
’Σ“ΣΘΚ κΡ Χε «“Μ÷÷¥”ΥφΜζΑ±ΜυΥαΒΡκΡΈΡΩβ÷–…Η―Γ≥ωά¥ ,Ω…“‘ΗΏ«ΉΚœΝΠΒΊ”κΑ–Έο÷ ΧΊ“λ–‘ΫαΚœΒΡΕΧκΡ–ρΝ–. ΥϋΒΡ÷ς“ΣΫαΙΙΑϋά®ΚψΕ®ΒΡ’Ι Ψ÷ßΦήΒΑΑΉΦΑΆ®ΙΐΝΫΕΥœό÷Τ–‘≤ε»κΒΡΗΏ±δκΡΜΖ. ΫΆΡΗΥΪ‘”ΫΜΦΦ θ≥Θ”Ο”Ύ…Η―Γ’κΕ‘œΗΑϊΡΎΧΊ“λ–‘Α–ΒΑΑΉΒΡκΡ ΧεΙΐ≥Χ ,…Η―Γ≥ωΒΡκΡ ΧεΆ®ΙΐΧΊ“λ–‘ΫαΚœ Ε±πΑ–±ξΖΔΜ”άύΥΤΓΑΗ…»≈Μυ“ρΓ±ΒΡΉς”Ο¥”Εχ”ΑœλΒΑΑΉΒΡ…ζΈο―ßΜν–‘.
Χε (aptamer) ‘¥”Ύά≠ΕΓ”ο aptus ,Φ¥≈δΕ‘ΓΔ ”Π÷°“β , « άϊ ”Ο ÷Η ΐ ΗΜ Φ· ≈δ Χε ΒΡ œΒ Ά≥ Ϋχ Μ· ΦΦ θ( systematic evolution of ligands by exponentialenrichment ,SELEX) ¥”Κ§”–¥σΝΩ–ρΝ–ΒΡΥφΜζΈΡΩβ÷– ,Ά®ΙΐΕύ¥Έ―≠ΜΖ―Γ‘ώΗΜΦ· ,Μώ»ΓΥυ–η“ΣΒΡΧΊΕ®ΫαΙΙΜρΙΠΡήΒΡ…ζΈο ≈δΉ”. ΥϋΨΏ”–Ωβ»ίΝΩ¥σΓΔ…Η―ΓΖΕΈßΙψΓΔ”κΑ–Έο÷ «ΉΚΆΝΠΗΏΓΔΧΊ“λ–‘«ΩΒ»”≈Βψ ,≤Δ“‘ΤδΈ»Ε®ΓΔ“Ή–ό ΈΒ»”≈”ΎΩΙΧεΒΡΧΊ–‘‘ΎΜυ¥Γ―–ΨΩΓΔΝΌ¥≤’οΕœΓΔ“©Έο―–÷ΤΖΫΟφΒΟ“‘ΙψΖΚ”Π”Ο.
κΡ Χε(peptide aptamers , pepaptamers) «“Μ÷÷ΨΏ”–”κΚΥή’Υα ΧεœύάύΥΤ–‘÷ ΒΡΕΧκΡ–ρΝ–:Φ¥άϊ”Ο SELEXΦΦ θ¥”»ΥΙΛΚœ≥…ΒΡΥφΜζΑ±ΜυΥακΡΈΡΩβ÷–…Η―Γ≥ωΒΡΩ…ΧΊ“λ–‘ΓΔΗΏ«ΉΚœΝΠΫαΚœΑ–Έο÷ ΒΡΗ¥ΚœΒΑΑΉ[1]. ≤ΔΆ®Ιΐ”κΑ–Έο÷ Ε±πΫαΚœ ,ΖΔΜ”ΤδΧΊΕ®ΒΡ…ζΈο―ßΙΠ–ß.
1 κΡ ΧεΒΡΙΠΡήΜυ¥Γ
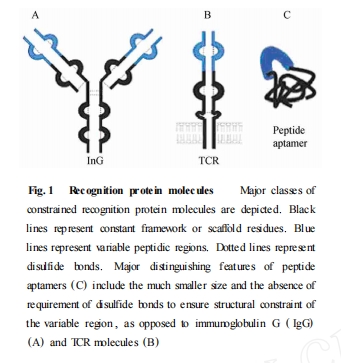
κΡ Χε“≤Ω…≥ΤΈΣ”’Εϋ (decoys) ΜρΜ·―ßΩΙΧεΖ÷Ή”(chemical antibodies) , «“Μ÷÷–¬Ϋϋ≥ωœ÷ΒΡ…ζΈοΙΠΡή–‘Ζ÷Ή”[2 ]. Ήν‘γΕ‘”ΎκΡ ΧεΒΡ…ηΦΤΝιΗ–ά¥‘¥”ΎΟβ“Ώ«ρΒΑΑΉΚΆ TœΗΑϊ ήΧεΥυΙ≤”–ΒΡ±Θ Ί«χΩρΦήΫαΙΙΚΆΗΏΕ»ΧΊ“λ–‘ΒΡΩ…±δ«χκΡΕΈΫαΙΙ. Τδ…ηΦΤ‘≠άμ «ΫΪ¥σΝΩ»ΥΙΛΚœ≥…ΒΡΥφΜζκΡ–ρΝ–≤ε»κΓΔ’ϊΚœ”ΎΒΑΑΉ÷ßΦή (proteinscaffolds) ÷°…œ , άϊ”ΟΥφΜζ«χ”ρΒΡ–≈Κ≈ΡΘ–ρ (signalmotif) ”κΧΊ“λ–‘Α–ΒΑΑΉ÷°ΦδΒΡœύΜΞΉς”Ο ,Ά®ΙΐΕύ‘ΣΜ·ΒΡΖ÷Ή”…ζΈο―ß’Ι ΨΦΦ θ (display technique) ,…Η―ΓΒΟΒΫ’κΕ‘Α–ΒΑΑΉΒΡΗΏ«ΉΚΆΝΠΓΔΗΏΧΊ“λ–‘ΒΡκΡ Χε. Υϋ «“ΜάύΡή‘ΎΧεΡΎ“‘ΧΊΕ®»ΐΦΕΫαΙΙΈ»Ε®¥φ‘ΎΒΡΕΧκΡ–ρΝ– ,ΗΏΕ»Ω…±δΒΡΙΠΡή–‘ΫαΙΙ”ρ (functional structuredomain ) «…Η―ΓΒΡΈο÷ Μυ¥Γ. ΥφΜζ–ρΝ–Ω…±δ«χ”ρ”κ÷ßΦήΒΑΑΉΝΫΕΥœύΜΞΉς”ΟΒΡ Ε±πΓΔ»ΎΚœΈΜΒψΙΙ≥…ΝΥκΡ ΧεΒΡΥΪ÷Ίœό÷Τ–‘ ,”–άϊ”Ύ…Η―ΓΚσΒΡ–ό ΈΓΔ±δΙΙ[3 ]. ”κΉ‘»Μ¥φ‘ΎΒΡΑ±ΜυΥακΡΤ§ΕΈœύ±» ,»ΥΙΛΚœ≥…ΒΡκΡ Χε÷ΜΑϋΚ§ΒΞ“Μ±δΝΩΒΡκΡΜΖ (loop) ΫαΙΙ ,«“ΝΫΕΥΒΡΝ§Ϋ”œό÷Τ–‘“≤≤Μ“άάΒ”ΎΕΰΝρΦϋ ,Εχ «άϊ”Ο÷ßΦήΒΑΑΉ≤–ΜυΒΡΕ®“εΈΜΒψœύΜΞ Ε±πΝ§Ϋ”ΒΡ ,”–άϊ”ΎΧαΗΏ‘ΎΜζΧεΡΎΒΡΈ»Ε®–‘[4 ](Fig. 1 ) .
2 κΡ ΧεΒΡ…Η―Γ‘≠άμΦΑΦΦ θ¬ΖœΏ
κΡ ΧεΒΡ…Η―ΓΝς≥Χ «“Μ÷÷Ήώ―≠―œΗώœό÷Τ–‘ΒΡΡΘΩι ΫΒΡΫχΜ·ΗΜΦ·Ιΐ≥Χ. ΡΩ«Α÷ς“Σ”– 3 ÷÷…ζΈο―ß’Ι ΨΖΫΖ®”Ο”Ύ…Η―Γ :άϊ”ΟΫΆΡΗΥΪ‘”ΫΜΦΦ θ¥”œΗΑϊΡΎ…Η―Γ≥ωΡή”κΉςΈΣΓΑ”ψΕϋΓ±ΒΡΒΑΑΉΑ–Έο÷ ΧΊ“λ–‘ Ε±πΒΡκΡ Χε ,ΖΫΖ®Φρ±ψΓΔΩλΫί[5 ];Ά®Ιΐ …ΨζΧε’Ι ΨΦΦ θ”≈Μ·ΩΙΧεΓΔΒΑΑΉΜρ «’κΕ‘ΜνœΗΑϊΒΡκΡ Χε…Η―Γ[6 ];≤…”ΟΡφΉΣ¬Φ≤ΓΕΨ’Ι ΨœΒΆ≥Ϋ®ΝΔΕύ÷÷œΗΑϊ±μ–ΆœύΙΊΒΑΑΉΒΡ…Η―ΓΤΫΧ®[7 ]. œ÷ΨΆ“‘ΡΩ«Α Ι”ΟΉνΙψΖΚΒΡΫΆΡΗΥΪ‘”ΫΜœΒΆ≥ΈΣάΐΫι…ήκΡ ΧεΒΡ…Η―ΓΙΐ≥Χ.
