
’Σ“ΣΘΚ¥©ΡΛκΡ «“ΜάύΨΏ”–ΧΊ ⥩ΡΛΙΠΡήΒΡΕύκΡΖ÷Ή” ,Ρή–·¥χΤδΥϋΖ÷Ή”…θ÷Ν≥§Ζ÷Ή”Ω≈ΝΘ¥©ΡΛΫχ»κœΗΑϊΡΎ≤Ω. ‘γΤΎ―–ΨΩ»œΈΣ ,ΤδΫχΑϊ «“Μ÷÷Έό–η ήΧεΓΔ“≤≤Μ¥φ‘Ύ±ΞΚΆΉ¥Χ§ΒΡΖ«Ψ≠ΒδΑϊΆΧ––ΈΣ. ΫϋΡξ―–ΨΩ±μΟς ,Τ䥩ΡΛΜζ÷ΤΩ…Ρή”κΤδΚ§”–ΒΡΑ±ΜυΥα÷÷άύ”–Κή¥σΙΊœΒ. œ÷‘Ύ ,¥©ΡΛκΡΒΡ¥©ΡΛΙΐ≥Χ≥ΤΈΣΨό–ΆΑϊ“ϊ––ΈΣ ,Υϋ”κ¥ΪΆ≥ΒΡΑϊΆΧ–Έ ΫΚήœύΥΤ. Β±»Μ ,ΜΙΩ…Ρή¥φ‘ΎΉ≈ΤδΥϋΒΡΫχΑϊΖΫ ΫΕχΟΜ”–±Μ÷ΛΟςΜρΖΔœ÷. ΙΊ”Ύ¥©ΡΛκΡΒΡ”Π”Ο“≤ «»ΥΟ«ΉνΗ––Υ»ΛΒΡ ,‘ΎΚήΕύΝλ”ρΒΡ―–ΨΩΕΦ‘ΎΫχ––≤Δ≤ΜΕœ»ΓΒΟΫχ’Ι. ≤Μ¬έ «…ζΈοΫγΜΙ «“Ϋ―ßΫγ ,¥©ΡΛκΡΕΦ±Μ»œΈΣΫΪ «“ΜάύΖ«≥Θ”–ΖΔ’Ι«±ΝΠΒΡΕύκΡΖ÷Ή”ΓΘ
¥©ΡΛκΡ « 20 άΦΆ÷–ΤΎΩΣ Φ»œ ΕΒΫΒΡ“ΜάύΨΏΧΊ βΙΠΡήΒΡΕύκΡΖ÷Ή” ,”ΔΈΡ―ß θΟϊ≥Τ“ΜΑψ–¥ΈΣ cell2penetrating peptides(CPPs) . ΥϋΟ«“ΜΑψ”… 30 Ηω“‘œ¬ΒΡΑ±ΜυΥα≤–ΜυΙΙ≥… ,ΨΏ”–…ζΈοΡΛ¥©ΆΗΙΠΡή ,ΜΙΩ…–·¥χΤδΥϋΖ÷Ή”…θ÷Ν≥§Ζ÷Ή”Ω≈ΝΘΫχ»κœΗΑϊ÷–[1 ,2 ]. ’ΐ «“ρΈΣ’β÷÷ΧΊ βΒΡΙΠΡή ,ΥϋΟ«±ΜΩ¥Ής…ζΈοΜν–‘Ζ÷Ή””––ßΒΡœΗΑϊΡΎΉΣ‘ΥΙΛΨΏ ,ΨΏ”–ΙψΖΚΒΡ”Π”Ο«ΑΨΑ.
GreenΒ»[3 ,4 ] Ήœ»ΖΔœ÷≤Δ÷Λ Β»ΥΟβ“Ώ»±œί≤ΓΕΨ(HIV21) ΒΡΖ¥ ΫΦΛΜνΒΑΑΉ TATΨΏ”–ΩγΡΛΉΣ“ΤΒΡΙΠΡή.ΚσΨ≠ Β―ι÷ΛΟς ,’φ’ΐΤπΩγΡΛΉΣ‘ΥΉς”ΟΒΡΤδ Β « TATΒΡ“ΜΕΈ±Μ≥ΤΈΣΒΑΑΉΉΣΒΦ”ρ (PTD) ΒΡΧΊ βΑ±ΜυΥα–ρΝ–(49ΓΪ57 ΈΜΑ±ΜυΥα) . ‘Ύ¥Υ÷°Κσ ,ΚήΕύΨΏΩγΡΛΙΠΡήΒΡΕύκΡΖ÷Ή”¬Ϋ–χ±ΜΖΔœ÷. œ÷‘Ύ ,Ε‘¥©ΡΛκΡΒΡ―–ΨΩ «…ζΈοΩΤ―ßΚΆΝΌ¥≤“Ϋ“©―ßΒ»Νλ”ρΒΡ»»Βψ ,ΙΊ”ΎΥϋΟ«ΒΡΫαΙΙΓΔ–‘÷ ΓΔ¥©ΡΛΜζ÷ΤΦΑΩ…Ρή¥φ‘ΎΒΡœΗΑϊΕΨ–‘Β»ΖΫΟφΕΦ’ΙΩΣΝΥœΗ÷¬Εχ…ν»κΒΡ Β―ι―–ΨΩ ,¥”Εχ ΙΈ“Ο«Ε‘ΤδΝΥΫβ‘Ϋά¥‘Ϋ»ΪΟφ.
