
’™“™: ƒњµƒ їщ”Џ∞я¬н”г≈Ѕљр…≠÷Ґ(Parkinson°ѓs disease£ђPD)ƒ£–Ќ£ђ—–Њњ“ї÷÷њє—хїѓїо–‘ ЃлƒYSQLENEFDR (Tyr-Ser-Gln-Leu-Glu-Asn-Glu-Phe-Asp-Arg)µƒњє≈Ѕљр…≠÷Ґїо–‘°£ЈљЈ® µ—йЈ÷ќ™њ’∞„ґ‘’’„й£ђ50 ¶ћmol°§L-11-Љ„їщ-4-±љїщ-1, 2, 3, 6-Ћƒ«вяЅа§(1-methyl-4-phenyl-1£ђ2£ђ3£ђ6-terahydropyridine£ђMPTP)”’µЉ„й“‘Љ∞50 ¶ћmol°§L-1 MPTPЇЌ≤їЌђ≈®ґ»(2°Ґ10°Ґ50 mg°§L-1)µƒ Ѓлƒє≤і¶јн„й°£ №ЊЂЇу4d(day post fertilization£ђdpf)£ђ”√”Ђєвѕ‘ќҐЊµєџ≤м≤ҐЉ«¬ЉЄч µ—й„й„™їщ“т∞я¬н”гґа∞Ќ∞Ј…сЊ≠‘™ЇЌƒ‘≤њ—™є№µƒЈҐ”э«йњц; 5dpf ±£ђ”√Zebrabox∞я¬н”г––ќ™Ј÷ќц“«Ј÷ќц“∞…ъ–Ќ∞я¬н”г––ќ™≤ҐЌ®єэqRT-PCRЉм≤в≈Ѕљр…≠ѕаєЎїщ“тµƒ±ніп±дїѓ°£љбєы ”лњ’∞„ґ‘’’„йѕа±»£ђMPTPі¶јн„й∞я¬н”г≥цѕ÷ґа∞Ќ∞Ј…сЊ≠‘™»± І°Ґƒ‘≤њ—™є№Ћр…Ћ°Ґ≈Ѕљр…≠„і––ќ™ЇЌ≈Ѕљр…≠ѕаєЎїщ“т±ніп“м≥£°£”лMPTPі¶јн„й±»љѕ£ђє≤і¶јн„й∞я¬н”г±нѕ÷≥цґа∞Ќ∞Ј…сЊ≠‘™»± ІЉх…ў°Ґƒ‘≤њ—™є№Ћр…ЋљµµЌ°Ґ≈Ѕљр…≠„і––ќ™їЇљвµƒ±н’ч£ђѕаєЎїщ“т±ніп«ч”Џ’э≥£°£љб¬џ њє—хїѓїо–‘ ЃлƒYSQLENEFDRЊя”–њє≈Ѕљр…≠÷Ґїо–‘°£
≈Ѕљр…≠÷Ґ(Parkinson°ѓs disease£ђPD) «÷–јѕƒк»Ћ≥£ЉыµƒµЏґюіу…сЊ≠ѕµЌ≥ЌЋ–––‘Љ≤≤°£ђ65Ћк“‘…ѕ»Ћ»Ї÷–їЉ≤°¬ Єяіп1.7%£ђљціќ”Џ∞Ґґы„»Ї£ƒђ÷Ґ[1]°£Є√≤°Ѕўі≤÷Ґ„і÷ч“™ќ™’р≤ь°Ґґѓ„ч≥ўїЇ°ҐЉ°«њ÷±µ»––ќ™–‘’ѕ∞≠£ђїєїб∞йЋж”–Ћѓ√я’ѕ∞≠°Ґ»ѕ÷™є¶ƒ№ѕ¬љµ°Ґ„‘÷ч…сЊ≠є¶ƒ№ќ…¬“µ»“їѕµЅ–Ј«––ќ™–‘’ѕ∞≠[1-2]°£PD÷ч“™≤°јнћЎ’чќ™ЇЏ÷ ґа∞Ќ∞Ј(dopamine£ђDA)…сЊ≠‘™µƒЉх…ўЇЌ–ќ≥…“‘¶Ѕ-synќ™÷ч“™≥…Ј÷µƒ¬Ј“„–°ће(Lewy-body£ђLB)[3]°£PDµƒЈҐ≤°їъ÷∆Єі‘”ґа±д£ђƒњ«∞…–ќі√ч»Ј°£µЂ”–—–ЊњЈҐѕ÷£ђPD”л„‘ …ѕµЌ≥”–єЎ£ђ„‘ …ѕµЌ≥µƒЋр…ЋїбµЉ÷¬µ∞∞„µƒїэјџЇЌЊџЉѓ£ђ≤ъ…ъѕЄ∞ыґЊ–‘ЇЌ…сЊ≠ЌЋ–––‘≤°±д°£“≤”–—–Њњ±н√ч£ђ—хїѓ”¶Љ§°Ґµ∞∞„√ЄћеЋр…Ћ“‘Љ∞ѕяЅ£ћеє¶ƒ№ќ…¬“µ» «PD«±‘ЏµƒЈҐ≤°їъ÷∆°£љьƒкјі“—Њ≠ЈҐѕ÷µƒ”лPDѕаєЎµƒїщ“т∞ьј®¶Ѕ-syn°Ґparkin°Ґdj1µ»[4-5]°£¶Ѕ-synЇЌparkinїщ“тµƒЌї±д єЈЇЋЎµ∞∞„√Єћељµљв£ђі”ґшµЉ÷¬ґЊ–‘µ∞∞„ЊџЉѓ≥Ѕїэ[3, 6]; parkinµƒїщ“тЌї±дїб‘м≥…ѕяЅ£ћеє¶ƒ№ќ…¬“[7]; dj1Њя”–њє—хїѓ”¶Љ§µƒ„ч”√[8]µ»°£
