
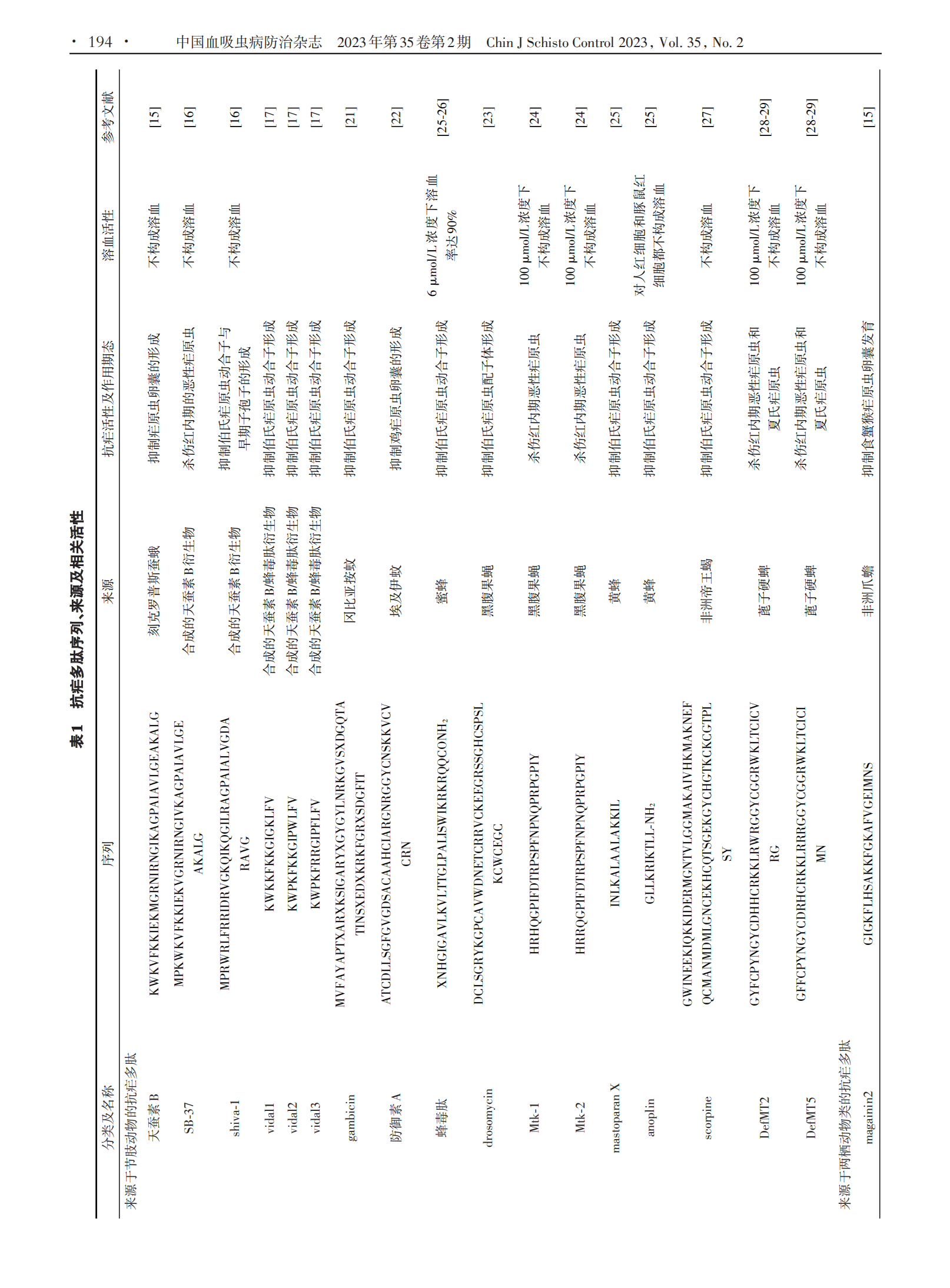
еЊвЊЃК ХБМВвРШЛЪЧШЋЧђживЊЕФЙЋЙВЮРЩњЮЪЬтжЎвЛЃЌШЋЧђШдгаНќвЛАыШЫПкУцСйИаШОЗчЯеЁЃФПЧАЃЌФЭвЉадГцжъЕФВЛЖЯГіЯжКЭРЉЩЂЪЙЕУДЋЭГПЙХБвЉж№НЅЪЇаЇЁЃвђДЫЃЌбаЗЂаТЕФПЙХБвЉЮяЦШдкУМНоЁЃПЙОњыФЪЧЛњЬхЬьШЛУтвпЗРгљЯЕЭГЕФживЊзщГЩВПЗжЃЌЮЊЕжгљжТВЁадШыЧжЬсЙЉСЫЕквЛЕРЗРЯпЃЌЦфгХЯШЙЅЛїЯИАћФЄЕФзїгУЛњжЦЪЙЦфВЛвзВњЩњФЭвЉадЃЌвђДЫБЛШЯЮЊЪЧаТаЭПЙХБвЉЮяЕФгХжЪКђбЁзЪдДжЎвЛЁЃБОЮФЖдОпгаПЙХБЛюадЕФПЙОњыФЁЊЁЊПЙХБЖрыФбаОПНјеЙНјаазлЪіЃЌВЂЖдЦфзїЮЊаТаЭПЙХБвЉЮяЕФбаОПЧАОАНјааеЙЭћЁЃ
ХБМВЪЧвЛжжгЩХБдГцИаШОв§Ц№ЁЂЭЈЙ§АДЮУДЋВЅЕФИаШОадМВВЁЃЌгыАЌзЬВЁЁЂНсКЫВЁвЛЦ№БЛСаЮЊШЋЧђШ§ДѓДЋШОВЁЃл1ЃнЁЃОнWHOБЈЕРЃЌ2021 ФъШЋЧђШдга 2.47 вкР§ХБМВВЁР§ЃЌ61.9ЭђР§ХБМВЛМепЫРЭіЃл2ЃнЁЃЖёадХБдГцЪЧЮЃКІШЫЬхзюЮЊбЯжиЕФХБдГцЃЌОјДѓВПЗжгыХБМВгаЙиЕФЫРЭіВЁР§ЖМЪЧгЩЖёадХБдГцИаШОв§Ц№ЃЌЦфжа90%ЗЂЩњдкШіЙўРвдФЯЗЧжоЕиЧјЃЌЖрЪ§ЮЊ5ЫъвдЯТЖљЭЏЁЃИУЕиЧјдаИОМАЦфЬЅЖљМЋвзИаШОХБМВЃЌЪЧдьГЩаТЩњЖљЬхжЪСПЙ§ЧсЁЂЦЖбЊКЭЫРЭіЕФжївЊдвђЃл3ЃнЁЃФПЧАЃЌХБМВвпУчаЇЙћгаЯоЃЌжївЊвРПППЙХБвЉЮяНјааЛЏбЇжЮСЦЃЌЕЋНќМИЪЎФъРДПЙХБвЉЕФВЛКЯРэЪЙгУЕМжТСЫХБдГцвЉЮяПЙадВњЩњКЭбИЫйРЉЩЂЃЌЪЙЕУАќРЈТШрЁЂМзЗњрЁЂЛЧАЗЖраСЁЂввАЗрзрЄЁЂппрЕШОЕфПЙХБвЉЮяж№НЅЪЇаЇЃл4ЃнЁЃ2001ФъЃЌWHOе§ЪНЭЦМівдЧрняЫиЮЊЛљДЁЕФСЊКЯгУвЉзїЮЊжЮСЦЖёадХБЕФвЛЯпгУвЉЃл5ЃнЃЌШЛЖјдкДѓфиЙЋКгДЮЧјгђГіЯжСЫХБдГцЖдЧрняЫиУєИаадЯТНЕЕФЯжЯѓЃл6ЃнЃЌЩѕжСдкФЯУРжоКЭЖЋФЯбЧЦеБщГіЯжСЫЖржиПЙаджъЁЃНижС2015Фъ2дТЃЌИУПЙадЧјгђвбРЉДѓЕНећИіДѓфиЙЋКгСїгђЃЌзюНќвЛаЉЗЧжоЙњМввВгаЖёадХБдГцЖдЧрняЫиВњЩњФЭвЉадЕФЯрЙиБЈЕРЃл7⁃8ЃнЁЃвђДЫЃЌбАеваТЕФЬцДњвЉЮяЦШдкУМНоЁЃ
