
’Σ“ΣΘΚΧλ»Μ¥ΈΦΕ¥ζ–Μ≤ζΈο «÷Ί“ΣΒΡ“©Έοά¥‘¥,Ζ«ΚΥΧ«ΧεκΡ(non-ribosomal peptide,NRP) «Ή‘»ΜΫγ÷–ΙψΖΚ¥φ‘ΎΒΡ¥ΈΦΕ¥ζ–Μ≤ζΈο,ΤδΕύ―υΒΡΜ·―ßΫαΙΙ ΙΤδΨΏ”–Εύ÷÷…ζΈοΜν–‘,»γΩΙ―ΉΓΔΩΙ÷ΉΝωΓΔΩΙ≤ΓΕΨΒ»ΓΘΜυ”ΎΖ«ΚΥΧ«ΧεΕύκΡΚœ≥…ΟΗ(nonribosomal peptide synthetases,NRPS)ΡΘΩιΜ·œΏ–‘Κœ≥…ΕύκΡΒΡ‘≠άμΕ‘Τδ¥ΏΜ·ΡΘΩιΫχ––ΗΡ‘λΓΔ÷ΊΉι,Ε®œρ…ηΦΤΕύκΡΒΡ…ζΈοΚœ≥…ΆΨΨΕ“‘ΜώΒΟΡΩΒΡΕύκΡ“―≥…ΈΣ“ΜΗω―–ΨΩ»»ΒψΓΘ»ΜΕχ‘”ΚœNRPS¥φ‘Ύ¥ΏΜ·ΡΘΩιΈόΖ®Φ”‘ΊΡΩ±ξΑ±ΜυΥαΜρΕύκΡΚœ≥…–߬ œ‘÷χΫΒΒΆΒ»÷νΕύΈ Χβ,œό÷ΤΝΥΤδ”Π”ΟΓΘΫϋΡξά¥,NRPSœΌή’θΘΜ·”ρ(adenylation domain,A”ρ)ΦΑΥθΚœΫαΙΙ”ρ(condensation domain,C”ρ)ΒΡΒΉΈο―Γ‘ώ–‘ΓΔNRPS―«ΜυΦδΕ‘Ϋ””ρ(docking domain,DD)ΚΆΡΘΩιΦδΝ§Ϋ”«χ(linker)ΒΡ―–ΨΩ“―»ΓΒΟΫœ¥σΆΜΤΤΓΘ¥”C”ρΕ‘ΒΉΈοΒΡ―Γ‘ώ–‘ΦΑ“‘≤ΜΆ§»ΎΚœ±ΏΫγΫχ––¥ΏΜ·ΒΞ‘ΣΧφΜΜΝΫΖΫΟφΫχ––Ήέ ω,Ϋι…ήNRPS¥ΏΜ·ΡΘΩι÷ΊΙΙΒΡ―–ΨΩΫχ’Ι,≤ΔΗ≈ ωΝΥΗςΧφΜΜΖΫΑΗΒΡ”≈Βψ”κΨ÷œό–‘ΓΘ
‘ΎΙΐ»ΞΒΡΦΗ °Ρξάο,¥ΈΦΕ¥ζ–Μ≤ζΈο «÷Ί“ΣΒΡ“©Έοά¥‘¥[1-2]ΓΘ”…Ζ«ΚΥΧ«ΧεκΡΚœ≥…ΟΗ¥ΏΜ·Κœ≥…ΒΡΖ«ΚΥΧ«ΧεκΡΦ“Ήε «¥ΈΦΕ¥ζ–Μ≤ζΈο÷–ΒΡ“Μάύ÷Ί“ΣΜ·ΚœΈο,“ρΤδΖαΗΜΕύ―υΒΡΜ·―ßΫαΙΙΚΆ”ΟΆΨ≥…ΈΣΩ…Ι©ΆΎΨρΒΡ“©Ωβ,»γΨΏ”–ΩΙΨζΜν–‘ΒΡ¥οΆ–ΟΙΥΊΓΔΕύπΛΨζΥΊΓΔΆρΙ≈ΟΙΥΊ,ΨΏ”–ΩΙ―ΉΚΆΟβ“Ώ“÷÷ΤΉς”ΟΒΡΜΖφΏΟΙΥΊA,ΨΏ”–ΩΙ÷ΉΝωΜν–‘ΒΡ≤©ά¥ΟΙΥΊ[3⇓-5](ΆΦ1)ΓΘ
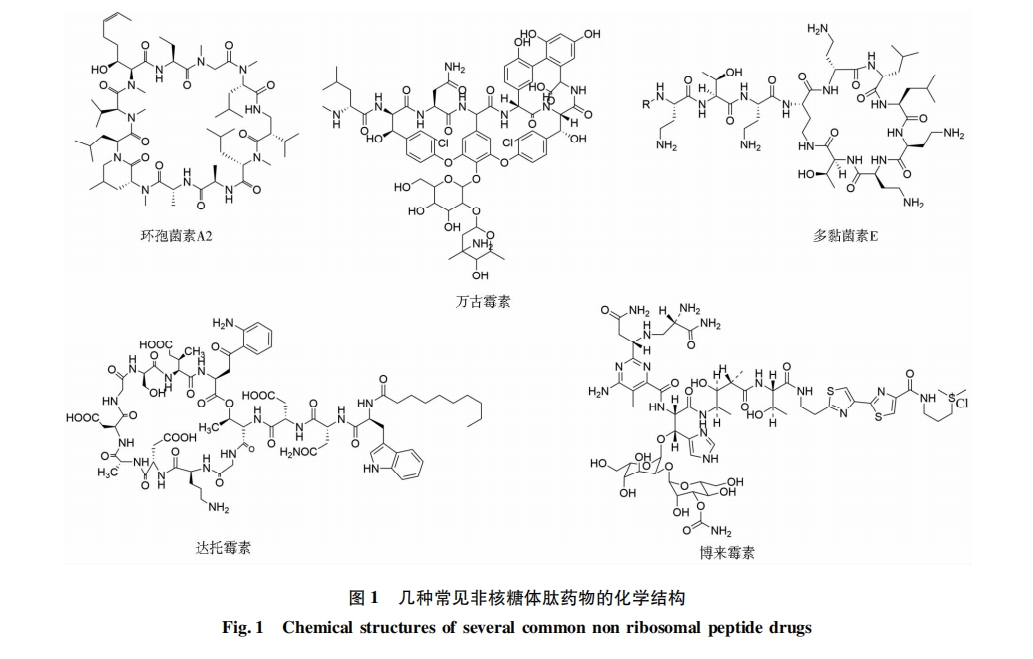
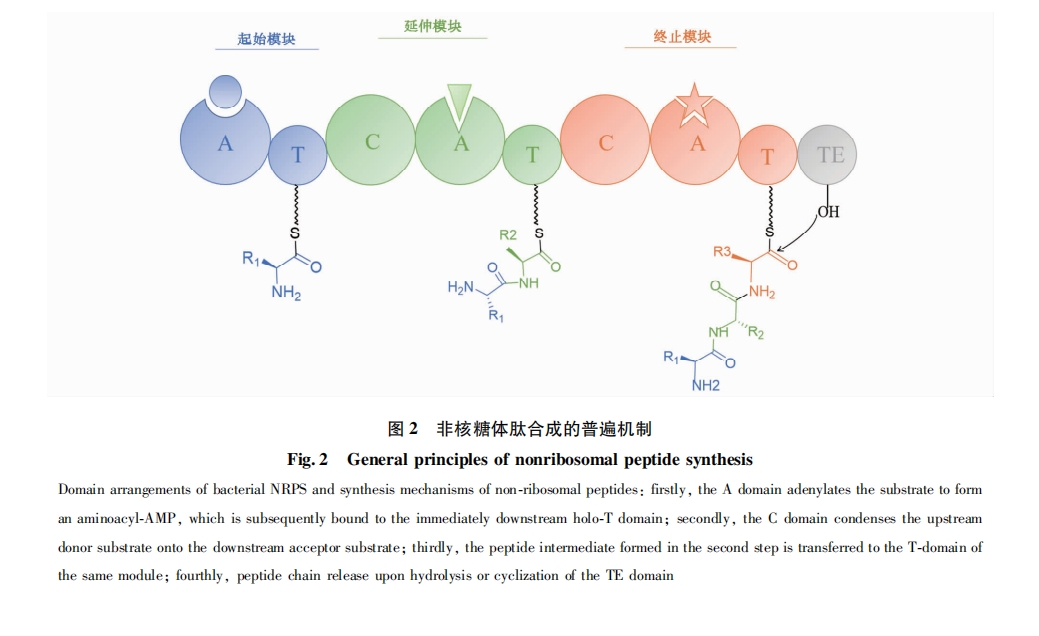
Ζ«ΚΥΧ«ΧεκΡ…ζΈοΚœ≥…Μζ÷Τ”–±π”Ύ¥ΪΆ≥ΒΡ“άάΒΚΥΧ«ΧεΚœ≥…ΒΡΒΑΑΉ÷ ΜρΕύκΡΓΘΤδ÷ς“Σ «Ά®Ιΐ“ΜΗωΨό¥σΒΡΖ«ΚΥΧ«ΧεκΡΚœ≥…ΟΗΫΪΑ±ΜυΥαœΏ–‘ΉιΉΑΤπά¥[6-7]ΓΘΖ«ΚΥΧ«ΧεκΡΚœ≥…ΟΗ «”…ΕύΗωΡΘΩιΉι≥…ΒΡΗ¥ΚœΟΗΟΗœΒ,ΟΩΗωΡΘΩιΗΚ‘π“Μ¬÷κΡΦϋΒΡ―”…λ,≤Δ«“ΡΘΩι”κΑ±ΜυΥαΒΡΉιΉΑΥ≥–ρ¥φ‘Ύ“Μ“ΜΕ‘”ΠΙΊœΒΓΘΤδ÷–ΡΘΩιΜ·¥ΏΜ·ΒΞ‘ΣΉνΜυ±ΨΒΡΙΠΡή”ρΑϋά®ΥθΚœ”ρΓΔœΌή’θΘΜ·”ρΓΔκΡΜυ‘ΊΧεΒΑΑΉΫαΙΙ”ρ(thiolation domain, T”ρ)[5,8 -9]ΓΘΖ«ΚΥΧ«ΧεκΡΚœ≥…ΟΗœΏ–‘¥ΏΜ·Κœ≥…ΕύκΡΒΡΜυ±ΨΙΐ≥Χ»γœ¬:(1) A”ρ¥”Α±ΜυΥαΒΉΈο≥Ί÷–ΧΊ“λ–‘ Ε±πΦΛΜνΑ±ΜυΥα≤ΔΫΪΑ±ΜυΥαœΌή’θΘΜ·–Έ≥…Α±θΘ-AMP[10],ΥφΚσΑ±θΘ-AMPΙ≤ΦέΫαΚœΒΫΨ≠ΝΉΥαΖΚθΘέœΜυ““ΑΖΜυΉΣ“ΤΟΗ[11-12](phosphopantetheine transferase, PPTase)ΜνΜ·ΒΡT”ρ…œ,–Έ≥…Α±θΘΝρ÷§;(2) T”ρΫΪΑ±θΘΝρ÷§ΉΣ“ΤΒΫC”ρ,‘ΎC”ρΗΟΑ±θΘΝρ÷§ΒΡ”ΈάκΑ±Μυ«ΉΚΥΫχΙΞ…œ“ΜΗω÷–ΦδΧεΒΡτ Μυ[13], ΙΤδΝρθΞΦϋΕœΝ―Εχ–Έ≥…κΡΦϋ,”…¥ΥΆξ≥…“Μ¬÷ΒΉΈοΥθΚœ;(3)ΒΎΕΰ≤Ϋ–Έ≥…ΒΡκΡΝ¥÷–ΦδΧεΉΣ“Τ÷ΝΆ§“ΜΡΘΩιΒΡT”ρ,ΉςΈΣœ¬“ΜΡΘΩιΒΡΙ©ΧεκΡΝ¥;(4)‘ΎΝρθΞΟΗΫαΙΙ”ρ(thioesterase