Ї£—у…ъќпћеƒЏЄїЇђґа÷÷…ъќпїо–‘їѓЇѕќп£ђЊя”–њє—хїѓµƒ…ъќпїо–‘£ђ“тіЋ‘Џ‘§ЈјЇЌ÷ќЅ∆¬э–‘Ј«іЂ»Њ–‘Љ≤≤°(»зPD°Ґ∞Ґґы„»Ї£ƒђ÷Ґµ»)Њя”–«±‘ЏµƒЉџ÷µ[9]°£÷оґа µ—й÷§ µ£ђњє—хїѓЉЅїтЊя”–њє—хїѓїо–‘µƒќп÷ ƒ№єї”––І«е≥эїо–‘„‘”…їщ£ђЊя”–…сЊ≠±£ї§„ч”√[10]°£‘Џ»ЋћеƒЏ£ђѕяЅ£ћеЇфќьЌ®≥£їб≤ъ…ъ“ї–©«њ—хїѓЉЅ£ђComplex Iµƒ“÷÷∆„ч”√їб‘цЉ”їо–‘—х(reactive oxygen species£ђROS)≥ђ—хїѓќпµƒ≤ъ…ъ£ђROS≥ђ—хїѓќпњ…–ќ≥…”–ґЊµƒф«„‘”…їщ£ђїт’я–ќ≥…єэ—х—«ѕхїщ£ђ’в–©Ј÷„”„ч”√”ЏЇЋЋб°Ґµ∞∞„÷ ЇЌ÷ђјаµ»ќп÷ ґш“э∆рѕЄ∞ыЋр…Ћ[7]°£њќћв„й«∞∆Џ—–ЊњЈҐѕ÷£ђјі‘і”ЏЇ£¬Ё»вћб»°ќп÷–µƒ“ї÷÷ ЃлƒYSQLENEFDRЊя”–љѕ«њµƒњє—хїѓƒ№Ѕ¶£ђњ…«е≥эћеЌв„‘”…їщЉ∞“÷÷∆∞я¬н”гћеƒЏ—хїѓЋр…Ћ[11]£ђµЂЇ£¬Ё‘іњє—хїѓїо–‘ ЃлƒYSQLENEFDR «ЈсЊя”–њєPDїо–‘»‘ќіњ…÷™°£ƒњ«∞≥£ЉыµƒPD÷ќЅ∆“©ќпґ‘”Џ»ЋћеґЉ”–їтґаїт…ўµƒЄ±„ч”√£ђ»зЈю”√„у–эґа∞ЌЉЂ“„“эЈҐќЄ≤њ≤їЅЉЈі”¶[12]; ґа∞Ќ∞Ј №ћеЉ§ґѓЉЅњ…”––ІЄƒ…∆їЉ’яµƒЅўі≤÷Ґ„і£ђµЂїб‘м≥…їЉ’яґс–ƒ°Ґ І√яµ»≤їЅЉЈі”¶[13]°£≤Ґ«“’в–©“©ќп÷їƒ№Єƒ…∆÷Ґ„і£ђ≤Ґ≤їƒ№—”їЇPDљш≥ћ°£ћм»їјі‘іµƒ ≥‘і–‘ ЃлƒYSQLENEFDR≥э∆д„‘…нЋщЊя”–Єяњє—хїѓїо–‘Ќв[11]£ђѕаґ‘ЄьЉ”∞≤»Ђ£ђЄь ”√”Џ≥§∆ЏЈю”√°£
∞я¬н”гће–Ќ–°«“ЌЄ√ч°ҐЈҐ”эЈ±÷≥њм°ҐЋ«—шЊ≠Љ√«“Њя”–ќё–иљв∆ ±гњ…єџ≤мЄч∆чєўЈҐ”э«йњцµƒ”≈ ∆[16]°£„о÷Ў“™µƒ «£ђ∞я¬н”гµƒїщ“т–тЅ–”л»Ћјаµƒїщ“т–тЅ–”–„≈ЉЂЄяµƒѕаЋ∆ґ»£ђЋщ“‘±ї»ѕќ™ «”≈ЅЉµƒљ®ЅҐ…сЊ≠ЌЋ–––‘Љ≤≤°ƒ£–Ќµƒґѓќп„ ‘і[14]°£Vmat°ҐFli1Ј÷±рћЎ“м–‘±ніп”Џ∞я¬н”гDA…сЊ≠‘™”л—™є№÷–£ђ“тіЋ„™їщ“т∞я¬н”гVmat:GFP°ҐFli1:GFPЈ÷±р”√”Џ∆јЉџDA…сЊ≠‘™°Ґƒ‘≤њ—™є№µƒЈҐ”э„іњц°£1-Љ„їщ-4-±љїщ-1, 2, 3, 6-Ћƒ«вяЅа§(1-methyl-4-phenyl-1£ђ2£ђ3£ђ6-terahydropyridine£ђMPTP) «“ї÷÷…сЊ≠ґЊ–‘ќп÷ £ђ‘Џ∞я¬н”г÷–≥£±ї”√јі”’µЉPDƒ£–Ќ°£
±Њ µ—й“‘∞я¬н”гќ™ƒ£–Ќ£ђ÷ч“™єџ≤м≤ҐЉ«¬Љ≤їЌђі¶јн„йµƒ∞я¬н”гґа∞Ќ∞Ј…сЊ≠‘™“‘Љ∞ƒ‘≤њ—™є№µƒЈҐ”э«йњц£ђ∞я¬н”г––ќ™“‘Љ∞PDѕаєЎїщ“т±ніп±дїѓ£ђ“‘іЋјіћљЊњњє—хїѓїо–‘ ЃлƒYSQLENEFDR «ЈсЊя”–њєPDїо–‘°£Њяће≤љ÷и»зFig 1Ћщ Њ°£
1 ≤ƒЅѕ”лЈљЈ®
1.1 ≤ƒЅѕ
YSQLENEFDR(іњґ»ќ™99.02%)°£„™їщ“т∞я¬н”гVmat :GFP£ђ„™їщ“т∞я¬н”гFli1 :GFP£ђ“∞…ъ–Ќ∞я¬н”гAB∆ЈѕµЊщ”……љґЂ °њ∆—І‘Ї…ъќп—–ЊњЋщ“©ќп…Є—°∆љћ®ћбє©°£
∞я¬н”г≈а—шЉ∞≈яћ•µƒ„Љ±Є£Ї≥…ƒкі∆–џ∞я¬н”гЈ÷њ™≈а—ш”ЏЋЃќ¬26 °ж∞я¬н”г—ш÷≥ѕµЌ≥÷–£ђ’’√чЇЏ∞µ14 h :10 hљїћжљш––£ђ√њ»’ґ® ±ќє ≥°£ µ—й«∞“їЌн£ђ—°‘с≥… м∞я¬н”г∞іі∆–џ±»јэ2 :2£ђЈ≈»л≤ъ¬—Є„÷–°£іќ»’‘з…ѕ8£Ї30≥й»°Єф∞е£ђ2 hЇу ’»°”г¬—£ђљЂЋј¬—ћё≥э£ђ—ш÷≥ЋЃЈіЄі≥еѕі3іќ£ђ“∆»лЇђ2 mg°§L-1—«Љ„їщјґµƒ—ш÷≥ЋЃ÷–£ђ÷√”Џ26 °жЇгќ¬≈а—шѕд÷–њЎєв≈а—ш£ђ“‘±ЄЇу–ш µ—й є”√°£
MPTP°Ґ—«Љ„їщјґ°Ґ1-±љїщ-2-Ѕтле(PTU)(√јєъSigmaєЂЋЊ); RNA ‘ЉЅЇ–(RN2802£ђ±±Њ©∞ђµ¬ј≥…ъќпњ∆ЉЉ”–ѕёєЂЋЊ); Јі„™¬Љ ‘ЉЅЇ–°Ґ”Ђєвґ®ЅњPCR ‘ЉЅЇ–(RR036A°ҐRR091A TaKaRaєЂЋЊ)°£
∞я¬н”г—ш÷≥Ћ«—шѕµЌ≥(±±Њ©∞Ѓ…ъњ∆ЉЉєЂЋЊ)°Ґће ””Ђєвѕ‘ќҐЊµ(µ¬єъ≤ћЋЊєЂЋЊ)°Ґ µ ±”Ђєвґ®ЅњPCR“«(¬ё ѕ’пґѕ≤ъ∆Ј”–ѕёєЂЋЊ)°Ґ≥ђќҐЅњЈ÷євєвґ»Љ∆(їщ“т”–ѕёєЂЋЊ)°ҐC1000 TouchћЁґ»PCR“«(Bio-RadєЂЋЊ)°ҐZebrabox∞я¬н”г––ќ™Ј÷ќц“«(ViewpointєЂЋЊ)µ»°£
1.2 ЈљЈ®
1.2.1 ЃлƒYSQLENEFDRµƒ÷∆±Є
їщ”Џњќћв„й“—љ®ЅҐµƒЋбіЉљюћбЈ®љбЇѕїо–‘µЉѕтЈ÷јліњїѓЈ®[10]£ђ÷∆±ЄЇ£¬Ёїо–‘ Ѓлƒ£ђЉі»°ѕг¬Ё„й÷ѓ£ђƒ•Ћй£ђ‘»љђ£ђЉ”»л10±ґЅњµƒЋб–‘ѕ°““іЉ»№“Ї(±щі„Ћбµч’ы»№“ЇpH÷µ÷Ѕ5)£ђ30 °жљЅ∞ићб»°6 h£ђјл–ƒ£ђ»°…ѕ≤г«е“Ї£ђЉх—є≈®Ћх÷ЅќёіЉќґ£ђ’эЉЇЌйЁЌ»°Ќ—≥э÷ђ»№–‘≤њЈ÷£ђѕ¬≤г»№“Їјдґ≥±£іж£ђЉіµ√і÷ґалƒћб»°ќп°£ЋжЇу£ђЌ®єэћеЌвїо–‘µЉѕтЈ÷јлЈ®£ђЈ÷јліњїѓЄїЉѓµ√µљЇ£¬Ё Ѓлƒ—щ∆Ј£ђЊ≠µ∞∞„ґ®–‘»Јґ®∆д∞±їщЋб„й≥…£ђ≤…”√єћѕаЇѕ≥…ЉЉ х£ђ÷∆±ЄYSQLENEFDR (Tyr-Ser-Gln-Leu-Glu-Asn-Glu-Phe-Asp-Arg)—щ∆Ј°£
1.2.2 µ—йЈ÷„йЉ∞і¶јн
љЂ №ЊЂЇу1 dµƒ≈яћ•љш––Ќ—ƒ§і¶јн£ђЈ≈”Џ6њ„∞е÷–£ђ√њњ„Ј≈»л20ћх∞я¬н”г£ђЈ÷ќ™“‘ѕ¬5„й£Їњ’∞„ґ‘’’„й°Ґ50 ¶ћmol°§L-1 MPTPі¶јн„й°Ґ50 ¶ћmol°§L-1 MPTP”л≤їЌђ≈®ґ»(2°Ґ10°Ґ50 mg°§L-1)њє—хїѓїо–‘ ЃлƒYSQLENEFDRє≤і¶јн„й°£ЅнЌв–и“‘0.003%(W/V)PTUі¶јн„™їщ“т∞я¬н”гVmat :GFP°ҐFli1 :GFP£ђƒњµƒ‘Џ”Џ“÷÷∆ЇЏ…ЂЋЎµƒ–ќ≥…£ђ±г”ЏЇу∆Џ‘Џѕ‘ќҐЊµѕ¬єџ≤мЉ«¬Љ°£Љ”Ќк“©Їу£ђљЂ∆дЈ≈»л≈а—шѕдƒЏ≈а—ш£ђ√њ»’її“©°£
1.2.3 ґа∞Ќ∞Ј…сЊ≠‘™Љ∞ƒ‘—™є№µƒЉм≤в
4 dpf ±£ђ‘Џ≤ћЋЊће ””Ђєвѕ‘ќҐЊµѕ¬£ђ—°»°≤їЌђ µ—й„йµƒ„™їщ“т∞я¬н”гVmat :GFP°ҐFli1 :GFPЈ÷±рєџ≤мDA…сЊ≠‘™”лƒ‘≤њ—™є№ЈҐ”э«йњц°£
1.2.4 ––ќ™—ІЉа≤в
5 dpf ±£ђљЂ“∞…ъ–Ќ∞я¬н”гABµƒ≤їЌђ„й±рЈ≈»л48њ„∞е÷–£ђ1ћх/њ„£ђЉ”—ш”гЋЃ1 mL/њ„£ђљЂ48њ„∞еЈ≈»лZebrabox∞я¬н”г––ќ™Ј÷ќц“«∞µѕд÷–£ђѕ» є∆д ”¶їЈЊ≥°£15 minЇуњ™ Љљш––––ќ™—ІЉм≤в£ђЉм≤в ±Љд…и÷√ќ™20 min°£‘Ћ”√Zeblab»нЉюљш–– эЊЁі¶јн£ђЉ∆Ћг√њћх”г”ќґѓ„№ЊајлЇЌ∆љЊщЋўґ»°£
1.2.5 ”Ђєвґ®ЅњPCR(quantitative real-time PCR£ђqRT-PCR)Љм≤вPDѕаєЎїщ“тµƒ±ніп
√њ„й»°5 dpfµƒ“∞…ъ–Ќ∞я¬н”гAB”„”г(n=20)љш––qRT-PCRЉм≤в°£RNAµƒћб»°≤ќ’’ ‘ЉЅЇ–Ћµ√ч й£ђјы”√≥ђќҐЅњЈ÷євєвґ»Љ∆≤вґ®ћб»°µƒRNAµƒ≈®ґ»£ђA260/A280”¶‘Џ2.0~2.5÷ЃЉд; RNAЈі„™ќ™cDNA≤ќ’’Јі„™¬Љ ‘ЉЅЇ–Ћµ√ч й; Real-time PCR≤ќ’’ µ ±”Ђєвґ®ЅњPCRЋµ√ч й°£Real-time PCRј©‘цљб шЇу д≥цґ‘’’„йЇЌіэ≤в„йƒњµƒїщ“тCTЉ∞ƒЏ≤ќїщ“тrpl13aµƒCT÷µ£ђ“‘ѕаґ‘ґ®ЅњЈ®”√2-¶§¶§CTЉ∆ЋгЄч„йїщ“тµƒѕаґ‘±ніпЅњ°£
1.2.6 эЊЁі¶јн
”√Graphpad prism 7.0»нЉюґ‘ µ—йљбєыљш––One-way ANOVAЉм—й£ђ≤Ґ“‘x°јs±н Њ°£
2 љбєы
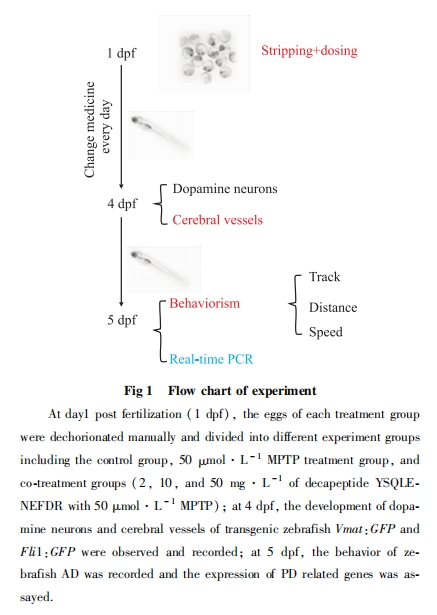
2.1 ЃлƒYSQLENEFDRґ‘∞я¬н”гDA…сЊ≠‘™µƒ±£ї§„ч”√
4 dpf ±∞я¬н”гDA…сЊ≠‘™“—ЈҐ”эЌк»Ђ°£і”∞я¬н”гDA…сЊ≠‘™«ш”т≥§ґ»јіњі£ђFig 2Ћщ Њ£ђ”лњ’∞„ґ‘’’„йѕа±»£ђMPTPі¶јн„йµƒDA…сЊ≠‘™≥§ґ»√чѕ‘Љх…ў«“Њя”–Ќ≥Љ∆—І“в“е; ”лMPTPі¶јн„йѕа±»£ђ≤їЌђ≈®ґ»µƒ ЃлƒYSQLENEFDR”лMPTPє≤і¶јн„й÷–DA…сЊ≠‘™≥§ґ»÷рљ•‘цЉ”«“ЊщЊя”–Ќ≥Љ∆—І“в“е°£і”∞я¬н”гDA…сЊ≠‘™«ш”т”Ђєв√№ґ»„№ЇЌјіњі£ђ”лњ’∞„ґ‘’’„йѕа±»£ђMPTPі¶јн„йµƒDA…сЊ≠‘™«ш”т”Ђєв√№ґ»„№ЇЌ√чѕ‘ѕ¬љµ«“≤о“мЊя”–Ќ≥Љ∆—І“в“е; ”лMPTPі¶јн„йѕа±»£ђ ЃлƒYSQLENEFDR”лMPTPє≤і¶јн„й÷–DA…сЊ≠‘™«ш”т”Ђєв√№ґ»‘Џ≈®ґ»ќ™2 mg°§L-1 ±”–¬‘ќҐµƒ…эЄя£ђµЂ≤о“мќёЌ≥Љ∆—І“в“е£ђґш‘Џ10 mg°§L-1ЇЌ50 mg°§L-1 ±√чѕ‘…эЄя°£„џ…ѕЋщ ц£ђ‘Џ ЃлƒYSQLENEFDR≈®ґ»ќ™50 mg°§L-1 ±ґ‘DA…сЊ≠‘™µƒ±£ї§„ч”√„оЉ—°£—–Њњљбєы±н√ч£ђ ЃлƒYSQLENEFDRґ‘DA…сЊ≠‘™Њя”–±£ї§„ч”√£ђ«“≥ ѕ÷≈®ґ»“јјµ–‘°£
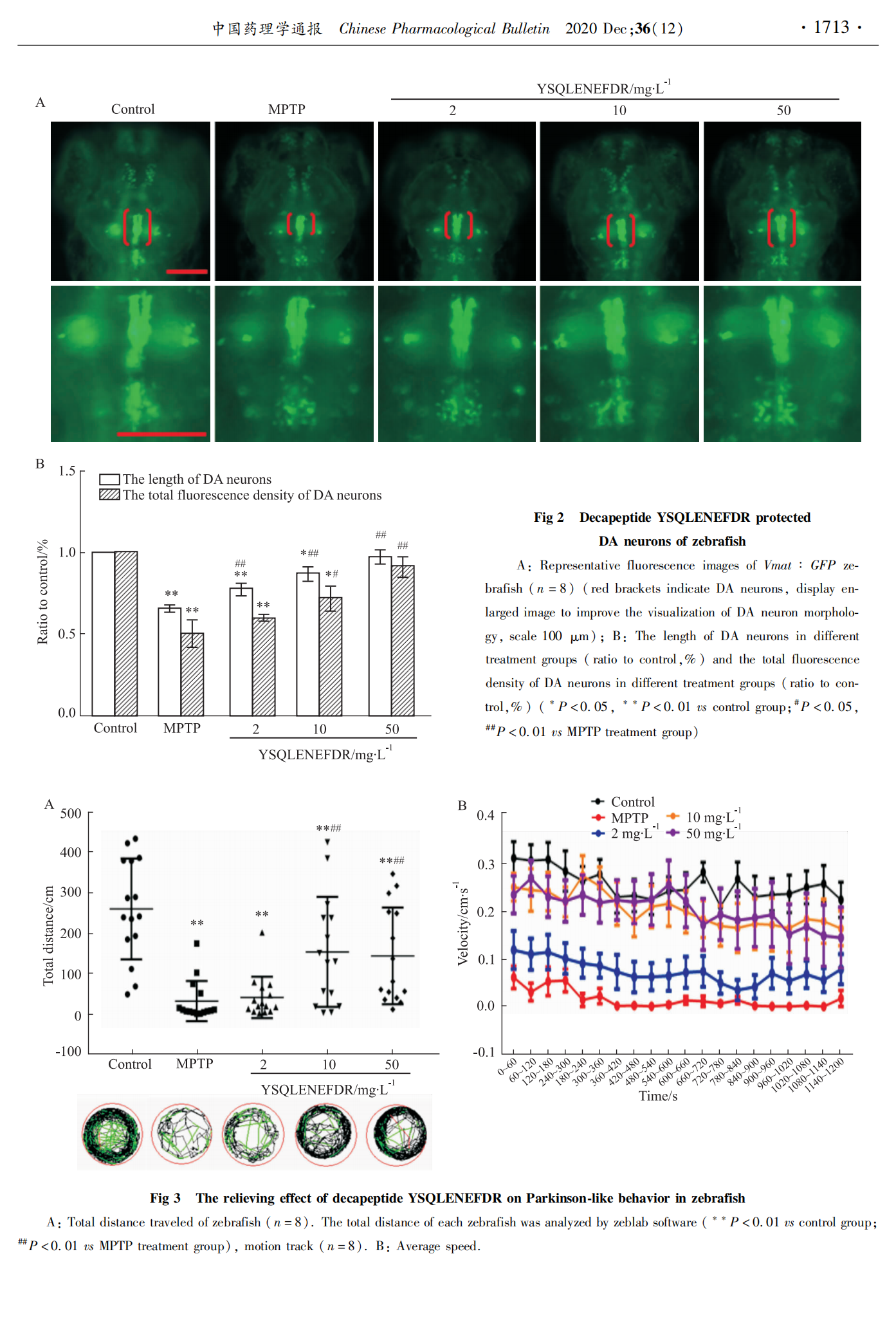
2.2 ЃлƒYSQLENEFDRњ…“‘їЇљв∞я¬н”гPD„і––ќ™
5 dpf ±£ђґ‘≤їЌђ µ—й„йµƒ“∞…ъ–Ќ∞я¬н”гAB(n=8)љш––––ќ™—І≤в ‘£ђљбєыЉыFig 3°£Fig Aќ™≤їЌђ„й±рµƒ“∞…ъ–Ќ∞я¬н”гABµƒ”ќґѓ„№ЊајлЇЌ‘Ћ––ємЉ££ђємЉ£ЌЉ÷–Їм…Ђѕя°Ґ¬ћ…Ђѕя°ҐЇЏ…ЂѕяЈ÷±р±н Њ”ќґѓЋўґ»µƒњм°Ґ÷–°Ґ¬эЋў°£”лњ’∞„ґ‘’’„йѕа±»£ђMPTPі¶јн„й∞я¬н”г”ќґѓ„№Њајл√чѕ‘Љх…ў(Fig 3A£ђMPTP„йЋщ Њ)£ђ∆љЊщЋўґ»±д¬э(Fig 3B£ђMPTP„йЋщ Њ); ”лMPTPі¶јн„йѕа±»£ђMPTP”л ЃлƒYSQLENEFDRє≤Ќђі¶јн„й‘Џ2 mg°§L-1 ±”ќґѓ„№Њајл¬‘ќҐ‘цЉ”µЂ≤о“м≤їЊя”–Ќ≥Љ∆—І“в“е£ђ10 mg°§L-1ЇЌ50 mg°§L-1є≤і¶јн„й”ќґѓ„№Њајл√чѕ‘‘цЉ”£ђ∞я¬н”гµƒ‘Ћґѓƒ№Ѕ¶ї÷Єі«“Њя”–Ќ≥Љ∆—І“в“е(Fig 3A£ђ2°Ґ10°Ґ50 mg°§L-1„йЋщ Њ)£ђ∆љЊщЋўґ»±дњм(Fig 3B£ђ2°Ґ10°Ґ50 mg°§L-1„йЋщ Њ)°£—–Њњљбєы±н√ч£ђ ЃлƒYSQLENEFDRњ…“‘їЇљв”…MPTP‘м≥…µƒ∞я¬н”гPD„і––ќ™£ђ«“Њя”–“їґ®µƒ≈®ґ»“јјµ–‘°£
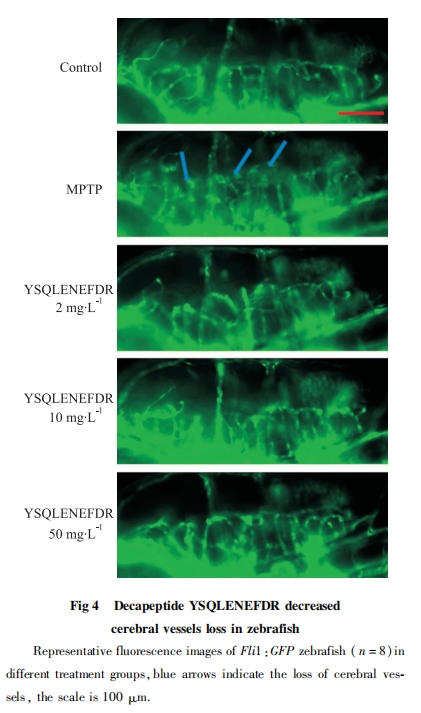
2.3 ЃлƒYSQLENEFDRњ…“‘’ьЊ»∞я¬н”гƒ‘—™є№Ћр…Ћ
4 dpf ±£ђњ’∞„ґ‘’’„й÷–∞я¬н”гƒ‘—™є№«еќъњ…Љы(Fig 4£ђcontrol„йЋщ Њ)°£”лњ’∞„ґ‘’’„йѕа±»£ђMPTPі¶јн„й÷–∞я¬н”гƒ‘—™є№—ѕ÷ЎЋр…Ћ£ђ—™є№√№ґ»√чѕ‘Љх…ў(Fig 4£ђMPTP„йЋщ Њ)°£”лMPTPі¶јн„йѕа±»£ђ ЃлƒYSQLENEFDR”лMPTPє≤і¶јн„й÷–ƒ‘—™є№√№ґ»Ћж„≈ ЃлƒYSQLENEFDRµƒ≈®ґ»‘цЉ”÷рљ•‘цЉ”£ђ∆д÷–50 mg°§L-1 ЃлƒYSQLENEFDR”лMPTPє≤і¶јн„й’ьЊ»їЎµƒƒ‘—™є№ эЅњ„оґа(Fig 4£ђ50 mg°§L-1„й)°£—–Њњљбєы±н√ч£ђ ЃлƒYSQLENEFDRњ…“‘Єƒ…∆”…MPTP‘м≥…µƒƒ‘—™є№Ћр…Ћ£ђ≤Ґ«“‘Џ“їґ®ЈґќІƒЏ£ђ≈®ґ»‘љЄяЄƒ…∆–Ієы‘љ√чѕ‘°£
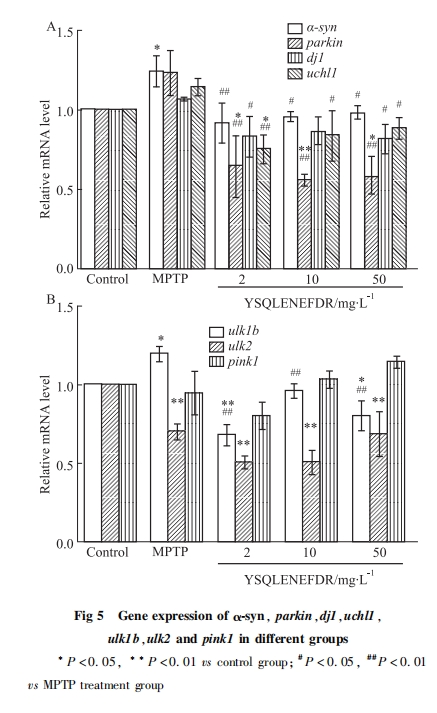
2.4 ЃлƒYSQLENEFDRґ‘PDѕаєЎїщ“т±ніпµƒ”∞ѕм
ќ™ЅЋљш“ї≤љ—–Њњ ЃлƒYSQLENEFDRњєPDїо–‘µƒƒЏ‘Џїъ÷∆£ђ≤…”√qRT-PCRЉм≤в≤їЌђ µ—й„й(n=20) PDѕаєЎїщ“т¶Ѕ-syn°Ґparkin°Ґdj1°Ґuchl1°Ґulk1b°Ґulk2ЇЌpink1‘Џ∞я¬н”гћеƒЏµƒ±ніпЋЃ∆љ°£”лњ’∞„ґ‘’’„йѕа±»£ђMPTPі¶јн„й÷–¶Ѕ-syn°Ґulk1bµƒ±ніп√чѕ‘…ѕµч£ђulk2µƒ±ніп√чѕ‘ѕ¬µч£ђґшparkin°Ґdj1°Ґuchl1°Ґpink1µƒ±ніп±дїѓќёЌ≥Љ∆—І“в“е°£”лMPTPі¶јн„йѕа±»£ђЊ≠ ЃлƒYSQLENEFDRі¶јнЇу£ђ¶Ѕ-syn°Ґparkin°Ґuchl1ЇЌulk1bµƒ±ніп√чѕ‘ѕ¬µч«“Њя”–Ќ≥Љ∆—І“в“е; dj1‘Џ2 mg°§L-1ЇЌ50 mg°§L-1 ±µƒ±ніп√чѕ‘ѕ¬µч£ђ10 mg°§L-1≤Ґќё√чѕ‘±дїѓ; ulk2°Ґpink1‘Џ2°Ґ10ЇЌ50 mg°§L-1Њщќё√чѕ‘±дїѓ°£љбєыѕ‘ Њ£ђЊ≠ ЃлƒYSQLENEFDRі¶јнЇу£ђ≥эulk2°Ґpink1ЌвЄчїщ“т‘Џ∞я¬н”гћеƒЏµƒ±ніпЋЃ∆љЊщ”–√чѕ‘±дїѓ£ђЋµ√ч ЃлƒYSQLENEFDRƒ№єї”∞ѕм”…MPTP‘м≥…µƒїщ“т“м≥£±ніп°£
3 ћ÷¬џ
PD «љціќ”Џ∞Ґґы„»Ї£ƒђ÷ҐµƒµЏґюіу…сЊ≠ЌЋ–––‘Љ≤≤°£ђ—ѕ÷Ў”∞ѕмїЉ’яµƒ…ъїо÷ Ѕњ£ђћЎ±р «јѕƒк»Ћ£ђЋщ“‘—–Њњ»зЇќ÷ќЅ∆ЇЌїЇљвPD ∆‘Џ±Ў––°£
±Њ µ—й÷Љ‘ЏћљЊњЇ£¬Ё‘іµƒЊя”–њє—хїѓїо–‘ ЃлƒYSQLENEFDRµƒњєPDїо–‘°£ µ—й÷–”√MPTP”’µЉµƒ∞я¬н”гPDƒ£–Ќ£ђ≥цѕ÷ЅЋґа∞Ќ∞Ј…сЊ≠‘™Љ∞ƒ‘≤њ—™є№»± І√чѕ‘£ђ––ќ™“м≥£ЇЌPDѕаєЎїщ“т±ніп“м≥£µ»ѕ÷ѕу£ђЋµ√ч∞я¬н”гPDƒ£–Ќљ®ЅҐ≥…є¶°£”лMPTPі¶јнµƒ∞я¬н”гѕа±»£ђMPTP”л ЃлƒYSQLENEFDRє≤і¶јн„йµƒ∞я¬н”гґа∞Ќ∞Ј…сЊ≠‘™µƒ»± ІЋж„≈ ЃлƒYSQLENEFDR≈®ґ»µƒ‘цЉ””–√чѕ‘µƒЄƒ…∆°£ ЃлƒYSQLENEFDRґ‘”Џ∞я¬н”гPDƒ£–Ќƒ‘≤њ—™є№Ћр…Ћ“≤±нѕ÷≥ц“їґ®µƒ’ьЊ»„ч”√°£MPTP”л ЃлƒYSQLENEFDRє≤і¶јн„йµƒ∞я¬н”г”ќґѓƒ№Ѕ¶√чѕ‘ї÷Єі£ђ”ќґѓ„№Њајл‘цЉ”£ђ∆љЊщ”ќґѓЋўґ»±дњм°£“‘…ѕЄчЉм≤вљбєы±н√ч ЃлƒYSQLENEFDRЊя”–њєPDїо–‘£ђ«“‘Џ≈®ґ»ќ™50 mg°§L-1 ±£ђ∆дїЇљвPDµƒƒ№Ѕ¶„оЉ—°£
µ∞∞„÷ «…ъќпћеƒЏ÷ч“™ЈҐї”є¶ƒ№µƒќп÷ £ђЋщ“‘Ќ®єэЉм≤вµ∞∞„÷ ±ніп±дїѓі”ґш—–Њњ“©ќп°ҐїѓЇѕќпЈҐї”„ч”√µƒ∞–µг°Ґ…ѕѕ¬”ќЌ®¬ЈєЎѕµ «±»љѕ÷±єџµƒЈљЈ®°£µЂ « –√ж…ѕЋщ џєЎ”Џ∞я¬н”гµƒњєће±»љѕ…ў£ђ‘Џ√ї”–Їѕ њєћеµƒ«йњцѕ¬£ђЉм≤вїщ“т±ніп±дїѓ «“їЄц±»љѕЈљ±гµƒЈљЈ®£ђїщ“тµƒ…ѕµчЇЌѕ¬µчњ…“‘Љдљ”Јі”≥µ∞∞„µƒ±дїѓєэ≥ћ°£љьƒкјіµƒ—–Њњ±н√ч£ђ≤ќ”лPDЈҐ…ъЈҐ’єµƒїщ“т∞ьј®¶Ѕ-syn°Ґparkin°Ґdj1°Ґuchl1°Ґulk1b°Ґulk2°Ґpink1°ҐLRRK2°ҐATP13A2°£іуЅњ¶Ѕ-synµ∞∞„ЊџЉѓ–ќ≥…LB£ђµЉ÷¬PD°£±Њ—–ЊњЈҐѕ÷Њ≠MPTPі¶јнЇу£ђ¶Ѕ-synµƒ±ніп√чѕ‘…ѕ…э£ђґш ЃлƒYSQLENEFDRі¶јнЇу£ђ¶Ѕ-synµƒ±ніп√чѕ‘ѕ¬µч£ђћб Њ ЃлƒYSQLENEFDRљµµЌЅЋ¶Ѕ-synµ∞∞„ЊџЉѓ°£parkin„чќ™E3ЈЇЋЎЅђљ”√Є£ђƒ№і”∞ы÷ „™“∆µљѕяЅ£ће÷–£ђЈЇЋЎїѓѕяЅ£ћеµ∞∞„£ђ–ќ≥…„‘ …ће£ђ єЋр…ЋµƒѕяЅ£ћеЌ®єэ„‘ …ће±їљµљв[6]°£Њ≠MPTPі¶јнЇу£ђparkinµƒ±ніп…ѕµч£ђћб ЊMPTPњ…ƒ№ґ‘ѕяЅ£ће‘м≥…Ћр…Ћ£ђїъће„‘ќ“±£ї§і”ґшћбЄяparkinµƒ±ніп°£ґш ЃлƒYSQLENEDRі¶јнЇу£ђparkinµƒ±ніп√чѕ‘ѕ¬µч£ђћб Њ ЃлƒYSQLENEFDRƒ№єїїЇљвѕяЅ£ћеЋр…Ћ°£dj1Њя”–њє—хїѓ”¶Љ§„ч”√£ђ‘Џ—хїѓ”¶Љ§ѕ¬£ђdj1їбѕтѕяЅ£ће«®“∆ЄїЉѓ£ђ‘ЏѕяЅ£ће÷–µƒdj1Њя”–Єь«њµƒѕЄ∞ы±£ї§„ч”√[8]°£Њ≠MPTPі¶јнЇу£ђdj1µƒ±ніп…ѕµч£ђґш ЃлƒYSQLENEFDRі¶јнЇу£ђdj1µƒ±ніп√чѕ‘ѕ¬µч£ђћб Њ ЃлƒYSQLENEFDRњ…ƒ№Њя”–њє—хїѓ”¶Љ§µƒ„ч”√°£uchl1≤ќ”лµчљвѕЄ∞ыµƒ‘ц…ъ°ҐЈ÷їѓЇЌµтЌц[15]°£Њ≠MPTPі¶јнЇу£ђuchl1µƒ±ніп…ѕ…э£ђћб ЊMPTPњ…ƒ№ґ‘ѕЄ∞ы‘м≥…Ћр…Ћ£ђїъће„‘ќ“±£ї§і”ґшћбЄяuchl1µƒ±ніп£ђґш ЃлƒYSQLENEFDRі¶јнЇу£ђuchl1µƒ±ніп√чѕ‘ѕ¬µч£ђћб Њ ЃлƒYSQLENEFDRƒ№єїїЇљвMPTPЋщ‘м≥…µƒѕЄ∞ыЋр…Ћ£ђґ‘ѕЄ∞ы”–±£ї§„ч”√°£ulk1bµƒєэ±ніпїб“÷÷∆„‘ …[15]£ђ”’ЈҐPD°£Њ≠MPTPі¶јнЇу£ђulk1bµƒ±ніп√чѕ‘…ѕ…э£ђћб ЊMPTPњ…ƒ№“÷÷∆„‘ …£ђ ЃлƒYSQLENEFDRі¶јнЇу£ђulk1bµƒ±ніп√чѕ‘ѕ¬µч£ђћб Њ ЃлƒYSQLENEFDRƒ№єїЉ§їо„‘ …°£ulk2ƒ№„и÷є≤ї»№–‘ЈЇЋЎїѓµ∞∞„ЊџЉѓќпµƒїэјџ[15]°£Њ≠MPTPі¶јнЇу£ђulk2µƒ±ніп√чѕ‘ѕ¬µч£ђћб ЊMPTPњ…ƒ№їбµЉ÷¬≤ї»№–‘ЈЇЋЎїѓµ∞∞„ќёЈ®љµљвґшіуЅњїэјџ£ђ єPDґсїѓ£ђ ЃлƒYSQLENEFDRі¶јнЇу£ђulk2µƒ±ніпќё√чѕ‘±дїѓ£ђћб Њ ЃлƒYSQLENEFDRњ…ƒ№≤ї «Ќ®єэµчљЏulk2µƒ±ніпјіїЇљвPD°£“‘Ќщ—–Њњ±н√чpink1є¶ƒ№“м≥£‘м≥…ѕяЅ£ћеЋр…Ћ[7]°£Њ≠MPTPі¶јнЇу£ђpink1µƒ±ніп√ї”–√чѕ‘±дїѓ£ђ ЃлƒYSQLENEFDRі¶јнЇу“≤≤Ґќіґ‘pink1µƒ±ніп‘м≥…√чѕ‘µƒ”∞ѕм£ђћб Њpink1‘ЏMPTP”’µЉµƒ∞я¬н”гPDƒ£–Ќ÷–ќіЈҐї”÷Ў“™„ч”√°£µчљЏPDµƒ÷Ў“™їщ“тїє∞ьј®LRRK2°ҐATP13A2µ»°£LRRK2ЇЌATP13A2Њщ≤ќ”лµчљЏґа∞Ќ∞Ј…сЊ≠‘™Ћр…Ћ°£LRRK2µƒЌї±дƒ№єї‘м≥…ґа∞Ќ∞Ј…сЊ≠‘™µƒЋр…Ћ[16]°£ATP13A2µƒєэ±ніпƒ№√чѕ‘Љх…ўґа∞Ќ∞Ј…сЊ≠‘™µƒ±д–‘ґ™ І[17]°£µЂ ЃлƒYSQLENEFDR «Јсƒ№Ќ®єэµчљЏґа∞Ќ∞Ј…сЊ≠‘™ѕаєЎїщ“ті”ґшїЇљвPD£ђ∆дЊяћеїъ÷∆»‘–иљш“ї≤љ—–Њњ°£
±Њ—–Њњ÷–MPTP”’µЉ„йµƒїщ“т¶Ѕ-synµ»±ніпѕ¬µч£ђulk1bµ»±ніп…ѕµч£ђ”л÷Ѓ«∞—–Њњїщ“т±ніп“ї÷¬£ђЌђ ±“≤÷§√чЅЋ∞я¬н”гPDƒ£–Ќљ®ЅҐµƒ≥…є¶°£≤їЌђ≈®ґ»µƒ ЃлƒYSQLENEFDRƒ№єїµчљЏ”лPDѕаєЎїщ“тµƒ“м≥£±ніп£ђ єЄчїщ“тµƒ±ніп«ч∆љ”Џњ’∞„ґ‘’’„й£ђЌ∆≤в Ѓлƒƒ№єїЌ®єэЉх…ў¶Ѕ-synµ∞∞„µƒЊџЉѓ£ђїЇљвѕяЅ£ћеЋр…Ћ£ђњє—хїѓ”¶Љ§„ч”√µ»£ђјіїЇљвPD£ђµЂЊяћеµƒїъ÷∆»‘–и“™љш“ї≤љµƒ—–Њњ°£
Ї£¬Ёјі‘іµƒњє—хїѓїо–‘ ЃлƒYSQLENEFDRƒ№єї±£ї§∞я¬н”гDA…сЊ≠‘™«“ґ‘”Џƒ‘≤њ—™є№Њя”–Єƒ…∆„ч”√£ђƒ№єїїЇљв≈Ѕљр…≠÷Ґ––ќ™£ђµчњЎ”л≈Ѕљр…≠÷ҐѕаєЎїщ“тµƒ“м≥£±ніп£ђЊя”–њє≈Ѕљр…≠÷Ґїо–‘°£
√в‘р…щ√ч£Ї±Њќƒќ™––“µљїЅч—Іѕ∞£ђ∞ж»®єй‘≠„ч’яЉ∞‘≠‘”÷ЊЋщ”–£ђ»з”–«÷»®£ђњ…Ѕ™ѕµ…Њ≥э°£ќƒ’¬±к„Ґ”–„ч’яЉ∞ќƒ’¬≥ці¶£ђ»з–и‘ƒґЅ‘≠ќƒЉ∞≤ќњЉќƒѕ„£ђњ…‘ƒґЅ‘≠‘”÷Њ
