
��ժҪ�� ��������һ��С���ӻ����ģ���Դ�㷺����������������͵����з�����Ҫ���ã��Ҿ��п������ԣ���Ϊ�����������ҩ�������ṩ�˿��ܡ�LL�\37����֯����ø�����ؼ�����Ψһ�������ڵij�Ա���Ը������Ծ��������Ծ������������á����Ĵ�LL�\37���Խṹ����з�����������LL�\37�ڿ��������ڶ��ء��������ϡ����ߵ��ڵȷ�������û��Ƽ��ٴ�����DZ����չ������Ӧ��ǰ����Ϊ��ҩ�з���ʳƷ�����Ͷ������������ṩ˼·��
����������������ռ����Ҫ�ĵ�λ�����ȹ������˵���������ũҩ��ʳƷ���Ӽ�����������ֳ�з�������Ҫ���ã���Ϊ���ŵĿ���ҩ��Ȼ������ 20 ����70������з�����Ч������ȴ��Ϊ���⡣���ڿ����صij���ʹ�ã�Խ��Խ���ϸ�����ֳ���ҩ�ԣ�������������ǿ�ġ�����ϸ������1~4�ݡ�ϸ����ҩ�Ե���ǿʹ������ʧȥЧ������ˣ������ܹ���������ص�������Ч��Ʒ�̲��ݻ���5��6�ݡ�
��������Դ�㷺�����緢�������������ϵͳ�С����ϸ������������ܶ���ߵ�ֲ��Ͳ��鶯����Ҳ��ȡ�����ƵĶ��ġ����ֻ����ľ�����Ȼ���߹��ܣ��������ֿ���ԭ�������ֵ���Ҫ�������ϣ���˱���Ϊ�����ģ�7��8�ݡ������ľ��п����㡢��Է�������С�����ȶ��Ըߡ�ˮ���Ժõ��ص㡣Ŀǰ�о����漰�����ӽṹ�����Ӹ��������û��Ƶȶ���9�ݡ��ӿ������Ʒ����������������������������壬ϸ����ͨ���������Կ��������ã����յ�����ҩ���IJ������������Ŀɾ�����������ʹϸ��Ĥ���ף����մﵽ��������Ч������˲��ײ�����ҩ�ԡ����ԣ������ļ�����ܳ�Ϊ�����ص����Ʒ��10�ݡ�
���У���Դ�Կ�����ϸ�����Ե������췴ӦС����ʾ��һ�����ٴ��������ƣ�11�ݡ�Ŀǰ��Դ����������֯����ø�����أ�cathelicidin�����塢�����أ�defensin�����塢�����鰱�ᵰ����ȣ�12�ݡ������ؼ�����һ����Դ�Կ����ģ��������Խ�ǿ���� 1~10 µg����ʾ���־����ԡ���֯����ø�����ؼ����Dz����ද�����еĿ����ģ�LL�\37 ����Ψһ�������ڵĿ����ģ����ж������á���֯�أ�histatin����һ����鰱��Ŀ����ģ��н�ǿ��ϸ���Ϳ�������ԣ�13�ݡ�����ҽѧ����������������Դ�������ѽ����ٴ������14�ݣ�������������1�\11��human lactoferrin1�\11��hLF1�\11���ѽ����ٴ����ڽΣ����ڹ�����ֲ���̿���Ⱦ���ƣ������ B�����HB�\107�������ٴ�ǰ�ڣ����ڴ���������Դ���������ĵ������protegrin analogue��IB�\367����������ٴ����飬�������Ĥ�ף�15~17�ݡ����ԣ���Դ�����ļ��п��ܳ�Ϊ�о��ȵ㡣
LL�\37 ��Ҫ�ϳɲ�λΪ������ϸ�������������Ԧ������ṹ�����ɾ���ϸ��������ϸ�����ǵ���ϸ����������ϸ���Ƚ��з��ڣ�18�ݡ�LL�\37�������Ƹ������ԣ�G+�����������ԣ�G-���������ص㣬���п��ڶ��ء��������á��ٽ�Ѫ���������õȹ��ܣ���Щ����ʾ������濹���صĿ����ԡ������������ں������٣��ҷ��봿�����踴�ӣ�������ȡ�����ð���ͳ���뷽���Ѳ��ʺϴ���������19�����ֽ����û��̻�������ɸѡ���������ߡ��߿������ԡ��Ͷ��Կ����ģ���������һ��������20�ݡ�
Ȼ��������ڿ����أ�LL�\37����������Ȼ�������ƣ������߿������Բ�������Ѫ���Ժ�ϸ�������ǹؼ����21�ݡ�Ŀǰ����Ŀ�Ļ����������ڽ��и�Ч�����ǽ�Ϊ������ֶΡ��ڽ������θ���ǰ��������Ҫ�ڻ������ı俹�������ƽṹǰ���£�ͨ���������û����������н�ȡ����ϻ�ѧ��ȷ������Ŀ�Ŀ����ģ��⽫��δ��������Ч�����ĵ���Ҫ������22��23�ݡ��ı俹���Ķ�ϸ��ϸ��Ĥ�Ľ�����ã��Կ������־����Ե����йؼ����á��������ȣ�24�ݷֱ��� Gln16��Asn26 �� Gln36 ȡ�� Glu16��Asp26 ��Glu36��ʹ�����+5.8 ���ӵ�+9.0���������������LL�\37 �����ˮƽ����δ�ı� LL�\37 �ռ�ṹ��Ga�\gnon �ȣ�25�ݷ�����������Ҳ�뿹�������йء�������������ͬ����£�����Խ���������Խ��Ŀ����Ŀ���Ч��Խ�á�
��ˣ����Ķ� LL�\37 �����ĵ�����ṹ���н�����������ͬ����Ĺؼ����ã���������ѧ���ԣ����������ڶ��ء����ߵ��ڡ��ٽ�Ѫ�����ɣ��ȷ�����в�������Ϊ������ĵĿ����ṩ�������ݣ�����LL�\37DZ��Ӧ�÷����ṩ˼·��
1 LL⁃37����ṹ�����﹦��
1.1 ����ṹ
�������ӿ�������18��human cationic antimicrobialprotein 18��hCAP18������֯���������ؼ����Ա������Ҫ������ʽ������������ϸ���У���غ�衢���衢����Ƥϸ����������Ƥ�����γ�ϸ����ʳ�ܡ������Ȳ�������Ҫͨ�����п������Ե� C ����֬�������ӣ�26�ݡ�hCAP18�Էǻ���ǰ�嵰����ʽ�����ڿ����У���һ�������·��ѳ����е�C�˻����ģ���LL �\37�ģ�27�ݡ������� hCAP18 ��������ø 3 ��������ѽ⡣��ʵ�������ֻҪ����ø 3 Ũ���㹻�ߣ�hCAP18����ȫ������LL�\37��28�ݡ�
LL�\37�� N��ǰ���е�����������л���L�������� 37 ����������ɣ���˵��� LL�\37��29�ݡ�LL�\37 �����������֯�����б���ر�������������Ƥ����������Ϳ�ǻ��Ƥϸ���С����⣬��Ҳ������һЩ�˿�����Һ����Һ��ѪҺ�У�30�ݡ�LL�\37 ���������������ڰ��� 1�� TAAA ���С�1������ 6��ת¼���ӣ�nuclear factor interleukin�\6��NF�\IL�\6�����λ�㡢3�� �� �� �� �� Ӧ �� �ӣ�acute phase response factor��APRF���������������ɵ���֢���ӽ�ϣ��Ӷ�����LL�\37�������ˮƽ��31�ݡ�
�ڴ�ˮ�У�LL�\37�γɲ�����������ṹ������Χ�����ı� �� �����ṹʱ�����γ������������ӽṹ��LL�\37���ڵ�C�˺�N�˦��\���������˵�������ͨ��ת�ۻ������ֿ����� 5��6��17�� 27λ�������������ͬһ�������죬�γ���ˮ���棬����ɲл�����ˮ������븺��ɷ��ӽṹֱ����������G+֬�ǡ�G-�ױ���ȣ�32�ݡ������庹Һ�У�hCAP18�� LL�\37Ƭ�β��KS�\30�� RK�\31�������ߵ� N�˰����Ậ���� LL�\37 �� 6/7�����н�ǿ�Ŀ������ԣ������������ߵ���������33�ݡ����⣬LL�\37��ȱ��N��4�ְ�����ʱ�����ܱ��ֿ������ԣ�������Ѫ����ɥʧ����Щ���������N�������Զ��ĵĹѾۡ���������Ѫ�Ϳ�����øˮ������Ҫ���á�����ʵ��С���о��˽ض��ĵĻ��ԣ����˽� LL�\37 ��С���������34�ݡ����������LL�\37����С�����ڰ�����17~29λ�� 18~29 λ֮�䣬���� LL�\37 �� C ���������п������������Ϳ����������á�
�о���ʾ����Ӻ��ͬԴ���� RL�\37 �������Ϊ+8������ LL�\37 ��+6����������Ҫ���� LL�\37��35�ݡ��������LL�\37�еļ���������Ŀ�����ı�����\�������������俹�����ԣ�36�ݡ������д�����ɺ���ˮ���������ĿҲ�����������йأ������й����������ص㣬��˿�ͨ�����·�ʽ���죺���ڽṹ��ӦΪ��״������״������״�ϱ��ַ��ӵ���ˮ�Ժ���ˮ�ԣ��ڴ����������븺�������ᡢ��ˮ������ˮ�����ᣬ�������γ�����״�ṹ�����ᣬ��߿����Ļ��ԣ������������Ժ���ˮ����������������ɱ�����Ϳ��ڶ���������
1.2 ���﹦��
1.2.1 �������� LL�\37�� G+�� G-���п������ã��ھ��������£�������� LL�\37 ��������ϸ���ӣ�����ϸ��ϸ��Ĥ������������������������37�ݡ�Ŀǰ�������ּ�˵֧��LL�\37�ܽ�ϸ��Ĥ�Ļ��ƣ���̺ģ�ͣ�ͼ 1���ǻ��� LL�\37 ���溬�������Ӱ�������ϸ��Ĥ��֬ͷ����ϵ����ۣ�������ϸ��Ĥ�����γɵ�̺�ṹ�����ƻ�ϸ��Ĥ�����ԡ��� LL�\37 �����ṹ����ɺ���ˮ�Է����仯ʱ���俹�������ܵ�Ӱ�죻���ο�ģ�ͣ�ͼ 2����Ҫ����Ĥ�������������ʣ�ʹϸ��Ĥ�γɻ��οף��Ӷ�й¶ϸ���������ʹϸ��������38�ݡ����ڲ��鶯����˵����ϸ��Ĥ������������״̬������LL�\37��Ĥ���ý�����ϸ�������ܵ����ˡ����⣬LL�\37 ����ǿ��Ƥϸ��ĤӲ�Ȳ�������ͨ�ԣ��Ӷ�����ϸ��������39�ݡ���Σ�LL�\37����˫���������ṹ����ˮ�������״ٽ��Ѿ����γɣ����忹�������Ѿ�����̬�� �� �����ṹ�����ȣ����������϶�ʱ���������Խ�ǿ��
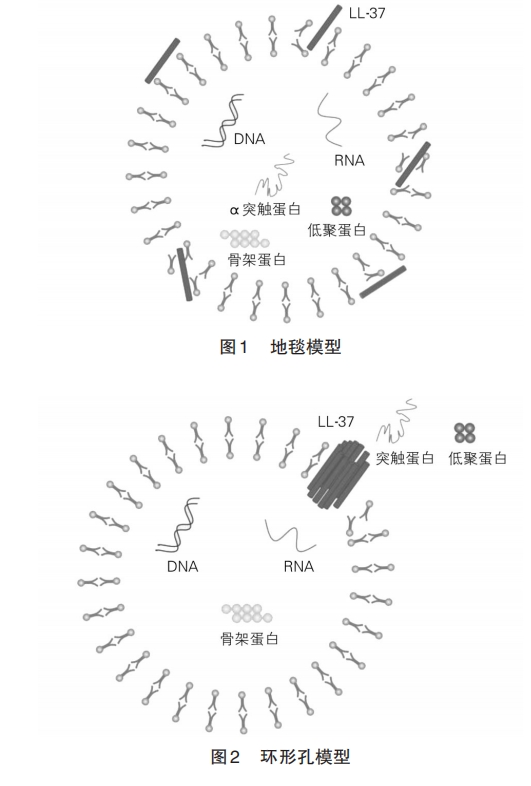