domain,TE”ρ)ΒΡΥ°ΫβΜρΜΖΜ·Ής”Οœ¬ΕύκΡ ΆΖ≈(ΆΦ2)ΓΘ¥ΥΆβ,“Μ–©ΡΘΩι÷–ΜΙΚ§”–≤νœρ“λΙΙ”ρΓΔ―θΜ·”ρΓΔΦΉΜυΜ·”ρΒ»,‘ωΦ”ΝΥΖ«ΚΥΧ«ΧεκΡΒΡΫαΙΙΕύ―υ–‘[14]ΓΘ≥ΐ20÷÷ΒΑΑΉ÷ ‘¥Α±ΜυΥαΆβ,Ζ«ΚΥΧ«ΧεκΡΫαΙΙ÷–ΜΙΩ…≤τ»κD-Α±ΜυΥαΓΔN-ΦΉΜυΜ·Α±ΜυΥαΓΔΠ¬-Α±ΜυΥαΓΔΠΝ-τ«ΜυΥαΓΔτ»ΥαΓΔΑΖΒ»[5⇓-7]ΓΘ
Μυ”ΎNRPSœΏ–‘Κœ≥…Μζ÷ΤΚΆΡΘΩιΜ·‘≠άμ,»ΥΟ«“Μ÷±≥Δ ‘≤…”ΟΉιΚœ…ζΈοΚœ≥…ΒΡΖΫΖ®Ε‘±ύ¬κΖ«ΚΥΧ«ΧεκΡΚœ≥…ΟΗΒΡ¥ΏΜ·ΒΞ‘ΣΫχ––≤ΌΉς[15],“‘―«ΜυΫΜΜΜ[16]ΜρΡΘΩι/ΫαΙΙ”ρΫΜΜΜ[17]/‘ωΦ”[18]/Φθ…Ό[19]Β»ΖΫ ΫΗΡ±δ‘≠”–Ζ«ΚΥΧ«ΧεκΡΒΡ…ζΈοΚœ≥…ΆΨΨΕ,ΜώΒΟ–¬ΒΡΖ«ΚΥΧ«ΧεκΡάύΜ·ΚœΈοΜρΤδ―ή…ζΈοΓΘΒΪ ««ΑΤΎΒΡ―–ΨΩ ΐΨί±μΟςΥφΜζ―Α’“NRPSΫαΙΙ”ρ±ΏΫγΦδΒΡΒΑΑΉ÷ Ά§‘¥«χ”ρΫχ––ΡΘΩιΫΜΜΜΒΡ≥…ΙΠ¬ ≤Δ≤ΜΗΏ:“ΜΖΫΟφ,Α¥“ΜΑψΙφ¬…Ϋχ––ΡΘΩιΫΜΜΜΚΆ‘”Κœ,ΆυΆυΒΦ÷¬ΡΩ±ξ≤ζΈο≤ζΝΩΒΡΦ±Ψγœ¬ΫΒ;Νμ“ΜΖΫΟφ,Κœ ΒΡΫΜΜΜ±ΏΫγΫω‘ΎΒΑΑΉ÷ –ρΝ–Ά§‘¥–‘ΫœΗΏΒΡNRPS÷°ΦδΫœ“Ή―Α’“,ΨΏ”–“ΜΕ®Ψ÷œό–‘[17]ΓΘΫϋΡξά¥,NRPS¥ΏΜ·ΡΘΩι÷ΊΙΙ―–ΨΩ»ΓΒΟΝΥΫœ¥σΒΡΆΜΤΤ,Έ“Ο«‘Ύœ¬ΈΡ÷–Ε‘œύΙΊΫχ’ΙΫχ––ΝΥ÷ΊΒψ¬έ ωΓΘ
1 ΡΘΩιΧφΜΜ
Ε‘NRPSΫχ––ΗΡ‘λ÷ς“Σ”–ΝΫΗωΡ―Βψ:Τδ“Μ «ΟΗΕ‘ΒΉΈο±δΜ·ΒΡ»ί»ΧΕ»;ΤδΕΰ «‘ΎΙΠΡή”ρ÷°Φδ÷ΊΉιΤ¥ΉΑΙΐ≥Χ÷–,»ΎΚœ±ΏΫγΕ‘ΒΑΑΉ÷ ’ϊΧεΫαΙΙ≤ζ…ζΝΥ”Αœλ¥”ΕχΫΒΒΆΝΥΟΗΜνΓΘ“ρ¥Υ“Σ Ι‘”ΚœNRPSΩ… Ε±πΦ”‘ΊΒΉΈοΚœ≥…ΡΩΒΡΕύκΡΦΑΧαΗΏ‘”ΚœNRPSΒΡ¥ΏΜ·–߬ ,ΙΊΦϋ‘Ύ”ΎΧαΗΏ‘”ΚœΡΘΩιΕ‘ΒΉΈοΒΡ―Γ‘ώ–‘“‘ΦΑΫΪ«Ε»κΒΑΑΉΕ‘’ϊΧεΒΑΑΉ÷ Ω’ΦδΫαΙΙΒΡ”ΑœλΦθ÷ΝΉν–ΓΓΘΈ“Ο«»œΈΣ÷Ν…Ό–η“ΣΩΦ¬«»ΐΖΫΟφΒΡΈ Χβ:“Μ «‘”ΚœΡΘΩιά¥‘¥ΒΡ―Γ‘ώΓΘΕ‘”ΠΆ§“ΜΑ±ΜυΥα≤–Μυ»ΎΚœ≤ΜΆ§NRPSά¥‘¥ΒΡΙΠΡήΡΘΩι¥ΏΜ·–߬ «Ζώ”–≤ν“λΓΘΕΰ «‘”Κœ±ΏΫγΒΡ―Γ‘ώΓΘΩ…“‘ΗυΨίΒΑΑΉ÷ –ρΝ–ΒΡ±Θ Ί–‘≈–ΕœΟΩ“ΜΗωΫαΙΙ”ρΒΡ±ΏΫγ,Ά®Ιΐ‘≠ ΦΡΘΩι”κΧφΜΜΡΘΩιΦδΒΑΑΉ÷ –ρΝ–ΒΡ±»Ε‘‘ΎΫαΙΙ”ρΝ§Ϋ”«χ”ρ―Α’“Ά§‘¥–‘ΫœΗΏΒΡ–ρΝ–ΉςΈΣ±ΏΫγΫΎΒψΫχ––ΧφΜΜΓΘ»ΐ «ΩΦ¬«ΥθΚœ”ρΒΡΒΉΈοΧΊ“λ–‘ΓΘΥδ»ΜœΌή’θΘΜ·A”ρ «ΒΉΈοΑ±ΜυΥα―Γ‘ώΒΡΒΎ“ΜΒάΟ≈Φς,ΒΪ «¥ΏΜ·ΡΘΩι÷–ΒΡΥθΚœΫαΙΙ”ρC”ρΕ‘ΒΉΈο“≤”–―Γ‘ώΧΊ“λ–‘ΓΘΆυΆυ‘ΎΕ‘NRPSΒΡAΫαΙΙΫχ––ΧφΜΜΚσ,œΌή’θΘΜυΜ·ΒΡΑ±ΜυΥα≤ΜΡή‘ΎC”ρΫχ––ΥθΚœΖ¥”ΠΕχ ΙκΡΝ¥ΈόΖ®―”…λ,«“C”ρΕ‘ΤδœύΝΎœ¬”ΈA”ρΒΡΒΉΈο―Γ‘ώ–‘≤ζ…ζ”ΑœλΓΘ
1.1 œΌή’θΘΜ·”ρΧφΜΜ