Ήœ» ,ΙΙΫ®Α±ΜυΥακΡΈΡΩβ :ΫΪ“ΜΕΈ≥ΛΕ»‘ΦΈΣ 60bp ΒΡΥφΜζΙ―ΚΥή’ΥαΤ§ΕΈ≤ε»κ±ύ¬κ÷ßΦήΒΑΑΉΒΡΩΣΖ≈‘ΡΕΝΩρ ,άϊ”ΟΝΫΕΥΒΡΗΏΕ»œό÷Τ–‘ΜΞœύ Ε±π»ΎΚœ , Ι÷ ΝΘ‘ΊΧεΉΣΒΦΚσΩ…“‘Έ»Ε®±μ¥οΚ§”–¥σ‘Φ 20 ΗωΥφΜζΑ±ΜυΥα–ρΝ–ΒΡ÷ßΦήΒΑΑΉ2κΡΈΡΩβΗ¥ΚœΈο (Fig. 2) . άμ¬έ…œ ,20 ΗωΥφΜζΑ±ΜυΥα–ρΝ–Ω…Ά®ΙΐΙΛ≥ΧΨζ÷ξ≤ζ…ζ‘Φ2 ΓΝ10ΒΡ8¥ΈΖΫΗω≤ΜΆ§κΡΕΈΫαΙΙΒΡΩΥ¬ΓΉ” ,ΉςΈΣ…Η―ΓΧΊ“λ–‘Α–Ζ÷Ή”ΒΡ≥θΦΕΈΡΩβ. Ά®Ιΐ≤ΜΆ§ΒΡ–ό ΈΩ…ΧαΗΏκΡ ΧεΒΡΈ»Ε®–‘ ,‘ωΦ”…ζΈοάϊ”ΟΕ» ,»γ‘Ύ±ύ¬κκΡ ΧεΒΡ≤ύ“μ–ρΝ–¥Π“ΐ»κΝ§Ϋ”ΒΑΑΉΉΣΒΦ”ρ (protein transductiondomain ,PTD) ,”–άϊ”ΎκΡ Χε‘Ύ…Η―ΓΙΐ≥Χ÷–ΒΡΆβ‘¥–‘…ψ»Γ[9 ]. ΡΩ«Α ,Ήν≥ΘΦϊΒΡ PTD «ά¥‘¥”Ύ HIV2TAT ΒΑΑΉΒΡ–·¥χ’ΐΒγΚ…ΒΡ±ύ¬κ–ρΝ–[10 ]Θ§ΥϋΩ…”κ÷ ΡΛ±μΟφ–·¥χΗΚΒγΚ…ΒΡΝρΥαΗΈΥΊΤœΨέΧ«œύΜΞΉς”ΟΫιΒΦ»ΎΚœΒΑΑΉΒΡœΗΑϊΡΎ…ψ[11-13 ].

