��ľ���һ�»��ܹ��ŽӲ�ͬ���ľ���ʱ�������Ͽ�������ȵط��Ӷ�����ö�ʶ��ͽ�Ϲ��̵�Ӱ�죬��ȷ�����������ǽ�ϵ���Ѿ�����������������Ч�ʡ�����һ�������������η��Ӷ˻���Ŀʱ�����ܳ������ڿռ�λ����������������ؽ��λ������õ��ǻ���ʹ�������η����������صĽ������������������ӡ�������֧�����������ø����յ�2��3-�������������ʱ����������Լ���������������2��3-�����������2������֮�������������3��̼ԭ�ӵľ��롣һ����Ϊ���ϳ���̼����ʹ�������������Ҹ������϶̵�̼����ʹ�������Խ��͡�����ĩ�˻���ͬʱ���а����Ǻ������ǵ����Ǵ�ʱ���������ͬ���ĵ��Ǵ�������Bouvier������[56]��һ��Ӧ��MARTINI������ģ�ͺͳ�ʱ��ķ��Ӷ���ѧģ��ϵͳ̽�����������ж����ǹ��ܻ��˼������δ���Ӷ���ѧ�Լ�����������������������صĽ��������Ӱ�죬ͼ4Ϊ����������η��ӵĶ�άʾ��ͼ��
����ʹ������Ϊ�ǻ����η��ӵĹǼܣ�Ҳ����ʹ���������η�����Ϊ֧�ܣ��γ����Ρ������Ρ������Լ�������̬���������η��ӣ���PAMAM���η��ӡ����л�״��������ѡ���Կ�Ѱַ���ܻ�ģ�壨Regioselectively Addressable Functionalized Templates��RAFTs���Լ����ƻ�״�ṹ�����η���[57-59]��
2.2 ����װ���IJ���
���ڶ������ЧӦ���̶��ڶ�۹Ǽ��е��ǻ�ͨ�����ֳ��ȿ����Ե����ǻ��ߵö�ĵ����ʽ��������������ǻ��������ڵ�����������ռ�ݶ�����λ��ʱ�����������ЧӦ���ܴ�״����ЧӦ���������ѿ�������ͬ��֧������ƾ��ж���ǻ�����Ľṹ��Ȼ�������ڽ�Ϊ���ӵĶ�۽ṹ�ͺϳɳ�������Ҫ���һ�ּ��ķ������Ʊ�����ǻ��ء������������ų����ӻ�ѧ������װ���ĵķ�չ������װ����Ϊ����ǻ���������ṩ��һ�ּ���ҿ�����ķ���������װ���IJ����������ϳɡ����ӽṹ��ȷ�Լ����е����������Ժ����オ���Ե�������ƣ�������ģ����Ȼ���е�����װ���̣����γɾ����������ȵĸ��ӽṹ���ܲ��ϡ��ڶ��ķ��ӵİ������������ǻ����ţ��ܹ��������в�ͬ���ܵ������೬��������װ���ϣ�ͼ5����
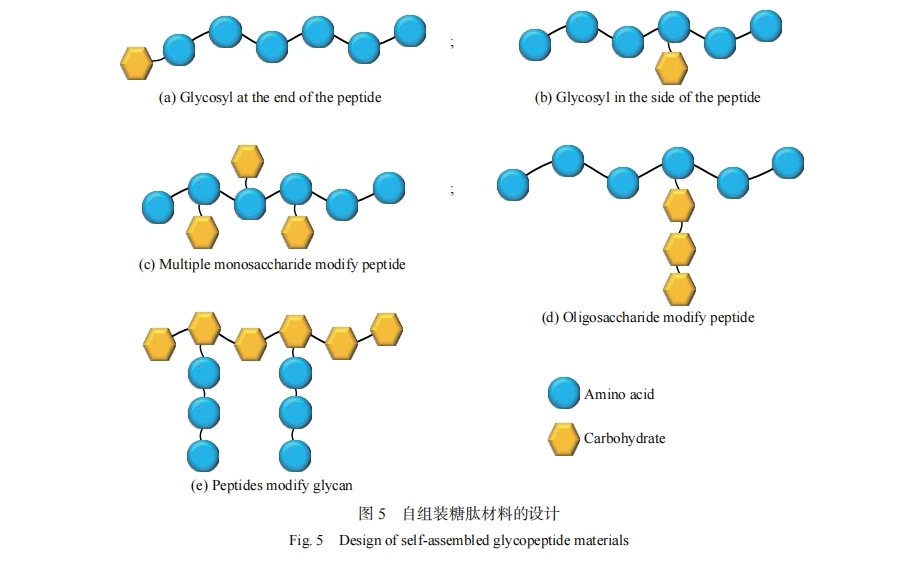
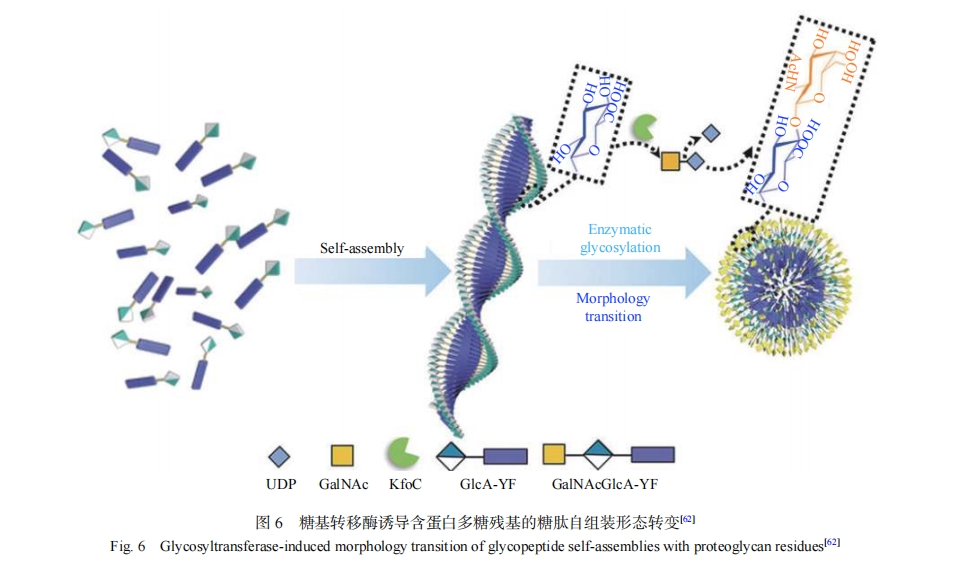
��������װ���ģ�һ�ּ���Ч�ķ�����������װ�������е�ĩ�˹��������ǻ��ţ�������һ�㲻��Ӱ�����ԭ�е�����װ���ܣ����η�����ҪΪ�����Ӧ����������[8]����˸�¶��ͨ�������Ӧ�����������������Ұ��ᣨTyr��-�Ӱ��ᣨVal��-�鰱�ᣨHis��-�춬���ᣨAsp��-���װ��ᣨCys�������ģ���¶�ǻ��������������ĵ�Nĩ�ˡ�Stupp������[26]����˵��ǻ�ͨ�������Ӧ����������ĩ�˽��й������ε����IJ��ϣ���������װ�γɳ��������ữ�������ṹ���ǻ�����Ӧ���Ľ�ϲ����ƻ����ṹ�����ڲ���-sheet�Ǽܡ�������ĩ�˻���������ǻ��ķ���һ�㲻��Ӱ�����ԭ�е�����װ���ܡ�Hudalla������[60, 61]ͨ����Q11��QQKFQFQFEQQ����N������N-�������������ǣ�N-Acetyl-D-Glucosamine��GlcNAc���õ�����GQ11��ͨ����Q11��GQ11�Բ�ͬ���ʵ���֮������װ�γɦ�-sheet������ά���ɹ��ı�������װ�������ǻ��ܶȡ��ǻ�������װ����汩¶������ʹ�ǻ��ܶȵĵ��ڱ�ü����Һ������ܵ��ⲿ�����и������صĵ��ڶ��ı��ǻ��ṹ��ʵ�ֶ�̬������������װ��ṹ��Ŀ�ġ��¹��̿�����[62]��Ƶ����ķ�����ˮ������װ�γ�������ά���ǻ���¶�ڱ��棬���ǻ�ת��øKfoC��PmHS2�������£�����ԭλ��Ч���γ����ռ��������µ�������������GalNAcGlcA-YF��GlcNAcGlcA-YF����װ��̬��������άת��Ϊ��������ͼ6����
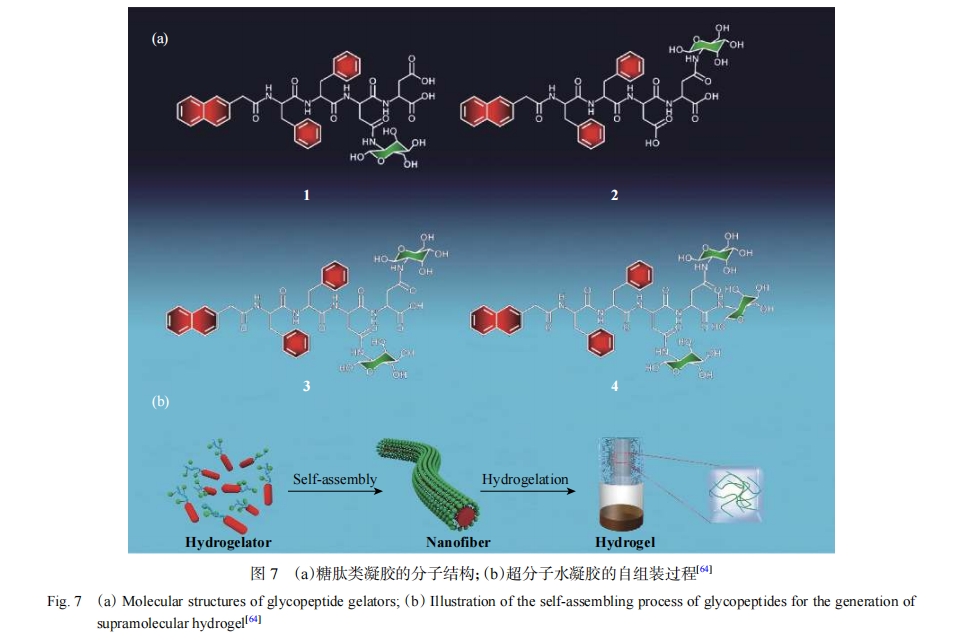
���⣬�ڶ������������뺬�в��������ŵİ�������Ϊ�ǻ�����λ�㣬���ǻ�ͨ����������Ĺ������ӡ�������������[63, 64]�����������-�������ᣨPhe��-Phe-Asp-Tyr���к�����-Phe-Phe-Ser-Tyr���в������ε��ǻ�������װ���ķ��ӣ�ͼ7�������dΰ������[65]��ƺϳ���ͨ��˿�������ż����¶�ǵ�����װ���ķ��ӣ���������װ�γ�������ģ�⸴�Ӷ��ǵĹ��ܡ��ڶ��IJ����Ͽ������ε����ǻ���Ҳ�����������������İ����ᣬ�Ӷ���һ��������ͬʱ���ζ���ǻ�������÷������[66]��Ƶ�������ؿ�ԭ𤵰��1��Tumor-associated Antigen Mucin 1��MUC1�����ķ���ͨ������2�����в����İ�������һ�������ϣ�ͬʱ����2��Tn��ԭ��������T����ϸ����λ�ĺͦ�-�۹Ȱ��ᣨ��-Polyglutamic acid����-PGA����ˮ�з����������װ���γ�����ż����������硣�ڶ����ϲ����������ε��ǣ����������νϳ��Ĺ���������Huang������[67]����������������Syndecan-1��˿���������������1����������
����������װ��Ϊ�������������ι����⣬�����Խ����dz�����Ϊ���壬�ڳ��������ν϶̵�����װ�ġ���������װ����������Ҫ��Դ������װ���ġ���ΰΰ������[68]��������ϸ�¶���dz���Ϊĸ�壬���������ζ���Q11����װ�����е����ķ��ӡ�����Q11����Ϊ��װ���������γɸ߶�����Ħ�-�۵�������ά��
����װ��������ż���γɵ�����װ�����ڻ���������������װ��Ϊ�Ļ����ϣ��־����ϳɵĻ�ѧ�ṹ�������γɵ���������װ�������ںϳɵ�ͬʱ��������װ�γɵĶ��ЧӦ�������γɵij�������װϵͳ�ڹ����������ͷ��ӽṹ����̬�ɵ�����װ��Ϊ��Ӧ�õIJ��Ϸ�����о�DZ����
2.3 ���ľۺ������
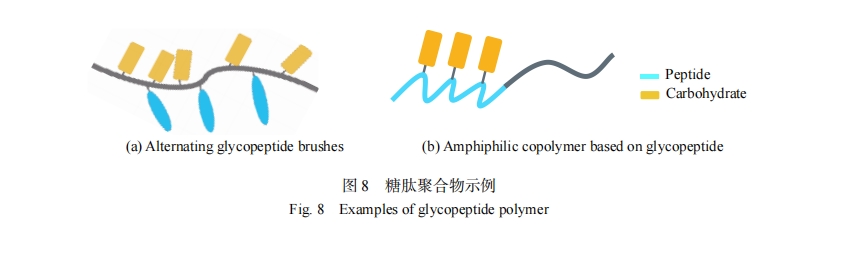
�ǻ������˸����Ļ�������������Ϲ����⣬������Ӱ����ṹ��Ϊ�˸��õ������ǻ���������װ�ĵ�Ӱ���Լ���Ȼ�������������װ���ƣ����õؿ����������Ĺ�������ҩ�ﴫ�ݹ���������ϡ����������֯���̵ȷ����Ӧ�ã��������͵ľۺ������ı�����������ͼ8ʾ����2�����ľۺ��
�ǻ���ͨ���������Ӽ����������ı䵰���ʵ���̬���ܣ��������������Խ鵼�����Ե�����ã�ͬʱ��ֹ�������Եľۼ���Hudalla��[69]�о��������ܵ��DZ�����Դٽ���������ά�ֲ���װ�ɸ����������硣��Ũ�ȵ�����������ά��ˮ����Ȼ��ɢ�������ǻ�����������ά�����ھۼ�����ӵ���������£�һЩ�ǻ�����������ά�����ϲ����С�������Ϊȡ�����ǻ��Ļ�ѧ���ʣ��������ǻ����������ڶ̳�������
���žۺ�������ǻ��ܶȵ����ӣ��ǻ����ǻ�֮����������¶�������Է���֮���������Լ��ۺ��������װ��Ϊ����Ӱ�졣����������[70]��ƵĽ��������Ƕ���ˢ��Alternating Amphiphilic Glycopolypeptide Brushes��AAGB�����и��ܶȵ��ǻ�������ͼ9�������ӳ�ʼ�γ���������������֮����ܶȵ��ǻ��������е��ǻ�-�ǻ�����û�ʹ��ʼ�γɵ����������������ںϡ�����������γ������ߣ�ʵ�ַּ�����װ��
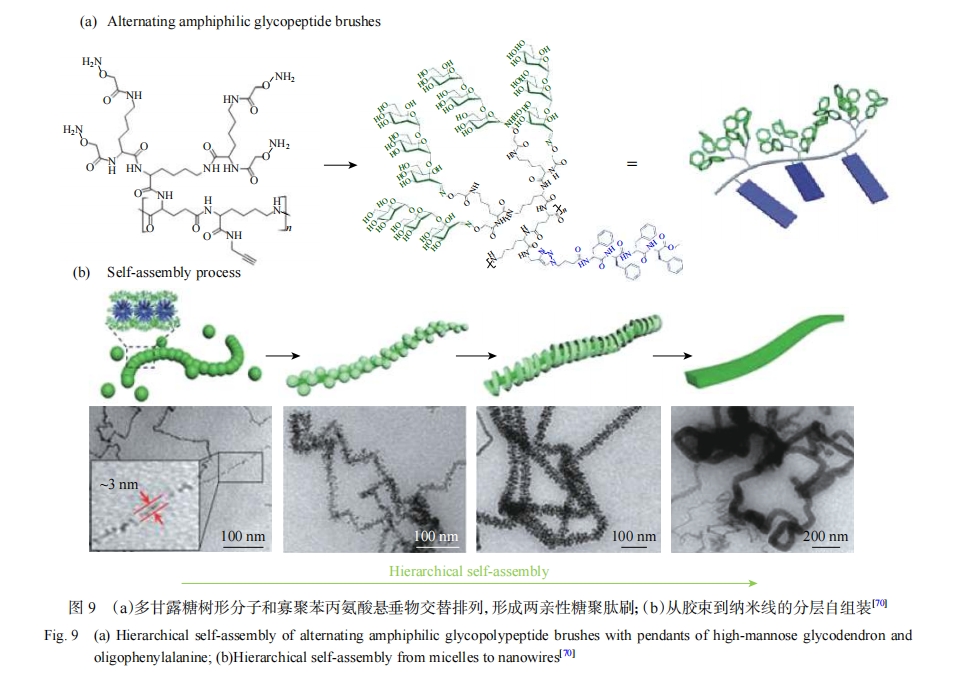
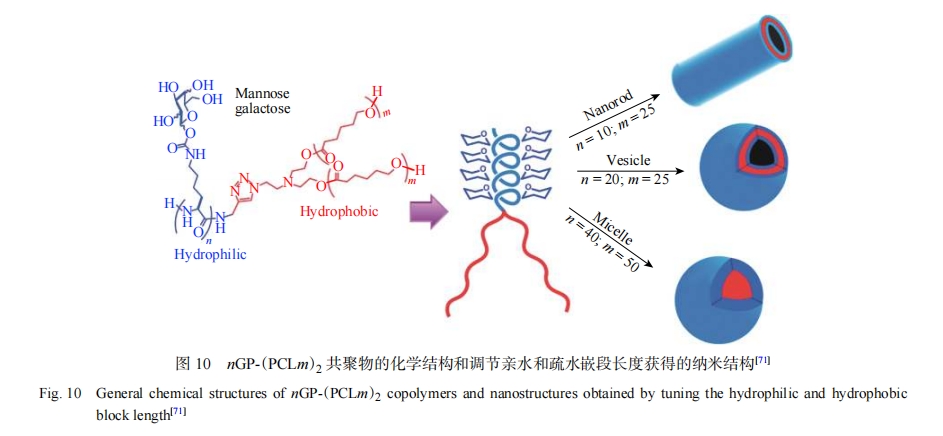
���⣬�ǻ�����Ҳ������Ϊ��ˮ���ֲ��빲�������װ��Gupta������[71, 72]��ƵĻ������ĵ������Թ�����������ǻ��ľ۸ʰ�������Ϊ��ˮ�Σ��ۣ���-������������ۻ������飨Polypropylene Oxide��PPO����Ϊ��ˮ�Σ��γ�������ά�����ݡ������Ȳ�ͬ��װ�壨ͼ10��������ʹ���Ƕ���Ƭ����Ϊ���ڹ�����ṹ�Լ���ˮ�Թ��ߵĹ������ۺ���Ҳ����������[73-76]��
3 ���ĸ߷��Ӳ��ϵ�����ҽѧӦ��
3.1 ����
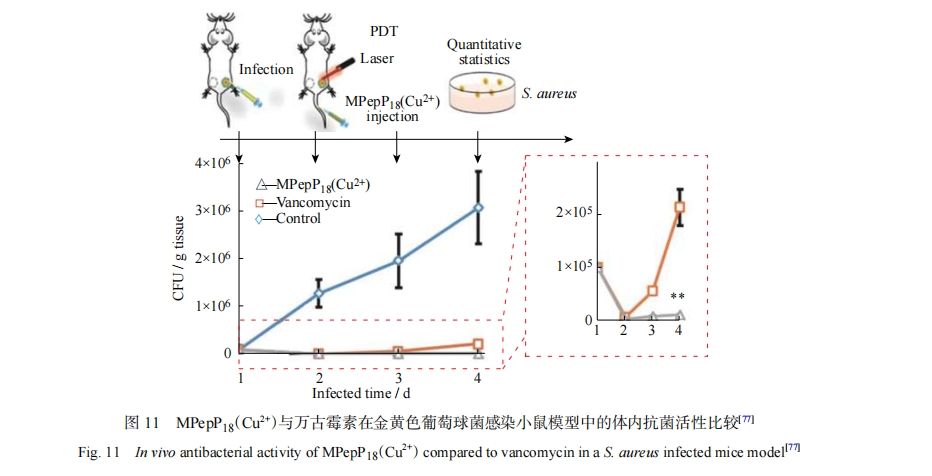
һЩϸ��������ɫ����������ܹ����벸�鶯��ϸ�������Ǿ���ϸ���������������棬�Ӷ���ֹ�����������ǵ����������ϸ�����溬�и�¶�����壬�ڵֿ���ԭ���Ⱦ��������Ҫ���ã�������һЩ���и�¶�ǵ��������ڰ������ϸ���ڵ�ϸ����Ⱦ�����Ľ������ϸ��������ϸ�������е���������������ʵ������װ���ڱ���Ⱦ����ϸ���ڳ����������Ӷ��ﵽ��Чɱ����Ŀ�ġ���������װ��������Ҳ���Եõ��Ŵ�������[77]��������и�¶�����ĵĹ�����Ӱ��������ΪMPC��MPCͬʱ����caspase-1ø��λ�㣬�����������ϸ����ø���γ�J�;ۼ��岢�����ھ���ϸ����ʹ�����ź���ǿ��������ϸ���ڽ��ɫ���������Ⱦ[8]������Ҳ���������С������Ρ�������ṹ�ĸ�¶������-Ҷ���ع�����ƣ�Photodynamic Therapy��PDT��ҩ�������ǿPDT��ЧӦ�ͻ������ԣ���������ϸ����ϸ���ڸ�Ⱦ��ͼ11����
ϸ����Ⱦ�����ÿ����ؽ������ƣ�Ȼ����ҩ�ԵIJ�����ʹ���й����߳��Ը�����������������ϸ�������ص����Ʒ�������Ϊ�ڽ���ϸ����Ⱦ�²�������ͬʱ����ֱ��Ӱ��ϸ���������������Ӷ�������ҩ�Գ��ֵĿ��ܣ�����ܵ��˹㷺��ע����������һ�ֵ����ʣ��������ǻ�������ʶ��ϣ���ϸ����Ⱦ�����Ĺ����з�����Ҫ���á�Ȼ������������-������þ��ж���ԣ��������װ���ı��㷺���ڰ��������صĿ������ơ����ĵ��ǻ�����ѡ�ð����ض�ϸ�������ص��ǣ����IJ����ṩ����װ�������������Ⱥ�Ĭ֢�������ڵ�������ά���γ��ǵ���������װ�Ľ�������ķ����еĶ��IJ��ֿ���ģ������װ�����γ����׳߶ȵ�����װ�塣����Щ��������У��ǻ�λ�ڶ���ĩ�ˣ�����װ�γɾۼ�����ǻ���¶�ھۼ�����档�ۼ����ɶ��������װ���ɣ���˾ۼ�������ж���ǻ����ţ������ṩ�������ã���ǿ����ϸ�������صĽ������������������ϸ���ĸ�Ⱦ�²�������������������[78]��Ʋ��ϳɵĸ�¶�����ε����ģ�����-Phe-Phe-Ser-Tyr��NMY�������۸�¶����������¶�ǽ�ϵ���Fim H𤸽�أ����IJ����ܹ��ڴ�ø�İ�������������������װ���Ը������Խ�ϴ˾���������ϸ��𤸽��Ĥ���ƻ�������ϸ��������
���˶�������װ�ṩ������⣬�������η��Ӿ��еĴ���ĩ�˻���ͬ�������ṩ����ԣ���ǿ�ǻ��������ص�����á�Reymond������[55]����˴��д��������Ǻ�/��������ĩ�˵��������η��ӣ�����ͭ�̼ٵ�������������LecA��LecB��������Ч����ͭ�̼ٵ�����������Ĥ���������η���ĩ�˻������������ӣ�ĩ���ǻ������ԺͿɼ�����С����Ч�Բ����������������������˺ϳ��Ѷȣ���˴�Ӧ�õ��������η���ĩ������һ����2��16���������ľۺ��﷽�棬Chan-Park������[79]����˺��о۸�¶�ǺͿ������ĵ��ı����;ۺ���������ı۶�ϸ������ɱ�����ã����۸�¶�DZ۶Դ˾�FimH�����ر��������������ǿ�˾ۺ����ϸ������������������������¶���մ��ڵ�����£��ۺ���Ҳ�ֿܵ�������
3.2 ��������
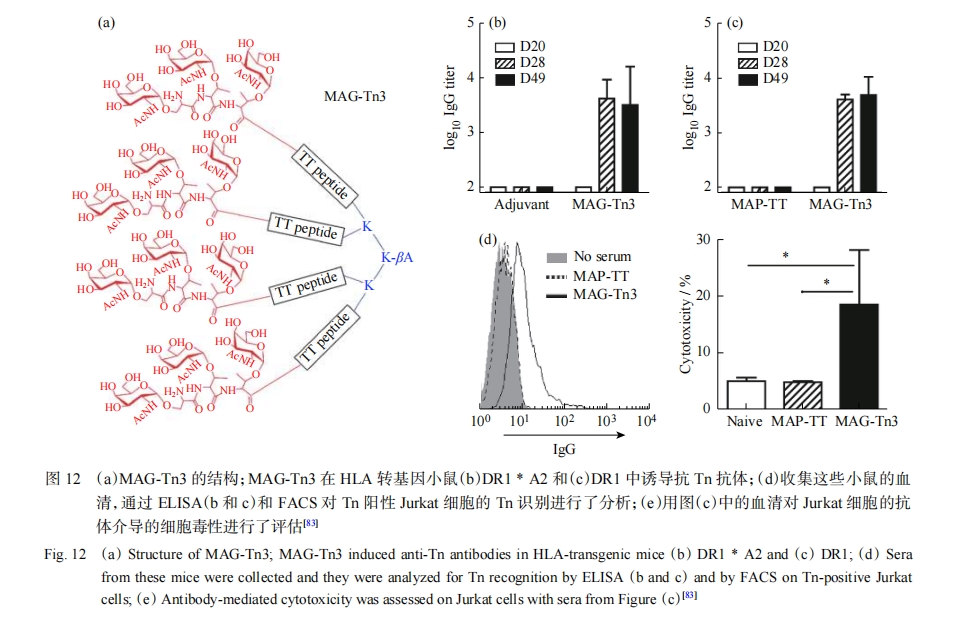
����ϸ���ϵ��ǵ��ף��ر���𤵰�ף����г�����������Ƥ����ϸ���ϵ�𤵰�����ڹؼ��ǻ�ת��ø�ı仯����𤵰�����������ؽض̣���Ϊ�������̼ˮ�����ԭ��tumor-associated carbohydrate antigens��TACA��������T��ԭ��Tn��ԭ������Һ�ữ�������[80]���Ѿ����ּ���𤵰��������ϸ���й��������������������о�������������ؿ�ԭ𤵰��1��MUC1����һ���dz���ǰ�����о��е㡣MUC1��һ��Ĥ���ף��������������о��������ڴ�Լ80%����Ƥ��ϸ������MUC1����������ǵ������ǵ������������ԣ�������ϸ���з��������MUC1ͨ����1����������Я������ϸ��������ϸ��2�ֿ�ԭ����������ԭ�����������յ���Խ�����֯�����߷�Ӧ����ˣ�ͨ����ѧ�����˹��ϳ�MUC1��������һ����Ч������������ķ�����Ŀǰ��Ҫ��3�����MUC1����������IJ��ԣ��ֱ��ǽ�MUC1������Tϸ����λż������MUC1���������嵰��ż������ƺ���������MUC1���ġ�������Ѿ��Ʊ���������MUC1����������ı�λΪ�б�ĺϳ��������磬ʹ�ö��ĵ�һ���干��ż�����ֿ�ԭ��Ȼ����������װ��������������װ���������η��Ӵ���ĩ�˻����γ����մ�ЧӦ�������ߡ�Dumy������[59]��������ѡ��Ĺ��ܻ�ģ�壨RAFTs����Ϊ���֧����ƿ������磬��Щż������ʾ�ɴص�Tn��ԭ���������������̼ˮ�����ԭ��Tumor-associated Carbohydrate Antigens��TACA�������Ժ�ѡ�����B��T��ԭ������ԭ�Խ���������������о���Kunz������[81]��ƺϳ���MUC1�����ظ�����ż��Tn����Һ��Tn��ԭ�Լ����˷���أ�TTox������Ұ���ͺ���MUC1ת����С�����յ���ǿ�ҵ�����Ӧ������÷������[82]���㶨��Ȼɱ��Tϸ����invariant natural killer T cell��iNKTϸ������ǿЧ��������-����������������-Galactosylceramide����-GalCer����MUC1���Ŀ�ԭ����һ�𣬲�������װ����Ϊ���͵���������֢���磬��û���ⲿ�������������������˸�ˮƽ������������IgG���塣Bay������[83]��ƺϳ���һ���������η��ӣ����ڱ���HLA-DR1��ת����С�����յ�Tn�����Կ���鵼����Tn��������ϸ����ɱ�ˣ�ͼ12����
3.3 ����֧��
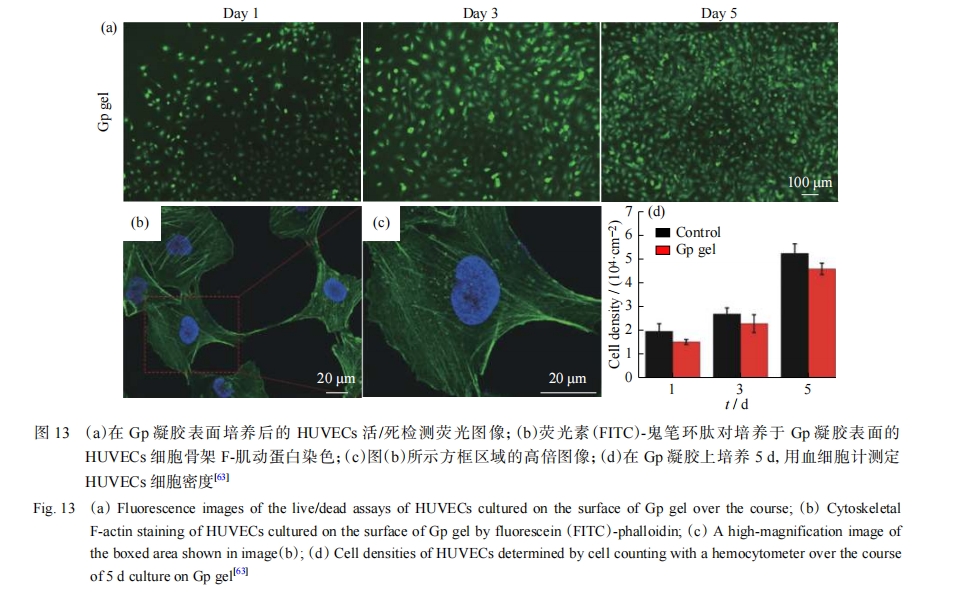
��������Դ����������Ǻ�����ɣ��������������Ժ������ԡ���ѡ���ض����ǻ��Ϳ�������װ�γ������Ķ��ĺ���������װ�γɵ���װ�廹����ģ��ϸ������ʣ�ECM���ǻ��������������Ϊ����ķ����������������ǵ�ע�⡣������������[63]��ƵĻ������ĵ�����װˮ�������沿��������ͨ����-��������ø���ϸ�����ź����꾲����Ƥϸ����������ͼ13������ΰΰ������[68]��ƵĦ�-sheet Q11�Ľ�֦���ϸ�¶��������װˮ������GPgel�������κ��������Ƽ��������Ҽ����˿����ϣ�Ҳ���Դٽ�����Ƥ����֯������Ѫ�ܡ�
3.4 ��֯��������
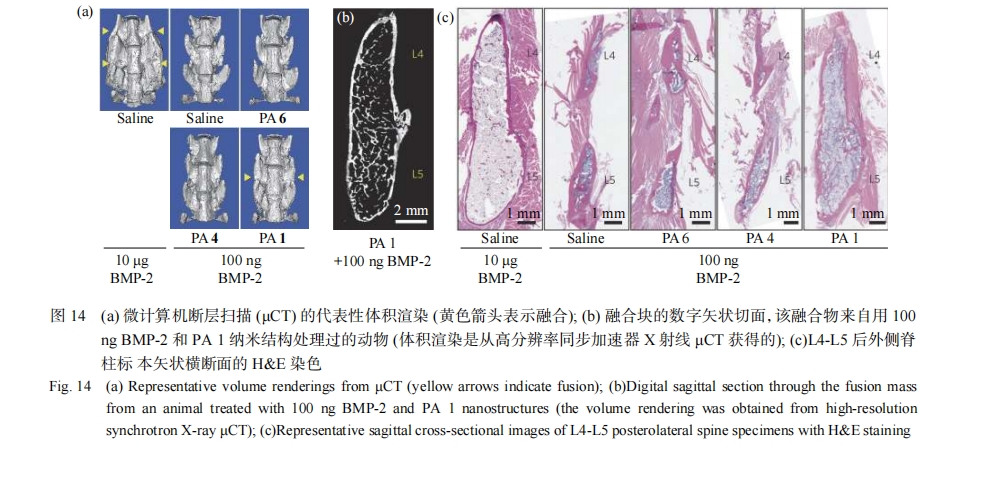
�ǰ����ǣ�glycosaminoglycans��GAG�����������������أ�Heparan Sulfate��HS�������������أ�Chondroitin Sulfate��CSs���������ᣨHyaluronic Acid��HA���ȣ����ڲ��鶯����֯���ձ���ڵ����ʶ��ǣ������������й㷺Ӧ�á���Ȼ�ǰ����Ǿ��и����Ժ������ԣ������������������ҽѧ������о���Ӧ�á�����װ���ĵ���ϳɽ�Ϊ���ҽṹ��ȷ�ɿأ�����װ���γɵ�������ά����ģ����Ȼ�ǰ�������̬����������������װ���ĵľ��ʵ;��ǹǼܱ�����������Stupp��������Ƶ����ữ���ṹ��������������װ�γ����ṹ˿״���ı���Ȼ���ữ���Ǹ���������ǿ�˹���̬��������2���źŴ��������Լ��ͼ����ٽ��˼����й�����������ͼ14����Guler������[84]��Ƶ�ģ����Ȼ��������ӵ�����������ά�����ʸ�ϸ������ã�������������ȻѪ���صķ�ʽ�յ����Ƿֻ���
4 �ܽ���ǰ��չ��
���������ĵĺϳɵõ��˹㷺��չ�����н��ǻ���ǰ���ӵ��������Ϲ����ǻ��������ᣬ�����ù���ϳ����������������ģ�黯ƴװ��Ϊһ���ܻ�ӭ�ĺϳɷ�ʽ��Ȼ�����ǻ���������ĺϳɺ�ʱ�ϳ�����Ϊ��ͨ����Ҫ�ಽ��Һ������Ҵ���������鲻�ǿ��̹��ģ���ʹ��������������̹������Ǽ۸�ͨ�����䰺����������Щ�ṹ��Ԫ�ڹ������ĺϳ��е�Ӧ�ã����⣬���ǻ����������������ĺϳ��ڼ䣬��ż����Ӧ��Ч�ʴ�ͣ����������ڴ��и߾��ǵİ������λ�����¡������IJ��ϵ���Ʒ��棬��Ȼ��Ҫ�����ء���̬�ص������ĵĽṹ����װ���Խ�һ��̽���ǻ��뵰��֮�������ü�����ù����еĶ�̬�仯����θ���Ч�غϳ�ģ�黯���ǻ��������ᣬ������ڹ���ϳ��е�ż��Ч�ʣ������غϳɽṹ�����������IJ��ϣ���Ȼ�д���һ��̽��������������ķ���֮�䡢�����뵰��֮�������ü��������װ���ܴ�����Ӱ�죻�������ڸ��ֻ������ض����Ľṹ�ȶ��Ե�Ӱ���Լ������ڴ˹����еĶ�̬�仯����Ȼ��һ����ս�����⣬��Ϊ�������ĵĽṹ�빦�ܣ���ƺͺϳɸ���ģ����Ȼ����/�ǵ��ķ��ӣ�������̽���ǻ��������ڸ��ӵ��������������ã��ۺ���������/�ǵ����ǻ���������Ҳ��һ���dz����������ķ���
��������������Ϊ��ҵ����ѧϰ����Ȩ��ԭ����ԭ��־���У�������Ȩ������ϵɾ�������±�ע���������³����������Ķ�ԭ�ļ��ο����ף����Ķ�ԭ��־
