
’™“™£ļ’ŻļŌňō¶Ńv¶¬3 «“Ľ÷÷ń‹Őō“ž–‘ ∂ĪūRGD–ÚŃ–Ķńń§ ‹ŐŚĶįį◊£¨∆š”Žļ¨RGD(Arg-Gly-Asp)ń£ŐŚĶńĶįį◊÷ ĹŠļŌĶńŐō“ž–‘‘ŕ÷◊ŃŲŌłįŻĶń’≥łĹ°Ę«®“∆°ĘĹĢ»ůľį÷◊ŃŲ—™Ļ‹–¬…ķ÷–∆ū÷ō“™◊ų”√£ģ”…”ŕ’ŻļŌňō¶Ńv¶¬3‘ŕ∂ŗ÷÷÷◊ŃŲŌłįŻĪŪ√śłŖĪŪīÔ∂Ý‘ŕ’ż≥£ŌłįŻ÷–ĶÕĪŪīÔĽÚ≤ĽĪŪīÔ£¨“Úīň∆š≥…ő™÷◊ŃŲ÷őŃ∆ĶńņŪŌŽį–Ķ„£ģ÷◊ŃŲ–¬…ķ—™Ļ‹ő™÷◊ŃŲĶń…ķ≥§ŐŠĻ©”™—Ý£¨“ÚīňĹŁńÍņī“÷÷∆÷◊ŃŲ—™Ļ‹–¬…ķ“≤≥…ő™÷◊ŃŲ÷őŃ∆Ķń÷ō“™Õĺĺ∂£ģ”–—–ĺŅŌ‘ ĺ£¨ľł÷÷RGD∂ĺňōĶįį◊≤ĽĶę“‘’ŻļŌňō¶Ńv¶¬3ő™į–Ķ„į–ŌÚĹŠļŌĶĹ÷◊ŃŲ≤ŅőĽī”∂ÝĺŖ”–÷ĪĹ”ŅĻ÷◊ŃŲŌłįŻ‘Ų÷≥°Ęū§łĹ°Ę«®“∆ľįĹĢ»ůĻ¶ń‹£¨∂Ý«“ňŁ√«ĽĻĺŖ”–ŅĻ÷◊ŃŲ—™Ļ‹–¬…ķĶń◊ų”√£¨“ÚīňRGD∂ĺňōĶįį◊Ņ…ī”…Ō ŲŃĹ∑Ĺ√ś“÷÷∆÷◊ŃŲ…ķ≥§”Ž◊™“∆£ģĪĺőńĺÕ’ŻļŌňō¶Ńv¶¬3ő™į–ŌÚĶńRGDŇšŐŚĹŠĻĻŐōĶ„ľį∆š‘ŕ÷◊ŃŲ÷őŃ∆÷–Ķńį–ŌÚ÷őŃ∆ļÕŅĻ—™Ļ‹–¬…ķ”¶”√ľį«įĺįľ”“‘◊Ř Ų£ģ
’ŻļŌňō(integrins) «“Ľ÷÷“ž‘ī∂ĢĺŘŐŚ◊ť≥…Ķńń§ ‹ŐŚľ“◊Ś£¨∆š∑÷◊””…2łŲ—«Ľý¶Ń°Ę¶¬◊ť≥…£¨őĽ”ŕŌłįŻĪŪ√ś£¨ľśĺŖū§łĹļÕ–ŇļŇīęĶľĻ¶ń‹£ģ∆ýĹŮő™÷Ļ“—∑ĘŌ÷”–18÷÷≤ĽÕ¨Ķń¶Ń—«ĽýļÕ8÷÷≤ĽÕ¨Ķń¶¬—«Ľý◊ť≥…Ķń24÷÷“ž‘ī∂ĢĺŘŐŚ£Ř1£›£ģ—–ĺŅĪŪ√ų£¨’ŻļŌňō‘ŕ–¬…ķ—™Ļ‹ńŕ∆§ŌłįŻļÕ∂ŗ÷÷÷◊ŃŲŌłįŻĪŪ√śłŖĪŪīÔ£Ř2£›£¨∆š‘ŕŌłįŻū§łĹ°Ę‘Ų÷≥°Ę∑÷ĽĮ°Ę◊™“∆°ĘĶÚÕŲĶ»Ļż≥Ő÷–∆ū◊Ň÷ō“™ĶńĶųŅō◊ų”√£¨Õ¨ Ī“≤‘ŕ—™Ļ‹–¬…ķļÕ÷◊ŃŲĶń«®“∆ĹĢ»ů÷–∑ĘĽ”÷ō“™Ļ¶ń‹£ģ—–ĺŅ÷§ Ķ£¨įŁņ®¶Ń2¶¬1°Ę¶Ń1¶¬1°Ę¶Ńv¶¬3°Ę¶Ńv¶¬5‘ŕńŕ∂ŗ÷÷’ŻļŌňōĺýń‹ ∂Īū≤ĘĹŠļŌRGD(Arg-Gly-Asp)ń£ŐŚ£¨”įŌž—™Ļ‹…ķ≥…Ļż≥Ő÷–ŌłįŻĶń…ķ≥§°Ę«®“∆ļÕ’≥łĹ£ģ≤ĽÕ¨÷÷Ķń’ŻļŌňō—«Ľý ∂Īū≤ĽÕ¨÷÷ņŗĶńECMĶįį◊(»ÁFig£ģ1)£ģ
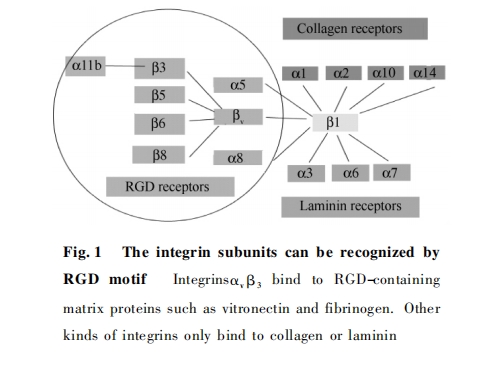
‘ŕ–¬…ķ—™Ļ‹ńŕ∆§ŌłįŻļÕ÷◊ŃŲŌłįŻĪŪ√ś£¨◊Óő™Ļ„∑ļłŖĪŪīÔĶń’ŻļŌňō «¶Ńv¶¬3£ģ¶Ńv¶¬3”ŽŌłįŻÕ‚Ľý÷ (extracellularmatrix£¨ECM)ĹŠļŌĶńŇšŐŚ£¨Õ®≥£ «ECMĶįį◊…ŌĶńRGDń£ŐŚ(RGDmotif)£ģ
RGDń£ŐŚ «ń≥–©Ķįį◊÷ ∑÷◊”÷–Ķń3Žń–ÚŃ–Arg-Gly-Asp£¨ő™ŌłįŻ ∂ĪūĶń◊Ó–°ĹŠĻĻĶ•őĽ£ģŌ÷“—∑ĘŌ÷£¨RGDń£ŐŚĻ„∑ļīś‘ŕ”ŕŌłįŻÕ‚Ľý÷ Ķń≤£Ń¨Ķįį◊(vitronectin£¨VN)°ĘŌňѨĶįį◊(laminin£¨LN)ľį≤„ѨĶįį◊(fibronectin£¨FN)÷–£¨ «ŌłįŻĪŪ√ś’ŻļŌňō(integrins)”ŽECMĶńĹŠļŌőĽĶ„£ģ
≥żŌłįŻÕ‚Ľý÷ ÷–ļ¨”–RGDń£ŐŚÕ‚£¨RGDń£ŐŚ“≤ń‹ĪĽ≤°∂ĺļÕŌłĺķĶ»≤°‘≠ŐŚňýņŻ”√£Ř3-5£›£ģ’‚–©≤°‘≠ŐŚÕ®ĻżRGD”Žňř÷ųŌłįŻĶńŐō∂®’ŻļŌňōĹŠļŌ£¨ĺ≠ĻżŌłįŻÕŐ …◊ų”√ĹÝ»Žňř÷ųŌłįŻ£ģīňÕ‚£¨‘ŕ…Ŗ∂ĺ°ĘňģÚőľįÚÁņŗ∂ĺŌŔ∑÷√ŕőÔ÷ ÷–“≤∑ĘŌ÷ŃňRGDń£ŐŚĶįį◊£ģ’‚–©ņī‘ī”ŕ∂ĮőÔ∂ĺŌŔĶńRGD∑÷√ŕĶįį◊£¨Ņ…∆ĺĹŤĶįį◊÷ “Ľľ∂ĹŠĻĻ…ŌŐō”–ĶńRGD∑÷◊”–ÚŃ–£¨≥…ő™ŌłįŻÕ‚Ľý÷ ”ŽŌłįŻ’ŻļŌňōľšĹŠļŌĶń«Ņ–ßĺļ’ý–‘ř◊ŅĻľŃ£¨ĹÝ∂ÝÕ®Ļż∑‚Ī’’ŻļŌňōĶńŌłįŻ–ŇļŇ◊™ĶľÕ®¬∑∂Ý“÷÷∆—™–°įŚĺŘľĮ£¨◊Ó÷’īÔĶĹ“÷÷∆—™Ļ‹–¬…ķ£¨ŅĻ÷◊ŃŲŌłįŻ‘Ų÷≥°Ęū§łĹ°Ę«®“∆ļÕĹĢ»ůĶ»◊ų”√£¨≤Ęń‹”’Ķľ÷◊ŃŲŌłįŻ∑Ę…ķĶÚÕŲ£Ř6£›£ģń£ń‚≤°‘≠ŐŚ£¨”√»ňĻ§ļŌ≥…≤ķőÔ—–ĺŅRGD”Ž’ŻļŌňōĶńį–ŌÚŌłįŻ ∂ĪūļÕŌłįŻńŕ…„◊ų”√£¨“—≥…ő™’Ô∂Ō÷őŃ∆÷◊ŃŲ£¨ľį∆š“©őÔ—–∑ĘĶńńŅĪÍ£ģ“‘’ŻļŌňōő™į–Ķ„ĶńRGDŇšŐŚ—–ĺŅ£¨“—≥…ő™÷◊ŃŲ≤°ņŪ—ßļÕ“©ņŪ—ß—–ĺŅ»»Ķ„£ģ
”…”ŕ¶Ńv¶¬3 «ĹŌő™Ļ„∑ļĪŪīÔ”ŕ–¬…ķ—™Ļ‹ńŕ∆§ŌłįŻļÕ÷◊ŃŲŌłįŻĪŪ√ś£¨∂Ý‘ŕĺ≤÷ĻĶń—™Ļ‹ńŕ∆§ŌłįŻļÕ’ż≥£ŌłįŻ÷–ĶÕĪŪīÔĶń’ŻļŌňō£¨“—≥…ő™ŅĻ—™Ļ‹–¬…ķļÕŅĻ÷◊ŃŲ“©őÔĶń◊ų”√į–Ķ„£¨’Ž∂‘∆šĹÝ––ĶńRGDŇšŐŚ—–ĺŅ“≤»°Ķ√ŃňĹŌļ√ĶńĹÝ’Ļ£¨
1 ÷◊ŃŲ—™Ļ‹–¬…ķ”Ž◊™“∆
—™Ļ‹–¬…ķ «÷łī”‘§Ō»īś‘ŕĶń—™Ļ‹÷––ő≥…–¬—™Ļ‹ĶńĻż≥Ő£ģ—™Ļ‹–¬…ķ»°ĺŲ”ŕ—™Ļ‹ńŕ∆§ŌłįŻ…ķ≥§“Ú◊”(vascular endothelial growth factor£¨VEGF)°Ę≥…Ōňő¨ŌłįŻ…ķ≥§“Ú◊”(fibroblast growth factor£¨FGF)°ĘĪŪ∆§…ķ≥§“Ú◊”(epidermal growth factor£¨EGF)”Ž—™Ļ‹–¬…ķ“÷÷∆∑÷◊”(»Á—™Ļ‹“÷ňō°Ęńŕ∆§“÷ňōĶ»)÷ģľšĶń∆Ĺļ‚£ģ∂Ý÷◊ŃŲ…ķ≥§”Ž◊™“∆łŖ∂»“ņņĶ”ŕ÷◊ŃŲ—™Ļ©Ķń ¬ Ķ£¨ ĻĶ√÷ŕ∂ŗĶńŅ∆—ßľ“Õ∂…Ū”ŕ—į’“ŅĻ—™Ļ‹–¬…ķ∑÷◊”£¨≤Ę÷∆∂®◊Ť÷Ļ÷◊ŃŲłī∑Ę”Ž◊™“∆≤Ŗ¬‘ĶńŅ∆—ß—–ĺŅ÷–°£
÷◊ŃŲ—™Ļ‹–¬…ķ”Ž…ķņŪ–‘—™Ļ‹–¬…ķĺŖ”–√ųŌ‘≤ĽÕ¨£¨’‚įŁņ®“ž≥£Ķń—™Ļ‹ĻĻ‘ž°Ęńŕ∆§ŌłįŻ”Ž÷‹őßŌłįŻŃ¨Ĺ”ĶńłńĪš°Ę∑ī≥£Ķń—™Ńų°Ę…ÝÕł–‘Ķń‘Ųľ”°ĘŌłįŻ≥… žĶń—”≥ŔĶ»£ģ∂ŗ ż÷◊ŃŲĶń‘Ų≥§∂ľ∆ū ľ”ŕőř—™Ļ‹–°ŃŲ£¨–¬—™Ļ‹Ķń–ő≥…ő™÷◊ŃŲ◊ť÷ĮŐŠĻ©ŃňĪō“™Ķń”™—Ý”Ž—ű∆Ý£¨ Ļ÷◊ŃŲĶ√“‘ŅžňŔ…ķ≥§£ģ
÷◊ŃŲŌłįŻÕ®Ļż◊™“∆ŌÚ»ę…Ūņ©…Ę£ģ◊™“∆Ķń∑Ę…ķĺ≠ĻżľłłŲ≤Ĺ÷Ť: ◊Ō»£¨÷◊ŃŲŌłįŻ ß»•E-ł∆’≥Ķįį◊“ņņĶĶńŌłįŻľš’≥łĹ£¨ĽŮĶ√«®“∆ĪŪ–Õ≤Ęī”ŌŗŃŕŌłįŻ÷–Õ—ņŽ£¨ī©ĻżĽý÷ ń§≤Ę«÷»ŽĶĹľš÷ Ľý÷ ÷–;»Ľļů£¨÷◊ŃŲŌłįŻī©»Ž—™Ļ‹ļÕŃ‹įÕĻ‹ī”∂ÝĹÝ»Ž—≠Ľ∑ŌĶÕ≥(’‚łŲĻż≥ŐĹ–ńŕ…Ý);◊Óļů£¨÷◊ŃŲŌłįŻňś—™Ńų‘ň––£¨ū§łĹ”ŕ—™–°įŚļÕį◊—™«Ú≤Ę–ő≥…ň®◊”£¨Õ£ŃŰ”ŕį–∆ųĻŔĶńőĘ—≠Ľ∑÷–£¨ī”—™Ńų÷–Õň≥Ų(’‚łŲĻż≥ŐĹ–Õ‚…Ý)£¨≤Ę‘ŕį–∆ųĻŔ◊ť÷Į÷–∑ŤŅŮ…ķ≥§£ģ
2 ¶Ńv¶¬3‘ŕ÷◊ŃŲ∑Ę’Ļ÷–ĶńĻ¶ń‹
ń‹ĻĽĶųĹŕ—™Ļ‹–¬…ķĶń–ŇļŇ∑÷◊”įŁņ®…ķ≥§“Ú◊” ‹ŐŚ°Ęņ“įĪňŠľ§√ł ‹ŐŚ°ĘGĶįį◊Ҿљ ‹ŐŚľį’ŻļŌňō£¨∂Ý’ŻļŌňō–ŇļŇÕ®¬∑‘ŕ÷◊ŃŲ—™Ļ‹–¬…ķ”Ž÷◊ŃŲ◊™“∆÷–∑ĘĽ”ĻōľŁ◊ų”√£ģ’ŻļŌňōĪŪīÔ”ŕ—™Ļ‹ńŕ∆§ŌłįŻĪŪ√ś£¨‘ŕ—™Ļ‹–¬…ķ∆ŕľšĶųĹŕŌłįŻ«®“∆”ŽīśĽÓ;∂Ý»ÁĻŻ’ŻļŌňōĪŪīÔ”ŕ÷◊ŃŲŌłįŻĪŪ√ś£¨ĹęÕ®Ļż‘Ų«Ņ÷◊ŃŲŌłįŻī©Ļż—™Ļ‹ĪŕĶńĹĢ»ůļÕ‘ň∂Į∂ÝīŔĹÝ÷◊ŃŲ◊™“∆£ģ”––ßĶń÷◊ŃŲĹĢ»ů–Ť“™∂‘ĹĢ»ů«į—ōī¶ĶńŌłįŻÕ‚Ľý÷ ĹÝ––≤Ņ∑÷ĹĶĹ‚£¨Ľý÷ Ĺū ŰĶįį◊√ł(MMPs) «īňĻż≥Ő÷–∆ū÷ų“™◊ų”√Ķń√ł£ģ¶Ńv¶¬3’ŻļŌňōń‹”ŽŌłįŻľš÷ ≥…∑›(»Á:ŌňѨĶįį◊°Ę≤£Ń¨Ķįį◊°Ę—™–°įŚ∑ī”¶Ķįį◊)…ŌĶńRGD–ÚŃ–ŌŗĹŠļŌ£¨Ņ…‘ŕ—™Ļ‹–¬…ķ∆ŕľš∑Ę…ķŌ‘÷Ý…ŌĶų£¨∂Ý‘ŕ≥ŃľŇĶńńŕ∆§ŌłįŻ÷–»īőřīň∑ī”¶£ģ
’ŻļŌňō¶Ńv¶¬3‘ŕ÷◊ŃŲĹĢ»ů”Ž◊™“∆÷–ĺŖ”–Őō“ž◊ų”√£¨ «“Úő™∆šĺŖ”–’–ńľļÕľ§ĽÓMMP-2ļÕŌň»‹√łĶńń‹Ń¶£¨ī”∂Ý∂‘Ľý÷ ń§ļÕľš÷ Ľý÷ ≥…∑›ĹÝ––ĹĶĹ‚£ģ—–ĺŅĪŪ√ų£¨’ŻļŌňō¶Ńv¶¬3‘ŕ÷◊ŃŲŌłįŻĪŪ√śĶńĪŪīÔ”Ž∂ŗ÷÷∂Ů–‘÷◊ŃŲ(įŁņ®∑őį©°Ę»ťŌŔį©°Ę¬—≥≤į©°ĘĹļ÷ ńłŌłįŻŃŲľįļŕ…ęňōŃŲ)Ķń∑Ę’ĻĹÝ≥Ő≥ ’żŌŗĻō£ģīůŃŅĶń—–ĺŅ÷§ Ķ£¨’ŻļŌňō¶Ńv¶¬3‘ŕ÷◊ŃŲ…ķ≥…°Ę◊™“∆Ļż≥Ő÷–∑ĘĽ”÷ō“™◊ų”√£Ř7-8£›£ģ
3 ”Ž¶Ńv¶¬3Őō“žĹŠļŌĶńRGDŇšŐŚľį∆šĻ¶ń‹
3.1 ľł÷÷»ňĻ§ļŌ≥…ĶńRGDŇšŐŚĶńĹŠĻĻ”ŽĻ¶ń‹
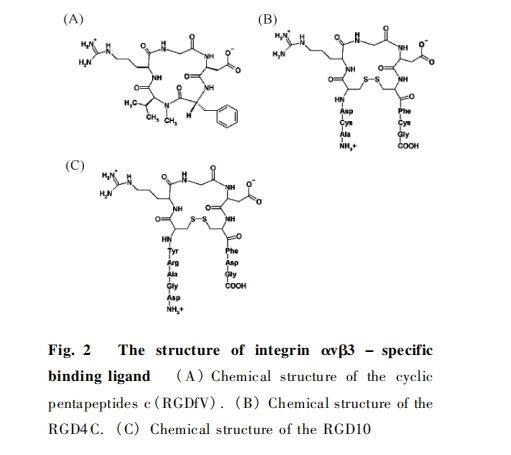
őř¬Ř «Őž»ĽĶńĽĻ «ļŌ≥…ĶńRGDŽń£¨∆š”Ž’ŻļŌňōĶń«◊ļÕ–‘ļÕŐō“ž–‘∂ľ»°ĺŲ”ŕRGDįĪĽýňŠ≤–Ľý≤ŗŃī£¨ŐōĪū «Őž∂¨įĪňŠĶń≤ŗŃī£Ř9£›£ģ≥żīň÷ģÕ‚£¨’‚–©≤–Ľý”Ž’ŻļŌňōľįRGD÷‹ĪŖĽýÕŇĶńŌŗĽ•◊ų”√“≤ń‹”įŌžŽńŃīĶń’ŘĶĢ£¨ī”∂ÝłńĪšRGD–ÚŃ–ĶńĻĻŌůŐōĶ„£ģRGDĶń»żő¨ĻĻŌů”–ņŻ”ŕRGD–ÚŃ–ÕĽ≥Ų∑÷≤ľ”ŕ“©őÔ∑÷◊”ĪŪ√ś£¨ Ļ∆š∑ĘĽ”łŁļ√Ķńį–ŌÚ◊ų”√£Ř10£›£ģīů∂ŗ żŌŖ–őRGDŽń‘ŕŐŚńŕ—≠Ľ∑ĶńįŽň•∆ŕĹŌ∂Ő£¨’‚ «“Úő™ŌŖ–őŽńŃīĶńŐž∂¨įĪňŠ≤–Ľý»›“◊ĪĽĶįį◊√łňģĹ‚£Ř11£›£¨∂ÝĽ∑ĽĮĶńĹŠĻĻ‘Úń‹ĻĽĪ‹√‚…Ō Ų«ťŅŲĶń∑Ę…ķ£¨ňý“‘Ľ∑–őRGDŽńņŗłŁľ”ő»∂®£¨«“ĺŖ”–łŁ«ŅĶń ‹ŐŚĹŠļŌ–‘ļÕ ‹ŐŚŐō“ž–‘£¨“ÚīňĽ∑ĽĮ≥…ő™ŐŠłŖRGDŽńĹŠļŌń‹Ń¶≥£”√Ķń∑Ĺ∑®£ģ
c(RGDf-N(Me)-V)”÷√ŻEMD121974ĽÚCilengitide£¨ «“Ľ÷÷Ľ∑ĽĮRGDŽń£ģCilengitideń‹‘ŕnmolŇ®∂»Ō¬ĺļ’ý–‘“÷÷∆≤£Ń¨Ķįį◊ļÕŌňő¨Ķįį◊‘≠”ŽŐō∂®’ŻļŌňōĶńĹŠļŌ£¨∆š”Ž’ŻļŌňō¶Ńv¶¬3ĶńĹŠļŌń‹Ń¶łŖ”ŕ”Ž¶ŃIIb¶¬3’ŻļŌňōĹŠļŌń‹Ń¶Ķń1000Ī∂£Ř12£›£¨’‚ňĶ√ų∆šń‹”Ž¶Ńv¶¬3Őō“žłŖ–ßĹŠļŌ£ģCilengitideĶńŌř÷∆–‘Ľ∑ĽĮĹŠĻĻļÕ≤Ś»ŽĶńDįĪĽýňŠ≤–ĽýłńĪšŃňRGD–ÚŃ–ĶńĻĻŌůŐōĶ„£¨ Ļ∆šłŁ”–ņŻ”ŕ”Ž’ŻļŌňō¶Ńv¶¬3ĶńĹŠļŌ£ģ‘ŕCilengitideĶńŌĶŃ–—‹…ķőÔ÷–£¨”…”ŕc(RGDFK)ĶńņĶįĪňŠ≤–Ľý(K) «ĹÝ––Ō¬“Ľ≤ĹĽĮ—ßĹŠļŌĶńņŪŌŽĹŠĻĻőĽĶ„£¨ňý“‘∆š≥£ĪĽ”√”ŕ“ĹŃ∆…ŌĶń“©őÔīę š£ģ
RGD4C «ŃŪ“Ľ÷÷ļ¨2łŲ∂ĢŃÚľŁĶńRGDŽń(ACDCRGDCFCG)£¨∆š∂‘÷◊ŃŲ–¬…ķ—™Ļ‹ńŕ∆§ŌłįŻĶń“÷÷∆ń‹Ń¶◊Ó«Ņ£¨∆š“÷÷∆ń‹Ń¶ «ļ¨Ķ•łŲ∂ĢŃÚľŁRGDĽ∑ŽńĶń20Ī∂£¨ «ŌŖ–őRGD∂ŗŽńĶń100Ī∂£Ř13£›£ģRGD4C“—ĪĽ◊ųő™ĽĮ—ß“©őÔĶń‘ōŐŚ£¨≥…Ļ¶”√”ŕ÷◊ŃŲ—™Ļ‹Ķńį–ŌÚ÷őŃ∆£Ř14£›£ģRGD4CŅ…◊ųő™į–ŌÚŇšŐŚĪĽ”√”ŕ‘ňňÕŌłįŻ‘Ų÷≥“÷÷∆“©őÔ£¨“≤Ņ…”√÷ō◊ťĶń∑Ĺ∑®Ĺę∆š’ŻļŌĶĹĶįį◊ĽÚ≤°∂ĺ÷–£ģĶę «£¨RGD4CĽĻĺŖ”–ĺ÷Ōř–‘£¨∆šń‹’ŘĶĢ≥Ų≤ĽÕ¨ĶńĽ∑◊īĹŠĻĻ£¨≥żŃňĶš–ÕĶń∂ĢĽ∑ĹŠĻĻ£¨ĽĻ”–∆šňŁĶńĶ•Ľ∑ļÕ∂ĢĽ∑ĹŠĻĻ£¨∂Ý∆šňŁĹŠĻĻ”Ž’ŻļŌňōĶń«◊ļÕŐō“ž–‘ĶÕ”ŕĶš–ÕĶń∂ĢĽ∑ĹŠĻĻ‘ľ10Ī∂£Ř15£›£ģ
◊ÓĹŁ£¨Õ®Ļż …ĺķŐŚĪŪ√ś’Ļ ĺŽńŅ‚ľľ ű∑ĘŌ÷Ńň“Ľ÷÷–¬ĶńĺŖ”–łŖ«◊ļÕŐō“ž–‘ĶńRGDŽń£Ř16£›£¨’‚÷÷RGDŽń√Ł√Żő™RGD10£ģRGD10ļ¨”–1łŲ∂ĢŃÚľŁ£¨Ķę∆šCRGDCĹŠĻĻ”ŽRGD4CĶńņŪĽĮ–‘÷ Ōŗň∆£¨“ÚīňŌ‘ ĺ≥Ų”ŽRGD4CŌŗÕ¨ĶńĹŠļŌń‹Ń¶£ģ
Õ®ĻżĹęRGDҚՌѨŔĶĹ≤ĽÕ¨‘ōŐŚŌĶÕ≥£¨Ņ…“‘ŐŠłŖRGDŇšŐŚĶń«◊ļÕń‹Ń¶ļÕ“©īķ∂ĮѶ—ߣģ—–ĺŅ£Ř17£›ĪŪ√ų£¨RGD∂ŗľŘĺŘļŌőÔ‘ŕnmolŇ®∂»Ō¬£¨”Ž»ň∆Íĺ≤¬Ųńŕ∆§ŌłįŻ(HUVEC)÷–Ķń¶Ńv¶¬3’ŻļŌňōĶń«◊ļÕѶĪ»Ķ•∂ņĶńRGDŐŠłŖŃň250Ī∂£ģ”√64CuĪÍľ«Ńň2÷÷RGDłīļŌőÔ°™°™°™RGDŽńňńĺŘŐŚļÕRGDŽńįňĺŘŐŚ£¨≤Ę‘ŕ’ŻļŌňō¶Ńv¶¬3łŖĪŪīÔĶńĹļ÷ ŌłįŻŃŲ∂ĮőÔń£–Õ÷–ĹÝ––PET≥…ŌŮ£ģĹŠĻŻŌ‘ ĺ£¨”ŽRGDňńĺŘŐŚŌŗĪ»£¨RGDįňĺŘŐŚ∂‘’ŻļŌňō¶Ńv¶¬3Ķń«◊ļÕѶļÕĹŠļŌŐō“ž–‘∂ľ√ųŌ‘ŐŠłŖ£Ř18£›£ģ”…īňŅ…ľŻ£¨RGD‘ŕŐŚńŕĶńő»∂®–‘ļÕ“©īķ∂ĮѶ—ßĻż≥Őń‹Õ®ĻżļŌ≥…RGD∂ŗĺŘŐŚĹÝ––ŐŠłŖ£ģīňÕ‚£¨∂ŗĺŘŐŚ≤ĽĹŲń‹īů∑ý∂»ŐŠłŖ«◊ļÕń‹Ń¶£¨ĽĻń‹īŔĹÝŌłįŻĶńńŕÕŐ◊ų”√£¨”Ž÷¨÷ ŐŚ°Ęń…√◊ŅŇŃ£°ĘĶįį◊÷ ļÕ∆šňŻĺŘļŌőÔ‘ōŐŚŌĶÕ≥ѨŔĶńRGD∂ŗľŘĺŘļŌőÔ£¨Ī»Ķ•∂ņĶńRGDŽńłŁ”–Ņ…ń‹Õ®Ļż ‹ŐŚĹťĶľĶńńŕÕŐ◊ų”√ĹÝ»ŽŌłįŻ£Ř19-20£›£ģ
3.2 RGD∂ĺňōĶįį◊ĶńŅĻ—™Ļ‹–¬…ķ◊ų”√
—™Ļ‹…ķ≥… «ĻŐŐŚŃŲ…ķ≥§ļÕ’≥łĹĶń÷ō“™Ļż≥Ő£ģľ§ĽÓ’ŻļŌ¶Ńv¶¬3Ņ…ń‹ī•∑ĘįŻńŕ–ŇļŇÕ®¬∑£¨īŔĹÝ—™Ļ‹Ķń…ķ≥§ļÕ∑÷ĽĮ£ģ“Úīň£¨“÷÷∆—™Ļ‹…ķ≥…ĪĽ»Ōő™ «“Ľ÷÷ļ‹”–«įÕĺĶń÷◊ŃŲ÷őŃ∆Õĺĺ∂£ģ
‘ŕ…Ŗ∂ĺ°ĘňģÚőľįÚÁņŗ∂ĺŌŔ∑÷√ŕőÔ÷ ÷–“—∑ĘŌ÷Ńň∂ŗ÷÷RGDń£ŐŚĶįį◊£ģ»•’ŻļŌňō «÷łņī‘ī”ŕ…Ŗ∂ĺĶńRGDń£ŐŚ∂ĺňōĶįį◊£¨ «ńŅ«įĪĽ—–ĺŅ◊Óő™Ļ„∑ļĶń“Ľ÷÷£¨∆š÷÷ņŗŅ…łýĺ›…Ŗ÷÷ņī‘ī∑÷ő™ ż ģ÷÷£ģTriflavin «ī”÷Ů“∂«ŗ…Ŗņŗ∑÷ņŽ≥ŲĶńRGD∂ĺňōĶįį◊£¨ń‹ĻĽ◊Ť÷ĻļÕ“÷÷∆»ň∆Íĺ≤¬Ųńŕ∆§ŌłįŻĶń’≥łĹļÕ«®“∆£¨≤Ę«“ Ķ—ť÷§√ų£¨∆šń‹Õ®Ļż“÷÷∆’ŻļŌňō¶Ńv¶¬3ĹÝ∂Ý”––ßĶń“÷÷∆—™Ļ‹…ķ≥…£Ř21£›£ģŃŪÕ‚£¨ī”Úů…ŖŐŚńŕ∑÷ņŽ≥ŲĶńContortrostatin£Ř22 RGD∂ĺňōĶįį◊£¨“≤ ««Ņ–ßĶń’ŻļŌňō¶Ńv¶¬3ř◊ŅĻľŃ£¨őř¬Ř «ŐŚńŕĽĻ «ŐŚÕ‚∂ľń‹«ŅѶ“÷÷∆—™Ļ‹…ķ≥…£¨“Ķ“—÷§√ų£¨”ŽdocetaxelŃ™ļŌ Ļ”√∂‘«įŃ–ŌŔį©”–ĹŌļ√Ķń“÷÷∆–ß”√,īňÕ‚£¨ĽĻ”–…Ŗ∂ĺobtustatin£¨ĹŲįŁļ¨41łŲįĪĽýňŠ∑÷◊”£¨ «“Ľ÷÷ĶÕ∑÷◊”ŃŅĶń’ŻļŌňō“÷÷∆ľŃ£ģ”…”ŕ’ŻļŌňō‘ŕ√‹ľĮ‘Ų÷≥ĶńőĘ—™Ļ‹ńŕ∆§ŌłįŻ…ŌłŖ∂»ĪŪīÔ£ģObtustainĶń’‚÷÷–‘÷ Ō‘ ĺ£¨∆š∑«≥£ ”ŕ∑Ę’Ļ≥…ő™ŅĻ—™Ļ‹…ķ≥…Õĺĺ∂ĶńŅĻ÷◊ŃŲ“©őÔ£Ř23£›£ģņī‘ī”ŕňģÚőņŗĶńDecorsin£¨ «ī”ĪĪ√ņňģÚőŐŚńŕ∑ĘŌ÷Ķńļ¨”–39łŲįĪĽýňŠĶńRGDń£ŐŚĶįį◊£¨Ornatin‘ŕňģÚőPlacobdellaornataŐŚńŕ∑ĘŌ÷£¨”ŽDecorsin”–40%ĶńÕ¨‘ī–‘£¨īŅĽĮĶń”Ž÷ō◊ťĶńOrnati ∂ľļ¨”–6łŲįŽŽ◊įĪňŠ≤–ĽýļÕ1łŲRGD–ÚŃ–£ģOrnatinń‹”––ßĶń“÷÷∆Ōňő¨Ķįį◊‘≠”Ž’ŻļŌňōĶńĹŠ£Ř24£›£ģņī‘ī”ŕÚÁņŗDermacentorvariabilisĶńRGDń£ŐŚ∂ĺňōĶįį◊Variabilin£¨ĹŲ”–5kD≤ĘįŁļ¨1łŲRGDń£ŐŚ£¨∆šń‹”––ßř◊ŅĻ’ŻļŌňō¶ŃIIb¶¬3ļÕ¶Ńv¶¬3£Ř25£›£ģSavignygrinī”»ŪŐŚÚÁOrnithodoros savignyi÷–∑ĘŌ÷£¨ňŁļ¨”–1łŲRGD–ÚŃ–Ķń—™–°įŚĺŘľĮ“÷÷∆ľŃ£Ř26£›£ģĶę «£¨≥żŃň‘ŕ…Ŗ∂ĺ÷–∑ĘŌ÷ĶńRGD∂ĺňōĶįį◊ĺŖ”–ŅĻ—™Ļ‹–¬…ķ◊ų”√£¨ŃŪÕ‚Ķń2ņŗRGD∂ĺňōĶįį◊≤Ę√Ľ”–÷ĪĹ”Ķń Ķ—ť÷§ĺ›£¨ĪŪ√ų∆š∂‘ŅĻ—™Ļ‹–¬…ķ“≤”––ßĻŻ£ģ
ĪĺŅőŐ‚◊ť∑ĘŌ÷Ķń…Ō Ųľłņŗ“‘Õ‚ĶńĶŕ4īůņŗRGDń£ŐŚ∂ĺňōĶįį◊(GenBankaccessionNo£ģACS71748.1)£¨ľīņī‘ī”ŕ∆Ŗ»ķų©Ņŕ«ĽŌŔĶńLj-RGD1£¨2£¨3Ķįį◊£Ř6£›£¨∆š÷–Lj-RGD1łĽļ¨įŽŽ◊įĪňŠļÕ1łŲRGDń£ŐŚ£¨Lj-RGD2”ŽLj-RGD3”–≤Ņ∑÷Õ¨‘ī–‘£¨∑÷Īūļ¨2łŲļÕ3łŲRGDń£ŐŚ;≤Ę«“Õ®Ļż Ķ—ť÷§√ų£¨rLjRGD3≤ĽĹŲĺŖ”–ŅĻ—™–°įŚĺŘľĮĽĻ”–ŅĻ—™Ļ‹–¬…ķĶńĻ¶ń‹(Fig£ģ3)£¨’‚–©Ļ¶ń‹ «Õ®Ļż“‘’ŻļŌňōő™į–Ķ„ř◊ŅĻ’ŻļŌňōŇšŐŚĶ√“‘ ĶŌ÷Ķń£¨ňĶ√ųLj-RGD3 «1÷÷Ķš–ÕĶńRGD∂ĺňōĶįį◊£Ř27-29£›£ģ
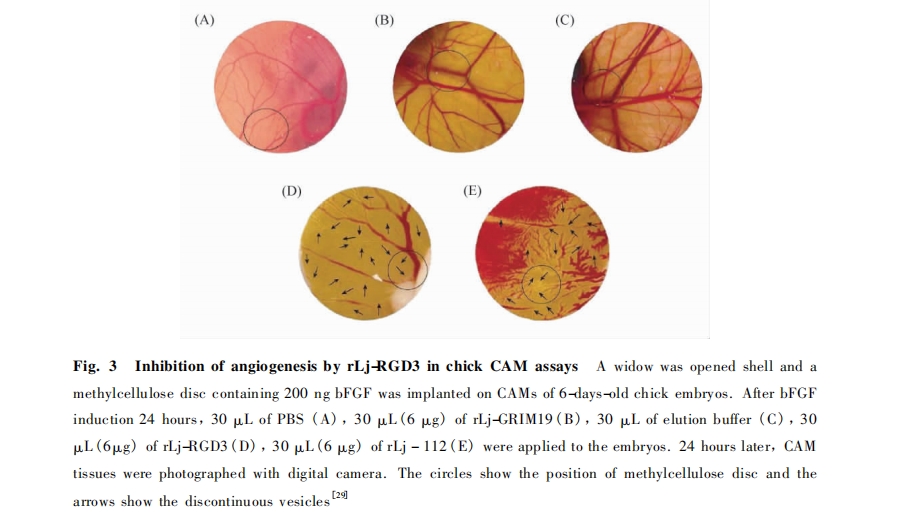
3.3 RGDĹťĶľĶń¶Ńv¶¬3į–Ōڳݓ©Õĺĺ∂
3.3.1RGDĹťĶľīęÕ≥ŅĻ÷◊ŃŲ“©őÔĶń¶Ńv¶¬3į–Ōڳݓ©Õĺĺ∂ ’ŻļŌňō¶Ńv¶¬3 «ņŪŌŽĶń÷◊ŃŲ÷őŃ∆į–Ķ„£¨∂Ý∆šŇšŐŚRGDŽńŅ…“‘Őō“žĹŠļŌ“©őÔ∑÷◊”≤ĘĹę∆š∂®ŌÚ‘ň š÷Ń÷◊ŃŲį–ŌÚ≤ŅőĽ£ģňý“‘£¨RGD–ÚŃ–”¶”√Õĺĺ∂≤Ľ÷Ľ «ŅĻ—™Ļ‹–¬…ķĪĺ…Ū£¨ĽĻ”–“©őÔį–ŌÚ◊ų”√£ģ
RGDŅ…‘ň‘ōįĘ√Ļňō(doxorubicin£¨DOX)ĶĹ—™Ļ‹–¬…ķńŕ∆§ŌłįŻ£ģŐŚÕ‚ ‘—ťĪŪ√ų£¨doxo-RGD4C(doxorubicin-RGD4CłīļŌőÔ)ļÕĶ•īŅĶńįĘ√ĻňōĺŖ”–ŌŗÕ¨Ķń÷őŃ∆◊ų”√£¨≥żÕ¨—ýĺŖ”–“÷÷∆–° ůŐŚńŕ÷◊ŃŲ…ķ≥§°Ę«®“∆ļÕĹĢ»ůĶńĻ¶ń‹Õ‚£Ř30£›£¨ĽĻń‹ĹĶĶÕ∂‘’ż≥£ŌłįŻĶń∂ĺłĪ◊ų”√£ģ’‚∆š÷–Ķń‘≠“ÚĪ„‘ŕ”ŕ÷◊ŃŲŌłįŻĪŪ√śłŖĪŪīÔń≥–©’ŻļŌňō£¨”»∆š «’ŻļŌňō¶Ńv¶¬3łŖĪŪīÔ”ŕ÷◊ŃŲŌłįŻĪŪ√ś£¨∂Ý’ż≥£ŌłįŻńŕ’ŻļŌňōĪŪīÔŃŅ‘Úļ‹ĶÕ£ģRGDŐō“ž ∂ĪūłŖĪŪīÔĶń’ŻļŌňō¶Ńv¶¬3£¨ī”∂Ý‘ž≥…÷◊ŃŲ÷őŃ∆÷–Ķńį–ŌÚ∂®őĽ£ģŌ÷“—≥…Ļ¶ļŌ≥…Ńňľł÷÷doxorubicinRGD4CłīļŌőÔ£¨“‘ŐŠłŖįĘ√ĻňōĶńį–ŌÚ÷őŃ∆;Õ¨ Ī£¨’‚–©łīļŌőÔĽĻĪĽ…Ťľ∆≥… ‹÷◊ŃŲ∑÷√ŕĶń√łľ§ĽÓĶń“©őÔ«įŐŚ£¨“‘ĹÝ“Ľ≤ĹĹĶĶÕ∆š∂ĺłĪ◊ų”√£¨≤ĘŐŠłŖ÷◊ŃŲį–ŌÚŐō“ž–‘£Ř31£›£ģņż»Á£¨Ō÷‘ŕ“——–ĺŅ≥Ų“Ľ÷÷ĺŖ”–Ōň»‹√łŐō“ž◊ų”√őĽĶ„Ķń“©őÔ«įŐŚ£¨ ĻĶ√“©őÔ«įŐŚ‘ŕ’ŻļŌňō¶Ńv¶¬3…ŌĽżĺŘļů”…Ōň»‹√łňģĹ‚≤Ę Õ∑ŇńłŐŚ“©őÔ£¨ī”∂Ý ĻįĘ√ĻňōĽ÷łīŌłįŻ“÷÷∆ĽÓ–‘£ģ’‚÷÷“©őÔ«įŐŚļÕŌň»‹√łĶń◊ų”√–ßĻŻ“—ĺ≠‘ŕŐŚÕ‚ŌłįŻŇŗ—ÝĶń Ķ—ť÷–Ķ√ĶĹ÷§ Ķ£ģľł÷÷RGDŽńį–ŌÚĶńįĘ√Ļňō“©őÔ«įŐŚ÷–£¨Ķ•Ľ∑ĶńRGD4CŇšŐŚĪ»∂ĢĽ∑ĶńRGD4C∂‘’ŻļŌňōĪŪŌ÷≥ŲłŁłŖĶń«◊ļÕ–‘£¨īňĹŠĻŻ”Ž3.1¬Ř ŲĶńĹŠĻŻŌŗ∑ī£Ř32£›£ģÕ¨ Ī£¨Ķ•Ľ∑RGD4CŽńĪ»∂ĢĽ∑RGD4CŽńňģ»‹–‘łŁľ—£¨“ÚīňŌ÷‘ŕĶńįĘ√ĻňōłīļŌőÔĺý”√RGDĶ•Ľ∑Žń÷∆Īł∂Ý≥…£ģ
īňÕ‚£¨◊Ō…ľīľ(PTX)“©őÔ«įŐŚ÷–ļ¨”–∂ĢľŘĶńc(RGDyk)ĹŠĻĻ”Ú£ģ Ķ—ť÷§ Ķ£¨łīļŌőÔPTX-RGD2‘ŕMDA-MB-435»ťŌŔ÷◊ŃŲ”Ž’ŻļŌňōĹŠļŌĶńĽżĺŘŃŅ£¨”ŽĶ•īŅĶńPTXĶńRGDŽń∂ĢĺŘŐŚŌŗÕ¨£Ř33£›£ģ“Úīň£¨∆šŅ…Õ®ĻżRGD”Ž’ŻļŌňō¶Ńv¶¬3ĹŠļŌ∂ÝŐŠłŖ÷◊ŃŲ÷őŃ∆Ķńį–ŌÚ–‘£ģ
RGD”Ž–ĮīÝįĘ√ĻňōĶń÷¨÷ ՌѨŔ“≤Ņ…”√”ŕ’ŻļŌňō¶Ńv¶¬3Ķńį–ŌÚ÷őŃ∆£ģ”–—–ĺŅ’Ŗ£Ř34£›∂‘RGDÓźň∆őÔ(RGDm)–ř őĶńő»∂®÷¨÷ ŐŚ(stabilizedliposomes£¨SL)–ĮīÝįĘ√Ļňō‘ŕ÷◊ŃŲ≤ŅőĽĶńĽżĺŘ£¨“‘ľįŌÚ÷◊ŃŲŌłįŻńŕīęĶ›Ķń«ťŅŲĹÝ––Ńň—–ĺŅ£ģŃų ĹŌłįŻ Ķ—ťļÕľ§Ļ‚Ļ≤ĺŘĹĻĶńĹŠĻŻŌ‘ ĺ£¨”ŽSL-DOXŌŗĪ»£¨ŌłįŻ∂‘RGDm-SL-DOXĶńĹŠļŌľį…„»°ĺýŌ‘÷Ý‘Ųľ”£ģ’‚ĪŪ√ų£¨RGDm–ř őĶńSLŅ…◊ųő™Őō∂®“©őÔį–ŌÚ‘ōŐŚ£¨∆šÕ®Ļż ‹ŐŚĹťĶľĶń∑Ĺ ĹĹę“©őÔ∑÷◊””––ßį–ŌÚ÷ŃŐō∂®÷◊ŃŲ≤ŅőĽ£ģ≥żīň÷ģÕ‚£¨“≤”–“‘ĺŘ““∂Ģīľ(PEG)◊ųő™Ń¨Ĺ”őÔѨŔRGDļÕ÷¨÷ ŐŚĶńĪ®Ķņ£ģŃŪ”–őńŌ◊Ī®Ķņ£¨…Ōőń÷–ŐŠĶĹĶńRGD10ĪĽņŗ÷¨∑÷◊”łńŃľļů£¨łŁ»›“◊ĹÝ»ŽįŁļ¨įĘ√ĻňōĶń÷¨÷ ŐŚ£Ř35£›£ģ∂ÝC26÷◊ŃŲń£–Õ Ķ—ť“≤÷§√ų£¨’‚–©÷¨÷ ŐŚĶńѨŔļÕńŕĽĮĪ»Ķ•īŅĶńįĘ√ĻňōĺŖ”–łŁļ√Ķń“÷ŃŲ–ßĻŻ£ģ
3.3.2RGDĹťĶľĶńļňňŠį–ŌÚ◊™»ĺ RGD“żĶľļňňŠ◊™»ĺĶńń‹Ń¶‘ŕŐŚńŕŐŚÕ‚∂ľĽŮĶ√÷§ Ķ£ģRGD “żĶľļňňŠ◊™»ĺĶń∑Ĺ Ĺ”–2÷÷:“Ľ «RGDŽńĻ≤ľŘĹŠļŌ”ŕPEI£¨»Ľļů‘ŔĹęRGD–ř őĶńPEI”ŽDNAĽÚsiRNA–ő≥…Ļ≤ĺŘőÔ;ŃŪ“Ľ÷÷ «ĹęRGDŽńŃī÷ĪĹ””ŽĺŘļŌőÔ÷ųŃīļŌ≥…‘ŕ“Ľ∆ū(RGD-PEG-PEI)£Ř36£›£ģRGDį–ŌÚĶńļňňŠ‘ōŐŚ‘ÚįŁņ®∑«≤°∂ĺņŗļÕ≤°∂ĺņŗ2÷÷£¨∑«≤°∂ĺņŗļňňŠ‘ōŐŚįŁņ®—ŰņŽ◊”ĺŘļŌőÔ°Ę—ŰņŽ◊”÷¨÷ ŐŚļÕ—ŰņŽ◊”ŽńņŗĶ»£¨≤°∂ĺņŗļňňŠ‘ōŐŚ‘Ú”–ŌŔ≤°∂ĺÕĺĺ∂ļÕ∑«ŌŔ≤°∂ĺÕĺĺ∂£Ř37£¨38£›£ģ
4ő Ő‚”Ž’ĻÕŻ
◊‘ī”∑ĘŌ÷—™Ļ‹–¬…ķ «÷◊ŃŲ…ķ≥§Ķń÷ō“™“Úňō£¨÷◊ŃŲ–¬…ķ—™Ļ‹ő Ő‚≥…ő™Ļę»ŌĶń÷őŃ∆į©÷ĘĶń”––ßńŅĪÍ£ģ¶Ńv¶¬3’ŻļŌňō‘ŕ–¬…ķ—™Ļ‹ĪŪ√śĪŪīÔ£¨Ķę≤Ľ «‘ŕECŌłįŻĪŪ√śĪŪīÔ£¨’‚÷÷”–ŌřĶńĪŪīÔ Ļ∆š≥…ő™ņŪŌŽĶńį–Ķ„£ģő™ŃňŅ™∑Ę’‚÷÷į–ŌÚ“©őÔ‘ň šĶńŐō–‘£¨ńŅ«įĶń—–ĺŅĹę¶Ńv¶¬3’ŻļŌňōĶń ∂Īū–ÚŃ–RGDŽń≤Ś»ŽĶįį◊÷ °ĘĺŘļŌőÔ°Ę÷¨÷ ŐŚ°Ę≤°∂ĺļÕ∆šňŻĶńĽý“Ú‘ň š‘ōŐŚ£¨Õ®Ļż∆šį–ŌÚ◊ų”√£¨Ĺę–°∑÷◊”“©őÔ°ĘļňňŠņŗ“©őÔĶ»‘ň š÷ŃŌłįŻńŕ£ģ∂Ý’‚÷÷ĽĮ—ßļŌ≥…ļÕ÷ō◊ťŐŚĺŘļŌőÔĶńÕĺĺ∂∂ľ «Ņ…––Ķń£¨∂ľń‹īŔĹÝ“©őÔį–ŌÚŃž”Ú÷–RGDĹťĶľĶń“©őÔ‘ň šĶńĻ„∑ļ”¶”√£ģ
√‚‘ū…ý√ų£ļĪĺőńő™––“ĶĹĽŃų—ßŌį£¨įś»®Ļť‘≠◊ų’Ŗľį‘≠‘”÷ĺňý”–£¨»Á”–«÷»®£¨Ņ…Ń™ŌĶ…ĺ≥ż°£őń’¬ĪÍ◊Ę”–◊ų’Ŗľįőń’¬≥Ųī¶£¨»Á–Ť‘ń∂Ń‘≠őńľį≤őŅľőńŌ◊£¨Ņ…‘ń∂Ń‘≠‘”÷ĺ