ПЙОњыФЪЧЩњЮяЬхФкВњЩњЕФвЛжжаЁЗжзгЛюадыФЃЌЪЧЛњЬхЬьШЛУтвпЗРгљЯЕЭГЕФживЊзщГЩВПЗжЃЌВЛНігаУтвпЕїНкЛюадЃЌЖјЧвОпгаПЙЯИОњзїгУЃЌЖдвЛаЉецОњЁЂдГцврОпгаЛюадЃЌЮЊЕжгљжТВЁадШыЧжЬсЙЉСЫЕквЛЕРЗРЯпЃл9⁃11ЃнЁЃетаЉЬьШЛКЭКЯГЩЕФЖрыФдкДѓаЁЁЂЕчКЩЁЂАБЛљЫсзщГЩЁЂЪшЫЎадКЭЖўМЖНсЙЙЩЯЖМВЛЭЌЃЌЦфжавЛаЉЖдИаШОХБдГцЕФКьЯИАћКЭАћФкМФЩњГцЯИАћФЄОпгабЁдёадзїгУЃл12ЃнЁЃПЙОњыФгХЯШЙЅЛїЯИАћФЄЕФзїгУЛњжЦЪЙЦфВЛвзВњЩњФЭвЉадЃЌвђДЫЃЌПЙОњыФГЩЮЊЖдПЙФЭвЉадХБдГцЕФКђбЁзЪдДЁЃЦљНёБЈЕРЕФПЙОњыФађСаГЌЙ§3 000ИіЃл13ЃнЃЌИљОнЦфЖўМЖНсЙЙЕФВЛЭЌЃЌЭЈГЃНЋПЙОњыФЗжЮЊІС⁃Тна§аЭПЙОњыФЁЂІТ⁃елЕўаЭПЙОњыФЁЂОпгаЛЗзДНсЙЙЕФПЙОњыФКЭЦЌВуНсЙЙПЙОњыФЁЃНќФъРДЃЌбаОПепУЧЗжРыКЭКЯГЩСЫЖржжОпгаПЙХБЛюадЕФПЙОњыФЁЊЁЊПЙХБЖрыФЃЌБОЮФДгПЙХБЖрыФЕФЗжРрКЭзїгУЛњжЦЖдЦфбаОПЯжзДНјааВћЪіЃЌВЂЬНЬжЦфзїЮЊаТаЭПЙХБвЉЮяУцСйЕФЬєеНКЭгІгУЧАОАЁЃ
1 ПЙХБЖрыФЗжРр
1.1 РДдДгкНкжЋЖЏЮяЕФПЙХБЖрыФ РЅГцИйЪЧЕиЧђЩЯзюДѓЕФЖЏЮяРрЃЌЪЧПЙОњыФЕФЙуЗКРДдДЃл14ЃнЁЃЦљНёЮЊжЙЃЌПЙОњыФЪ§ОнПтжаСаГіЕФ3 087ИіПЙОњыФжага305ИіРДдДгкРЅГцЃл13ЃнЁЃЦфжаЃЌРДздЬьВЯЖъЕФЬьВЯЫиBЪЧЕквЛИіБЛЗЂЯжОпгаПЙХБЛюадЕФПЙОњыФЃЌПЩИЩШХТбФвЗЂг§ЮЊцпзгЃЌЦфАыЪ§жТЫРМССПЃЈLD50ЃЉЮЊ0.5 ~ 1.0 ІЬg/ІЬLЃл15ЃнЁЃЫцКѓбаОПепШЫЙЄКЯГЩСЫЬьВЯЫиBЕФбмЩњЮяShiva⁃1КЭSB⁃37ЃЌЧАепгыЬьВЯЫиBОпга40%ЭЌдДадЃЌЕЋЛюадЪЧЬьВЯЫиBЕФ2БЖЃЛКѓепгыЬьВЯЫиBИпЖШЭЌдДЃЌЛюадЯрЕБЃл16ЃнЁЃVida1 ~ Vida3 ЪЧвЛРрКЯГЩЕФЬьВЯЫи B/ЗфЖОыФбмЩњЮяЃЌ100 µmol/L Vida3ЖўОлЬхЖдВЎЪЯХБдГцЖЏКЯзгвжжЦТЪДя100%ЃЌЭЌЕШХЈЖШЕФVida1 ~ Vida3ЕЅЬхЖдВЎЪЯХБдГцЖЏКЯзгвжжЦТЪвВПЩДяЕН70%зѓгвЃл17ЃнЁЃ
гааЉРЅГцБОЩэОЭЪЧвЛаЉМФЩњГцВЁЕФДЋВЁУННщЃЌШчАДЮУЪЧХБМВЕФДЋВЅУННщЃЌетаЉДЋВЅУННщдкНјЛЏЙ§ГЬжааЮГЩСЫЧПДѓЕФЗРгљЯЕЭГРДЕжгљаЏДјМФЩњГцЫљжТЕФЮЃКІЃЌШчЩЯЕївЛбѕЛЏЕЊКЯУИБэДяЁЂдкЖдХБдГцОпгаФЭЪмадЕФЮУзгжавжжЦМФЩњГцЗЂг§ЕФКкЛЏАќФвзїгУЃл18⁃19ЃнЃЌвдМАПЩФмдкетжжФЭЪмаджаЦ№живЊзїгУЕФПЙОњыФВњЩњЕШЁЃдкШіЙўРвдФЯЗЧжоЕиЧјЕФЖёадХБдГцжївЊУННщжаЗЂЯжСЫЗРгљЫиЁЂЬьВЯЫиКЭИдБШбЧУЙЫиЕШЖржжПЙОњыФЃл20⁃21ЃнЁЃИдБШбЧАДЮУПЙОњыФжївЊдкжаГІбЊЯИАћбљЯИАћжаБэДяЃЌдкИаШОХБдГц 2 h ФкЦфЫЎЦНЯджјЩ§ИпЁЃgambicin ЪЧРДдДгкИдБШбЧАДЮУЕФвЛжжОУтвпгеЕМЕФПЙОњыФЃЌжївЊБэДягкАДЮУЧАЮИЪвЁЂаиЧЛКЭИЙВПЃЌ10 µmol/L gambicinгыВЎЪЯХБдГцЬхЭтзїгУ24 hЃЌМДПЩЪЙЦфЖЏКЯзгаЮГЩЪ§НЕЕЭ 54.6%Ѓл21ЃнЁЃАЃМАвСЮУЕШЦфЫћЮУУНврПЩВњЩњОпгаПЙХБЛюадЕФПЙОњыФЃЌЦфПЩВњЩњ3жж40ИіАБЛљЫсГЄЖШЕФЗРгљЫиЃЈDefA⁃CЃЉКЭЬьВЯЫиAвдгІЖдЯИОњИаШОЁЃдкЙ§БэДяЬьВЯЫиAКЭЗРгљЫиAЕФзЊЛљвђАЃМАвСЮУжаЃЌМІХБдГцТбФвдіжГЪмЕНЯджјвжжЦЃл22ЃнЁЃ
вЛаЉРДдДгкЦфЫћМФЩњГцВЁРЅГцУННщЕФПЙОњыФврОпгаПЙХБЛюадЃЌДгКкИЙЙћгЌжаЗжРыЕФdrosomycinОЭЪЧЦфжажЎвЛЁЃЖд drosomycin МАЦфбмЩњыФНјааЕФПЙВЎЪЯХБдГцANKAГцжъХфзгЬхЗЂг§ЪдбщНсЙћЯдЪОЃЌЦфбмЩњыФdrosomycins⁃2дк20 µmol/LМССПЯТЯдЪОГі30%ЕФвжжЦзїгУЃЌЖјдаЭыФ drosomycin дкЯрЭЌХЈЖШЯТЯдЪОГі70%вдЩЯЕФвжжЦзїгУЃл23ЃнЁЃMtk⁃1КЭMtk⁃2ЪЧРДдДгкКкИЙЙћгЌЕФПЙОњыФЃЌдкНсЙЙЩЯЪєгкИЛКЌЬиЖЈАБЛљЫсЕФЦЌВуНсЙЙЁЃетСНжжПЙОњыФдкЬхЭтЖдЖёадХБдГц3D7ГцжъБэЯжГіНЯИпПЙХБЛюадЃЌдк50 µmol/LвЉЮяХЈЖШЯТДяЕН75% ~ 80%вжжЦТЪЃЌЧвСНепдкЬхЭтЖдДѓЪѓКЭжэКьЯИАћОљЮоШмбЊЛюадЃл24ЃнЁЃanoplinЪЧвЛжжДгЛЦЗфЖОвКжаЗжРыДПЛЏГіРДЕФПЙОњыФЃЌНігЩ10ИіАБЛљЫсзщГЩЃЌгыmas⁃toparan⁃X ОпгаИпЖШЭЌдДадЃЌдЄВтЦфЭЌЮЊСНЧзад ІС⁃Тна§НсЙЙЃЌЪЧЦљНёЮЊжЙдкздШЛНчжаЗЂЯжЕФзюаЁЯпадІС⁃Тна§ПЙОњыФЁЃCarter ЕШЃл25ЃнЗЂЯж anoplin ПЩвдЯджјвжжЦВЎЪЯХБдГцЖЏКЯзгаЮГЩЁЃЗфЖОыФЪЧвЛжжРДдДгкУлЗфЕФПЙОњыФЃЌгыanoplinЯрЭЌЃЌЗфЖОыФврОпгавжжЦВЎЪЯХБдГцЖЏКЯзгаЮГЩЕФзїгУЃЌдк 50 µmol/L ХЈЖШЯТзїгУ 30minМДПЩДяЕН100%вжжЦТЪЃл25⁃26ЃнЁЃscorpineЪЧвЛжжРДдДгкЗЧжоЕлЭѕаЋЕФРызгЭЈЕРзшжЭМСЃЌЖдВЎЪЯХБдГцХфзгКЭЖЏКЯзгОљгаЯджјвжжЦзїгУЃЌЦфАыЪ§гааЇМССПЃЈED50ЃЉЗжБ№ЮЊ10 µmol/LКЭ0.7 µmol/LЃл27ЃнЁЃ
РДдДгкЦфЫћНкжЋЖЏЮяЕФПЙОњыФврОпгаПЙХБЛюадЁЃвЛаЉРДдДгкБЭзггВђчЕФЗРгљЫидкЬхЭтОпгаПЙХБЛюадЃЌетаЉЗРгљЫидкНсЙЙЩЯЪєгкАыызАБЫсЮШЖЈЕФІС⁃Тна§КЭІТ⁃елЕўПЙОњыФМвзхЃЌдкЬхЭтПЩЯджјвжжЦЖёадХБдГцЩњГЄЃЌетжжвжжЦзїгУГЪМССПвРРЕадЃЌЦфжаDefMT2КЭ De⁃fMT5 дкЬхЭт 50 µmol/L вЉЮяХЈЖШЯТЖдЖёадХБдГцЕФвжжЦТЪПЩДяЕН65%КЭ85%Ѓл28ЃнЁЃЪѓХБФЃаЭЪЕбщНсЙћБэУїЃЌЫќУЧдкЬхФкЭЌбљОпгаПЙХБЛюадЃЌаЁЪѓИаШОЯФЪЯХБдГц24 hКѓОблПєКѓЕЅМССПЃЈ6 mg/kgЃЉзЂЩфИјвЉКѓ12 hЃЌгыЖдеезщЯрБШЃЌЖрыФДІРэзщаЁЪѓЬхФкдГцУмЖШЯджјНЕЕЭЃЌдкЬхЭтЖдЖёадХБдГцЮоПЙХБЛюадЕФDefMT6дкЬхФкврЯдЪОГіСЫвЛЖЈЕФПЙХБЛюадЃл29ЃнЁЃ
1.2 РДдДгкСНЦмЖЏЮяРрЕФПЙХБЖрыФ вЛаЉРДдДгкСНЦмЖЏЮяЦЄЗєЕФПЙОњыФЖдВЛЭЌЖёадХБдГцГцжъКьФкЦкОпгаПЙХБЛюадЁЃdermaseptinsЪЧДгЫїЭпвЖХнЭмЦЄЗєжаЗжРыГіРДЕФвЛРрдкНсЙЙКЭЙІФмЩЯЯрЙиЕФПЙОњыФМвзхЃЌЫќЪЧвЛжжЯпзДбєРызгЖрыФЃЌгЩ28 ~ 34ИіАБЛљЫсВаЛљзщГЩЃЌдкМЋадШмМСжавдСНЧзІС⁃Тна§аЮЪНДцдкЃЌжївЊЭЈЙ§НщЕМСНЧзІС⁃Тна§гыжЪФЄСзжЌЯрЛЅзїгУРДЗЂЛгЦфПЙХБЛюадЃЌЖдЖёадХБдГцКьФкЦкЖрИіЦкЬЌЦ№зїгУЃл30ЃнЁЃЦфжа dermaseptin S3ЃЈDS3ЃЉКЭ dermaseptin S4ЃЈDS4ЃЉдкЬхЭтЖдТШрУєИаГцжъHB3ОпгаПЙХБЛюадЃЌЦфАыЪ§вжжЦХЈЖШЃЈIC50ЃЉЗжБ№ЮЊ8.8 µmol/LКЭ2.2 µmol/LЁЃДЫЭтЃЌDS3КЭDS4ЭЌбљЖдТШрПЙадГцжъ Ibrahim БэЯжГівЛЖЈГЬЖШ ЕФ ПЙ ХБ Лю ад ЃЌIC50 жЕ Зж Б№ ЮЊ 1.5 µmol/ L КЭ 2.5µmol/LЃЛЕЋDS4дкЬхЭтОпгаШмбЊЛюадЃЌдкХЈЖШИпгк8.5µmol/LЪБПЩдьГЩКьЯИАћШмбЊЃл30ЃнЁЃЮЊНЕЕЭDS4ШмбЊЛюадЃЌбаОПШЫдБЩИбЁГі dermaseptin K4K20⁃S4ЁЂderma⁃septin K4⁃S4ЃЈ1⁃13ЃЉa КЭ dermaseptin NC7⁃P ЕШвЛЯЕСаDS4бмЩњЮяЃЌЯрЙибаОПБэУїетаЉбмЩњЮяПЙХБЛюадгыЦфздЩэаджЪгаЙиЃЌЪшЫЎадИќЧПЕФбмЩњЮяОпгаИќИпПЙХБЛюадЃЌЦфжавЛаЉбмЩњЮядкПЙХБЛюадгыШмбЊЛюаджЎМфБэЯжГіЯджјбЁдёадЃл31⁃32ЃнЁЃmagainin2ЪЧвЛжжДгЗЧжозІѓИжаЗжРыГіРДЕФгЩ23ИіАБЛљЫсзщГЩЕФПЙОњыФЃЌОmagain⁃in2жЮСЦЕФИаШОЪГаЗКяХБдГцЕФАДЮУГІТбФваЮГЩЪ§СПЯджјМѕЩйЃл15ЃнЁЃ
1.3 РДдДгкЮЂЩњЮяЕФПЙХБЖрыФ вЛаЉРДдДгкЮЂЩњЮяЕФПЙОњыФвВОпгаПЙХБЛюадЁЃЦфжаЃЌДгЫПзДецОњХрбјвКжаЗжРыГіРДЕФЛЗцпОњЫиAЪЧЕквЛИіБЛЗЂЯжОпгаПЙХБЛюадЕФЛЗзДПЙОњыФЃЌЦфВЛНідкЬхЭтЖдЖёадХБдГцОпгаПЙХБЛюадЃЌдкЬхФкЖдВЎЪЯХБдГцКЭдМЪЯХБдГцвВЭЌбљОпгаПЙХБЛюадЃл33ЃнЁЃthiostrepton ЪЧвЛжжРДдДгкСДЧђОњЕФЛЗзДПЙОњыФЃЌПЩЭЈЙ§ЕїНкЕААзУИЛюадКЭХБдГцЖЅЖЫИДКЯЬхЕААзЗвыРДвжжЦзгцпзгЛюадЃл34⁃35ЃнЃЌЦфIC50жЕЮЊ1.8 ~ 17 µmol/LЃл36ЃнЁЃbalgacycleamides A ~ CЪЧДгЭТЬЮЂФвдх EAWAG 251 жаЗжРыГіЕФ 3 жжаТДѓдгЛЗПЙОњыФЃЌЦфжа balgacycleamides A КЭ balgacycleamides B ЖдЖёадХБдГцТШрПЙадОњжъK1ОпгаПЙХБЛюадЃЌЦфIC50жЕЗжБ№ЮЊ 9.0 µmol/L КЭ 8.2 µmol/LЃЌВЂЧветаЉЛЗзДПЙОњыФдк150 µmol/LХЈЖШЯТЖдДѓЪѓГЩМЁЯИАћЯЕL6ЯИАћЮоЯИАћЖОадЃл37ЃнЁЃДЫЭтЃЌДгжВЮяШщИЫОњХрбјвКжаЗжРыДПЛЏЕФгЩ 4 жжыФзщГЩЕФПЙОњыФ LR14 вВОпгаПЙХБЛюадЃЌЕБЬхЭтвЉЮяХЈЖШДяЕН 42 ІЬg/mL ЪБЃЌЦфЖдЖёадХБдГцУєИаГцжъ3D7ЕФвжжЦТЪПЩДяЕН99%ЃЛЭЌЪБЃЌЦфЖдТШрПЙаджъ RKL19 врОпгаПЙХБЛюадЃЌЧветжжПЙОњыФдкЬхЭтВЛЛсдьГЩКьЯИАћШмбЊЃл38ЃнЁЃ
1.4 ЦфЫћКЯГЩЕФПЙХБЖрыФ NK⁃2ЪЧЪзРрОпгаПЙМФЩњГцЛюадЕФКЯГЩПЙОњыФжЎвЛЃЌгЩ39 ~ 65ИіАБЛљЫсВаЛљзщГЩЁЃгЋЙтБъМЧЙлВьЗЂЯжЃЌNK⁃2ПЩгыХБдГцМАЦфИаШОЕФКьЯИАћНсКЯЃЌЕЋВЛгыНЁПЕКьЯИАћНсКЯЃЌНјвЛВНжЄУїСЫ NK⁃2 ОпгабЁдёадСбНтЛюадЃл12ЃнЁЃD⁃HALO⁃rev вВЪЧвЛжжКЯГЩЕФПЙОњыФЃЌгЩ26ИіАБЛљЫсзщГЩЃЌдкЬхЭтЖдЖёадХБдГцгаПЙХБЛюадЃЌФмЙЛдкНЯЕЭХЈЖШЯТДЉЭИИаШОКьЯИАћВЂЩБЫРКьЯИАћФкМФЩњГцЃЌЦф IC50 жЕдМЮЊ 0.1ІЬmol/LЃл39ЃнЁЃIDR⁃1018ЪЧвЛжжКЯГЩЕФЬьШЛЗРгљЕїНкыФРрЫЦЮяЃЌгЩ12ИіАБЛљЫсВаЛљзщГЩЃЌОпгаУтвпЕїНкзїгУМАПЙОњЛюадЁЃетжжКЯГЩЖрыФЛЙгадЄЗРФдаЭХБзїгУЃЌдЄЗРадОВТізЂЩфКѓЃЌ56%ЕФХБдГцИаШОаЁЪѓПЩУтЪмФдаЭХБгАЯьЃл40ЃнЁЃзюНќЃЌвЛжжКЯГЩЕФІТЗРгљЫиDEFB130БЛЗЂЯждкЬхФкЭтЖМОпгаПЙХБЛюадЃЌетжжЗРгљЫидкНсЙЙЩЯЪєгк ІС⁃Тна§КЭ ІТ⁃елЕўЕФПЙОњыФМвзхЃЌдкЬхЭтЖдЖёадХБдГцЕФIC50жЕЮЊ43 ~ 49 µmol/LЃЛЦфбмЩњыФsDEFB130дкЬхЭтЖдЖёадХБдГцврОпгавЛЖЈПЙХБЛюадЃЌЕЋНЯдЖрыФЛюадНЯЕЭЃЌIC50жЕЮЊ90 ~ 93 µmol/LЃЌВЂЧветСНжжПЙОњыФдк200 µmol/LХЈЖШЯТЖМВЛЛсдьГЩКьЯИАћШмбЊЃл41ЃнЁЃЙњФкгабаОПЭХЖгКЯГЩСЫвЛжжРДдДгкЩпзщжЏЕААзУИЕФПЙОњыФLZ1ЃЌЦфЖдКьФкЦкЖёадХБдГцБэЯжГіСЫНЯЧПвжжЦзїгУЃЌIC50жЕЮЊ 3.045 µmol/LЃЛLZ1 ЖдВЎЪЯХБдГцврВњЩњСЫЯджјЕФЬхФкПЙХБЛюадЃЌЧвГЪЪБМфКЭМССПвРРЕадЃЌЭЌЪБЛЙЯдЪОГіПЙбззїгУЃЌПЩвдМѕЧсИЮЙІФмЫ№КІЃл42ЃнЁЃЦфЫћКЯГЩЕФОпгаПЙХБЛюадЕФПЙОњыФЛЙга pohlianin CМАЦф3ИібмЩњыФЁЃpohlianin CЪЧвЛжжИЛКЌИЪАБЫсЕФЛЗзДПЙОњыФЃЌдкЬхЭтЯдЪОГіжаЕШПЙХБдГцЛюадЃЌЦфРрЫЦЮяНсЙЙИќМгМђЕЅЃЌЕЋЦфПЙХБЛюадВЂЮДЪмЕНгАЯьЃЌЩѕжСЕУЕНСЫдіЧПЃл43ЃнЁЃ
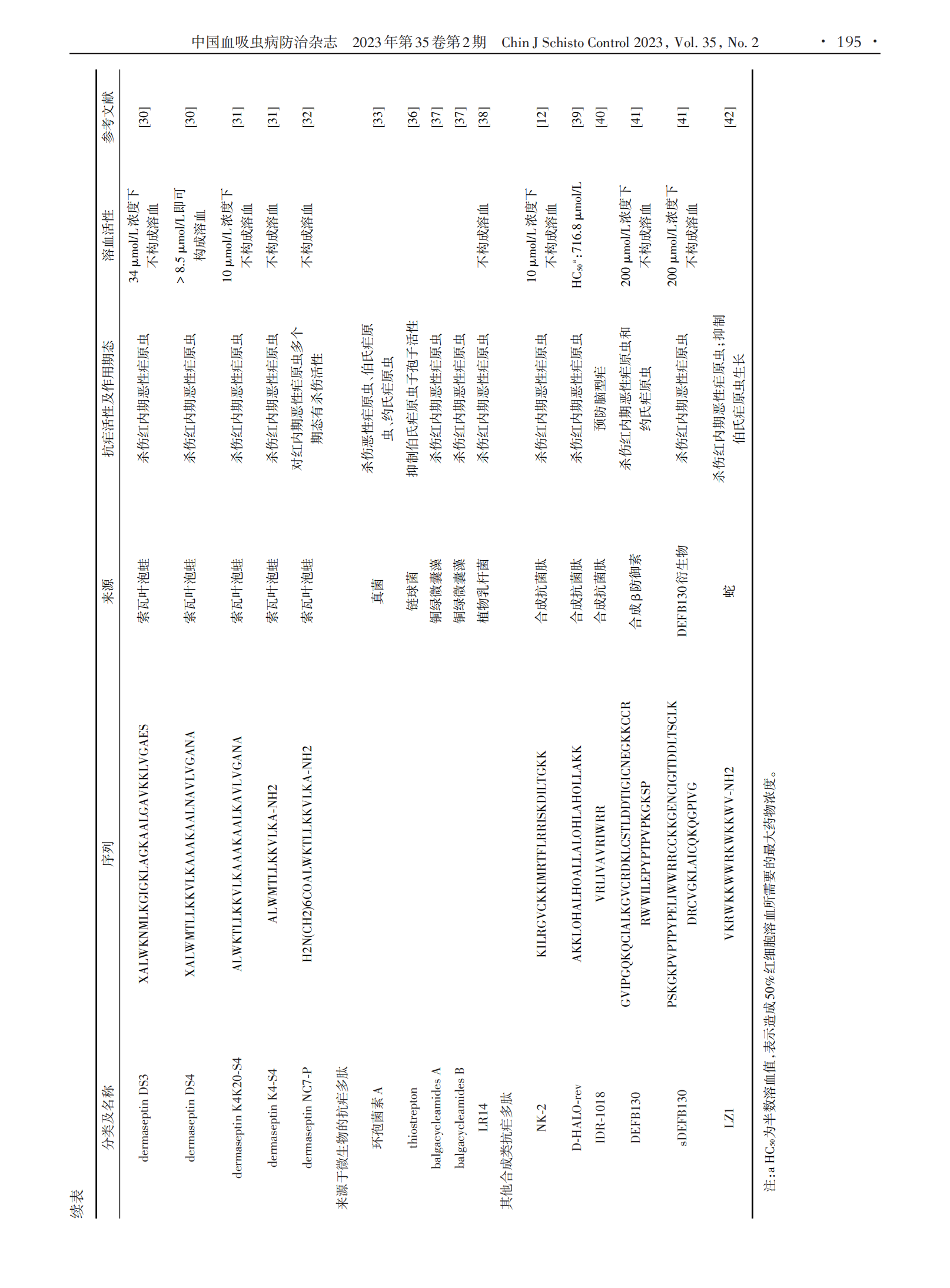
Бэ1ЖдвдЩЯЬсМАЕФПЙХБЖрыФађСаЁЂРДдДМАЯрЙиЛюадНјааСЫзмНсЁЃ
2 ПЙХБЖрыФзїгУЛњжЦ
ФПЧАПЙХБЖрыФЕФзїгУЛњжЦЩаЮДЭъШЋВћУїЃЌНсКЯПЙОњыФзїгУЛњжЦКЭвбгаПЙХБЖрыФбаОПЃЌЦфПЙХБЛњжЦжївЊАќРЈФЄЦЦЛЕЛњжЦКЭЗЧФЄЦЦЛЕЛњжЦЁЃ
2.1 ФЄЦЦЛЕЛњжЦ ПЙХБЖрыФЭЈГЃЪЧСНЧзадбєРызгЖрыФЃЌПЩвдгыДјИКЕчЕФМФЩњГцФЄЯрНсКЯЃЌЭЈЙ§ЦЦЛЕМФЩњГцФЄЭъећадЃЌдьГЩЯИАћФкШнЮяаЙТЉЃЌЕМжТХБдГцЫРЭіЃл44ЃнЁЃГ§СЫжБНгзїгУгкХБдГцФЄЃЌПЙХБЖрыФЛЙПЩвдЭЈЙ§зїгУгкИаШОХБдГцЕФКьЯИАћФЄРДЗЂЛгЦфПЙХБзїгУЁЃКьЯИАћБЛХБдГцИаШОКѓЃЌЦфЯИАћФЄЗЂЩњЯджјБфЛЏЃЌФЄЩЯСзжЌѕЃввДМАЗдіЖрЃЌЖјШмТбСзжЌКЭЧЪСзжЌКЌСПМѕЩйЃл45ЃнЃЌЧАепЭЈЙ§СзжЌѕЃЫПАБЫсЭбєШКЯГЩЃЌЪЧПЙХБЖрыФЕФзюжеАаБъЁЃгабаОПНвЪОСЫПЙОњыФNK⁃2ОпгаСзжЌѕЃЫПАБЫсвРРЕадСбНтзїгУЃЌетБэУїаЁбєРызгПЙХБЖрыФПЩФмЖдИаШОХБдГцЕФКьЯИАћОпгабЁдёадЃл12ЃнЁЃ
2.2 ЗЧФЄЦЦЛЕЛњжЦ Г§СЫФЄЦЦЛЕЛњжЦЃЌПЙХБЖрыФЛЙПЩвдЭЈЙ§гАЯьАћФкЯИАћЦїЁЂКЫЫсМАПЙОњЮяжЪЕФКЯГЩЕШЗНЪНРДЩБЩЫХБдГцЃл45ЃнЁЃШчthiostrepton ПЩЭЈЙ§ЕїНкЕААзУИЛюадКЭХБдГцЖЅЖЫИДКЯЬхЕААзЗвыРДДяЕНЩБЩЫХБдГцЕФаЇЙћЃл35ЃнЁЃГ§СЫжБНгЩБЫРХБдГцЭтЃЌПЙХБЖрыФЛЙЗЂЛгУтвпЕїНкЙІФмЃЌЫќУЧгЩАќРЈжаадСЃЯИАћКЭОоЪЩЯИАћдкФкЕФУтвпЯИАћЗжУкЃЌВЂВЮгыИаШОЕФЛКНтЙ§ГЬЁЃШчвЛаЉПЙХБЖрыФЭЈЙ§МѕЩйДйбзЯИАћвђзгЗжУкРДБмУтЯИАћвђзгЗчБЉКЭзщжЏЫ№ЩЫЁЂУтвпЯИАћФММЏКЭМЄЛюЁЂДйНјбЊЙмЩњГЩвдМАвжжЦЙ§СПЛюадбѕЪЭЗХЕШРДИФБфЫожїУтвпЗДгІЃЌДгЖјЕїНкбЯжиЪмЫ№ЦїЙйЫ№ЩЫЃл40, 46ЃнЁЃLZ1ПЩвдИФЩЦХБдГцИаШОаЁЪѓИЮдрЫ№ЩЫЃл42ЃнЃЌIDR⁃1018ПЩвдМѕЩйХБдГцИаШОв§Ц№ЕФЩёОбзжЂЃл40, 47ЃнЁЃ
3 ПЙХБЖрыФЕФгІгУЧАОАМАЦфУцСйЕФЬєеН
3.1 ПЙХБЖрыФЕФгІгУЧАОА ДЋЭГЕФПЙХБвЉДѓЖрЪЧаЁЗжзгЛЏКЯЮяЃЌгаЬиЖЈЕФАаБъЗжзгЃЌГЄЦкЪЙгУПЩФмЪЙЦфАаБъЗжзгЭЛБфДгЖјВњЩњПЙадЁЃЮЊбгЛКвЉЮяПЙадТћбгЃЌИДЗНПЙХБвЉВЛЖЯЭЦГіЃЌЕЋОјДѓЖрЪ§ИДЗНзщЗжОљЮЊвбжЊПЙХБвЉЃЌШчИДЗНняМзУбЕФжївЊзщЗжЪЧЧрняЫибмЩњЮяЁЂПЦЬЉИДЕФгааЇГЩЗжЪЧЫЋЧтЧрняЫиКЭСзЫсппрЃл48ЃнЁЃХБдГцЖдетаЉХфЮщгУвЉвбВњЩњЛђЖрЛђЩйПЙадЃЌвђДЫИУВпТдВЂВЛФмДгИљБОЩЯНтОідкШЋЧђПьЫйТћбгЕФПЙХБвЉПЙадЮЪЬтЁЃДЫЭтЃЌвВгавЛаЉзїгУЛњжЦВЛЭЌЁЂНсЙЙаТгБЕФаТвЛДњПЙХБвЉЮяВЛЖЯгПЯжЃЌБШШчФмЙЛвжжЦХБдГцЖўЧтШщЧхЫсЭбЧтУИЃЈPfDHODHЃЉЕФDSM265Ѓл49ЃнЃЌвдМАвжжЦPаЭбєРызгзЊдЫЬх ATP УИ 4ЃЈPfATP4ЃЉЕФ KAE609Ѓл50ЃнЃЌЛЙгаЦфЫћвЛаЉзїгУЛњжЦВЛУїШЗЕФЛЏКЯЮяЁЃЕЋетаЉЛЏКЯЮяРрвЉЮяЕФЙВадЖМЪЧеыЖдФГИіЬиЖЈАаБъЃЌЮоЗЈБмУтПЙадГіЯжЁЃвђДЫЃЌбАеваТаЭПЙХБвЉЮяЪЧСйДВХБМВжЮСЦКЭЗРжЙХБМВДЋВЅЕФЦШЧаашЧѓЃЌОпгаживЊЕФПЦбЇбаОПвтвхЁЃ
НќФъРДЃЌЖрыФзїЮЊжЮСЦвЉЮявбдНРДдНЪмЕНЙизЂЁЃЫцзХЩњЮяММЪѕгыЖрыФКЯГЩММЪѕЕФШеещГЩЪьЃЌдНРДдНЖрЕФЖрыФвЉЮяБЛПЊЗЂВЂгІгУгкСйДВЁЃЖрыФвЉЮяФПЧАвбЙуЗКгІгУгкжзСіЁЂЬЧФђВЁЁЂАЌзЬВЁЕШМВВЁдЄЗРКЭжЮСЦЃЌОпгаЙуРЋЕФПЊЗЂЧАОАЃл51ЃнЁЃЖјПЙОњыФзїЮЊУтвпЯЕЭГЕФЕквЛЕРЗРЯпЃЌПЩвдгааЇЪЖБ№ШыЧжЕФЭтдДВЁдЬхЃЌАќРЈМФЩњГцЁЃДЫЭтЃЌЩњЮяЬхЛсНјЛЏГівЛЯЕСаПЙОњыФЖдПЙВЁдЮЂЩњЮяИаШОЃл9ЃнЃЌвђДЫЃЌЬьШЛПЙОњыФЪЧКмКУЕФПЙХБЖрыФзЪдДЁЃЖрыФвЉЮяЕФжЪСППижЦЫЎЦННгНќгкаЁЗжзгЛЏКЯЮяЁЂЛюадНгНќгкЕААзжЪРрвЉЮяЃЌЖјЦфзїгУЛњжЦВЛЭЌгкаЁЗжзгЛЏКЯЮяЃЌВЛШнвзВњЩњПЙадЁЃДЫЭтЃЌдкСйДВгІгУКЭЩњВњжЦБИЩЯОпгаИпЛюадЁЂЕЭМССПЁЂЕЭЖОадЕШгХЕуЁЃ
3.2 ПЙХБЖрыФбаЗЂУцСйЕФЬєеН ЫфШЛФПЧАвбОЗЂЯжСЫДѓСПОпгаПЙХБЛюадЕФЖрыФЃЌЕЋетаЉЖрыФЕФПЙХБЛюадбаОПЩаДцдкОжЯоадКЭЦЌУцадЁЃШч gambicin дкАДЮУЬхФкБэЯжГіПЙХБЛюадЃЌЕЋЮУЬхФквЛаЉЬьШЛыФБОЩэОЭЛсгАЯьМФЩњГцЕФЩњДцзДЬЌЃЌвђЖјКмФбШЗЖЈДЫРрЖрыФЪЕМЪЕФПЙХБЛюадЃл21ЃнЁЃЖдгкзїгУгкЬхЭтХрбјЕФКьФкЦкХБдГцЖрыФЃЌЖрЪ§ШБЗІЬхФкгааЇадбщжЄЁЃетРрЖрыФЕФПЙХБЛюадашвЊИќЯЕЭГЕФбаОПЃЌвдШЗЖЈзюгааЇКЭзюОпбЁдёадЕФЖрыФНсЙЙЃЌВЂЧвашвЊдкЪѓХБФЃаЭжаНјаабщжЄЁЃДЫЭтЃЌзїгУЛњжЦЕФНвЪОНЋгажњгкжИЕМЖрыФЩшМЦКЭвЉЮябаЗЂЃЌЖјЖрЪ§ПЙХБЖрыФЕФзїгУЛњжЦЩаВЛУїШЗЁЃ
ЫфШЛЖрыФдкПЙвЉадЗНУцОпгагХЪЦЃЌЕЋФПЧАМЋЩйгаЩцМАМФЩњГцЖдЖрыФПЙадЕФбаОПЁЃЩаВЛШЗЖЈЖрыФдкХБдГцжаЪЧЗёИќЁАПЙвЉЁБЃЌетШЁОігкЖрыФПЙХБдГцЕФзїгУЛњжЦЁЃЩаВЛУїШЗЫќУЧЪЧЖдХБдГцЃЈЛђЪмИаШОЕФКьЯИАћЃЉЕФЯИАћФЄЗЂЛгЯрЖдЗЧЬивьадзїгУЃЌЛЙЪЧОпгаЦфЫћЬивьадАаБъЁЃВњЩњПЙвЉадХБдГцЕФЧуЯђБиаыЭЈЙ§ГЄЦкЪЕбщРДШЗЖЈЁЃДгЬхЭтЖЬЪБМфХрбјЛђФіГнЖЏЮяжаКмФбЗжРыКЭМјЖЈПЙвЉХБдГцЭЛБфЬхЃЌвдМАСйДВЪ§ОнЕФШБЗІЃЌЖМЪЧПЙХБЖрыФФЭвЉадбаОПжаУцСйЕФРЇФбЁЃ
ПЙХБЖрыФЕФбаЗЂГ§СЫгіЕНвдЩЯЦПОБЭтЃЌЖрыФРрвЉЮяЛЙгаЩњВњГЩБОИпЁЂИјвЉВЛБуЁЂЮШЖЈадВЛИпЕФШБЕуЁЃЕЋЫцзХЮДРДММЪѕЕФЗЂеЙгыИјвЉЭООЖЕФЗсИЛЃЌШЫУЧНЋЛсПЫЗўетаЉШБЕуЃЌВЂЧвНЋаТаЫыФРрММЪѕгІгУЕНжЦвЉжаЃЌАќРЈЖрЙІФмыФЁЂЯИАћДЉЭИыФКЭЖрыФХМСЊвЉЮяЃЌНЋДѓДѓЭиеЙжЮСЦадЖрыФвЉЮяЕФгІгУЗЖЮЇЁЃ
4 Нсгя
ЫфШЛЖрыФЕФСйДВгІгУЕНФПЧАЮЊжЙвЛжБЪмЕНЯожЦЃЌЕЋвЛЖЮЪБМфвдРДЃЌЖрыФвЛжБЪЧПЙОњКЭЦфЫћПЙОњЛЏКЯЮяЕФРДдДЁЃЦфжавЛаЉвЉЮягыЦфЫћПЙОњвЉЮяЯрБШОпгаЧБдкгХЪЦЃЌАќРЈзїгУЗЖЮЇЙуЁЂЩБУ№ЛюадПьЃЌвдМАДцдкКЯГЩКЭЩњЮяЗНЗЈРДЩњГЩжжРрЗБЖрЕФРрЫЦЮяЁЃвВаэзюЮќв§ШЫЕФЬиЕуЪЧЃЌЖдЖрыФЕФПЙвЉадГіЯжЕУКмТ§ЃЌЖјЖрыФвЉЮяЮДРДЕФЗЂеЙНЋМЬајНЈСЂдкЬьШЛыФЕФгХЪЦЩЯЃЌВЂгУЯжДњММЪѕНјааИФСМЃЌИФЩЦЦфЛЏбЇКЭЮяРэадФмШѕЕуЁЃвђДЫЃЌПЙХБыФзїЮЊаТаЭПЙХБвЉЮяЃЌПЩЭћЮЊПЙвЉадХБдГцжЮСЦДјРДаТЕФЯЃЭћЁЃ
Утд№ЩљУїЃКБОЮФЮЊаавЕНЛСїбЇЯАЃЌАцШЈЙщдзїепМАддгжОЫљгаЃЌШчгаЧжШЈЃЌПЩСЊЯЕЩОГ§ЁЃЮФеТБъзЂгазїепМАЮФеТГіДІЃЌШчашдФЖСдЮФМАВЮПМЮФЯзЃЌПЩдФЖСддгжО