‘ΎΖ«ΚΥΧ«ΧεκΡΉΑ≈δΙΐ≥Χ÷–, Ήœ»”…œΌή’θΘΜ·”ρ¥”Ω…άϊ”ΟΒΡΑ±ΜυΥαΒΉΈο≥Ί÷–―Γ‘ώœύΕ‘”ΠΒΡΧΊ“λΑ±ΜυΥαΚœ≥…œύ”ΠΒΡΑ±θΘ-AMP[9]ΓΘ–μΕύ―–ΨΩ±μΟς,NRPSΟΩ“Μ¥ΏΜ·ΡΘΩι―”…λΑ±ΜυΥαΒΡΧΊ“λ–‘÷ς“Σ»ΓΨω”ΎA”ρΚΆC”ρ,Τδ÷–A”ρ «ΒΉΈοΑ±ΜυΥα―Γ‘ώΒΡΒΎ“ΜΒάΟ≈ΦςΓΘA”ρœΌή’θΘΜ·Α±ΜυΥαΒΉΈοΒΡΧΊ“λ–‘ΒΦ÷¬¥ΏΜ·ΡΘΩι―”…λΑ±ΜυΥα≤–ΜυΒΡ≤ν“λΓΘ“ρ¥Υ,Ήν‘γΒΡΡΘΩιΧφΜΜΦ·÷–‘ΎA”ρΓΘStachelhausΒ»[20]”Ύ1995Ρξ‘ΎΩί≤ί―ΩφΏΗΥΨζ(Bacillus subtilis)÷–”Ο“λ‘¥A”ρΧφΜΜ÷§κΡΩΙ…ζΥΊSurfactinΚœ≥…ΟΗSrfA-C÷–Leu7ΧΊ“λ–‘A”ρ(SrfA-A7),≥…ΙΠΒΟΒΫΝΥPhe7ΓΔOrn7ΓΔCys7ΓΔVal7»Γ¥ζΒΡœΒΝ–SurfactinάύΥΤΈο (ΆΦ3a)ΓΘΥφΚσ,ΗΟΆ≈Ε”ΦΧ–χ”Ο“λ‘¥AT”ρΧφΜΜΚœ≥…ΟΗSrfA-A÷–Leu2ΧΊ“λ–‘A”ρ(SrfA-A2),ΒΟΒΫΝΥOrn2»Γ¥ζΒΡSurfactinάύΥΤΈο[21]ΓΘΥδ»Μ…œ ωA”ρΦΑAT”ρΧφΜΜΨυΒΟΒΫΝΥΡΩΒΡ≤ζΈο,ΒΪΗΟ Β―ι÷–SurfactinάύΥΤΈοΒΡ≤ζΝΩΨυΟςœ‘œ¬ΫΒΓΘ¥ΥΆβ,≤Δ≤Μ «Υυ”–ΒΡA”ρΧφΜΜΨυΡήΜώΒΟ‘ΛΤΎ≤ζΈοΓΘάΐ»γ,AckerleyΚΆLamont[22]Ζ÷±π≤…”Ο“λ‘¥ΒΡœΌή’θΘΜ·L-ThrΓΔCysΓΔValΚΆSerΒΡA”ρΧφΜΜΆ≠¬ΧΦΌΒΞΑϊΨζ(Pseudomonas aeruginosa PAO1)Ζ«ΚΥΧ«ΧεκΡΚœ≥…ΟΗΒΎ4Ηω―«ΜυPvdDΡΘΩι“ΜΒΡA”ρ(L-Thr),ΫωœΌή’θΘΜ·L-ThrΒΡA”ρΧφΜΜ”–…ΌΝΩ≤ζΈοpyoverdine≤ζ…ζ,Τδ”ύ‘”ΚœΒΡNRPSΨυΈόΙΠΡή (ΆΦ3b)ΓΘ¥Υ Β―ι÷–Ζ«ThrΧΊ“λ–‘œΌή’θΘΜ·”ρΧφΜΜΚσΈόΕύκΡ≤ζΈοΚœ≥…Ω…Ρή «”…”ΎC”ρΒΡΒΉΈοΧΊ“λ–‘ΓΘ1999ΡξBelshawΒ»[23]‘ΎΕ‘C”ρ―–ΨΩΚσΧα≥ωΦΌœκ,C”ρΒΡCΕΥ≤ΩΖ÷ΈΣ ήΧεΈΜΒψ,ΫαΚœΑ±θΘ-S-PCP;NΕΥ≤ΩΖ÷ΈΣΙ©ΧεΈΜΒψ,ΫαΚœ…œ”ΈΒΡκΡΜυ-S-PCP, ήΧεΈΜΒψΕ‘ ήΧεΒΉΈοΑ±θΘ-S-PCP”–«Ω―Γ‘ώ–‘,Ι©ΧεΈΜΒψΕ‘Ι©ΧεΒΉΈο≤ύΝ¥Α±ΜυΥαΫω”–»θ―Γ‘ώ–‘ΓΘ“ρ¥Υ‘”ΚœA”ρ Ε±πΒΡCysΓΔValΚΆSerΈόΖ®Ϋχ»κ…œ”ΈC”ρΫαΙΙ,ΫχΕχΈόΖ®”κ…œ”ΈκΡΝ¥ΥθΚœ,ΒΦ÷¬κΡΝ¥―”…λ÷–Εœ[22]ΓΘ»ΜΕχBelshawΒ»―–ΨΩΒΡ≤ΜΉψ÷°¥Π‘Ύ”ΎΫω“‘“ΜΗωΑ±ΜυΥαΦ¥Α±θΘ-S-PCPΉςΙ©ΧεΒΉΈο,ΈόΖ®÷ΛΟςC”ρΙ©ΧεΈΜΒψΕ‘ΕύκΡΒΡ―Γ‘ώ–‘,Β±Ι©ΧεΒΉΈοΈΣκΡ-S-PCP ±,Ι©ΧεΈΜΒψ―Γ‘ώ–‘≤ΔΖ«»γ¥Υ[24]ΓΘ
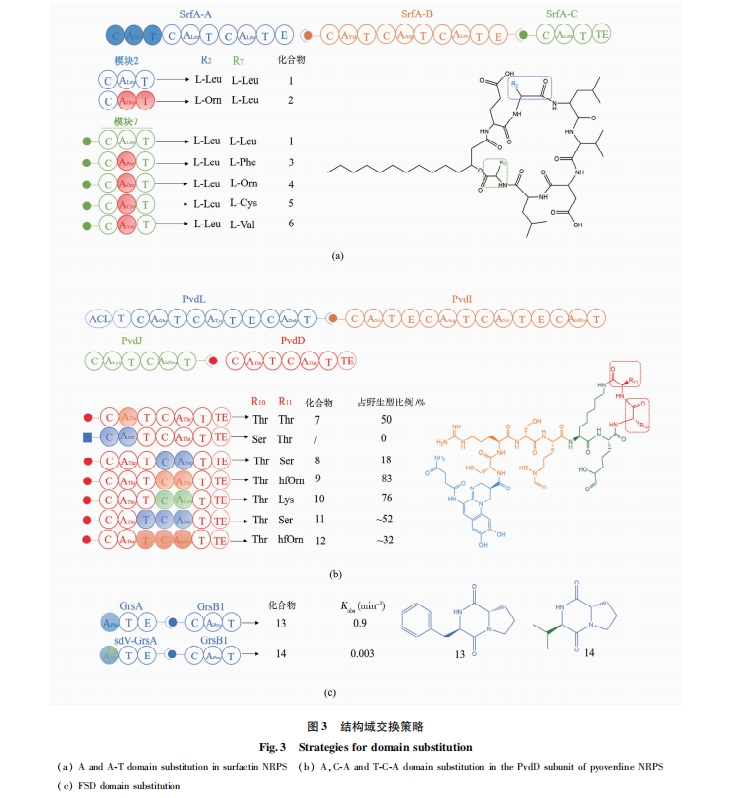
»γ«Α ω,A”ρ¥”Α±ΜυΥαΒΉΈο≥Ί÷–―Γ‘ώœύΕ‘”ΠΒΡΧΊ“λΑ±ΜυΥαΚœ≥…œύ”ΠΒΡΑ±θΘ-AMPΓΘ”–±®Βάœ‘ Ψ,AΫαΙΙ”ρ÷–”–10ΗωΖ«Ν§–χ±ύ¬κΒΡΙΊΦϋΑ±ΜυΥα≤–ΜυΨωΕ®ΝΥΤδΒΉΈο Ε±πΒΡΧΊ“λ–‘[25]ΓΘ“ρ¥Υ,≥ΐΆξ’ϊΒΡA”ρΧφΜΜΆβ,ΫϋΡξά¥”–―–ΨΩ≥Δ ‘ΧφΜΜA”ρΡΎΑϋΚ§ΙΊΦϋΜν–‘Α±ΜυΥα≤–ΜυΒΡΗΏΕ»±Θ ΊΒΡ“ΜΗωΜΤΥΊ―θΜΙΒΑΑΉ―υ―«”ρ(flavodoxin-like sub domain,FSD)[26-27]ΓΘΫΜΜΜFSDΡήΉν–ΓΒΊΤΤΜΒA”ρΒΡ’ϊΧεΫαΙΙ,Έ§≥÷ΚΆΤδΥϋΫαΙΙ”ρΦδΒΡΙΊΦϋœύΜΞΉς”Ο, Ι‘”ΚœΒΡNRPS±Θ≥÷‘≠”–¥ΏΜ·–߬ [26]ΓΘάΐ»γ,KriesΒ»[27]”Ο9Ηω≤ΜΆ§ΒΉΈοΧΊ“λ–‘ΒΡFSDΧφΜΜΕ‘±Ϋ±ϊΑ±ΥαΧΊ“λ–‘ Ε±πΒΡgramicidin SΚœ≥…ΟΗΤπ ΦΡΘΩιGrsAΒΡFSDΓΘΤδ÷–,ΧφΜΜγ”Α±ΥαΧΊ“λ–‘FSDΒΡGrsAœΌή’θΘΜ·Μν–‘«ΩΓΘ≤Δ«“‘ΎΧμΦ”Η§Α±ΥαΧΊ“λ–‘GrsB1ΡΘΩιΚσ,GrsA- GrsB1ΉςΈΣ“ΜΗωVal-ProΕΰΆΣΏΏύΚΚœ≥…ΟΗΖΔΜ”Ής”Ο,Κœ≥…ΝΥD-Val-L-ProΕΰΆΣΏΏύΚ (ΆΦ3c)ΓΘThongΒ»[28]”ΟCRISPR-Cas9Μυ“ρ±ύΦ≠ΦΦ θ,ΧφΜΜenduracidinΚœ≥…ΟΗΒΡFSD,ΒΟΒΫΝΥ“ΜœΒΝ––¬ΒΡ÷§κΡ,”–ΒΡ≤ζΝΩΫ”Ϋϋ“Α…ζ–ΆΨζ÷ξΓΘ
1.2 ΡΘΩι’ϊΧεΧφΜΜ
‘ΎNRPS÷–≤…”ΟΡΘΩι’ϊΧεΧφΜΜΡήΙΜ±Θ÷Λ¥ΏΜ·ΒΞ‘ΣΡΘΩιΫαΙΙΚΆΙΠΡήΒΡΆξ’ϊ–‘ΓΘάΐ»γ,”Ο÷§κΡlichenysin…ζΈοΚœ≥…Μυ“ρ¥Ί÷–ΒΡGln Ε±πΡΘΩιCATΫΜΜΜsurfactinΚœ≥…Μυ“ρ¥Ί÷–ΒΡGluΡΘΩι,ΒΟΒΫΝΥΗΏ≤ζΓΔΗΏΜν–‘ΒΡ–¬–Ά÷§κΡ[29](ΆΦ4a)ΓΘ”Ο¥οΆ–ΟΙΥΊNRPS÷–ΡΘΩι11CASerTΫΜΜΜΡΘΩι8 CAAlaT,ΒΟΒΫ8Κ≈ΈΜD-Ser»Γ¥ζΒΡ¥οΆ–ΟΙΥΊάύΥΤΈο;”ΟΡΘΩι8 CAAlaTΧφΜΜΡΘΩι11 CASerT,ΒΟΒΫ11Κ≈ΈΜD-Ala»Γ¥ζΒΡ¥οΆ–ΟΙΥΊάύΥΤΈο[17]ΓΘ’βΝΫ÷÷–¬Μ·ΚœΈοΨυΕ‘ΫπΜΤ…ΪΤœΧ―«ρΨζΨΏ”–“©άμΜν–‘,ΒΪ≤ζΝΩΫωΈΣ“Α…ζ–ΆΨζ÷ξΒΡ15%ΚΆ45%Ήσ”“ΓΘ”–―–ΨΩ“‘T-C”ρΦδΒΡLGG(H/D)S(I/L)±Θ Ί–ρΝ–ΈΣ»ΎΚœΈΜΒψ,ΫΪ“ΜΗωΗΤάκΉ”“άάΒΒΡ÷§κΡΩΙ…ζΥΊ(calcium dependent antibiotic,CDA)Κœ≥…ΟΗΡΘΩι13 CATrpTΚΆA54145 NRPSΡΘΩι13 CAIleTΧφΜΜ¥οΆ–ΟΙΥΊNRPSΡΘΩι13CAKynT,Ζ÷±πΜώΒΟΝΥ13ΈΜTrpΚΆIle»Γ¥ζΒΡ¥οΆ–ΟΙΥΊάύΥΤΈο,«“Ε‘‘”ΚœNRPS¥ΏΜ·–߬ ”ΑœλΫœ–Γ,–¬Μ·ΚœΈο≤ζΝΩΉνΗΏΫ”Ϋϋ‘≠≤ζΝΩΒΡ67%[30]ΓΘ
1.3 CA”ρΧφΜΜ
‘Ύ1.1Υυ ω“λ‘¥A”ρΧφΜΜPvdDΡΘΩι“ΜL-Thr A”ρΒΡΆ§ ±, AckerleyΚΆLamont[22]”Ο“λ‘¥CA”ρ(Αϋά®L-Υ’Α±ΥαΧΊ“λΒΡCA”ρ)ΧφΜΜPvdDΡΘΩι“ΜCA”ρ,ΒΪ“βΆβΒΡ «Υυ”–‘”ΚœΟΗΨυΈόΜν–‘ΓΘCA”ρΫΜΜΜ ßΑήΒΡ‘≠“ρ”–:(1)‘”ΚœΒΡ“λ‘¥CA”ρΈόΖ®”κPvdDΡΘΩι“ΜΒΡΧλ»ΜT”ρ”––ßΜΞΉς,ΒΦ÷¬‘”ΚœΟΗΈόΜν–‘;(2)œ÷”–άμ¬έœ‘ Ψ,―«ΜυΦδΒΡΕ‘Ϋ””ρΩ…÷ΗΒΦΖ«ΚΥΧ«ΧεκΡΚœ≥…ΟΗ’ΐ»ΖΒΡœΏ–‘≈≈Ν–,≤ΔΫιΒΦ―«ΜυΦδΒΡΜΞΉς,ΗΟ Β―ι÷–PvdDΡΘΩι“ΜΒΡCA”ρΧφΜΜΚσΈόΖ®”κ…œ”Έ―«ΜυPvdJ’ΐ»ΖΕ‘Ϋ”[31⇓-33](ΆΦ3b)ΓΘ
TanovicΒ»[34]‘ΎΕ‘Ωί≤ί―ΩφΏΗΥΨζ÷–ΒΡ÷§κΡΩΙ…ζΥΊΚœ≥…ΟΗΉνΡ©ΕΥΡΘΩιSrfA-CΒΡΫαΙΙ―–ΨΩΚσΖΔœ÷,ΤδΥθΚœ”ρ(C”ρ)ΚΆœΌή’θΘΜ·”ρ(A”ρ)÷°Φδ”–“ΜΕΈΚ§32ΗωΑ±ΜυΥαΒΡΝ§Ϋ”«χ,‘ΎSrfA-CΨßΧεΫαΙΙ÷–,ΗΟΝ§Ϋ”«χ”κC”ρΓΔA”ρΨυΫτΟήœύΝ§ΓΘ“ρ¥Υ,»ΥΟ«»œΈΣ‘ΎΡΘΩιΧφΜΜ ±–η±Θ≥÷CA”ρΫαΙΙΆξ’ϊ,C”ρ”κA”ρ”ΠΉςΈΣ≤ΜΩ…Ζ÷ΗνΒΡ’ϊΧε≤≈Ρή±ήΟβΤΤΜΒ‘”ΚœΟΗΒΡΫαΙΙΚΆΜν–‘ΓΘΜυ”Ύ¥Υœ÷œσ,CalcottΒ»[35]ΦΧ–χ”Ο“λ‘¥CA”ρΧφΜΜPvdD―«ΜυΒΎΕΰΗωΡΘΩιCA”ρΓΘ”κΗΟΆ≈Ε”‘γΤΎΧφΜΜPvdD―«ΜυΒΎ“ΜΗωΡΘΩιCA”ρ≤ΜΆ§,ΧφΜΜPvdD―«ΜυΒΎΕΰΗωΡΘΩιCA”ρΩ…±ήΟβΤΤΜΒPvdD”κ…œ”Έ―«ΜυPvdJΦδΒΡœύΜΞΉς”Ο(ΆΦ3b)ΓΘΫαΙϊœ‘ Ψ,”–ΝΫΗω‘”ΚœNRPSΨΏ”–ΫœΚΟΒΡ¥ΏΜ·Μν–‘,≤ΔΜώΒΟΝΥΝΫ÷÷Thr11ΚΆLys11»Γ¥ζΒΡpyoverdineάύΥΤΈο,≤ζΝΩΖ÷±π «“Α…ζ–ΆΒΡ83% ΚΆ76%,Τδ”ύ6Ηω‘”ΚœΟΗΨυΈόΜν–‘[35]ΓΘ
2 »ΎΚœ±ΏΫγ
‘Ύ≥θΤΎ―–ΨΩ÷–,¥”±Θ÷ΛΡΘΩιΙΠΡήΆξ’ϊ–‘ΒΡΫ«Ε»ΩΦ¬«,―–ΨΩ’ΏΆ®≥Θ“‘AΫαΙΙ”ρΜρ“ΜΗωΆξ’ϊΒΡCAT(E)ΡΘΩιΒΞ‘ΣΕ‘NRPSΫχ––ΧφΜΜΒ»ΗΡ‘λ[27-28,36]ΓΘάΐ»γ,“‘T-C”ρΦδΝ§Ϋ”«χΈΣ»ΎΚœ±ΏΫγΧφΜΜ¥οΆ–ΟΙΥΊΖ«ΚΥΧ«ΧεκΡΚœ≥…ΟΗΒΡΆξ’ϊCATΡΘΩι[30]ΓΘ»ΜΕχΆξ’ϊCAT(E)ΡΘΩιΫΜΜΜ¥φ‘Ύ≤ζΝΩΦ±Ψγœ¬ΫΒΒΡΈ Χβ,«““Σ«σΒΑΑΉ÷ –ρΝ–ΨΏ”–ΗΏΆ§‘¥–‘,”–“ΜΕ®ΒΡΨ÷œό–‘ΓΘάμΫβΫαΙΙ”ρΦδΒΡΝ§Ϋ”«χ‘ΎΖ«ΚΥΧ«ΧεκΡΚœ≥…ΟΗ¥ΏΜ·ΒΉΈοΜνΜ·ΓΔΦ”‘ΊΓΔΥθΚœΙΐ≥Χ÷–ΒΡΉς”Ο,Ε‘”ΎΆ®ΙΐΡΘΩιΧφΜΜΜώ»ΓΫœΚΟ¥ΏΜ·Μν–‘ΒΡ‘”ΚœNRPS”–÷Ί“ΣΉς”ΟΓΘ
2.1 “‘A-T”ρΦδΝ§Ϋ”«χΈΣ»ΎΚœ±ΏΫγ
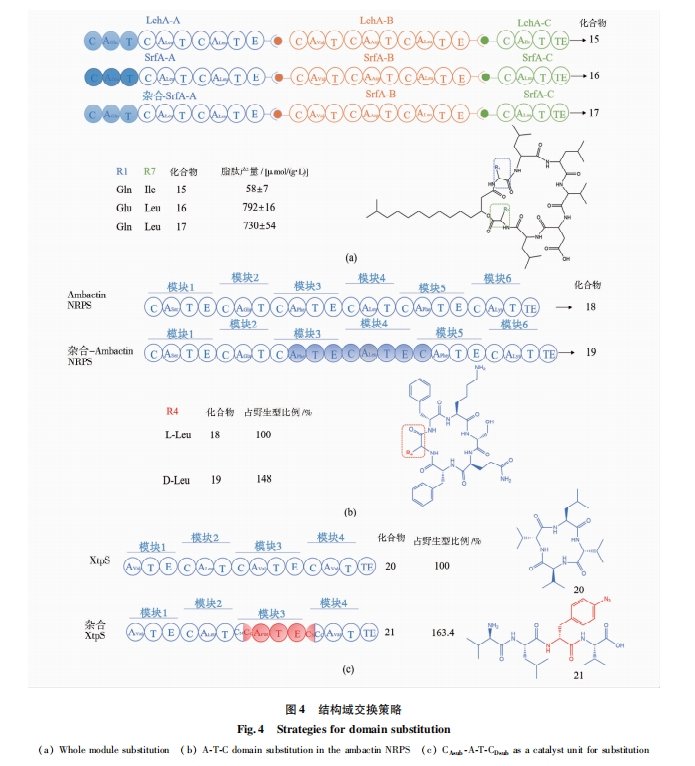
MillerΒ»[37]Ά®Ιΐ…ζΈο–≈œΔ―ßΖ÷ΈωΝΥ≤ΜΆ§NRPSΒΡA-T”ρΦδΝ§Ϋ”«χΚσ,‘ΎA-T”ρΒΡΝ§Ϋ”«χΖΔœ÷ΝΥ“ΜΕΈ±Θ ΊΒΡLPxP–ρΝ–,ΗΟLPxP–ρΝ–”κA”ρCΕΥΒΡ―«”ρœύΜΞΉς”Ο“‘Έ»Ε®A10Μυ–ρ÷–ΒΡ±Θ ΊάΒΑ±Υα;¥ΥΆβ,ΗΟΜυ–ρ”κA”ρCΕΥΒΡ―«”ρœύΜΞΉς”ΟΩ…ΡήΜα–≠ΒςPCP”ρΒΡ‘ΥΕ·”κA”ρΒΡΙΙœσ±δΜ·ΓΘΫΪLPxP–ρΝ–÷–ΒΡPro961ΆΜ±δΚσΒΦ÷¬≤ζΈοΚœ≥…ΥΌ¬ œ¬ΫΒΫϋ“ΜΑκ[37]ΓΘΆ§ ±”–―–ΨΩΫΪΖΔΙβΗΥΨζ(Photorhabdus luminescens)÷–ΒΞΡΘΩιΒεάΕΚœ≥…ΟΗIndCΒΡT”ρ”Οά¥Ή‘Ν¥ΟΙΨζΒΡBpsAΟΗΒΡT”ρΧφΜΜΚσ,ΤΤΜΒΝΥΒεάΕΚœ≥…;Νν»ΥΨΣΤφΒΡ «,ΫΪBpsAΟΗΒΡT”ρ”κA-T”ρΦδΒΡΝ§Ϋ”«χ“ΜΤπΫΜΜΜ ±Μ÷Η¥ΝΥΒεάΕΚœ≥…[38]ΓΘ…œ ω Β―ι±μΟς―Γ‘ώ≤Δ±ΘΝτΚœ ΒΡΫαΙΙ”ρΦδΝ§Ϋ”«χΕ‘Ρ≥–©ΡΘΩιΧφΜΜΜώΒΟ”–Μν–‘ΒΡ‘”ΚœNRPS÷ΝΙΊ÷Ί“ΣΓΘCalcottΒ»[39]‘χ”ΟCA”ρΧφΜΜPvdDΒΎΕΰΗωΡΘΩι(1.3),»ΜΕχΫω…Ό ΐ‘”ΚœNRPSΨΏ”–ΫœΚΟ¥ΏΜ·Μν–‘,¥σΕύ ΐ‘”ΚœNRPSΈόΙΠΡή[35]ΓΘΥφΚσ,ΗΟΆ≈Ε”“‘A-TΦδΝ§Ϋ”«χΈΣ»ΎΚœ±ΏΫγΧφΜΜΚœ≥…ΟΗPvdD÷–T-C-AThr11¥ΏΜ·ΒΞ‘Σ(ΆΦ3b)ΓΘΥδ»Μ≥…ΙΠΜώΒΟΝΥΝΫ÷÷Ser11ΚΆfhOrn11»Γ¥ζΒΡpyoverdine―ή…ζΈο,ΒΪ «’βΝΫΗω―ή…ζΈοΒΡ≤ζΝΩΕΦΫœΒΆΓΘ
2.2 “‘C-AΦδ±Θ Ί–ρΝ–ΈΣ»ΎΚœ±ΏΫγΧφΜΜA-T-C”ρ
ΫϋΡξά¥Β¬ΙζΖ®άΦΩΥΗΘ¥σ―ßBodeΆ≈Ε”ΖΔœ÷CAΫαΙΙ”ρ÷°ΦδΒΡΝ§Ϋ”«χΚ§“ΜΕΈ±Θ Ί–ρΝ–LLLxxWNxTΓΘάϊ”Ο¥Υ±ΏΫγΫΎΒψ,‘ΎNRPSΝΫΗωΡΘΩι(CAT-CAT)÷°Φδ―Γ‘ώAT-C«χ”ρΈΣΧφΜΜΒΞ‘Σ(exchange unit,XU)Ϋχ––ΡΘΩιΧφΜΜΡήΜώΒΟΜν–‘ΫœΗΏΒΡ‘”ΚœNRPSΒΑΑΉ,Ρ≥–©ΧφΜΜΉιΚœΖ«ΚΥΧ«ΧεκΡΒΡ≤ζΝΩ…θ÷Ν±»“Α…ζ–ΆΧαΗΏΝΥ48%[40](ΆΦ4b)ΓΘ“‘A-T-CΫαΙΙ”ρΉςΈΣΫΜΜΜΒΞ‘ΣΫχ––ΡΘΩιΧφΜΜΚΆΉιΚœΥδ»Μ»ΓΒΟΝΥ÷Ί¥σΆΜΤΤ,Εχ«“Ω…“‘Ϋχ––ΕύΡΘΩιΆ§ ±ΧφΜΜΚΆΉιΉΑ,ΒΪ’βΗωΖΫΖ®Ψ÷œό–‘Οςœ‘: Ήœ»,CAΫαΙΙ”ρ÷°ΦδΒΡ±Θ Ί–ρΝ–LLLxxWNxT‘ΎΚήΕύNRPSΒΑΑΉ–ρΝ–÷–≤Δ≤Μ¥φ‘ΎΓΘΤδ¥Έ,œ÷”–άμ¬έœ‘ ΨC”ρΕ‘Ϋχ»κΗΟΫαΙΙ”ρΒΡΑ±θΘ-S-PCP ήΧεΨΏ”–ΚήΗΏΒΡΒΉΈο―Γ‘ώ–‘,Β±‘”ΚœΒΡA”ρ Ε±πΒΡΑ±ΜυΥα”κ…œ”ΈΒΡC”ρ≤ΜΤΞ≈δ ±,κΡΝ¥ΈόΖ®―”…λΒΦ÷¬ΈόΡΩ±ξ≤ζΈο≤ζ…ζΓΘ“ρ¥Υ,‘ΎΕ‘NRPSΫχ––ΗΡ‘λ ±,≤ΜΫω–η“ΣΩΦ¬«‘”ΚœNRPSΒΡ¥ΏΜ·–߬ ,ΜΙ–η“ΣΩΦ¬«C”ρΕ‘ΒΉΈοΒΡ―Γ‘ώ–‘,±ήΟβ‘”ΚœΒΡA”ρ Ε±πΒΡΑ±ΜυΥα”κ…œ”ΈC”ρ≤ΜΤΞ≈δ ±κΡΝ¥ΈόΖ®―”…λΒΦ÷¬ΈόΡΩ±ξ≤ζΈο≤ζ…ζΓΘ
2.3 ΧφΜΜCAsub-A-T-CDsub¥ΏΜ·ΒΞ‘Σ
Ά®ΙΐΕ‘C”ρΒΡΒΑΑΉ÷ ΫαΙΙΖ÷Έω,―–ΨΩ»Υ‘±ΖΔœ÷C”ρΒΡNΕΥ «ΧαΙ©κΡΝ¥ΒΡΙ©Χε”ρ(CDsub),CΕΥ « ήΧε”ρ(CAsub),ΥϋΟ«Ζ÷±πΫ” ήκΡΜυ-S-PCPΚΆΑ±θΘ-S-PCPΓΘΤδ÷– ήΧε”ρ(CAsub)Ε‘Α±θΘ-S-PCP”–ΧΊ“λ Ε±πΉς”Ο[41]ΓΘάΐ»γ,KaniusaiteΒ»[42⇓-44]Ε‘Χ«κΡάύΩΙ…ζΥΊ(glycopeptide antibiotics,GPAs)NRPSΒΡΗΡ‘λ Β―ι“≤±μΟς,C”ρΕ‘ ήΧεΒΉΈοΑ±θΘ-S-PCP”–«Ω―Γ‘ώ–‘ΓΘ“≤”–―–ΨΩœ‘ Ψ,C”ρ≤ΜΫωΕ‘ ήΧεΒΉΈοΑ±θΘ-S-PCP”–―Γ‘ώ–‘,Ε‘Ι©ΧεΒΉΈοκΡΜυ-S-PCPΒΡ¥σ–ΓΚΆΉι≥…“≤”–―Γ‘ώ–‘[24,45⇓ -47]ΓΘΕ‘C”ρΒΡΫαΙΙΫβΈω“≤ΖΔœ÷ΤδΨΏ”–ΝΔΧεΚΆ≤ύΝ¥―Γ‘ώ–‘[48]ΓΘΜυ”Ύ¥Υάμ¬έBozh®Ιy®ΙkΒ»[49]”Ύ2019ΡξΫχ“Μ≤ΫΩΣΖΔΝΥΒΎΕΰ¥ζΒΡΧφΜΜΖΫΑΗ:”ΟCAsub-A-T-CDsubΉςΈΣΜυ±ΨΒΡΧφΜΜΒΞ‘Σ(exchange unit condensation domain,XUC)Ϋχ––’ϊΧεΧφΜΜΓΘΗΟΖΫΑΗ±ήΟβΝΥ‘”ΚœA”ρΚΆC”ρ÷°ΦδΑ±ΜυΥαΒΉΈο Ε±π≤ΜΤΞ≈δΒΡΈ Χβ,Οςœ‘ΨΏ”–ΗϋΙψΒΡ ”ΟΖΕΈß,«“ΗΟ¥ΏΜ·ΒΞ‘Σ±ήΟβΝΥΤΤΜΒΉν÷ς“ΣΒΡΫαΙΙ”ρ-ΫαΙΙ”ρœύΜΞΉς”Ο[50],”–ΒΡΕύκΡ≤ζΝΩ±»“Α…ζ–ΆΧαΗΏΝΥ63.4%[49](ΆΦ4c)ΓΘάϊ”ΟΗΟΖΫΖ®Ω…“‘ΙΙΫ®Εύ÷÷ΥφΜζΉιΚœΒΡάύΥΤΧλ»ΜΒΡΖ«ΚΥΧ«ΧεκΡΩβΓΘ»ΜΕχCAsub-A-T-CDsubΉςΈΣΫΜΜΜΒΞ‘ΣΒΡΨ÷œό–‘‘Ύ”Ύ:ά¥‘¥”ΎΆ§ τΒΡΡΘΩιΦδ“Ή»ΎΚœ,Εχά¥‘¥”Ύ≤ΜΆ§ τΒΡΡΘΩιΦδ»±ΖΠΦφ»ί–‘;TEΫαΙΙ”ρΕ‘Α±ΜυΥαΈΜ÷ΟΓΔκΡΝ¥≥ΛΕ»ΜρΜΖκΡ¥σ–ΓΒΡΧΊ“λ–‘œό÷ΤΝΥ»Ϊ–¬ΒΡΖ«Χλ»ΜΓΔΖ«ΚΥΧ«ΧεκΡ“‘ΦΑΜΖκΡΓΔΥθκΡ–Έ≥…[51]ΓΘ
2.4 άϊ”ΟΚœ≥…ά≠Ν¥‘ΎC-A±ΏΫγ¥ΠΗΏ–ß‘”Κœ¥ΏΜ·ΡΘΩι
ΨΓΙή“‘CAsub-A-T-CDsubΈΣΧφΜΜΒΞ‘ΣΩ…“‘ΙΙΫ®ΫαΙΙΕύ―υΒΡΖ«ΚΥΧ«ΧεκΡΩβ,ΒΪ «NRPSΒΡΨό–ΆΫαΙΙΚΆ±Θ ΊΒΡ÷ΊΗ¥–ρΝ– ΙΒΟΕ‘ΤδΫχ––Μυ“ρ≤ΌΉς±»ΫœΗ¥‘”ΚΆάßΡ―;¥ΥΆβ,Ε‘ΙΠΡή”ρΫαΙΙΝΥΫβ≤ΜΉψ”– ±ΜαΒΦ÷¬»ΥΟ«‘ΎΕ‘NRPSΫχ––¥ΏΜ·ΒΞ‘ΣΧφΜΜ÷ΊΉι ±ΤΤΜΒΝΥ”ρΦδΒΡΕ‘Ϋ””ρ,“≤œό÷ΤΝΥΧφΜΜΕ‘Ϋ””ρ’β“ΜΖΫΑΗ[40,52 -53]ΓΘ2021Ρξ,ReinkeΒ»[54]ΚΆThompsonΒ»[55]”Ο“ΜΕ‘»ΥΙΛΚœ≥…ΒΡΕΧκΡΉςΈΣΓΑά≠Ν¥Γ±(synthetic zippers,ZPs)Χφ¥ζNRPSΫαΙΙ”ρΦδΒΡΕ‘Ϋ””ρ,ΫιΒΦ‘”Κœ¥ΏΜ·ΡΘΩιΦδΒΡœύΜΞΉς”ΟΓΘ“ΜΕ‘ά≠Ν¥“‘Ζ«Ι≤ΦέΉς”ΟœύΜΞΫαΚœ,≤Δ“‘C-AΦδΒΡ±Θ Ί–ρΝ–LLLxxW]-[NxTΉςΈΣ≤ε»κΈΜΒψ,”…¥ΥΫιΒΦΝΫΗω―«ΜυΜρ¥ΏΜ·ΡΘΩιΦδΒΡΫαΚœΓΘBozhueyuekΒ»[56]”Ο¥ΥΖΫΖ®Κœ≥…ΝΥ¥σΝΩΕύκΡ,”–ΒΡΉιΚœΖ«ΚΥΧ«ΧεκΡΒΡ≤ζΝΩ…θ÷Ν±»Τ’Ά®»ΎΚœΖΫ ΫΧαΗΏΝΥ8±Ε (ΆΦ5b)ΓΘ
