
ժҪ��������Ӧ�������������Լ�����أ�ͬʱ����ʳƷ��ܵ�����֮һ���ܹ�����ʳƷӪ����ֵ�����̻����ڡ������������п��������Ե��İ�������ɡ�����˳�ռ乹���ѱ��㷺��������������о��IJ������룬�ĵĹ�Ч��ϵ����Ϊ�����俹�������Ƶĺ������⡣��ǰ�о��У���½�ض����з���Ŀ������ģ��俹�������û����ѻ���������Ȼ������Ե����ʺ����ĸ��ߵ�������Դ���俹�����ĵĹ�Ч��ϵ�о����١���ˣ����Ļ��ڿ������ĵ���ת��������ɻ�����ԭ��ת��������ɻ������ϴ������������ӵ����û��ƣ�������Դ�������ĵ�һ���ṹ�Ͷ����ṹ�Ի��Բ�������ҪӰ������������ص���������ᡢ�����就���ᡢ������ˮ������Ͳ����������ᡢ��������ͼ�������Կ��������Ե�Ӱ�죬����ָ�������������ʳƷ���з��������չ�������IJ�Ʒ�������г���
��������reactive oxygen species��ROS��������������³´�л�ͺ�����������Ȼ����[1]�������ɻ���������������û�б���ȫ���ʱ�����ǿ��Թ��������ķ��ӣ�ʹ��ʧȥ���ӣ�ȱ�ٵ��ӵķ��ӻṥ���������ӣ������������[2]��Ŀǰ����Ȼ��ϳɿ�������[3-5]������Ϊ��Դ�Կ�������ά�����ڻ���֬��������ԭƽ�⡢Ԥ�������Ƽ����������DZ�ø��������ṹ�����ȶ����������Σ�յ����߷�Ӧ���Ӷ���������[6]��
������Ȼ��Դɸѡ�ͻ�þ�����ȷ�ṹ�Ŀ�����Ч�õ��IJ�Ʒ���ڳ�ΪʳƷ��ҽ����ҵ�Ķ�����2021���ҹ������ܲ�������3561��֣�ռˮ��Ʒ�ܲ�����1/2����[7]������ӹ������в�����Լռ����50%~70%�ĸ�����[8]��������ͷ��Ƥ������������Ƭ�����������[9]���㵰����һ�ּ��ѵĿ���������Դ�����塢��������ԭ�ϵ�����ȣ����൰����Arg��Lys��Gly������IJ�ͬ��λ�Ʊ���ˮ��Һ������Ϊ���谱�����������Դ���������ױ���������[10]����ˣ�������������Դ��ȣ��㵰���в�ͬ�İ�����������ˮƽ[10]����Ϊ�Ʊ�����ͬ�ṹ�Ŀ������Ĵ�������Ŀ����ԡ�
����������Կ��������Ʊ������봿���ͼ������з�����о��϶࣬��ǰ�о���Ա��Ҫͨ��ø���[11]���������ҿ����볬����[12]����[13]����ѹ����糡[14]�ͳ��ٽ�ˮ��ȡ[14]��Ԥ����������ϣ��Ի�ø��ߵIJ�����Ͷ����͵ijɱ�����ǰ���ڿ������Ļ���λ�㡢�ṹ�����빦�ܷ�����һ������ʾ���������乹Ч��ϵ�����û����Դ���̽���Ρ�һֱ���������������ͺ����б���Ϊ�Ǿ����ĵĿ��������ܵ���Ҫ����[15]���������д��ڵ�20�ְ�������������ɻ���Ӧ����1�����˽���������������ĵ����û��ƣ�˵���˰�����Ľṹ�����뿹���������Ĺ�ϵ�����⣬�������ĵ�����úͶ����ṹӰ�쿹�����Ļ��Եĸߵͣ�Эͬ��������ǿ���忹�����ĵĻ��ԣ������û�������忹�����ĵ�����[16]��������һ���ṹ�������[17]�����ǻ����֮�������ã�����Ӱ������Ϊ��������������ǿ��[16]��ˮ�⼰Ԥ�����ֶλ�ʹ����ṹ������ṹ���߷���ת���������������ĵĶ����ṹ�������ĵĻ��Գ�����ػ���ء�
��û��һ�ֵ�һ�ļ�ⷽ���ܷ�ӳ�����������е����û��ƣ����������ͨ��������ɻ�������֬�����������Ͻ������ӡ����������������ø�ı����[18]�������������У��ĵ�������Ź�������ROS��Ӧ�������ɰ������ṩ��ԭ�ӣ���ʹ��ԭ�ӵ�λ�úͷ�ֵ�����仯������������ɻ�[11,19-20]��ͼ1��ӳ�˸���������Ŀ��������ƣ����������ˮ������Ŀ��������ƶ�����a��b��d�������就����Ŀ��������ư���a��b��������������Ŀ��������ư���a�������������a��b��c�������������a��c�����������˹��������༰�丱���ﵰ�������ĵĽṹ�����ɻ��ͽ������ӵ�Ӱ�죬�ص������ܽ��˿������ĵĹؼ�����λ������Ŷ������û��Ƶ�Ӱ�죬���ڶ�����ϳɸ��Կ�������ָ���µķ�չ�������߸��������ü�ֵ��
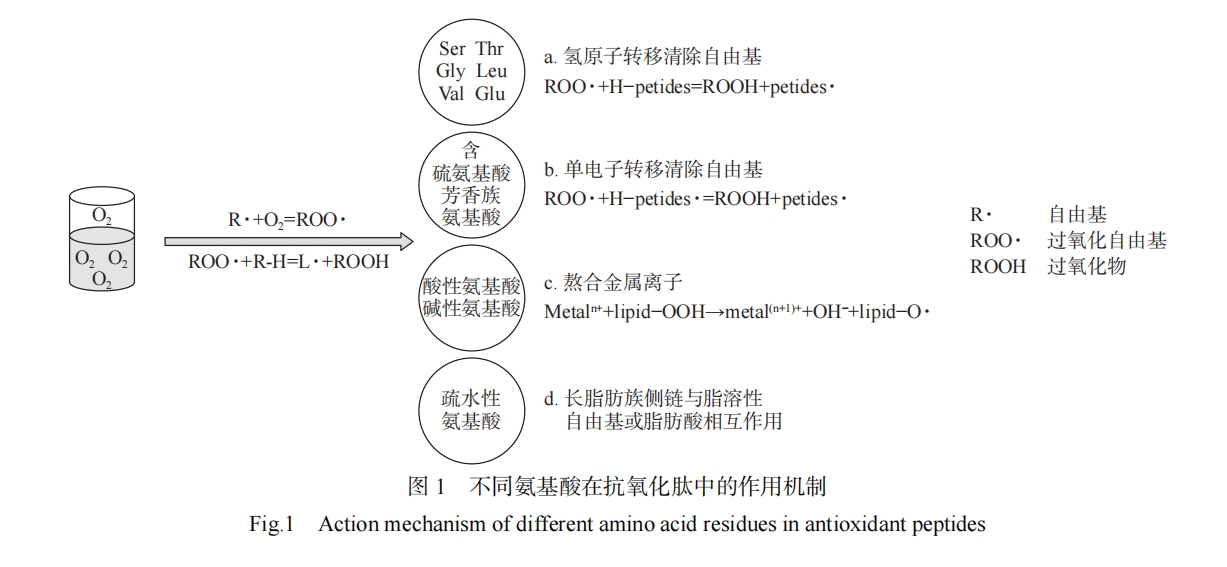
1 һ���ṹ��������ɻ������Ĺ�ϵ
1.1 ������
Met��Cys�Dz���ӵ���ϻ���sulfhydryl group��-SH���İ����ᣬ-SH�ı�¶������ǿ��������ɻ�������[21]��Met��һ�ָ߶���ˮ�İ����ᣬ�俹�������ƼȰ���ͼ1�е�b��d;�������㵰����GGGAGMLLK��Gly-Gly-Gly-Ala-Gly-Met-Leu-Leu-Lys�������е�Met�ṹ��Sԭ�ӿ����ṩ���ӴӶ���������Met����[22]��Met����й���ṹ����ά�ֿ��������ȶ���[22]�������㼡�⵰����LMGQW��Leu-Met-Gly-Gln-Trp���������ɻ�����������Oxygen radi-cal absorbance ability��ORAC��������GSH�������е�Met�ǿ������ľ߱����ԵĹؼ��ṹ[23]��Cys�Ŀ��������ư���a��b;�����������DCGY��Asp-Cys-Gly-Tyr����NYDEY��Asn-Tyr-Asp-Glu-Tyr�����ǻ����ɻ�������ԣ�hydroxy radical scavenging activity��HRSA����IC50�ֱ�Ϊ27.6��38.4 ��g/mL����DCGY�����е�Cys������-SH�ṩ��ԭ�������ɻ�ֱ�����ã�ʹ��DCGY���и�С��IC50ֵ[24]�������㼡�⵰���м����õ���HRDRLCVVQ��His-Arg-Asp-Arg-Leu-Cys-Val-Val-Gln����Cys���е�-SH���ṩ��Ծ����ԭ�������ɻ�ֱ�ӷ�Ӧ��Cys����й���ṹ����ά�ֿ��������ȶ���[22]�����⣬����CQV��Cys-Gln-Val����QCV��Gln-Cys-Val����QVC��Gln-Val-Cys����QCA��Gln-Cys-Ala��������ǻ����ɻ���ʵ���У�Cys�Ŀ�����������ͨ��-SH���Ķ�����ɵģ�������-SHʧȥ���Ӻ���ԭ�Ӻ����������ɻ�[25]����Բ�����⵰����HDHPVC��His-Asp-His-Pro-Val-Cys����HEKVC��His-Glu-Lys-Val-Cys����DPPH���ɻ����������DPPH radical scavenging activity��DRSA���ͻ�ԭ��������Ȼ��������VC��GSH���ĵĿ�������������������ˮ������Cys��His�й�[26]��
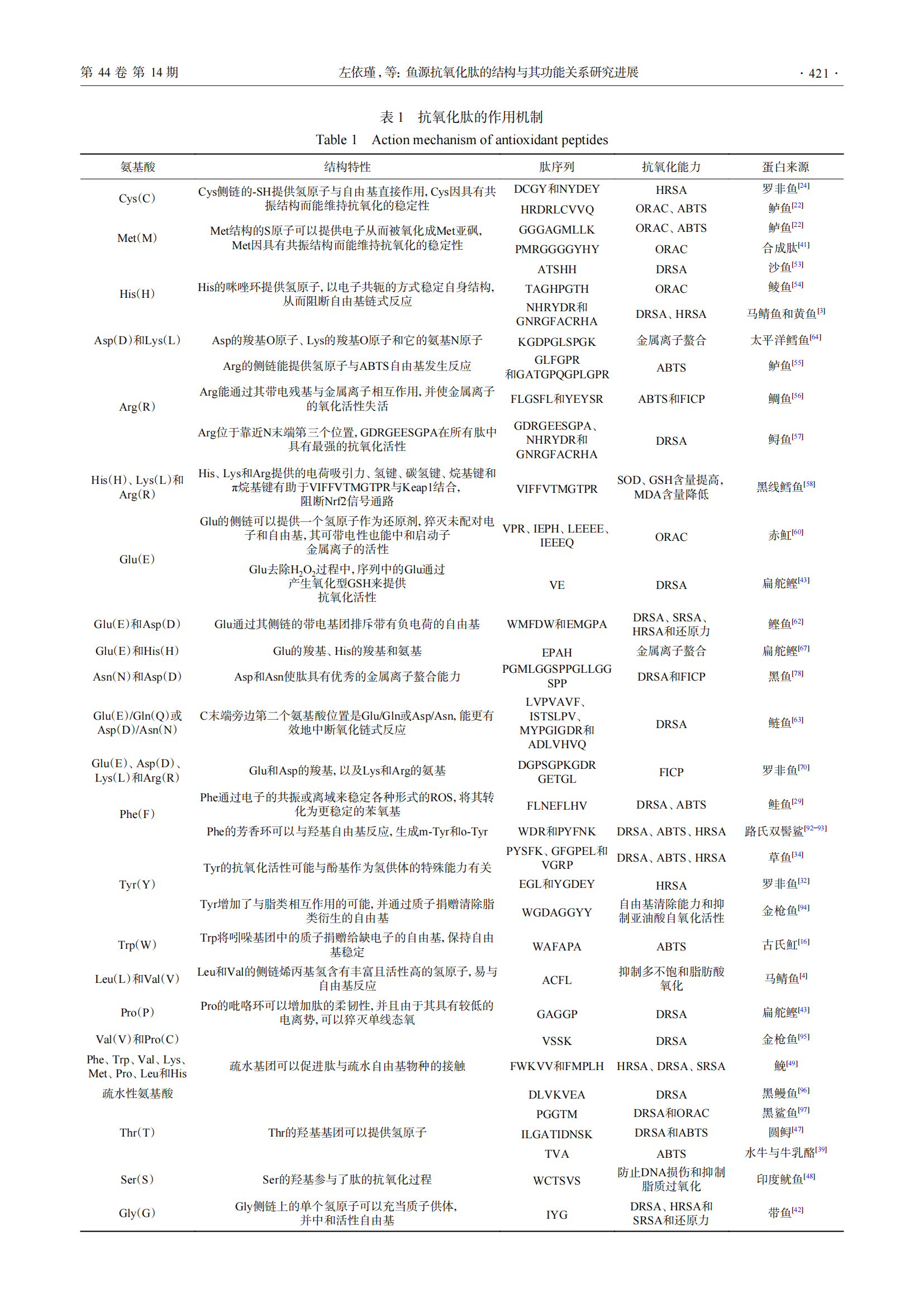
1.2 �����就����
Phe��Tyr��Trp��ͨ���䷼�㻷�����ǻ���������Ÿ����Ŀ��������ԣ����ǵ����û�����ͼ1�е�b;�������õ�����ת��ȥ�����ɻ�[25]����Tyr�����û��ƻ�����a;����Phe�Ľṹ�к��з��㻷�ͷ��ǻ���������ͨ�����ӵĹ�����������ȶ�������ʽ��ROS������ת��Ϊ���ȶ��ı��������������ɻ��鵼�Ĺ�������ʽ��Ӧ�Ĵ���[27-28]����������������FLNEFLHV��Phe-Leu-Asn-Glu-Phe-Leu-His-Val�����нϺõ�DRSA��ABTS��ABTS radicals scavenging activity��ABTS�����ɻ�������Ժͻ�ԭ�����������е�Phe��Ϊ����������λ��ʹ�IJ���������Ч��[29]��Tyr�ڽṹ����p-�㶹�����ƣ�p-�㶹����һ�ַ��㻷�ϴ���һ���ǻ��Ҿ���ǿ���������Եķ������[30]��������Tyr�Ŀ���������������ǻ�����Ϊ���������ɻ�������TyrҲ��Ŀ����ˮ�����к�ǿ����[31]����������Ƥ��YGDEY��Tyr-Gly-Asp-Glu-Tyr�����н�ǿHRSA���ԣ������еĻ���λ������ΪC�˺�N�˴��ڵ�Tyr[32]��������Ƥ����������ȡ�õ�����AY��Ala-Tyr�����нϺõ�������������������ferrous ion chelating capacity��FICP������ԭ����DRSA�������������������ǿ�ȶ��ԣ�C��Tyr�Ĵ�����ǿ�����ɻ����������[33]������Ƥ��������PYSFK��Pro-Tyr-Ser-Phe-Lys����GFGPEL��Gly-Phe-Gly-Pro-Glu-Leu����VGRP��Val-Gly-Arg-Pro��������PYSFK������ǿ��DRSA��ABTS���ɻ������������鹦�������еķ����就����Tyr�ķ��ǻ��������ã����������в��������就�����VGRP�Ŀ�������������[34]��Trp��һ�ֲ�����������ŵķǼ��Է����就����[35]��������ȹ������ɻ����ȶ�����������������κ��淴Ӧ�����ɻ��鵼�Ĺ���������Ӧ�Ĵ��������ܵ�����[36]��Trp����������е����Ӿ�����ȱ���ӵ����ɻ����Ӷ��������ɻ��ȶ�������ĵ����ɻ��������[36]����������WAFAPA��Trp-Ala-Phe-Ala-Pro-Ala����GSH�и�ǿ�Ŀ��������ԣ�����Trp��������Ҫ����[16]��
1.3 ������ˮ������
��ˮ���������Leu��Val��Gly��Pro��Met��Phe��Trp��Ala��Ile����ˮ��������������������ˮ�ͽ����е��ܽ��[16]�����Ӷ�֬�������ɻ�������֬����Ŀɼ��Ժ������[37-38]���ٽ�������֬�����������ɻ��ĵ����̣������������˵IJ���[36]����ͼ1�����û���d��ʾ������Leu��Ala���ĵĿ������������������ij�֬�����[35]���ܹ�����֬�������������������ã�������������ACFL��Ala-Cys-Phe-Leu������Leu��Ala�������ƶ����֬���������������Ȼ����������-������[4]�����⣬Leu��Val�IJ���ϩ�����⺬�зḻ�һ��Ըߵ���ԭ�ӣ����������ɻ���Ӧ[39]����ˣ�Leu��Val����ͼ1��a��b���������ơ������������QRPPEPR��Gln-Arg-Pro-Pro-Glu-Pro-Arg����EKVWKYCD��Glu-Lys-Val-Trp-Lys-Tyr-Cys-Asp����VGLPGLSGPVG��Val-Gly-Leu-Pro-Gly-Leu-Ser-Gly-Pro-Val-Gly�������ǵ�DRSA����ʷֱ�Ϊ55.1%��55.1%��59.1%���������к�������Val��VGLPGLSGPVG��DPPH���ɻ������Ч�����[40]��
Gly��Proͨ��������ṹ�����Ŀ��������ԣ�����Gly��Pro��Ӱ���ĵĶ�������[40-41]������Ӱ�칦�ܻ��ԡ�Gly�Ŀ�����������ͼ1�е�a;������������Ϊ�����壬ͨ��ת����ԭ��������ɻ���Gly������һ����ԭ����ɣ�Ϊ�ĵ������ṩ�߶�����ԣ�����ı��ĵĿռ�ṹ��Wu��[41]�������źϳ�������PMRGGGGYHY��Pro-Met-Arg-Gly-Gly-Gly-Gly-Tyr-His-Tyr����Gly�ļ��٣��ļ��γɵķ��������������Խ�٣��������ĸ���Ļ���λ�㱩¶������ORAC����ǿ�����㼡�⵰����IYG��Ile-Tyr-Gly��֮���Ծ��нϸߵ����ɻ������������ԭ������������ģ����ϵ�е�����֬�ʹ���������������ΪGly�����ϵĵ�����ԭ�ӿ��Գ䵱���ӹ��壬�кͻ������ɻ��������������̵����Ƹ��������ĵĻ���λ��Ӵ����е㣬�������ķ����������[42]��Pro�����յ������ṹ���������Ӱ����ᷢ�����õĿ�����[40]��Pro�����û�����ͼ1�е�b;�������õ�����ת��ȥ�����ɻ�������䵰����GAGGP��Gly-Ala-Gly-Gly-Pro�����нϺõ�DRSA����[43]�������е�Pro��һ���������ӵ�������ͪ����������ϵ͵ĵ����ƣ�������������������ԣ�Ҳ���Դ�����̬��[43-44]����·��˫�������ǵ�������ȡ�õ�����Pro����GPE��Gly-Pro-Glu����������ģ����ϵ�б��ֳ������ڿ�������BHT��֬�ʹ��������ƻ��ԣ��䷼������ڿ�������Ӧ�г䵱���Ӹ���[45]��
1.4 ������������
��ͼ1��a;����ʾ��Asn��Gln��Thr��Ser������Ϊ��壬ͨ���ṩ��ԭ���谭��������Ӧ��������pH�����£�Asn��Gln�����ṩ��ԭ�ӣ�����������������ɵ����ɻ��������ĵĿ���������[20]��Thr��Ser��������Ϊ������к����ɻ���������Ƥ�м����õ��ĺ���Thr������PGGTM��Pro-Gly-Gly-Thr-Met������������ǿDRSA��ORAC[46]����Բ�൰���м����õ����������У���������Thrʹ��ILGATIDNSK��Ile-Leu-Gly-Ala-Thr-Ile-Asp-Asn-Ser-Lys��������ǿ��DRSA��ABTS���ɻ��������[47]��ӡ���������WCTSVS��Trp-Cys-Thr-Ser-Val-Ser��Ser���ǻ������˸��ĵĿ��������ԣ���ֹDNA���˺�����֬�ʹ�����[48]�����ң�һͬ������Ĺ���SVNVPLY��Ser-Val-Asn-Val-Pro-Leu-Tyr���������仹ԭ����������YRIVPL��Tyr-Arg-Ile-Val-Pro-Leu����ԭ����ǰ�ߵ�N�˴���Ser[30]��
1.5 ��������
His��Lys��Arg�ǿ������������г����ļ������ᡣHis�����û��ư���ͼ1�е�a��b��c;��[25]��His��������ź�Arg�IJ������ṩ��ԭ�������ɻ���Ӧ��ʹ�ķ��ӿ��������á�His����������ǿ������ĵ���Ҫ����λ��[28]����Ϊ��������Ϊ����֬�ʹ������ɻ�����[49]��ͬʱHis����ͨ�������ӽ����ɻ�ת��Ϊ�ȶ����ӣ�ͬʱͨ������ṹ�����������ȶ���[50]���������������������������ͪ��������ͪҲ���п���������[51]��Chen��[52]���֣�����D-Hisȡ����������PHH��Pro-His-His���е�L-His�ᵼ��������½����Ʋ���������λ�á�������乩���������йء�ɳ�㼡�⵰����ATSHH��Ala-Thr-Ser-His-His�������к�������His����1 mg/mL��Ũ���£����Ķ�DPPH���ɻ�����������ߴ�90%���ϣ��������His������ŵĹ���������[53]������Ƥ������TAGHPGTH��Thr-Ala-Gly-His-Pro-Gly-Thr-His������ORAC���ԣ�������His�������ṩ��ԭ�ӣ��Ե��ӹ���ķ�ʽ�ȶ������ṹ���Ӷ�������ɻ���ʽ��Ӧ[54]��Lys�Ŀ�����������ͼ1�е�c;����ʾ��������ɵļ�����������෴��ɵĽ������ӽ�ϣ��谭������ʽ��Ӧ�Ľ��С�Arg�Ŀ��������ư���ͼ1�е�a��c;����Arg�IJ������ṩ��ԭ����ABTS���ɻ�������Ӧ��������Ƥ��GLFGPR��Gly-Leu-Phe-Gly-Pro-Arg����GATGPQGPLGPR��Gly-Ala-Thr-Gly-Pro-Gln-Gly-Pro-Leu-Gly-Pro-Arg�����н�ǿ�Ŀ����������������ĵ�Cĩ�˰�����Arg�����Ŀ���������[55]������������FLGSFL��Phe-Leu-Gly-Ser-Phe-Leu����YEYSR��Tyr-Glu-Tyr-Ser-Arg�����н�ǿ��ABTS���ɻ����������FICP��������Ϊ�����е�Arg��ͨ�������л��������������ã���ʹ��������ʧȥ��������[56]������Ƥ������GDRGEESGPA��Gly-Asp-Arg-Gly-Glu-Glu-Ser-Gly-Pro-Ala���Ǽ����л�õ��������о�����ǿ��DRSA���Ե���[57]��Arg���������еĴ��ں�λ�þ������ĵĿ��������ԣ���Argλ�ڿ���Nĩ�˵ĵ�����λ��ʱ���нϺÿ���������[3]���������ľ��нϵ�DRSA���Ե�ԭ����������ȱ��Arg������ͼ1�еĿ����������⣬His��Lys��ArgҲ����ϸ���ڿ���������������ɯ��[58]���÷��ӶԽӷ��ֺ�������Ƥ�м�������VIFFVTMGTPR��Val-Ile-Phe-Phe-Val-Thr-Met-Gly-Thr-Pro-Arg����Keap1��������ȶ��������е�His��Lys��Arg���ṩ�ĵ���������������̼�����������ͦ�����������ڶ��߽��ܽ�ϣ���Ϻ�ת¼���ӣ�NF-E2-related factor 2,Nrf2���ź�ͨ·��ʵ�ֶ��˱�Ƥ�����γ�ϸ����HaCaT���Ŀ���������ЧӦ��
1.6 ��������
�����������Glu��Asp���俹����������ͼ1��a��c;����ʾ��Glu��Asp����������ɻ����������Ͻ������ӣ������������ӵĴ��������ã��ֿ�֬������[20]��Glu�IJ����������ṩһ����ԭ����Ϊ��ԭ����������ɻ�[59]��������ǵ�����VPR��Val-Pro-Arg����IEPH��Ile-Glu-Pro-His����LEEEE��Leu-Glu-Glu-Glu-Glu����IEEEQ��Ile-Glu-Glu-Glu-Gln������ǿ���ɻ���������������е�Glu���ĵ���Ҫ����λ��[60]��������ˮ��Һ��ORACֵ����ݮ��ƻ���ͳ��ӵ�ˮ����ORACֵ���ƣ����м����Ķ�������ж�����IEE��Ile-Glu-Glu����IEEE��Ile-Glu-Glu-Glu�����У�IEE�ظ������Ǹ߿��������ԵIJ���ԭ��[61]������䵰����VE��Val-Glu������ȥ��H2O2�����У������е�Gluͨ���������������ģ�glutathione��GSH�����ṩ����������[43]��������ͷ������WMFDW��Trp-Met-Phe-Asp-Trp����EMGPA��Glu-Met-Gly-Pro-Ala���������е�Asp��Glu������ǻ����ɻ��Ĺ��������Źؼ�����[62]�����㼡�⵰����MYPGIGDR��Met-Tyr-Pro-Gly-Ile-Gly-Asp-Arg����ADLVHVQ��Ala-Asp-Leu-Val-His-Val-Gln�������н�ǿDRSA��Cĩ���Աߵڶ���������λ����Glu��Asp�ܸ���Ч�ض��ж�������ʽ��Ӧ�������к�ROS�IJ�������������ǿ�ĵ����ɻ�������Ժͻ�ԭ����[63]��
2 һ���ṹ������������������Ĺ�ϵ
���������������Ca2+��Fe2+��Zn2+�Ƚ�������ͨ����λ���۽�ϻ����������[3,64-66]����������������������Ȼ������γ����Ӽ������������İ��������γ���λ��[60]��Zhang��[64]��ˮ��̫ƽ��������еõ���Ca2+��ϵ�KGDPGLSPGK��Lys-Gly-Asp-Pro-Gly-Leu-Ser-Pro-Gly-Lys����ͨ�����ӶԽӼ��������������õ������Խ��λ����Asp-3���Ȼ�Oԭ�ӡ�Lys-10���Ȼ�Oԭ�ӡ�Lys-10�İ���Nԭ�ӡ�Chen��[67]�Ѵӱ���䵰����EPAH��Glu-Pro-Ala-His������Ca2+����λ����Glu���Ȼ���His���Ȼ��Ͱ�������ͼ2��ʾ��WALTERS�ȷֱ��С���̫ƽ�������еõ�������FVDVT��Phe-Val-Asp-Val-Thr����ʮ����AGPHGPPGKDGR��Ala-Gly-Pro-His-Gly-Pro-Pro-Gly-Lys-Asp-Gly-Arg�����������ϵ�N�˵İ�����C�˵����ᡢ�ļ���C=O���ź�His���ǰ����Ų���������������ӵ�����[68]����ǰ���������������������ĵĽ��λ����Ҫ����ˮ�����ᣨArg��Asn��Asp��Cys��Gln��Glu��His��Lys��Ser��Thr��Tyr���й�[69]��Lin��[70]������Ƥ�ȵ���øˮ��Һ�з������4����GPAGPAGEK��Gly-Pro-Ala-Gly-Pro-Ala-Gly-Glu-Lys����DGPSGPKGDR ��Asp-Gly-Pro-Ser-Gly-Pro-Lys-Gly-Asp-Arg����GLPGPSGEEGKR��Gly-Leu-Pro-Gly-Pro-Ser-Gly-Glu-Glu-Gly-Lys-Arg����DGPSGPKGDRGETGL��Asp-Gly-Pro-Ser-Gly-Pro-Lys-Gly-Asp-Arg-Gly-Glu-Thr-Gly-Leu��������FICP��Fe2+��Ҫ����������Glu��Asp���Ȼ����Լ�Lys��Arg�İ�����ϡ�Sampath��[3]��������ͻ����зֱ�����õ���NHRYDR��Asn-His-Arg-Tyr-Asp-Arg����GNRGFACRHA��Gly-Asn-Arg-Gly-Phe-Ala-Cys-Arg-His-Ala������ͬ��Ȼ��������EDTA�����൱��FICP������NHRYDR����������ϵ�еĿ���������������Ȼ��������a-�����ӣ������ĵĿ�����������Ҫ����������His�Ĺ��ס�Mendis��[71]�Ӿ��������м�������FDSGPAGVL��Phe-Asp-Ser-Gly-Pro-Ala-Gly-Val-Leu����NEPLQAGQPGQPGR��Asn-Gly-Pro-Leu-Gln-Ala-Gly-Gln-Pro-Gly-Glu-Arg��û�н��������������������ƺ�����������ȱ��His��Ե�ʡ�����Ե�[65]������������ֺ���Ƥ������Zn2+�Ľ��λ����C=O��N-C-O�����⣬λ��N�˵�Pro�����ĵĶ����ṹ���ص�����ǿ����Ϊ�����������ϼ�������[72]�����Ҹ���Pro������Ϊ���пռ�λ�裬�ֿܵ���������ǿ����Ϊ�����������ϼ����ȶ���[73]��
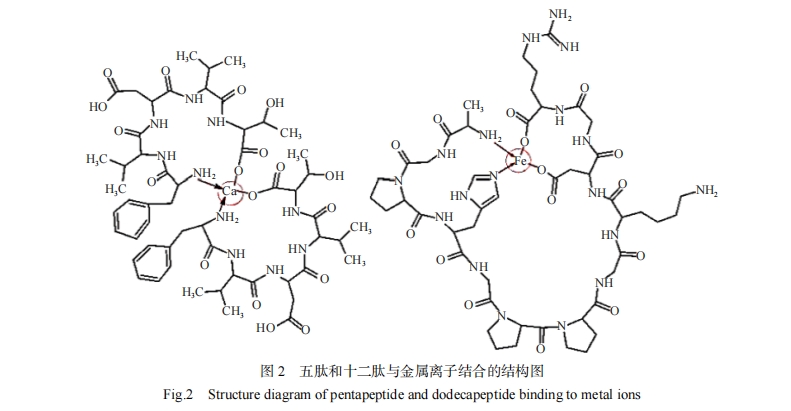
����������Ļ���������������ϻ����йأ�������ļ�����������ϵ��Ȼ�ʧȥ��ԭ������������γ������[60]���������������������ϵİ���ʧȥ��ԭ�ӣ���ԭ��δ�����ĵ��Ӷ���������ӽ�ϣ�������������Fe2+�Ļ�ԭ��ʽ�뿹��Ѫ������[61]�����ɾ������ݽṹ���ȶ���������[73]������������Ȼ���Asp�������-�Ȼ���Glu�������-�Ȼ���������ӽ��[73]��His������Cys���ϻ���Trp���������Thr���ǻ�[1]�Լ�Ser���ǻ�[74]�����ṩ����Ľ������ӽ��λ�㡣
3 �����ṹ�뿹�������ԵĹ�ϵ
�ĵĶ����ṹ������-��������-�۵�����-ת�ǡ����������Դ�ڦ�-���Ķ���WYSLAMAASDI��Trp-Tyr-Ser-Leu-Ala-Met-Ala-Ala-Ser-Asp-Ile����ORAC���Աȿ�������BHAǿ���������Ӧ�ĵ�Ħ�����백����Ļ������и����ԣ�������YVEEL��Tyr-Val-Glu-Glu-Leu��ȴ�����෴���䰱���������ORAC�ȹ��ĵ�2��[75]����˵���ļ����������Ŀռ�ṹ�ȿ��ܽ���Ҳ������߶��ĵĿ��������ԡ�
3.1 ��-�����ṹ
��-������Ҫͨ���ʻ�����-CO���Ͱ����⣨NH-�����ֻ���֮��ķ���������γɣ�������Խ����ĸ������γɦ�-�����ṹ[28]����-�����ṹ������нϸߵ������ԣ����������ڽ��淢�ӿ���������[76]����-�������ĵĿ����������з����Źؼ����ã�����Ƥˮ��Һ���нϺõ����ɻ�������������ú��������������ṹ�����Ц�-��������ռ����ߣ����������ռ����ͣ�ˮ��Һ��ABTS��HRSA�ͳ������ɻ�������ԣ�superoxide anion radical scavenging capacity��SRSA���ֱ�Ϊ1.86��1.65��2.72 mg/mL[77]��Ҳ���о�������ֳ���-��������Խ�ͣ��ľ��еĿ�����������ǿ�������չ��ײⶨ����������ˮ����Ķ����ṹ����-����ռ�����Ϊ47.88%�������ռ����С��Ϊ10.3%����200 W������������-��������������0.78%�����������������12.33%��ø������DRSA��IC50ֵ���ͣ�˵����������������[12]��
3.2 ��-�۵��ͦ�-ת�ǽṹ
��-�۵�ͨ��������֮��ά���ȶ��Ե���������γɣ�����ṹ��-�����ͦ�-�۵���Ҫλ�ڵ������ڲ�λ�ã���-ת�ǽṹ��������ṹ����[12]���ĵĿ�������������ά�ṹ������-ת�ǵĹ����йأ���������Һ�з�������ĸ��ľ��еĦ�-ת�ǽṹ��ռ�ܶ��Ķ����ṹ��75%�����м���������PGMLGGSPPGLLGGSPP��DRSA������GSH�൱����SDGSNIHFPN��FICP������EDTA�൱[78]����������Ƥ��������Ƥ�ĵĶ����ṹ��Ҫ�Ԧ�-�۵�Ϊ�����������پ�����øˮ���-�۵���������������ͣ���-�����ͦ�-ת�Ǻ������ӣ�����������Ƥ�ĵ�DRSA�������7.89%���ӵ�28.77%������������Ƥ�ĵ�DRSA����ʼ�������������Ƥ������������-���������������������DRSA����ʺ��ܻ�ԭ��������[79]�����⣬Yang��[80]��Ϊ�ϸߵĦ�-�۵������ͽϵ͵Ħ�-�����������ĵĿ�����������һ��������ԡ�
3.3 ����Ȧ�ṹ
����Ȧ��δ�۵�������������������������й�[12]����������Ȧ�ĺ���������ṹ�Ĺ�ϵ�������Ȼȹ�ϵ�������ߵ��������ʹ�ĵĽṹ������ɢ����¶����Ļ���λ�㣬�����ں����ɻ����[81]������ṹ������ṹת����ʹ�Ŀ�����������ǿ[82]���������������ȡ�ĵ;��ķ۾��н�ǿABTS�ͻ�ԭ����ʹ���������Բ��ɫ����������ṹ����-������ƽ��ʽ��-�۵���������ṹռ����С�����������ռ���������ѹ����糡����������ʵ�����Ac-QWFCT��Ac��ʾN��������Gln����������Ȧ�����뿹�������Գ��෴����[83]�����ĵ����������Ϊ32.5%ʱ���������ѵĿ��������ԡ���Ө��[84]���ú�����ס��˴Ź�����ס�Zeta��λ��Բ��ɫ������ѹ����糡�����Ĺ���KWFH��Lys-Trp-Phe-His��������KWFH�������ʻ���C=O�ͱ����������ӷ����仯����λ�ı�ʹ�����ṹ��ø�������DRSA����ʽ�PEF����ǰ��ǿ��13.92%��
4 һ���ṹ���ĵ������
4.1 ������֮��������
����������е����ӻ�ȱʧ������ǿ�ĵĿ��������ԣ�Ҳ���������ĵĿ��������ԣ����뿹�����ĵķ��������ļ�������й�[85]��Gopinatth��[17]�ӽ�ǹ��Ǹɵ�������ȡ����APTBP����������һ���滻Trpʹ�ĵ�ABTS�������63.1%�����ĵ����˶��滻Trp��ABTS���Խ���10.3%���������а�����֮���ЭͬЧӦ���俹���������з�����Ҫ���á�ͨ�����ɻ�ת�Ʒ�Ӧ���������İ�������Խ����ɻ����˽������ڽ��İ����ᣬʹ��Ӧ������ȶ�[86]��Liu��[87]��Ϊ������Tyr�ķ��ǻ��Ļ���Ӱ������Ŀ��������ܣ���Tyr����DD��Asp-Asp����DQ��Gln-Asp��֮��ʱ���������Ի����ӣ���ΪDD��DQ�еİ����������������ŵ����������˷��ǻ������������ܶȣ�����ǿ�˹�Ӧ���ͷ������ӵ�������ͬ����Bamdad��[86]��Ϊ����QPYPQ�е�P��Y�Ľ������ж��ȶ����ɻ��л������á�Najafian��[88]�������㣨L. plantarum��ˮ��Һ�м�������֮����ͬ�ṹ����PYP��Pro-Tyr-Pro������AIPPHPYP��Ala-Ile-Pro-Pro-His-Pro-Tyr-Pro�������ĵ�DRSA��ABTS�ͻ�ԭ����IC50�ֱ�Ϊ1.38��0.873��0.456 mg/mL����ˣ��������еİ�������ڵ�����ö��ĵĻ�����һ��Ӱ�죬��Ӱ��̶Ȼ��д���һ���о���
4.2 ��֮��������
�����������ĵĿ��������Բ�ͬ����������WAFAPA��Trp-Ala-Phe-Ala-Pro-Ala����MYPGLA��Met-Tyr-Pro-Gly-Leu-Ala�����ABTS���ɻ���ʵ��ֵ���ڼ��������ֵ��������֮��DZ�ڵ�Эͬ�����ܰ��������ڹ�����ʳƷ�з��Ӹ��õĿ���������[16]��ͬ�����ڰ���������NKVKGELD��EMSAGLHE��WRKKDPLND�У����ܵ�����NKVKGELD��DRSA���Ժ��������������������ķֱ���Ϻ�Ŀ�����������ǿ[89]�����������Ӷ���FLGSFLYEYSR��ABTS���ɻ������Ϊ2.448 mmol/L Trolox����һͬ���������������Ϻ����Ŀ�������������1.962 mmol/L Trolox[56]��Halim��[90]��ʵ���з��֣�10000 Da�����㵰�׳���Һ��DRSA���Գ�����3000��5000 Da����Һ�����������о���Ա��Ϊ����Һ�ķ��������俹�������Գɸ����[91]��������ĵ������˵������������ΨһӰ�����Ŀ��������Ե����ء�
5 ����
�������ĵĻ��Թ�����������ռ�ṹ��һ���ṹ�а�������š�����ԡ�����ˮ�ԡ�����úͶ����ṹ�Կ������Ļ��Բ�����ҪӰ�졣His��Arg��Lys��Glu��Asp��Ϊ���Դ��簱������������������Ե�ɵ����ɻ��ͽ������ӣ������Ŀ�������������������ϻ��������就����ı��������ǻ���������ţ�������ˮ������ij�֬�����������������������ǻ�����������ͼ���������Ȼ���Щ�����Ŷ��Կ������ĵĻ���������Ӱ�졣������˵��Gly��Asp��Glu��Leu��Val��Cys��Ser��Thr��Arg��Tyr��Glu������ɻ������û������ṩ��ԭ�ӡ�Ile��Leu��Ala��Lys��Val��Met����ˮ������Ŀ��������û�����Ҫ�����֬�����ɻ�����ǿˮ�ͽ�����ܽ�ȣ��ӻ�֬��������Met��Tyr��Trp��His��Phe��Pro��Trp�����û�����ͨ���ṩ�����ȶ����ɻ���
ø��ˮ���ƻ������༡������ﵰ�Ĺ��죬���Բ�λ��¶�����忹����������ǿ�йء����������絰��ø��ø��������ø�ⷽʽ����ø���ֶα���Ϊ�ǿ��Ƹ���ø����̵Ĺؼ������ڵ���ø��ԭ�ϵ��Ŀɼ��Ժ��и��ļ��������ԣ����»�õĿ������ĵĽṹ�в��졣���ݸ��ֵ���ø��ø��λ�㣬�����ĵĹ�Ч��ϵ����Ԥ����ʵĵ���øˮ�⵰�����ͷų�����ǿ���Կ������ĵĿ����ԣ�����������ั�������ü�ֵ�ʹ��ƿ������Ĺ������������ݡ�
��������ĵĽṹ�������������ڼ������������кͶ����ṹ����Ӧ���뿪�����ͼ����ṹ�ķ��������������俹�������ԵĹ�Ч��ϵ���ܹ����ͳɱ������Ӿ���Ч�档���⣬δ��̽���������İ��������ӡ����١��滻�����δ������Լ�̽�����������С�λ�á���ɺͲ����ǻ����̶ȶԿ��������Ե�Ӱ�죬������Ϊ�������ĵ�Ӧ���ṩ���ۻ�����ָ�����Ϳ������Ŀ����г������⣬����������Ϣѧ������ͬԴ��ģ�������ĵİ��������н��з������ܹ����õ��˽��乹Ч��ϵ���з���������������ϸ���й�Ч��ϵ����Ӧ���ߣ������ڽ�һ�������ĵĿ��������ƣ�ָ���������ĵĶ��ƻ���ơ�
��������������Ϊ��ҵ����ѧϰ����Ȩ��ԭ����ԭ��־���У�������Ȩ������ϵɾ�������±�ע���������³����������Ķ�ԭ�ļ��ο����ף����Ķ�ԭ��־