1.2.2 ���ڶ������� ֬���ǣ�lipopolysaccharide��LPS����Ҫλ�� G�\ϸ����Ĥ�ϡ����������Ҫ�У����� LPS �յ��� Toll �����壨Toll�\like receptor��TLR���\������ ��B��nuclear factor kappa�\B��NF�\��B���ź�ͨ·�У�LL�\37 ��������á����⣬LL�\37 ���� LPS �յ��� NF�\��B�ǵ�λ p50�� p65��ת¼��41�ݡ���˿��ԭ�����ø��mitogen �\activated protein kinase��MAPK��ͨ��������Ӷ����Ӵ���Ӧͨ�����ԣ�42�ݡ���ѡ���Եص��ڻ���ת¼�����ʹ���Ӧ���� p50�������������ө\����tumor necrosis factor�\����TNF�\�����ı�������� LPS �� LPS ��ϵ��ף�LPS�\binding pro�\tein��LBP���������ֻ����� 2��myeloid differentiationprotein 2��MD�\2���� TLR ���������ϣ���������ͨ·�ļ��������Щ���Ƶ��ڣ���LPS����IJ�����Ӧ�ɱ�LL�\37��Ч���ơ�
1.2.3 ���ߵ������� ���ȶ�״̬�����£�������ϸ�������ṹ�Ե������Խ��������������������ϸ�����������������֢��λ�γ�������ϸ������ɱ�����磨neutrophil extracellular traps��NET�������ṩ��Ũ�ȿ������ӣ����ٿ�������ϸ����Ⱦ��43��44�ݡ��������������֢�� NET �����������ͷ� LL�\37��ͬʱLL�\37Ҳ�ٽ�����������ϸ���Լ���������ʽ�γ� NET���� NETͨ�� C1q��Ѫ�岹�壩/LL�\37����������LPS����ľ���ϸ������IL�\6��45��46�ݡ������о�������LL�\37 ��Դ�ij�����ͻ״ϸ����maturationof dendritic cell��mDC����LPS������һ�����͵ĸ�����Tϸ����helper T cell��Th1���յ�ϸ�������ף�IL�\12��IL�\6 �� TNF�\�� �������ӣ�IL�\4 �������ͣ������⣬��mDC�̼��£�Tϸ�������������ʦø����أ�interfer�\on gamma��IFN�\�ã��ĺϳ��������ӡ���ˣ�LL�\37�ƺ�����������ϵͳ����Ӧ������ϵͳ֮���������ͨ������ LL �\37 ������Ѫ������ϸ����peripheralblood mononuclear cells��PBMC���е����ã��о���Ա����LL�\37Ҳ�ٽ�������Tϸ������������Ӱ��Tϸ���������֢��ֲ��Ѫ���أ�phytohemagglutinin��Pha������ݱ����£�LL�\37�Ŀ��յ���ϢTϸ����ֳ���������ʹ� PBMC �������ӱ��IFN�\�ã�TNF�\�������������ķ�����������Ⱦ����ʱ���÷�Ӧʼ�տ��ƴ���Ӧ���Ա�����������������֢��Ӧ��47�ݡ����ң�LL�\37 ���� TLR 3 �ı���ٽ����ڸ���ˮƽ�������ӣ����᰷2��3�\˫����ø��indoleamine2��3�\dioxy�\genase��IDO���ݡ�IL�\10��ת���������Ӧ£�transforminggrowth factor�\�£�TGF�\�£����������ʸ�ϸ���Թ��ȴ̼���Tϸ�������ƹ��ܡ���ˣ�LL�\37�����ṩ�Ի� �� �� �� �� ���� ͨ �� �� �� ; �� �� �� �� �� �� �����ã�48�ݡ�
1.2.4 �������� Ѫ�����ɡ���Ƥϸ��Ǩ�ƺ���ֳ������άϸ�����ƽ�ԭ��������Ƥ�������ϵ� 3���ؼ����أ������������ͨ�������Ƥ�����������壨epidermal growth factor receptor��EGFR���鵼��LL�\37�Ĵ��ڿɼ���G����ż�����壨G protein�\coupledreceptors��GPCR���� EGFR ��ʽ����ٳɽǻ�ϸ��Ǩ�ƣ��ٽ���������LL�\37�����յ�Ѫ�����ɣ��ɽ���Ƥǰ��ϸ���ۼ���������Ҫ����λ��������Ƥϸ�����ٽ���Ƥϸ����ֳ���γ�Ѫ�����ṹ��49�ݡ�
�����������⣬LL�\37�����п�����������������ã��⽫ΪLL�\37Ͷ�뵽�ٴ������ṩ���ݡ�
2 LL⁃37�ٴ�����DZ��
2.1 �����Լ���
����������� 1��herpes simplex virus 1��HSV�\1��������������Ƶ�һԱ���������ڿ�ǻ����ֳ���Ĥ��50�ݡ����о���ʾ LL�\37 �� HSV ����������Ӱ�졣�������п��������ã��ھ� LL�\37 �����ĸ�Ⱦ HSV�\1 �����γ�ϸ���У���ʾ 5 ����Ҫ�����ش̼�����۸������յ��������ظ�����1��interferon�\induced protein with tetratricopeptide repeats 1��IFIT1���������ش̼�����ø���� 20 kDa��interferon stimulatedexonuclease gene 20 kDa��ISG20���ȣݱ������ӣ�51�ݡ�������ϸ������ʵ���У�vero ϸ���������̺���ϸ������ LL�\37 Ԥ������Ȼ���� HSV�\1 ��Ⱦ��Ҳ��HSV�\1�����������٣�52�ݡ���ѧ����֬����LL�\37���� HSV�\1��Ⱦ�� HaCaT ϸ�����������������γ�ϸ������������֣�������LL�\37������ȣ�����֬����LL�\37 ���Ը�С�������ڸ��������ױ����ա���ˣ�֬���� LL�\37 ΪԤ�� HSV�\1 ��Ⱦ�ṩ��˼·�����п��ܳ�ΪDZ��ҩ���53�ݡ�
���ؼ��Ժ���ϵͳ�ۺ�����״����2�ͣ�severeacute respiratory syndrome coronavirus 2��SARS�\CoV�\2�����������е��¹ڷ�����Ҫ��ԭ�壬���ְ�Ĥ����RNA�����ɵ��¼��Ժ���ϵͳ����������ʱ����������54~56�ݡ����ݽ��������ά���� D �ϵ� LL�\37�������ֱ�ӻ��������� SARS�\CoV�\2����ֹ�临�ƣ����ӿ���ϸ�����ӵIJ��������ٴ������߷� Ӧ��57�� ���� �� �� �� �� �� ϸ ����mesenchymal stemcells��MSC���������������� SARS�\CoV�\2 ��Ⱦ���ٴ��������ڽ����С�MSC�������ֿ��������ӣ����� �� �� �� LL �\ 37���� �� �� �� �� �� �� ʹ �� �� �� ʧЧ��58��59�ݡ�Ȼ����LL�\37�� SARS�\CoV�\2�Ļ����в���ȷ�����һ��̽����
������������Ebola virus��EV����һ������˿״�����Ƶĵ���RNA�������н�ǿ��Ⱦ�Ժͽϸ������ʣ�60�ݡ�������֣�LL �\37 �� LL �\37 ���壨GI �\20 ��17BI��������������ˮ���Կ��ײ����ۿɱ���EV�\�ǵ��ף�glycoprotein��GP���ݺ�Ұ���� EV ��Ⱦ������Ҫ���á���ЩLL�\37ҩ��ͨ����HeLaϸ��ϵ���˹�����ϸ�����е���֯����øb�鵼��EV�ǵ��ӹ��������� EVϸ�����룬�����ܿ��Ʋ������ơ�����Ҫ���ǣ����ֺ���D�\��������������Ķ���ø����еֿ������� L�Ϳ����ĸ���Ч��61�ݡ���Щ������һ��̽�������ڶ���ģ����Ԥ�� EV ��Ⱦ���ã��Լ��뿹����ҩ�����õĿ����ԡ�
2.2 �����Լ���
��ֱ������colorectal cancer��CRC���ǵڶ����������İ�֢��ռ��֢������ 9.2%��62�ݡ�LL�\37�����᳦�Ĥ�б���ǿ�ң����ڽ᳦ֱ����֯�б����µ�����LL�\37ˮƽ�ѱ���Ϊ�ǽ�ֱ�����������־�ͬʱ��LL�\37�ɽ������Ƶ���Bϸ���ܰ����\2����b�\cell lymphoma�\2��Bcl�\2��ˮƽ����ߴٵ��� Bax/Bak��Bcl�\2�����Ǽ����Ա��ˮƽ��LL�\37Ҳ������P53 �ϵ������������ӣ�p53 upregulated modulatorof apoptosis��PUMA�����������ƻ��� p53 �ı��PUMA �� p53 ��ֱ�Ӱе㣬Ҳ�ǽ�ֱ����ϸ�������ĵ������ӡ��ڽ᳦��ϸ���У�LL�\37 �����յ������յ����ӣ�apoptosis inducing factor��AIF��������øG��endonuclease G��EndoG�����������쵰��ø��cysteinylaspartate specific proteinase��caspase���������Ե������ڽ᳦��ϸ���У�һ�ֻ���LL�\37�Ļ��Ƽ���GPCR�\p53�\Bax/Bak/Bcl�\2�źż��������� AIF/endog�鵼�ĵ�����63��64�ݡ���LL�\37�����ڴ�����������magneticnanoparticles��MNP������ʱ���俹������������ߣ���ʾ��һ��DZ����65�ݡ�
��θ��ϸ���У�LL�\37 ����������������θ��֯�����о���ʾ LL�\37 ͨ������̬�������ף�bonemorphogenetic protein��BMP���ź�ͨ·���ڵ���ø�弤���66�ݡ�BMP�źŴ���Smad��Sma�\ and Mad�\relatedprotein��1/5/8��ļ�������ữ����Smad4�γ���Դ�����壬�� Smads ����������ϵ�ϸ�����ڵ��ػ���p21Waf1�ı���ˮƽ���ڸ�;���У�LL�\37Ҳ���յ���ˮƽ��ϸ�����ڵ��� E2��p21Waf1��ϸ�����ڵ���E2 ����ˮƽ�ĸı������ G0/G1��ϸ���������͡����ԣ�LL�\37 ͨ������ø�������Ļ��Ƽ��� BMP �ź�ͨ·������θ��ϸ����ֳ��
��ǻ�۰���oral squamous cell carcinoma��OSCC������ͷ���������������������黯������ʾ����������ǻ�Ĥ��֯��ȣ�LL�\37 �ͱ�������֯ѧ�ֻ����ܰͽ�ת���йء���˵�� LL�\37 ������ OSCC ����һ�������������ӡ�hCAP18��һЩ��ǻ��ϸ��ϵ�еı�������漰������ CAMP �����������DNA ����״̬��68�ݡ�Ҳ���о�������hCAP18 ��OSCCSAS �\H1 ϸ�����յ����쵰��ø�������Ե��� ����������������άϸ���� HaCaT ϸ�������ޣ�69�ݡ��������������һЩ��ϸ�����и������̴���֬������Ҳ�����ǿ��� LL�\37 �Բ�ͬ��ϸ��ѡ���ԵĹؼ����أ��Ӷ�Ϊ LL�\37 ���������ṩ��һ���µĿ����ԣ�70��71�ݡ�
���⣬LL�\37 ��ѪҺ����������Ҳ����һ���������ã����ƿ���������ϸ������ͨ·�йء���Щ���Ϊ�����ٴ������ṩ�˷���
3 LL⁃37Ӧ��ǰ��
3.1 ��ҩ
��Ӧ��Ƥ�ף�atopic dermatitis��AD������TLR2��LL�\37�ĵͱ��PѪ��LL�\37�ĵ�ˮƽ��ʾAD�����������ߺ�Ƥ�����Ϲ���ȱ�ݵ����⡣ά����D���ܼ���ά����D���壬ͨ��TLR2;���ٽ�LL�\37����Ӷ��� AD �������ã�72�ݡ�����δ�������LL�\37�й�ҩ�︨��ά����D�Կ���Ӧ��Ƥ�ס�
��������Ѫ֢��Ҫ�ɸ�Ⱦ����������ҩ���IJ�����������ѡ����Խ��Խ���֣������������ֿ�����Ϊ��Ҫ�����о���ʾά����D������忹���� LL�\37 �ı���Ӷ�����������Ⱦ��������Ϊ�����������ṩ�������ϡ��⽫Ϊ��ƿ��������Ʒ��ҩ���ṩ˼·��73�ݡ�
3.2 �����������Ӽ�
LL�\37 �������ҿ������Ըߣ�ϸ�����ײ�����ҩ�ԣ������к��ڶ��أ�������Ϊ�����ص��������Ʒ�����ѡ����ʵı���ģ��������LL�\37������������������и�Ч��������Ӫ���Ķ����������Ӽ��дٽ����ã�ʹ��ӵ��һ���ľ���Ч�档
3.3 ������
LL�\37 �� CP1 ��ͨ�����̴��ģ�����ģ��俹������ǿ����Է�������С��121�����30 min�������Ըߣ�ˮ���Ժã���ȫ�������ײ�����ҩ�ԡ���ţ�����������������Ķ�������һ���������á�ϸ������ʵ����������ǶԴ����ϸ��������ֻ��Ϊ�����ĵİ�ȫ���ṩ���ݣ���һ����ȫ��Ӧ���д���һ���о���ͼ3����74�ݡ�
4 չ��
������LL�\37��Է�������С���ṹ���⣬����ѧ���ܶ�����������������ܵ����������������ײ�����ҩ�Ե����ʣ�ʹ����п��ܳ�Ϊ���������Ʒ�����⣬��������ʳƷ�е����Ա��֣�Ҳ�п���ʹ���Ϊ��һ��ʳƷ���Ӽ��������ĵ�ʹ�ã��ɸ���Ч�ط�ֹ�����߲����У���������ҵ��ʧ���⽫����ٽ��ҹ�������ֳҵ������չ��ʹ�ÿ����Ŀ���ǿ����Լ����ֿ��������ټ��������Ϳ�����ʹ�ã��Ӷ�����ҩ���ڶ������ڼ����Ʒ�еIJ�����Ϊ�ҹ�����������ṩ�˸���ȫ������ʳƷ�����ڿ����ľ��й������ԣ���G+��G-���кܺ���ɱ���ã���˿ɹ㷺Ӧ����������ֳҵ�������ľ��й���Ӧ��ǰ�������û��̼����������鿹���IJ�Ʒ���кܺõ��ٴ�Ч��������Ҳ�в�����ҵ�������˿����IJ�Ʒ��������ؿ����ģ����ڶ��������еõ��˽Ϻ�Ӧ��Ч���������ĵ��о��ȳ��Ʊش���һЩ��ҵ����ɫ��������ҽҩ���棬������Ŭ��������ɫ����Ч���Ͷ�����Ⱦҩ���ũ��ҵ���棬��������Ϊ�����������Ӽ�Ҳ��Ͷ��������������ʳƷ��ҵ����Ҳ���ֳ���DZ���������о����룬��������Ϊ���Ӽ��������еĹ㷺Ӧ���ս���ɿ��ܡ���������δ����չ�о�������DZ����Ӧ�������ӣ����ھ���DZ�ڵ����Ӧ�ü�ֵ��
��������������Ϊ��ҵ����ѧϰ����Ȩ��ԭ����ԭ��־���У�������Ȩ������ϵɾ�������±�ע���������³����������Ķ�ԭ�ļ��ο����ף����Ķ�ԭ��־