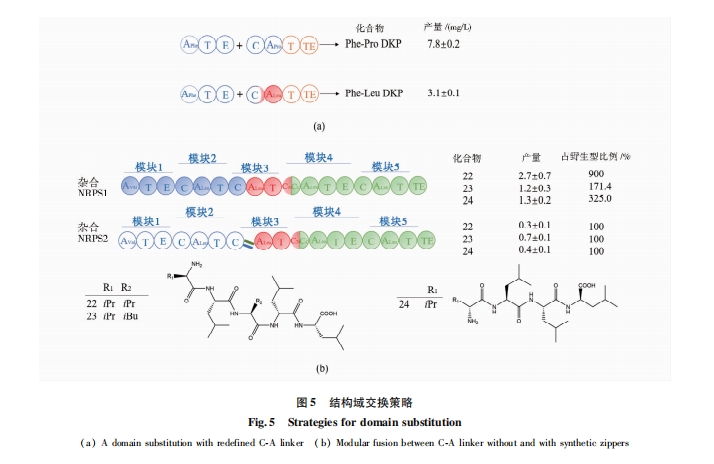
¥ΥΖΫΖ®±ήΟβΝΥΩΥ¬ΓΚΆΖ«ΚΥΧ«ΧεκΡΚœ≥…ΟΗ¥σ–ΓΒΡœό÷Τ,Ω…ΥφΜζΉιΚœ»Έ“βΒΡNRPSΡΘΩι,ΈΣΩλΥΌ Βœ÷–¬ΗΏΆ®ΝΩ…ζΈοΉιΚœΖΫΖ®ΓΔΩΣΖΔΕύκΡάύ“©ΈοΤΧΤΫΝΥΒά¬ΖΓΘ
3 C”ρ”κA”ρΒΡœύΜΞΉς”Ο
3.1 C”ρΗΡ±δA”ρΒΡΜν–‘ΚΆ―Γ‘ώ–‘
¥”…œ ωΨ≠ΒδΒΡNRPS…ζΈοΚœ≥…άμ¬έΩ…÷Σ,A”ρΧΊ“λ–‘ Ε±πΑ±ΜυΥαΒΉΈοΫχ––œΌή’θΘΜυΜ·,C”ρ‘ΎΑ±ΜυΥαΥθΚœΙΐ≥ΧΕ‘ ήΧεΑ±θΘ-S-PCPΒΡ÷÷άύΫχ“Μ≤Ϋ–ΘΕ‘―ι÷Λ(proofreading),A”ρΚΆC”ρ‘ΎΒΉΈο―Γ‘ώΙΐ≥Χ÷–ΗςΉ‘ΕάΝΔΖΔΜ”Ής”Ο,Ι≤Ά§»Ζ±ΘΥυΚœ≥…Ζ«ΚΥΧ«ΧεκΡΫαΙΙΉ®“ΜΓΘΒΪΫϋΡξΒΡ―–ΨΩ±®Βάœ‘ ΨC”ρΕ‘ΤδœύΝΎœ¬”ΈA”ρΒΡΒΉΈο―Γ‘ώ–‘≤ζ…ζ”ΑœλΓΘMayerΒ»[57]ΫΪA”ρΫχ––ΒΞΕά±μ¥οΚσ,ΥϋΩ…“‘œΌή’θΘΜυΜ·Εύ÷÷Α±ΜυΥαΒΉΈο,ΒΪΚΆC”ρΝΣΚœ±μ¥οΚσ,A”ρ÷ΜΕ‘ΨΪΑ±ΥαΨΏ”–œΌή’θΘΜυΜ·Μν–‘;ΒΞΕά±μ¥οA”ρ ±Ε‘Χλ»ΜΒΉΈοά“Α±ΥαΚΆ…ΪΑ±ΥαΨυΈόΜν–‘,”κC”ρΝΣΚœ±μ¥οΚσΕ‘’βΝΫ÷÷Α±ΜυΥαΨΏ”–œΌή’θΘΜυΜ·Μν–‘ΓΘ±μΟςC”ρ”κA”ρΒΡœύΜΞΉς”ΟΗΡ±δΝΥA”ρΒΡΒΉΈοΧΊ“λ–‘,«“Ρ≥–©A”ρ÷Μ”–‘ΎC”ρ¥φ‘ΎΒΡΧθΦΰœ¬≤≈ΨΏ”–Μν–‘ΓΘ
¥ΥΆβ,ΚήΕύΖ«÷§κΡάύNRPSΒΡΤπ ΦΡΘΩι≤Δ≤ΜΚ§”–C”ρ,Τπ ΦΡΘΩιΒΡA”ρœΌή’θΘΜυΜ·Α±ΜυΥαΚσΆ®ΙΐT”ρ÷±Ϋ”ΉΣ“ΤΒΫœ¬“ΜΡΘΩιΒΡC”ρΫχ––Α±ΜυΥαΥθΚœΓΘΒΪ «»τ≤…”ΟNRPS―”…λΡΘΩιA”ρ÷±Ϋ”Χφ¥ζΤπ ΦΒΞ‘ΣA”ρ,ΡΩ«Α…–Έ¥ΦϊκΡΝ¥―”…λ≥…ΙΠΒΡ±®ΒάΓΘΫϋΤΎBozh®Ιy®ΙkΒ»[49]ΫΪC”ρ(ΜρC”ρCΕΥΒΡ ήΧε”ρCAsub)ΚΆ―”…λΡΘΩιA”ρΙ≤±μ¥ο ±,≥…ΙΠΒΊΫΪ―”…λΒΞ‘ΣΒΡA”ρΉΣ±δΈΣΤπ ΦΒΞ‘ΣA”ρ,≤ζ…ζΝΥΡΩ±ξΖ«ΚΥΧ«ΧεκΡΜ·ΚœΈοΓΘ
”–―–ΨΩ‘ΎΫΜΜΜAT”ρ ±Α―C”ρCΕΥΉςΈΣ‘”ΚœΈΜΒψ,‘”ΚœΒΑΑΉΨΏ”–ΫœΗΏ≤ζΝΩ[58]ΓΘ”κΉν‘γΒΡAT”ρΧφΜΜΆ®≥ΘΒΦ÷¬≤ζΝΩœ¬ΫΒ[21,59 -60]œύΖ¥,ΗΟ Β―ι÷–AT”ρΒΡ≥…ΙΠΫΜΜΜ±μΟςC-AΦδΒΡœύΜΞΉς”ΟΫγΟφΕ‘A”ρΖΔΜ”Μν–‘ΙΠΡήΚή÷Ί“Σ,Β±‘”ΚœΒΡ“λ‘¥A”ρΜρAT”ρΜρAT-C”ρ≤ΜΑϋΚ§¥ΥœύΜΞΉς”ΟΫγΟφ ±,“λ‘¥A”ρΈόΖ®”κ…œ”ΈC”ρ’ΐ»ΖΜΞΉς,“ρ¥ΥA”ρΜν–‘…Ξ ß,‘”ΚœNRPSΈόΖ®ΖΔΜ”ΙΠΡήΓΘ…œ ω Β―ι±μΟς‘ΎNRPS÷–ΟΩ“ΜΉι¥ΏΜ·ΡΘΩιΒΡC”ρΚΆA”ρ≤ΔΖ«ΗςΉ‘ΕάΝΔΖΔΜ”Ής”ΟΆξ≥…“Μ¬÷Α±ΜυΥα―”…λ,C”ρ”κA”ρΖΔ…ζΫΜΜΞΉς”ΟΆξ…ΤΝΥA”ρœΌή’θΘΜυΜ·ΒΡΙΠΡήΓΘ
3.2 ΙΊ”ΎC”ρΒΉΈο―Γ‘ώ–‘ΒΡ’υ“ι