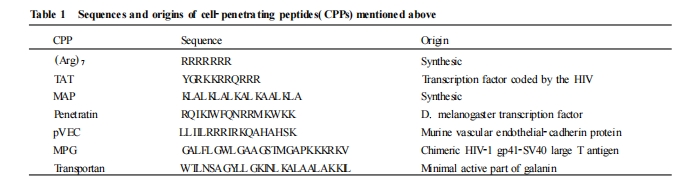
1 ¥©ΡΛκΡΒΡΧΊΒψΦΑΖ÷άύ
ΒΫΡΩ«ΑΈΣ÷Ι ,“―ΖΔœ÷ΝΥΚήΕύ÷÷¥©ΡΛκΡ ,ΟΩΡξΜΙ‘Ύ≤ΜΕœ‘ωΦ”. ΨΓΙή‘ΎΜ·―ßΫαΙΙ…œΩ…Ρή¥φ‘ΎΫœ¥σ≤ν±π ,ΒΪ“ΜΑψΕΦ¥χ”–ΫœΕύΒΡ’ΐΒγΚ…ΚΆΨΏ”–ΝΫ«Ή–‘.ΗυΨί≤ΜΆ§ΒΡΖ÷άύ±ξΉΦ ,Ω…ΫΪ¥©ΡΛκΡΖ÷ΈΣ≤ΜΆ§ΒΡ÷÷άύ. ¥”ΫαΙΙΧΊΒψ…œΥΒ ,‘γΤΎ”–»ΥΫΪΤδΦρΒΞΖ÷ΈΣ : ΔΌ≤ΜΨΏ”–Βδ–ΆΫαΙΙΒΡ¥χ¥σΝΩ―τάκΉ”ΒΡ¥©ΡΛκΡ ,»γ TATΚΆ penetratins; ΔΎά¥‘¥”ΎΒΑΑΉ–≈Κ≈–ρΝ–ΒΡΝΫ«ΉΠΝ¬ί–ΐκΡ[5 ]. ΗυΨίΤδά¥‘¥ΧΊ’ςΫχ––Ζ÷άύ ,÷ς“ΣΖ÷ΈΣ 3 άύ : ΔΌά¥‘¥”ΎΒΑΑΉΒΡ¥©ΡΛκΡ ,»γpenetratinΓΔTATΚΆpVEC Β».ΥϋΟ«Ά®≥ΘΨΏ”–ΉΣ‘ΥΒΑΑΉΉν–ΓΒΡ”––ßΤ§ΕΈ ,Φ¥ΒΑΑΉΉΣΒΦ≤ΩΖ÷ΜρΡΛ“ΉΈΜ–ρΝ– ; ΔΎΡΘ Ϋ (model) ¥©ΡΛκΡ ,»γMAP ΚΆ (Arg) 7 Β». ΥϋΟ« «ΈΣΝΥ–Έ≥…»ΖΕ®ΒΡΝΫ«Ή–‘ΠΝ¬ί–ΐΫαΙΙΜρΡΘΡβ“―÷Σ¥©ΡΛκΡΫαΙΙΕχ»ΥΙΛΚœ≥…ΒΡ ; Δέ»ΥΙΛ…ηΦΤΚœ≥…ΒΡ¥©ΡΛκΡ ,»γ MPG ΚΆ Transportan Β».ΥϋΟ«Ά®≥Θ ««ΕΚœΕύκΡ ,Κ§”– 1 Ηω ηΥ°≤ΩΖ÷ΚΆ 1 Ηω«ΉΥ°≤ΩΖ÷[6 ].
2 ¥©ΡΛκΡΒΡΡΎΜ·Μζ÷Τ
Β―ι÷ΛΟς ,≤ΜΆ§ΒΡ¥©ΡΛκΡ¥©ΡΛΡήΝΠ «”–≤ν±πΒΡ.Α±ΜυΥαΙΙ≥…ΓΔΖ÷Ή”¥σ–ΓΓΔΖ÷Ή”ΙΙœσΦΑΗΚ‘ΊΈοάύ–ΆΒ»ΕΦΕ‘¥©ΡΛΡήΝΠ”–”Αœλ.
Φν–‘Α±ΜυΥαΒΡ¥φ‘Ύ±Μ»œΈΣ «¥©ΡΛκΡΒΡ“ΜΗω±ξ÷Ψ. Ά®≥Θ¥σΝΩΒγΚ…ΒΡ¥φ‘Ύ≈≈≥ΐΝΥ¥©ΡΛκΡΆ®Ιΐ±ΜΕ·‘Υ δΆ®Ιΐ÷§ΥΪ≤ψΒΡΩ…Ρή. Β±”ΟΤδΥϋΒΡΑ±ΜυΥα≤–Μυ¥ζΧφΨΪΑ±Υα ±Ε‘¥©ΡΛκΡΒΡΡήΝΠ”ΑœλΫœ¥σ. »γάϊ”Ο±Μ±ϊΑ±Υα»Γ¥ζΒΡΕύκΡ retro2pVEC ,D2pVEC ΚΆ pVEC Ϋχ––±»Ϋœ―–ΨΩ ± ,ΖΔœ÷ N ΕΥΒΎ 5 ΈΜ ηΥ°–‘Α±ΜυΥα±Μ±ϊΑ±Υα»Γ¥ζ ±Τδ“ΉΈΜΡήΝΠœ¬ΫΒΟςœ‘ ,Β± Arg6 ,Arg8 Μρ Ser17±Μ±ϊΑ±Υα»Γ¥ζ ± ,Τδ±ΜœΗΑϊ…ψ»κΡήΝΠ‘ω«Ω[7 ].
Geisler Β»[8 ]±»ΫœΝΥ≤ΜΆ§≥ΛΕ»ΒΡΨΏ Δρ–ΆΕύΨέΗ§Α±Υα¬ί–ΐΙ«ΦήΒΡΝΫ«Ή¥©ΡΛκΡΫχΑϊΡήΝΠ ,ΖΔœ÷≥Λ–ρΝ–ΒΡP14LRR±»ΫœΕΧΒΡ P11LRR Ϋχ»κ MCF27 œΗΑϊΒΡΡήΝΠΗΏ 7ΓΪ12 ±Ε ,±» TAT ΗΏ 35 ±Ε. άϊ”Ο”ΪΙβΦΦ θΦΑ CD(circular dichroism) ΙβΤΉ―ßΦΦ θ ,―–ΨΩ 2 ÷÷¥©ΡΛκΡtransportan ΚΆ pAntp ΦΑΤδΆΜ±δΧε”κ≤ΜΆ§ΒγΚ…ΟήΕ»ΝΉ÷§Ρ“≈ίΒΡœύΜΞΉς”Ο Β―ι±μΟς ,pAntp ΦΑΤδΆΜ±δΧε“ΐΤπΒΡΡΛΜ묓≥ΧΕ»”κΥϊΟ«ΒΡΕΰΦΕΫαΙΙ”–ΙΊ. ‘Ύ¬ί–ΐΉ¥Χ§ ,ΕύκΡΕ‘ΡΛΗ…»≈Κή–Γ. Β± pAntp ΦΑΤδΆΜ±δΧε±δΈΣΠ¬ΫαΙΙ ± ,“ΐΤπΡΛΜ묓. Ε®œρ CD ±μΟς ,Β± transportan ΚΆpAntp ΈΣ¬ί–ΐΉ¥Χ§ ± , ÷Μ¥φ‘Ύ”ΎΡ“≈ίΡΛ±μΟφ , ΒΪpAntp ΒΡΝΫ÷÷ΆΜ±δΧεΡήΫχ»κΡΛΡΎ[9 ]. Ά®Ιΐ―–ΨΩ 3 ÷÷÷ς“ΣΒΡΝΫ«Ή–‘¥©ΡΛκΡ PΠ¬,PΠΝΚΆ Pep21 –Έ≥…ΩΉΒάΒΡΡήΝΠ ,“≤÷ΛΟςΩΉΒάΒΡ–Έ≥…”κΕύκΡΫαΙΙΧΊ’ς”–ΚήΚΟΒΡœύΙΊ–‘ ,ΧΊ±π «ΥϋΟ«ΒΡΙΙœσ±δΜ·[10 ].
¥ΥΆβ ,ΗΚ‘ΊΈοΒΡ¥σ–ΓΚΆάύ–Ά“≤Μα”Αœλ¥©ΡΛκΡΒΡ¥©ΡΛΡήΝΠ. “ΜΑψ«ιΩωœ¬ ,–ΓΖ÷Ή”ΒΡΗΚ‘ΊΈο≤Μ”ΑœλΡΎΜ·(internalization) ΒΡ–ß¬ ,ΒΪ¥σΖ÷Ή”ΗΚ‘ΊΈο ± ,ΡΎΜ·–߬ Οςœ‘ΫΒΒΆ[6 ]. Ε‘ Pep21 ‘Υ δΡήΝΠΒΡ―–ΨΩΖΔœ÷ ,Ω≈ΝΘΉ¥ΗΚ‘ΊΈοΒΡΩ≈ΝΘ¥σ–ΓΚΆΨυ‘»–‘ΕΦ÷±Ϋ”“άάΒ”Ύ Pep21ΠΑΗΚ‘ΊΈοΉι≥…±»¬ ,≤Δ”κ…ζΈο–߬ ÷±Ϋ”œύΙΊ[11 ]. “ρ¥Υ ,≤ΜΆ§ΒΡ¥©ΡΛκΡ‘Ύ–·¥χ≤ΜΆ§Ζ÷Ή” ±≤…»ΓΒΡΫχΑϊΖΫ ΫΩ…Ρή”–≤ν“λ ,Ά§ ±ΤδΫχΑϊ ±Φδ“≤Μα≤ΜΆ§.
‘γΤΎΙέ≤λΖΔœ÷ ,¥©ΡΛκΡΒΡΡΎΜ·Ιΐ≥ΧΩ…‘Ύ 4 ΓφΒΡΒΆΈ¬ΜΖΨ≥œ¬ΖΔ…ζ ,Εχ«“Έό±ΞΚΆΉ¥Χ§. “ρ¥Υ»œΈΣ ,ΗΟΙΐ≥Χ≤ΔΖ«Ψ≠ΒδΒΡΑϊΆΧΉς”Ο (endocytosis) ,“≤≤Μ–η ήΧεΒΑΑΉΒΡΫιΒΦ[12ΓΪ14 ]. ΒΪ «ΉνΫϋΒΡ―–ΨΩΫαΙϊ±μΟς ,ΑϊΆΧΉς”Ο‘ΎΡΎΜ·Ιΐ≥Χ÷–Ω…Ρή’Φ”–÷Ί“ΣΒΊΈΜ.
Nakase Β»[15 ] Ά®ΙΐΕ‘ TAT ΚΆΤδΥϋΗΜΚ§ΨΪΑ±ΥαΕύκΡΒΡ―–ΨΩ»œΈΣ ,‘γΤΎ Ι”ΟœΗΑϊΙΧΕ®Μ·ΦΦ θΓΔ”ΪΙβœ‘ΈΔΨΒΦΦ θΦΑΝς ΫœΗΑϊΦΤΝΩΦΦ θΒ»Ϋχ–– Β―ι ± ,Κ§”–ΨΪΑ±ΥαΕύκΡΒΡΡΎΚ≠ΧεΡΛΨ≠ΙΧΕ®Μ·Ιΐ≥ΧΜα±δΒΟΖ«≥Θ¥ύ»θ ,ΦΪ“ΉΤΤΝ―ΚΆ–Ι¬Ε ,¥”Εχ Ι¥©ΡΛκΡΟ÷…ΔΖ÷≤Φ”ΎœΗΑϊ÷ ΚΆœΗΑϊΚΥ¥Π. ’ΐ «’β–©ΦΌœσΒΡ≥ωœ÷ Ι»ΥΟ«»œΈΣ ,¥©ΡΛκΡ¥©ΡΛ≤Μ «Ψ≠ΒδΒΡΑϊΆΧΆΨΨΕ. ΒΦ …œ ,’β–©ΕύκΡ¥χ”–¥σΝΩ―τάκΉ”≤Δ±ΜΡΛ«ΩΝ“Έϋ“ΐ ,ΚήΡ―¥”ΗΏ«ΉΚΆΝΠΒΡœΗΑϊ±μΟφΆξ»ΪάκΩΣ. ‘ΎΒΆΈ¬œ¬ ,”ΟΗΜΚ§ΨΪΑ±ΥαΕύκΡ¥ΠάμœΗΑϊ ± ,≤ΔΟΜ”–ΡΎΆΧΉς”Ο≥ωœ÷ ,ΒΪ”…”ΎΙΧΕ®≤ΌΉςΕχ«αΈΔΈ…¬“ΒΡ÷ ΡΛΜα ΙœΗΑϊ±μΟφΈϋΗΫΒΡΕύκΡΧ”Ά―Ϋχ»κΑϊ÷ ,¥”Εχ≥ωœ÷ΝΥΕύκΡΡΎΜ·Ϋχ»κΑϊ÷ ΚΆΚΥΒΡΦΌœσ. άϊ”ΟΆξ’ϊΒΡΜνœΗΑϊΫχ––÷Ί–¬―–ΨΩ±μΟς ,‘ΎΧΊΕ®ΒΡ«ιΩωœ¬ ,ΡΎΆΧΆΨΨΕ «’β–©ΕύκΡΡΎΜ·ΒΡ÷ς“ΣΆΨΨΕ. Ε®ΈΜ”ΎœΗΑϊΡΛΒΡΒΑΑΉΨέΧ« ,Αϋά®ΝρΥα““θΘΗΈΥΊ ,±Μ»œΈΣ‘ΎΡΎΆΧΙΐ≥Χ÷–Τπ÷Ί“ΣΉς”Ο ,’β“ΜΧΊ βΡΎΆΧΙΐ≥Χ±Μ≥ΤΈΣΨό–ΆΑϊ“ϊ(macropinocytosis) . ’βΗωΙΐ≥ΧΤπ Φ”ΎœΗΑϊΦΛΜνΚΆ÷ ΡΛΒΡ±Ώ‘Β≤®Ε· ,¥”ΕχΆΧ …¥σΝΩΒΡΑϊΆβ“ΚΫχ»κΨό–ΆΑϊ“ϊ≈ί. ‘ΎΆξ’ϊΜνœΗΑϊ÷– ,≤ΜΫχ––ΙΧΕ®Μ·¥Πάμ ± ,»‘”–…Ό≤ΩΖ÷ΕύκΡΫχ»κΑϊ÷ ΚΆΚΥ÷ήΈß ,ΥΒΟςΩ…ΡήΜΙ”–“Μ–©ΤδΥϋΒΡΡΎΜ·Μζ÷Τ¥φ‘Ύ.
TATΚΆΤδΥϋΒΑΑΉΉΣΒΦ”ρ“‘Ψό–ΆΑϊ“ϊΆΨΨΕ Βœ÷ΉΣ‘ΥΒΡΡΘ–Ά «“ΜΗωΕύΫΉΕΈΙΐ≥Χ ,Αϋά® TAT ”κœΗΑϊ±μΟφΫαΚœΓΔ¥ΧΦΛΨό–ΆΑϊ“ϊ≈ί…ψ»κ TAT ΦΑΤδΗΚ‘ΊΈοΓΔΡΎΚ≠ΧεΆ―άκΫχ»κœΗΑϊ÷ ÷–Β»“ΜœΒΝ– ¬Φΰ. ΒΎ“Μ≤Ϋ÷– TATΫαΚœΒΫœΗΑϊ±μΟφ «Ά®ΙΐœΗΑϊ±μΟφΤ’±ι¥φ‘ΎΒΡΨέΧ«Ν¥Άξ≥…ΒΡ. TAT¥ΧΦΛΨό–ΆΑϊ“ϊ≈ίΒΡ≥ωœ÷ΤδΜζ÷Τ…–≤Μ÷ΣΒά ,Ω…Ρή”κœΗΑϊ±μΟφΒΑΑΉΒΡΫαΚœ”–ΙΊ ,“≤Ω…ΡήΆ®ΙΐΒΑΑΉΨέΧ«ΜρΧ«÷§ά¥ Βœ÷. Ά®ΙΐΨό–ΆΑϊ“ϊ≈ίΒΡ…ψ»κ ,Υυ”–ΒΡœΗΑϊΕΦ–Έ≥…“ΚΧ§ΡΎΆΧ–Έ Ϋ ,’βΕ‘”Ύ TAT ΚΆΕύΨέΨΪΑ±ΥαΒΡΉΣΒΦ «±Ί–ηΒΡ. ΉνΚσ“Μ≤Ϋ «¥”Ψό–ΆΑϊ“ϊ≈ίΧ”Ά―Ϋχ»κœΗΑϊ÷ ,’βΗωΙΐ≥ΧΩ…Ρή“άάΒΡΎΚ≠Χε÷– pH ΒΡœ¬ΫΒ ,ΑιΥφΤδΥϋ“ρΥΊ ΙΡΎΚ≠ΧεΡΛΖΔ…ζ«αΈΔΈ…¬“ ,¥”Εχ ΆΖ≈ TATΦΑΤδΗΚ‘ΊΈο[16 ].
»»ΝΠ―ß―–ΨΩœ‘ Ψ ,Ζ«ΝΫ«ΉΒΡΚΆ÷–Β»ΝΫ«ΉΒΡ¥©ΡΛκΡ”κΑ±ΜυΤœΨέΧ«( GAGs) ΨΏΗΏ«ΉΚΆΝΠ ,ΒΪ”κ’φΚΥœΗΑϊΒΡ«ΉΚΆΝΠΚήΒΆ. “ρ¥Υ ,¥©ΡΛκΡ”ΠΗΟΡή”κΑϊΡΛ≥…Ζ÷ ,»γ“θάκΉ”÷§άύΚΆ GAGs Β»Ά®ΙΐΨ≤ΒγΫαΚœ ,‘ΎΒΆΈΔΡΠΕϊ≈®Ε»œ¬“‘÷±Ϋ”ΆΗΡΛΒΡ¥Έ“ΣΆΨΨΕΫχΑϊ. ’β“≤÷ß≥÷ΝΥΧεΡΎ Β―ι―–ΨΩΥυΫ“ ΨΒΡ¥©ΡΛκΡΒΆΈΔΡΠΕϊ≈®Ε» ±ΑϊΆΧΉς”Ο «÷ς“ΣΒΡ…ψ»ΓΆΨΨΕ[17 ]. Β±¥©ΡΛκΡ≈®Ε»¥ο 100ΠΧmolL “‘…œΒΡΗΏ≈®Ε» ± ,…ζΈοΡΛ¥χ”–ΫœΕύΒΡ“θάκΉ”≥…Ζ÷,‘ΎΒΆΈ¬ ±÷±Ϋ”“ΉΈΜΜα±»ΫœΟςœ‘.
¥©ΡΛκΡΒΡΫχΑϊΆΨΨΕΩ…Ρή”–Εύ÷÷ ,÷Ν…ΌΑϋά®÷±Ϋ”“ΉΈΜΚΆΡΎΆΧΉς”Ο. ÷Ν”ΎΡΡ÷÷Μζ÷ΤΗϋ”––ß ,“ΉΈΜ≥ωœ÷”–ΕύΩλ“άάΒ”Ύ ΒΦ «ιΩω ,»γ¥©ΡΛκΡάύ–ΆΓΔ≈®Ε»ΓΔΈ¬Ε»ΓΔœΗΑϊΡήΝΩΥ°ΤΫΓΔΗΚ‘ΊΒΡ¥φ‘Ύ ,ΑϊΡΛ÷–÷§άύΉι≥…Β». Τδ÷–“ΜΗωάΐΉ”ΨΆ «άϊ”ΟΒΞ≤ψΡ“≈ίΚΆΗΤΜΤ¬ΧΥΊ”ΪΙβΜυΆ≈―–ΨΩ¥©ΡΛκΡ Tp10 ΦΑΤδΆΜ±δΧε ±ΖΔœ÷ ,ΡΎΜ·Μζ÷Τ”κ «Ζώ”–¥σΒΡΗΚ‘ΊΈο”–“ΜΕ®ΙΊœΒ. ‘Ύ»±ΖΠ¥σΒΡ«ΉΥ°–‘ΗΚ‘ΊΈο ± ,ΕύκΡ”’ΒΦΡΛΈ…¬“ ΙκΡ÷±Ϋ”“ΉΈΜΫχ»κΑϊΡΎ ;Β±¥χ”–¥σΒΡ«ΉΥ°–‘ΗΚ‘ΊΈο ±ΡΛΈ…¬“≥ΧΕ»ΫœΒΆ ,ΡΎΆΧΉς”ΟΥΤΚθ «Έ®“ΜΒΡ“ΉΈΜΜζ÷Τ[12 ].
3 ¥©ΡΛκΡΕ‘œΗΑϊΒΡ”Αœλ
“ΣΫΪ¥©ΡΛκΡ”Ο”ΎΝΌ¥≤“©ΈοΒΡ‘Υ δΙΛΨΏ , Ήœ»±Ί–κ»Ζ»œκΡ±Ψ…μ≤ΜΜαΕ‘’ΐ≥ΘœΗΑϊ‘λ≥…≤Μάϊ”Αœλ ,‘Ύ’βΖΫΟφ“―Ϋχ––ΝΥ“Μ–©―–ΨΩ ,ΒΪ¥σΕύ «ΧεΆβ Β―ι ,ΧεΡΎ―–ΨΩΫœ…Ό. Ά®≥Θ―–ΨΩ÷ς“ΣΦ·÷–‘ΎΝΫ≤ΩΖ÷ : ΔΌΕ‘œΗΑϊΡΛΚΆœΗΑϊΤςΒΡΕΨ–‘”Αœλ ; ΔΎ¥©ΡΛκΡ”κœΗΑϊ≥…Ζ÷œύΜΞΉς”ΟΕχ‘λ≥…ΒΡΕΨ–‘”Αœλ[6 ].
“Μ–©ΡΘ Ϋ¥©ΡΛκΡ ,»γ MAP ,ΦΑ»ΥΈΣ…ηΦΤΒΡ¥©ΡΛκΡ ,»γ transportan ,ΥϋΟ«‘ΎΫαΙΙ…œ”κΩΙΨζκΡάύΥΤ ,Ε‘œΗΑϊΡΛΒΡΈ»Ε®–‘”–Ϋœ¥σΒΡ”Αœλ. Saar Β»[19 ] ―–ΨΩΝΥ 5 ÷÷¥©ΡΛΕύκΡ pAntp (43ΓΪ58) ΓΔpTAT(48ΓΪ60) ΓΔpVEC(615ΓΪ632) ΚΆ MAPΓΔtransportan10 Β»Ε‘ 2 ÷÷»ΥΑ©œΗΑϊ÷ξK562 ΚΆ MDA2MB2231 ΦΑ”άΨΟ÷ςΕ·¬ωΡΎΤΛœΗΑϊΒΡœΗΑϊΡΛΕΨ–‘. ‘ΎΥυ”–œΗΑϊ÷ξ÷– ,pAntp (43ΓΪ58) ΓΔpTAT(48 ΓΪ 60) ΚΆ pVEC ( 615 ΓΪ 632 ) –Ι ¬Ε ; Εχ MAP ΚΆtransportan10 Ω…“ΐΤπΟςœ‘ΒΡœΗΑϊ–Ι¬Ε. Β± K562 ΚΆMDA2MB2231 Αϊ¥Π”Ύ 10ΠΧmolΠΑL MAP ΚΆ transportan10÷– 10 min ± ,Φ¥Ω… Ι 40 %»ιΥαΆ―«βΟΗ–Ι¬Ε. ”––©―–ΨΩ±μΟς ,¥©ΡΛκΡΒΡΕΨ–‘ «”κΤδ÷÷άύΓΔ≈®Ε»ΚΆΗΚ‘ΊΈοΟή«–œύΙΊΒΡ. Β± TAT(48ΓΪ57) ≈®Ε»¥ο 100ΠΧmolΠΑL “‘…œ ±»‘Μυ±Ψ…œΕ‘Υυ”–œΗΑϊΕΦΈόΚΠ ,Εχ Antp (43ΓΪ58) »¥±μœ÷≥ωΟςœ‘ΒΡΕΨ–‘. ‘Ύ≤ΜΩΦ¬«ΗΚ‘ΊΈο–ρΝ– ± ,”κ TAT(48ΓΪ57) ΚΆ Antp (43ΓΪ58) ΫαΚœΕύκΡ≈®Ε»¥σ”Ύ 10ΠΧmolΠΑL ±¥ΞΖΔΟςœ‘ΒΡ≥ΛΕ»“άάΒΕΨ–‘. Ϋχ“Μ≤Ϋ ΐΨίΜΙ±μΟς ,ΕΨ–‘ΒΡΖΔ…ζ”κΑϊΡΎΝΫΗω”ΠΦΛ–≈Κ≈Ά®¬ΖΒΡΦΛΜν”–ΙΊ. Υδ»ΜΑϋΚ§ΒΑΑΉΉΣΒΦ”ρΒΡ TAT κΡ (48ΓΪ85) ‘ΎΧεΆβ≈ύ―χ Β―ι÷–Έ¥±μœ÷≥ω…ώΨ≠ΕΨ–‘ ,ΒΪΚ§ΠΝ¬ί–ΐΕΈ(37ΓΪ60) ΒΡ TAT κΡΕ‘ HeLa œΗΑϊΒΡ«®―”’’…δΜαΒΦ÷¬60 %œΗΑϊΜΒΥά. Εχ«“ TAT ΕύκΡ (43ΓΪ60) Υδ÷Μ±μœ÷10 %ΓΪ15 %ΕΨ–‘ ,œΗΑϊΕΧ ±Φδ≈ύ―χ‘Ύ 20ΓΪ100ΠΧmolΠΑLΒΡ PTD ≈®Ε»œ¬“≤Έ¥±μœ÷»ΈΚΈ≤ΜΝΦΉς”Ο ,Εχ 500ΠΧmolΠΑL PTD ‘ΎΚήΕΧΒΡ ±ΦδΡΎΦ¥Ω…ΒΦ÷¬≥…œΥΈ§œΗΑϊΙΠΡήΗΡ±δ. Χα Ψ PTD ≈®Ε»≥§Ιΐ 100ΠΧmolΠΑL Ω…ΡήΜαΒΦ÷¬ΕΨ–‘[21ΓΪ24 ]. ”…“‘…œΉ ΝœΩ…“‘Ω¥≥ω ,“Σ≥δΖ÷ΖΔΜ”¥©ΡΛκΡΕά“ΜΈόΕΰΒΡΉς”Ο ,Ε‘ΤδΕΨ–‘”ΑœλΒΡΫχ“Μ≤Ϋ―–ΨΩ «Ζ«≥Θ±Ί“ΣΒΡ.
4 ¥©ΡΛκΡΒΡ”Π”Ο«ΑΨΑ
Μυ”Ύ¥©ΡΛκΡΒΡΧΊ βΙΠΡή ,ΙΊ”ΎΥϋΟ«”Π”ΟΖΫΟφΒΡ―–ΨΩ’ΐ‘Ύ―ΗΥΌ’ΙΩΣ. ¥©ΡΛκΡΩ…“‘–·¥χΒΡΓΑΜθΈοΓ±≤Μ÷Μ «–ΓΖ÷Ή”Έο÷ ,ΜΙΩ…“‘ «ΕύκΡΓΔΒΑΑΉ÷ ΓΔΚΥΥα ,…θ÷Ν≥§Ζ÷Ή”Ω≈ΝΘ.
4. 1 ‘Υ δΕύκΡΚΆΒΑΑΉ÷
Morishita Β»[25 ]άϊ”ΟΕύΨέΨΪΑ±Υα≥…ΙΠΫΪ“»ΒΚΥΊΖ÷Ή”¥χ»κΝΥœΗΑϊ. Β―ι±μΟς ,ΒΞΕά“»ΒΚΥΊ¥φ‘Ύ ±Ιέ≤λ≤ΜΒΫ≥ΠΈϋ ’ ,ΒΪ‘ΎΆ§ ±Ηχ“‘ D2(Arg) 6 ΚΆ L2(Arg) 6 ±≥ œ÷ΦΝΝΩ“άάΒ–‘Οςœ‘ΒΡ“»ΒΚΥΊ≥ΠΈϋ ’. D2(Arg) 6 ±» L2(Arg) 6 Ε‘“»ΒΚΥΊΈϋ ’”ΑœλΗϋ¥σ. ‘ΎΕύΨέΨΪΑ±Υα÷– D2(Arg) 8 Ε‘“»ΒΚΥΊ≥ΠΈϋ ’±μœ÷≥ωΉν«ΩΒΡ”Αœλ. Εχ«“ ,¥Υ ±≤Δ≤Μ–η“ΣΫΪΕύΨέΨΪΑ±Υα”κ“»ΒΚΥΊΫχ––Ι≤ΦέΝ§Ϋ”.Ω…“‘œκœσ»γΙϊ’β÷÷ΖΫΖ®≥…ΙΠ ,ΫΪΈΣΧ«Ρρ≤ΓΜΦ’ΏΒΡ÷ΈΝΤ¥χά¥ΕύΟ¥…νΩΧΒΡ”Αœλ. ΤδΥϋΕύκΡΜρΒΑΑΉ ,»γ…ώΨ≠‘Σ”Σ―χ“ρΉ” ( GDNF)[26 ] ΓΔΠ¬2Ακ»ιΧ«ή’ΟΗ[27 ] ΓΔ«Ω–ßΈΗΟΗ“÷ΥΊ A(PepA)[28 ] Β»ΕΦ±Μ±®ΒάΩ…Ά®Ιΐ”ꥩΡΛκΡΝ§Ϋ”Εχ”––ßΫχ»κœΗΑϊΡΎ≤Ω ,GDNF …θ÷ΝΩ…“‘Ά®Ιΐ―ΣΡ‘ΤΝ’œ.
4. 2 ‘Υ δ siRNA ΚΆ DNA
RNA Η…»≈ «―–ΨΩΜυ“ρΙΠΡήΓΔΜυ“ρΒςΩΊΚΆΜυ“ρ÷ΈΝΤΒΡ”––ßΖΫΖ®. ΥΪΝ¥ RNA ”Ο“‘”’ΒΦ–ρΝ–ΧΊ“λ–‘Μυ“ρ≥ΝΡ§. ΈΣ ΙΧΊΕ®Μυ“ρ≥ΝΡ§ ,Κœ ΒΡ siRNA ‘Υ δœΒΆ≥ «±Ί–ηΒΡ. Κ§ 9 ΗωΨΪΑ±ΥαΒΡ¥©ΡΛκΡΡή“‘Ζ«Ι≤Φέ–Έ Ϋ”κEGFPsiRNA –Έ≥…Η¥ΚœΈο ,≤ΔΫΪΤδΉΣ‘ΥΫχ»κΈ»Ε®±μ¥ο¬Χ…Ϊ”ΪΙβΒΑΑΉΒΡ GC2EGFP œΗΑϊ÷ξ÷–. Ϋχ»κΚσ ,siRNA≥ωœ÷‘ΎœΗΑϊΒΡΚΥ÷ή«χ ,≤ΔΩ…Ήν÷’ Βœ÷ΤδΡΩ±ξΜυ“ρegfp ΒΡ≥ΝΡ§[29 ]. Unnamalai Β»[30 ] “≤”Ο CPP-siRNA ΑϋΉΑΒΡ‘Υ δΖΫ Ϋ≥…ΙΠΫιΒΦ RNAi Ϋχ»κ―Χ≤ίœΗΑϊ. άϊ”Ο…ηΦΤΚœ≥…ΒΡΨό¥σΖ÷÷ßΒΡ¥©ΡΛκΡΖ÷Ή”ΜΙΡή≥…ΙΠΒΡΫΪDNA ÷ ΝΘΖ÷Ή”ΉΣ»ΨΫχ»κΕύ÷÷≤Η»ιΕ·ΈοœΗΑϊ÷– ,ΈΣDNA Ζ÷Ή”ΒΡ‘Υ δΧαΙ©ΝΥ–¬ΒΡ―Γ‘ώ ,«“–߬ ±»¥ΪΆ≥ΖΫΖ®ΗϋΗΏ[31 ]
4. 3 ≥§Ζ÷Ή”Ω≈ΝΘΒΡ‘Υ δ
Sethuraman Β»[32 ] ‘ΎΦΪΕΥ pH ΟτΗ–ΨέΚœΈοΚΆ¥©ΡΛTATΒΡΜυ¥Γ…œΖΔ’ΙΝΥ“Μ÷÷–¬ΒΡ’κΕ‘Υα–‘ ΒΧεΝωΒΡ“©ΈοΑ–œρ‘Υ δœΒΆ≥. ‘Υ δœΒΆ≥Αϋά® 2 ΗωΉιΖ÷ : ΔΌΚ§”– ηΥ°ΚΥ–ΡΒΡ”…ΕύΨέ L2»ιΥα(PLLA) ΙΙ≥…ΒΡΨέΚœΫΚΆ≈ ,Τδ«ΉΥ°ΆβΩ«ΑϋΚ§Ψέ““Εΰ¥Φ (PEG) Ν§Ϋ”ΒΡ TAT ΈΔΆ≈ ; ΔΎΦΪΕ» pH ΟτΗ–ΒΡ”…ΕύΨέ“λΕΓœ©θΘΜυΜ«ΑΖΒΊΥς–Ν(PST) ΚΆ PEG(PSD2b2PEG) ΙΙ≥…ΒΡΥΪ÷ΊΙ≤ΨέΧε. Β―ι÷ΛΟς ,TAT≤ΜΫω“ΉΈΜΫχ»κœΗΑϊΜΙΩ…≥ωœ÷‘ΎΚΥ÷ήΈß. ΥΒΟς’β―υΒΡ‘Υ δœΒΆ≥Ω…Ρή–·¥χ»ΈΚΈ ηΥ°“©ΈοΫ”ΫϋœΗΑϊΚΥ. Ε‘¥ΥœΒΆ≥Ϋχ“Μ≤ΫΒΡ Β―ι’ΐ‘ΎΫχ––÷– ,ΥϋΈΣ÷ΉΝωΑ–œρ÷ΈΝΤ¥χά¥ΝΥœΘΆϊ.
άϊ”Ο»ΎΚœκΡ diINF27 Ι≤Αϋ±Μ–Έ≥…Οβ“Ώ÷§÷ ΧεΜΙΡή Βœ÷÷§÷ ΧεΒΑΑΉΒΡΑϊ÷ ‘Υ δ. ―–ΨΩ±μΟς ,ΨΓΙήάϊ”Ο±μΟφ±Μ¥©ΡΛκΡ TAT –ό ΈΒΡ÷§÷ Χε≤ΜΡή Ι÷§÷ ΧεΩ≈ΝΘ÷±Ϋ”“ΉΈΜ¥©ΙΐœΗΑϊΡΛ ,ΒΪœΗΑϊΆ®ΙΐΡΎΆΧΉς”ΟΕ‘÷§÷ ΧεΒΡ…ψ»κ±»Έ¥–ό ΈΒΡ÷§÷ ΧεΧαΗΏΝΥ[33 ].
5 Ϋα”ο
¥©ΡΛκΡΒΡ―–ΨΩ «ΡΩ«ΑΒΡ“ΜΗω÷ΊΒψΚΆ»»Βψ ,Έ“Ιζ“≤ΩΣ’ΙΝΥ“ΜœΒΝ–ΙΊ”ΎΤδΫαΙΙΧΊ’ςΓΔ¥©ΡΘΜζ÷Τ”κΜν–‘ΓΔ«±‘ΎœΗΑϊΕΨ–‘Β»ΒΡ―–ΨΩ[34ΓΪ37 ],≤Δ»ΓΒΟΝΥ“ΜΕ®ΒΡ≥…Ιϊ. Υυ”–ΒΡ―–ΨΩΕΦ±μΟς ,¥©ΡΛκΡΩ…”––ßΫΪΕύκΡΓΔΒΑΑΉ÷ ΦΑ DNA ΓΔRNAΓΔ≥§Ζ÷Ή”Ω≈ΝΘΒ» , Ά®ΙΐΈό ήΧεΫιΒΦΓΔΈόΚΡΡήΒΡΖΫ Ϋ ,ΒΦ»κΕύ÷÷œΗΑϊ÷– , «“‘Ύ“ΜΕ®≈®Ε»ΖΕΈßΡΎ≤ΜΜα‘λ≥…œΗΑϊΥπ…Υ. Υδ»ΜΕ‘”Ύ¥©ΡΛκΡΫχ»κΜνœΗΑϊΒΡΨΏΧεΜζ÷Τ…–ΈόΕ®¬έ ,ΒΪΩ…“‘ΩœΕ®ΤδΉςΈΣ…ζΈοΜν–‘Ζ÷Ή””––ßΒΡœΗΑϊΡΎΉΣ‘ΥΙΛΨΏ ,±ΊΫΪ‘ΎœΗΑϊ…ζΈο―ßΓΔΜυ“ρ÷ΈΝΤΓΔ“©ΈοΧεΡΎΉΣ‘ΥΓΔΝΌ¥≤“©–ßΤάΦέ“‘ΦΑœΗΑϊΟβ“Ώ―ßΒ»Εύ÷÷―–ΨΩΝλ”ρΖΔΜ”≤ΜΩ…Χφ¥ζΒΡΉς”Ο. ΑιΥφΉ≈―–ΨΩΒΡΫχ–– ,”»Τδ «Ε‘Τ䥩ΡΛΜζ÷Τ―–ΨΩΒΡ≤ΜΕœ…ν»κ ,Έ“Ο«ΜΙΩ…Ρή»ΥΙΛΚœ≥…ΗϋΕύΒΡΓΔ¥©ΡΛΡήΝΠΗϋ«ΩΕχΕΨ–‘Κή–Γ…θ÷ΝΈόœΗΑϊΕΨ–‘ΒΡ¥©ΡΛκΡ , “‘”Ο”ΎΦ≤≤Γ÷ΈΝΤΒ»ΤδΥϋΖΫΟφ.
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷Ψ
