
ժҪ��ϸ������Ĥ���µ�ϸ����ҩ�������ܵ��˹㷺��ע��������Ĥ����һ��������ƺ�ɱ��ϸ������Ĥ�������ƵĿ�������,������Ϊ����Ŀ�ϸ������Ĥ�����Ϳ���ҩ��Ϳ�����Ĥ��������Ĥ����ּ������á�������Ĥ�Ķ�����Ĥ�γɵĸ�Ԥ���ü�����ء�������Ĥ��Ŀǰ���ڵ����⼰����˼·�Լ�������Ĥ��δ����Ӧ�������չ��������
ϸ������Ĥ��ϸ��𤸽��������߷�������沢�����ڵĵ����ʡ����ǡ�eDNA��Extracellular DNA����ˮ�ϻ��ʣ�������Ǹ�����,Extracellular polymeric substances,EPS������������ۼ���[1]�����������Ĥ����һ��ϸ������,����Ĥϸ�����������ڳ����������,��EPS���dz��еĻ�����ʩ,Ϊ����Ĥϸ���ṩ��Ҫ�ı������о�����,����Ĥϸ�����Բ����ȸ���ϸ���ߴ�10-1 000���Ŀ�������ҩ[2]��������������ḻ�����ࡢ���������Ժ�ǿ������ߵ����ص��ܵ�ѧ�ߵĹ㷺��ע��2008��,Hancock��[3]�״α�������Դ��������LL-37��������ϸ������Ĥ�γɵ���������,һЩ���п�����Ĥ���ԵĿ������ı��о�����̷���,Ҳ����һ���ֿ������ij�֮Ϊ������Ĥ�ģ�Anti-biofilm peptides�������ľͿ�����Ĥ�ĵ�����о���չ����������,�����������˲���Ŀǰ�ѱ����Ŀ�����Ĥ�ģ���1����
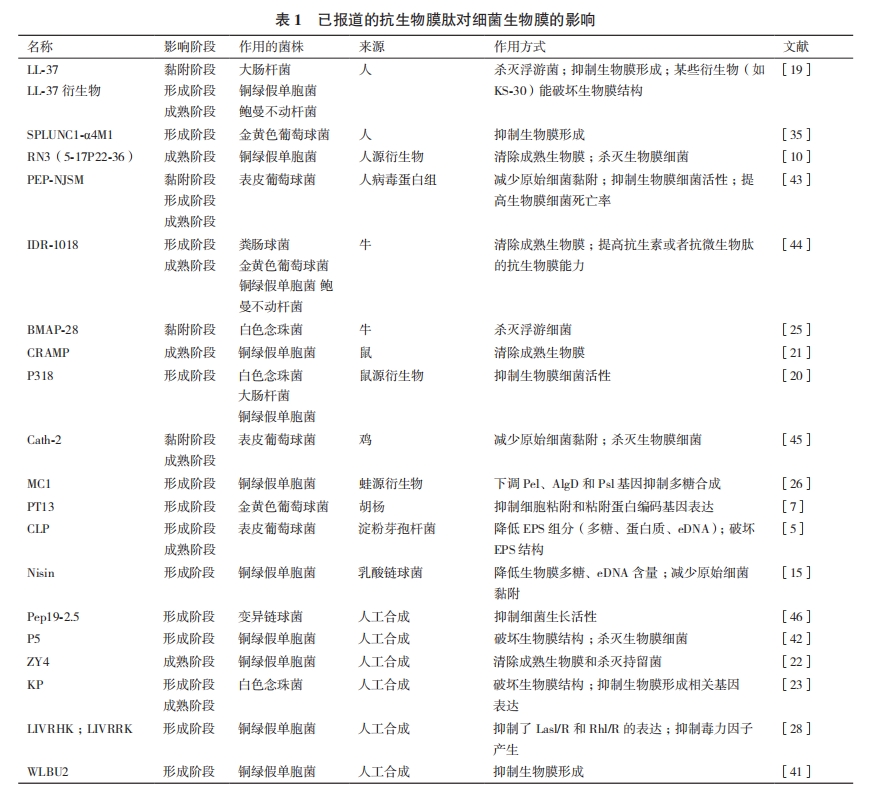
1 ������Ĥ��������Ĥ����ּ�������
1.1 ������Ĥ���뵰���������
����Ĥ�İ�������а�������������,������ë��IV�;�ë�Լ�������ϸ�����ڵ�ø���絰��ø���ȡ���ͭ�̼ٵ�����Ϊ��,���ë��IV�;�ë���ж�������ѧ����,����ͨ�����ֶ����о�֤�����Ա�ëΪ�п�ԭ�ܹ�Ԥ��ͭ�̼ٵ��������µĸ�Ⱦ������֮��,IV�;�ë����������Ĥ�������ɷ�����ʹ������Ĥ�����ȶ�[4]��Gowrishankar��[5]���ֻ�����CLP�ܹ�ʹ��Ƥ�������������Ĥ���ǡ������ʡ�eDNA��������,����������в���ȷ��Duperthuy ��[6]���ֻ��һ�������ǿOMVs���ױ�������ǿ��������е�Bap1���ס�Bap1����������LL-37��OMVs�����OmpT����,ʹ������Ĥ�������LL-37Ũ�Ƚ��͵���ЧŨ������,ʹ������Ĥϸ������LL-37�����á�����ϸ����ø��������Ĥ����ɳɷ�,Ҳ��ϸ���Ķ���������ɳɷ�֮һ��Akeel��[7]���ָ���������Ŀ�����Ĥ��PT13�ܹ����ٽ��ɫ��������������ӵIJ���������������ɱ������Ĥϸ�����õĿ�����Ĥ��Ҳ��ͨ��ɱ��ϸ���ķ�ʽ���ٶ������ӵIJ�����
1.2 ������Ĥ���������������
����Ĥ�еĶ��ǽṹ�����ǵ��ǻ����Ǵ���Ӿۺ���,��Ҫ����ά������Ĥ�ĹǼܽṹ�����ǻ�����ͨ��������ѧ���������ش�������������ϸ�����������á����ƿ������Ļ��ԡ��Լ����������,������ϸ�����Ͽ��Ļ���������[8]��
��ͬ���²��������İ������Ҳ�в��졣��������²������������Ӱ������,��ͭ�̼ٵ�����������ǣ�Alg��,�����²���Ҳ����������Ӱ������,���Ƥ���������ϸ�������𤸽�أ�PIA��������,ͬ��ϸ�������İ������Ҳ�����в��졣����,�����ײ���KpTs101�������������,��KpTs113���������Ӱ������[9]��һЩ������Ĥ���ܹ�ֱ������������Ĥ�ڻ��°�����Ƕ��ѡ���Pulido��[10]�ϳ������RNase3����������,��ͨ���о�֤ʵ�˻���λ��ȱ��ͻ����������ױ�������ͬ�Ŀ�����Ĥ����,���Ҿ��б��ױ���������Խ��EPS���ԡ�����,�ƻ�������ǽṹҲ�ǿ�����Ĥ�����õ���һ����,��EPS�뻷����CLP����ú���,CLP��ʹ��Ƥ�������EPS��ˮ,��ʹ�ǻ���������ֹ�����ռ�����[5]��ֵ��ע�����,��������뿹����Ĥ�ļ�ľ�������ûᵼ������Ĥ��ϸ�����ڿ�����Ĥ�ĵĵֿ�����ǿ���������ӿ�������LL-37��hBD3��PIA�ľ����ų������ܹ���������ϸ���Ӵ�[11]����KpTs113������ɵİ�������ܴ�ʹ�����ӿ�����Ĥ��BMAP-27������������,����KpTs101��������DZ��ֳ���ǿ���յ�������Ĥ��������������[9]��
1.3 ������Ĥ����eDNA�����
ϸ����DNA��eDNA����ϸ�������ͷš����ڻ�����ϸ���ܽ���ͷ��ڰ�������е�DNA,��ϸ���������ʮ����Ҫ�����á���1����������Ĥ����ά������Ĥϸ��𤸽���ۻ�������Ĥ�ȶ��Ե���Ҫ��ɳɷ�[12];��2��Ӫ�����ã�������Ϊ�˾�Ψһ���ס�����̼��Ӫ����Դ[13];��3���������ã��������Ͻ������ӣ���Mg2+���������أ��簱��������ҩ�[14];��4������pH;eDNA������Ĥ�е��ۻ���ʹ����Ĥ�ֲ������ữ,����pHֵ��ٽ�ͭ�̼ٵ���������ҩ���Ͳ���[12]��һЩ���������ܹ���������Ĥ��eDNA�����½�����ֱ����֮���,ʹ��eDNA�������书�ܡ�Wang��[15]��������������ģ�nisin���ڸ�Ũ���������ܹ���������Ĥ��eDNA�ĺ���,�ҳ�Ũ�������Ա仯��Jones��[14]������Դ�������ģ�hBD-3���ܹ�ֱ����eDNA���,��ͬʱ�ƻ��˿�������hBD-3��������ԡ����ܺ��ӵ���,Lewenza��[16]������eDNA��Ͻ��������ӣ���Mg2+����,�ܹ�����PhoPQ��PmrAB˫���ϵͳ������ϸ������ṹ�����仯������ɱ��ڸǣ����ͽṹ�ػ�ϸ����,ʹ�������ӿ�����Ĥ���������������ϸ��Ĥ��
1.4 ������Ĥ��������Ĥϸ�������
����Ĥ��һ�����С�Ģ��������Ʈ��������3D�ṹʵ��,���������Ĥϸ��Ҳ��˾��ְ[4]�������Ǹ�������Ĥ���ۻ������Ϸ���EPS������Ĥ������,���븡��̬ϸ��������������һ��������ǰ������������ϸ��������ͱ仯�ı��ͽṹ�ػ�ϸ��,�ܹ������߶ȵĿ�������ҩ�������һЩ�ײ�����Ĥϸ��,�����䲻�ϵ��ܵ���Ӫ������������pH�ļ��˻�����ս[17],���һЩϸ��Ϊ�������ɴ�ת����һ�֡��ʹ�л�״̬�������ǡ�����״̬����ʹ���һЩ�����ڻ�����λ�㡢mRNA��������ȴ�л���ؿ����ظ߶���ҩ,���Ҹ�����Ĥϸ�����ڲ����������Ŵ��仯������²����Կ����ص���ҩ[18]��
��������Ĥ�����о���������,һЩ�ܹ���Բ�ͬ������Ĥϸ���Ŀ�����Ĥ����̱����֡�LL-37���о���㷺����Դ������Ĥ��,LL-37��������Ƭ�Ρ������ﶼ��֤ʵ���п�����Ĥ���ԣ�������������Ĥ�γɺ��ƻ���������Ĥ������ɱ������Ĥϸ��[19]��De Brucker��[20]��������Դ������Ĥ��CRAMP�����־�Ũ����,�ܹ����ư�ɫ���������Ĥ���γɡ�Chen��[21]���ּ�Դ������Ĥ��Cath-2�ܹ���ͭ�̼ٵ�������������Ĥ��ɱ������Ĥϸ��,��CRAMP����������Ĥ�����Ե�������á�Mwangi��[22]���ֿ�����Ĥ��ZY4�ܹ�ɱ���������˾�����Ĥ�еij�������Paulone��[23]������һ�ֺϳɿ�����Ĥ��KP,�ܹ�ͨ����������Ĥϸ������Ĥ�γ���ػ���ı������ƻ���ɫ�������������Ĥ��
2 ������Ĥ�Ķ�����Ĥ�γɵĸ�Ԥ���ü������
����Ĥ���γ���һ����������̬�Ĺ���,��Ҫ��Ϊ𤸽�Ρ��γɽΡ�����κ��ͷŽ�[24]�����γɵľ�����̳����ܵ���������뻷���Ĺ�ͬ���á�
2.1 ����ϸ��𤸽
�²����γ�����Ĥ��һ������Ҫ���ھ���ϸ������𤸽��һ��������߷�������档��ϸ��δ𤸽ʱ����𤸽���ι�ʱ����Ĥ�γ�ʱ������������ܵ�Ӱ�졣���,���ÿ������ĸı�ϸ����𤸽������𤸽�IJ��ϱ�����ܹ���������Ĥ���γɡ�Scarsini��[25]���ÿ�����Ĥ��LL-37������������Ծ۱���ϩ��ͪ�����𤸽�Ӷ���ֹ�����������Ĥ���γɡ�����һЩ������Ĥ��Ϳ���о����ڱ���Ӧ�ü�չ����������
2.2 ��Ԥ����Ĥ��ֵ��γ�
ͨ��ֱ�ӵ�������Ĥ��ֻ���ı���Ҳ��Ӱ������Ĥ���γɹ��̡�Yu��[26]���ֿ�����Ĥ��MC1����ͨ���µ�ͭ�̼ٵ�����pelA��AlgD��PslA�������Ա��������ٰ�����Ǻϳ�����������Ĥ���γɡ�Akeel��[7]���ָ���������Ŀ�����Ĥ��PT13�ܹ�����ϸ��𤸽��ϸ��𤸽���ױ��������EPS�IJ���,��������Ĥ���γɡ�
2.3 �������ܶȸ�Ӧϵͳ
�ܶȸ�Ӧ��Quorum sensing,QS��ϵͳ�����ڶ���ϸ��֮��,������ϸ�����ֶ������Ӽ�����Ĥ���γ���ػ���[27]��ͭ�̼ٵ�������QSϵͳ��Ҫ��2���˿���ᣨAHL������LasI/R��RhlI/Rϵͳ��2���AHL������PQS��IQSϵͳ���,������Ĥ��صĶ�Ϊǰ��[27]��Taha��[28]�ϳ���2�ֿ�����Ĥ��,�����ʾ,���ܼ���ͭ�̼ٵ�����PAO1����Ĥ���γɺͶ������Ӳ���,��ͨ��ӫ�ⶨ��PCR�ⶨQS��ػ���仯,������lasI��lasR��rhlI��rhlR�Ļ������ˮƽ�������͡�Ciulla��[29]�����˽��ɫ�������������ĤQSϵͳ����RNAIII�����ģ�RAP������е��ף�TRAP���鵼��,RNAIII�����ģ�RIP���ܹ���RAP�����Ӷ�����TRAP�����ữ,������ϸ��𤸽������Ĥ�γɡ����������Ĥ�IJ�������ɱ������,��˿���Ϊҩ�����Ч����������ֱ������������Ĥ���γ�,���Իָ������غ�����ϵͳ������Ĥϸ����ɱ�˻��ԡ�
2.4 ������c-di-GMP
3,5-���������յ����ᣨc-di-GMP���ǹ㷺������ϸ���ڵĵڶ���ʹ,���������ѧ�������,ͬʱҲ������Ĥ���γ�ֱ����ء�c-di-GMP�������ӿ��Դٽ�ϸ������𤸽���ۼ���EPS�ķ������յ�����Ĥ�γ�,������������Ĥ��ɢ,��˿���ͨ������ϸ���ڵ�c-di-GMPˮƽ���ٽ��Ѿ��γɵ�����Ĥ�ֽ���ɢ[30]����һ����Ļ��ƿ���������ǵ��γɹ������,Whitney��[31]֤ʵ��Alg44����һ�����ж���ģʽ��PilZ�۵�����,�ܹ���c-di-GMP���,���������εķ��ڡ����Foletti��[32]���ָ��������������Gup-Gup-Nap-Arg��ˮ�н��c-di-GMP������������ӵ�и��ߵ�ѡ����,����������ͭ�̼ٵ���������Ĥ����������һ����Ϊ������Ĥ�ĵĽṹ�Ż��ṩ�˲ο���
3 ������Ĥ��Ŀǰ���ڵ����⼰���˼·
���ڿ�����Ĥ�ľ�����������Ĥ�γɻ�����Ѿ����������Ĥ���ص�,Խ��Խ����о�����Ϊ,�����Ƿ���ϸ������Ĥ��Ⱦ��DZ������ҩ��,��Ŀǰ���������������ȱ��ʹ�����Ӧ�����ٴ���
3.1 �����ȶ��Բ�
��Ȼ���������ں����ױ�����øˮ���Լ�Ѫ����˥�ڶ̡����,�����߿�����Ĥ�������ڵ��ȶ���������Ҫ���������Ľṹ��������Ҫ��һ��,Ŀǰ����ͨ����ֱ���Ļ�������L-�Ͱ��������ΪD-�Ͱ����ᡢ�ı�������е������߶�������ᡢ�ı����������/��ˮ�ԡ�����ҩ��ѧ���������Ľ��а������ֶο���߿�����Ĥ�ĵ��ȶ��Ժ��ӳ�Ѫ����˥��[33]��
3.2 ϸ�����Խϴ�
������Ĥ�������������ϸ�����ֳ�һ������Ѫ�Ի�ϸ�����ԡ�Ϊ��,��ζԿ�����Ĥ�Ľ��������Ե�������Ҫ��һ�����ǽ���ԭ���ĵ�ϸ������,��Kang��[34]ͨ����Pseudin-2����һϵ������,ͨ��ȡ��ɫ�����˿������������ˮ��,��õ�Pse-Anal6��Pse-Anal7������������ϸ�����Ժ���Ѫ�ԡ�����,�䱻������ȡ��������Ľض���Pse-T3��Pse-T4Ҳ�ܼ���ϸ�����Ժ���Ѫ�ԡ���һ�������ڽϵ�ϸ�����ԵĻ�����,���ӿ�����Ĥ�ĵĿ�����Ĥ����,��Yu��[35]��SPLUNC1-��4�����ṹ������������������ɫ����ĺ����ϳ�SPLUNC1-��4M1,��һ�ı������������ĵĿ�����Ĥ����,����û�������ĵ���Ѫ�Ժ�ϸ�����ԡ�����֮��,Kłodzi��ska��[36]���ø�����������ɵ�������������������Ĥ��DJK-5��,���������е�DJK-5��δ�������ĵĶ��Խ�����4��,����Ӱ���ĵĿ������ܡ�˵��ͨ������ҩ��ѧ���ֶ�Ҳ���Խ��Ϳ�����Ĥ�ĵĶ������á�
3.3 �ϳɷ��ð���
����������Ȼ�����,������Դȴʮ�����ޡ��о�������Դ�����ӻ��Գɷֿ����ϵ�������Դ�Կ�����Ĥ�ĵı���[37],���Ƿ��봿���Ѷȴ��ʵ�,����ֱ������ʹ����Ҫ����Ȼ��ѧ�ϳɵķ������Խ��������Ĥ�IJ����͵�����,�������ɱ��ߡ����û��̵ķ������������Ĥ���ƺ��Ǹ�������ѡ��Ŀǰ,����������Ҫ�ı���ϵͳ����ԭ�˱���ϵͳ���˾�����ϵͳ����������ϵͳ���ݲ�ѿ�߸˾�����ϵͳ������˱���ϵͳ�Ͷ�ֲ�����ϵͳ��,����Ȼ�����ڱ�������жԵ���ø��Ϊ����,�����ʵ͵��³ɱ��ߵ����⡣����ͨ��������ǿø�ȶ���,�����ת��ȷ��,��ȡ�ʵ�������߲�Ʒ�����ʵȰ취,������߿�����Ĥ�ĵ�����Ч�ʡ����ͳɱ�,����ʵ�ֹ�ģ���Ͳ�ҵ��[38]��
3.4 ������ҩ
���ڿ�����Ĥ�������IJ��㡢�����ɱ������Լ�ר���������Ĥ��ҩ�↑�����ڽϳ���ȱ�ݡ����,�����п����صĻ����Ͽ�չ�뿹����Ĥ�ĵ�������ҩ�����ش����塣Dosler��[39]ͨ���о�3�ֿ�����Ĥ�ĺ�5�ֿ�����������ҩ,���������Ĥ���뿹��������ʹ��������Ԥ�������ӻ���ҩ�Եij��֡����Dosler��[40]�ַ��ֿ�����Ĥ�ļ�ʹ��1/10MICŨ��������,Ҳ��������ǿ�����ص�ɱ������,����ijЩ�����ص���С����Ĥ����Ũ�ȣ�MBEC��������8����Lashua��[41]������WLBU2�����ӿ�����,���ײ�ù�ء�����ɳ�ǡ�ͷ����ड�������������ʱ������ǿ�˿����ض�����Ĥϸ����ɱ�˻��ԡ������������в���ȷ,�Ʋ����£���1��������Ĥ��������EPS��ͨ�ԡ���Martinez��[42]���ù��۽�����ɨ����������,P5������Ĥ�Ŀ����ƻ�ͭ�̼ٵ���������Ĥԭ�нṹ������ʱ���ϱ���������ɱ����ϸ�����ԵĿ�����,��ʹ��������Ĥ�ھֲ�Ũ�����߶���Эͬ����;��2��������Ĥ�Ļָ�����Ĥϸ����л���ԡ�����Ĥϸ���������ڻ��Խϵ������ǿ��������״̬,��ͨ�������ضԵʹ�л���Ե�ϸ�����ý���,�Ӷ����¸߶ȵĿ�������ҩ[18],������Ĥ�Ŀ��ܴ�ʹ����Ĥϸ��������̬���¸���,������ҩ֮���ʹ����Ĥϸ���Կ����ظ�������;��3��������Ĥ��Ӱ������Ĥ����̶�,�����ڿ����ط���ɱ�����á���ǰ������,ijЩ������Ĥ��ֱ�Ӳ�������Ĥ�γɵĵ��ع���,�䱾����������ɱ������,���ɽ�������Ĥ�ĸ߶Ƚṹ��,����ʹ�ÿ����غ�������ں��߷���ɱ�����á�
4 Ӧ�ü�չ��
����������Ĥ������²�����ҩ�Բ��ϼӾ�,������Ĥ������������ḻ�����û��ƶ��ؼ����ײ�����ҩ�Ե��ص���ܵ��㷺�Ĺ�ע������������һЩ�������ı�Ӧ�����ٴ�ʵ��������,��Xoma��˾�ϳɵ�Neuprex��Ӧ������Ĥ��˫���Ѫ֢������EntoMed SA ��˾�ϳɵ�Thanatin �����ı�Ӧ�������ߵ��»���ȫ���������Ⱦ�Ͷ�����ҩ����Ⱦ��Micrologix ���\����˾�ϳɵ�MBI-�ı�Ӧ���������ص�Ѫ����Ⱦ����ǻ��Ⱦ��Demgen��Dow��ҩ��˾�ϳɵ�Histatin �����ı�Ӧ���������ס���ǻ��Ⱦ�Ϳ�ǻ�������Ⱦ[37]��
δ��������̽������չ������Ĥ�ĵ�������ܡ�������Ĥ�Ľ����Զ�����ʽӦ���������еĸ�������,��ϸ������Ĥ����ĸ�Ⱦ�����Ž�����һ�����
��������������Ϊ��ҵ����ѧϰ����Ȩ��ԭ����ԭ��־���У�������Ȩ������ϵɾ�������±�ע���������³����������Ķ�ԭ�ļ��ο����ף����Ķ�ԭ��־