Ϋϋ20Ρξά¥,―–ΨΩ’ΏΟ«Τ’±ι»œΈΣA”ρΚΆC”ρ‘ΎΒΉΈο―Γ‘ώΚΆΓΑ–Θ’ΐΓ±÷–ΕΦΤπΉ≈ΙΊΦϋΉς”Ο,’β“ΜΦΌΥΒΦΪ¥σΒΊΗ¥‘”ΝΥNRPSΒΡάμ–‘Μ·ΉιΚœ…ζΈοΚœ≥…ΓΘΆ®Ιΐ…ζΈο–≈œΔ―ßΖ÷ΈωA”ρΒΡΒΑΑΉ÷ –ρΝ–,ΖΔœ÷ΤδΕ‘Α±ΜυΥαΒΉΈοΒΡ―Γ‘ώ”–Κή¥σ±Θ Ί–‘,…θ÷ΝΩ…“‘Ά®ΙΐA”ρΒΑΑΉ÷ –ρΝ–¥σ÷¬‘Λ≤βΤδΑ±ΜυΥαΒΉΈοΒΡ÷÷άύΓΘΒΪ «C”ρΒΡΒΑΑΉ÷ –ρΝ–Ε‘”ΠœύΆ§Α±θΘ-S-PCPΒΉΈοΈόΧΊ βΙφ¬…Μρ±Θ Ί–‘ΓΘ‘ΎΧλάΕ…ΪΝ¥ΟΙΨζ(Streptomyces coelicolor)÷–“ΜΗωΗΤάκΉ”“άάΒΒΡ÷§κΡΩΙ…ζΥΊΚœ≥…ΟΗΒΡΒΎ“ΜΗωC”ρ(CDA-C1)ΗΚ‘π¥ΏΜ·Ι©Χε2,3-epoxyhexanoyl ACP”κΥΩΑ±Υα-S-PCPΒΡΥθΚœΓΘΒΪ «Φ”ΡΟ¥σ¬σΦΣΕϊ¥σ―ßMartin SchmeingΩΈΧβΉιΆ®ΙΐΈΙ―χΤδΥϊ ήΧεΒΉΈο(±ϊΑ±Υα-,ΝΝΑ±Υα-,ΦΉΝρΑ±Υα-S-PCP) ±ΖΔœ÷,’β–© ήΧεΒΉΈο“≤Ω…“‘”κΙ©Χε2,3-epoxyhexanoyl ACPΫχ––ΥθΚœ[61]ΓΘ¥ΥΆβ,SchoppetΒ»[62] ΒΡΉν–¬―–ΨΩΫαΙϊ“≤±μΟςC”ρΕ‘ΒΉΈοΨΏ”–ΩμΖΚΒΡ―Γ‘ώ–‘ΦΑΝΔΧεΫαΙΙΒΡ»ί»Χ–‘ΓΘCalcottΒ»[63]ΒΡ―–ΨΩ“≤Ε‘C”ρΒΡ ήΧεΒΉΈοΧΊ“λ–‘’β“Μάμ¬έΧα≥ωΝΥ÷ “…ΚΆΖ¥≤ΒΓΘAckerleyΒ»[22]ΚΆCalcottΒ»[35]Ά®Ιΐ―Γ‘ώ”––ßΒΡ÷ΊΉι±ΏΫγ,‘Ύpyoverdine NRPS÷–ΫΜΜΜA”ρΒΡ≥…ΙΠ¬ ΚήΗΏ,pyoverdineάύΥΤΈοΒΡ≤ζΝΩ”≈”ΎΗΟΆ≈Ε”œ»«Α“‘CAΉςΫΜΜΜΒΞ‘Σ ±ΒΡ≤ζΝΩ(ΆΦ5a)ΓΘ
Ψί¥Υ,ΫαΚœ…ζΈο–≈œΔ―ßΖ÷Έω,―–ΨΩ’Ώ»œΈΣCΫαΙΙ”ρ ήΧεΒΉΈοΒΡΧΊ“λ–‘≤Μ «ΉηΑ≠Ε‘NRPSΙΛ≥ΧΜ·ΗΡ‘λΒΡ“ρΥΊΓΘCalcottΒ»[63]ΫΪPheATE- ProCATΡΘ–Ά÷–ProCATΡΘΩιA”ρ“‘…œ ω÷ΊΉι±ΏΫγΫΜΜΜΈΣL-LeuΒΡA”ρΚσ,≥…ΙΠΚœ≥…ΝΥD-Phe- L-Leu,Ζ¥≤ΒΝΥC”ρ ήΧεΒΉΈοΧΊ“λ–‘’β“ΜΦΌΥΒ,÷ΛΟς‘ΎΫΜΜΜNRPS¥ΏΜ·ΡΘΩι ±,Ά®Ιΐ―Γ‘ώ”––ßΒΡ÷ΊΉι±ΏΫγΦ¥Ω…ΫΜΜΜA”ρ, Βœ÷Ε‘Ζ«ΚΥΧ«ΧεκΡ…ζΈοΚœ≥…ΆΨΨΕΗΡ‘λ,Μώ»Γ–¬ΒΡΖ«ΚΥΧ«ΧεκΡΓΘCalcottΒΡ Β―ιΕ‘Τ’±ιΙΪ»œΒΡC”ρ ήΧεΒΉΈοΧΊ“λ–‘’β“Μάμ¬έΧα≥ωΝΥΨό¥σΧτ’ΫΓΘ»ΜΕχΤδΕΰκΡΚœ≥…÷–Ϋω“‘ Ε±πL-LeuΒΡA”ρΫΜΜΜProCAT,“ρ¥Υ¥φ‘Ύ≈Φ»Μ–‘,»±…ΌΕ‘ΤδΥϊΑ±ΜυΥαΧΊ“λΒΡA”ρΫΜΜΜΫαΙϊΓΘ
4 ΉήΫα”κ’ΙΆϊ
…ηΦΤΖ«ΚΥΧ«ΧεκΡΒΡ…ζΈοΚœ≥…ΆΨΨΕ,“‘≤ζ…ζΨΏ”––¬ΫαΙΙΜρΗϋΚΟ“©άμΜν–‘ΒΡ–¬–ΆΜ·ΚœΈο,“Μ÷± «Χλ»Μ≤ζΈοΚœ≥……ζΈο―ßΒΡΡΩ±ξΓΘΜυ”ΎΖ«ΚΥΧ«ΧεκΡΚœ≥…ΟΗΒΡœΏ–‘¥ΏΜ·Μζ÷Τ,άμ¬έ…œΕ®œρ÷ΊΉι¥ΏΜ·ΡΘΩιΩ…“‘ΜώΒΟ»Έ“βΑ±ΜυΥα–ρΝ–ΉιΚœΒΡΕύκΡΓΘ»ΜΕχ‘”ΚœNRPSΒΡ¥ΏΜ·–߬ œό÷ΤΝΥΤδΩ…–––‘ΚΆ ”ϖ‘ΓΘΥφΉ≈Ε‘NRPSΗςΙΠΡή”ρΒΡΫαΙΙ―–ΨΩ»’“φ…ν»κ,―Γ‘ώΚœ ΒΡ¥ΏΜ·ΒΞ‘ΣΚΆάμœκΒΡ÷ΊΉι±ΏΫγ Ι‘”ΚœNRPSΩ… Ε±πΦ”‘ΊΡΩΒΡΒΉΈοΚΆΧαΗΏΤδ¥ΏΜ·–߬ ≥…ΈΣΩ…ΡήΓΘ“ρ¥Υ,Κœάμ…ηΦΤΙΙΫ®≤Ώ¬‘,‘ –μ¥ΏΜ·ΒΞ‘Σ»Έ“β÷ΊΉι,¥”ΕχΗΏ–ßΚœ≥…Ή‘»ΜΫγ…–Έ¥¥φ‘ΎΒΡΜ·ΚœΈο,Ε‘‘ωΦ”Ζ«ΚΥΧ«ΧεκΡΦ“ΉεΜ·ΚœΈοΕύ―υ–‘ΓΔΩΣΖΔ–¬“©ΨΏ”–÷Ί¥σ“β“εΓΘ
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷Ψ