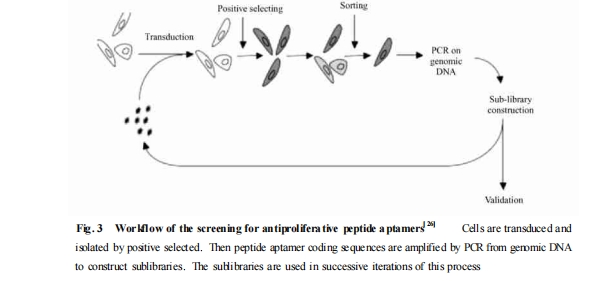
Τδ¥Έ ,ΫΆΡΗΥΪ‘”ΫΜœΒΆ≥…Η―ΓκΡ Χε :“ΜΑψΒΡΫΆΡΗΨζ÷ξΨυ¥χ”– 2 Ηω“‘…œΒΡ―Γ‘ώ–‘”Σ―χ»±œίΜυ“ρ ,Α–ΒΑΑΉΉςΈΣΓΑ”ψΕϋΓ±ΒΑΑΉ (baits) ”κΉΣ¬Φ“ρΉ”ΒΡ DNA ΫαΚœ”ρœύ»ΎΚœ ,ΥφΜζΑ±ΜυΥαΈΡΩβΉςΈΣΓΑ≤ΕΜώΓ±ΒΑΑΉ (preys)Κρ―ΓΈΡΩβ”κΉΣ¬ΦΦΛΜν“ρΉ”»ΎΚœ ,¥ΌΫχΜυ“ρΒΡΜνΜ·±μ¥ο. »γΙϊΈΡΩβ÷–¥φ‘Ύ”κΑ–ΒΑΑΉœύΕ‘”ΠΒΡκΡ Χε ,‘ρΡήΆ®Ιΐ–Έ≥…Η¥ΚœΈο ,“ΐΖΔ DNA ΫαΚœ”ρΝΎΫϋ«χ”ρΒΡΙΙœσ±δΜ· ,ΉΣ¬Φ“ρΉ”ΜνΜ·¥”Εχ÷±Ϋ”ΒΦ÷¬ΝΥœ¬”Έ±®ΒάΜυ“ρΒΡΉΣ¬Φ±μ¥ο. Χτ»Γ―τ–‘Ψζ÷ξΧα»ΓΡΩΒΡ DNA Τ§ΕΈ ,άϊ”Ο‘Λœ»…ηΦΤΒΡ“ΐΈοΫχ–– PCR ά©‘ω ,≤ζΈοΦΧ–χ≤ε»κ÷ßΦήΒΑΑΉ–ρΝ– ,Ϋ®ΝΔκΡ ΧεΒΡ¥ΈΦΕΗΜΦ·ΈΡΩβ. Ά®Ιΐ ΐ¬÷…Η―ΓΚσ ,κΡ Χε”κΑ–Έο÷ Ζ¥”ΠΒΡ«ΉΚΆΝΠΓΔΧΊ“λ–‘Ψυ”–Οςœ‘ΧαΗΏ ,…Η―ΓΙΐ≥Χ»γ Fig. 3 Υυ Ψ. άϊ”Ο“‘”’ΒΦΤθΚœΓΔ»»ΒψΝλΚΫΓΔΒγΖϋ»ΠΚœΈΣΜυ¥ΓΒΡΟή¬κΤ·“ΤΦΦ θ(codon2shuffling) Ϋ®ΝΔΒΡΈΡΩβΩ…“‘÷±Ϋ”Ε‘Έ¥÷ΣΒΑΑΉΫχ––ΙΠΡήΖ÷Έω[14 ]. Rao Β»[15 ] Ά®Ιΐ‘Λœ»…ηΦΤΒΡΒγΚ…ΤΪ“–…Η―Γ≥ω’κΕ‘Ζ÷÷ΠΗΥΨζΒΡΙΠΡήΒΑΑΉ HupB ΒΡκΡ Χε ,Ω…‘ΎΧεΆβ÷±Ϋ”ΡΘΡβ HupB ΒΡΕΰ‘ΣΫαΙΙ ,ΈΣΡΆ“©–‘Ζ÷÷ΠΗΥΨζΒΡ“÷Ψζ―–ΨΩΧαΙ©ΝΥΜυ¥Γ.
3 κΡ ΧεΒΡΧΊ–‘
3. 1 κΡ ΧεΕύ‘Σ…ζΈοΜν–‘
”κΜυ“ρ«Ο≥ΐΓΔRNA Η…»≈ΦΦ θ[16 ] œύ±» ,κΡ Χε“‘ΤδΨΪœΗΒΡΒςΩΊΜζάμ‘Ύ―–ΨΩΗςΧθ–≈Κ≈ΉΣΒΦΆΨΨΕ÷–“ΐΤπΙψΖΚΒΡΙΊΉΔ. ΥϋΕ‘Ά§‘¥Α–Έο÷ ΨΏ”–ΦΪΗΏΒΡΧΊ“λ–‘ ,“ρ¥ΥΩ…”Ο”Ύ«χΖ÷Ά§“ΜΒΑΑΉΦ“ΉεΒΡ≤ΜΆ§≥…‘± ,…θ÷Ν «≤ΜΆ§Β»ΈΜΜυ“ρ±δ“λ≤ζ…ζΒΡ“λΙΙΧε ,ΤδΉς”ΟΒΡΜζ÷ΤΈΣΖβ±’ΒΑΑΉΒΡΨέΚœΈΜΒψ ,“÷÷Τ–≈Κ≈Ζ÷Ή”ΒΡ‘Υ δΓΔΜνΜ·ΓΔ¥ζ–Μ ,“‘ΦΑΗς÷÷ΟΗΉς”ΟΒΉΈοΒΡ¥ΏΜ·ΦΛΜν[17 ]. Zhao Β»[18 ]…Η―Γ≥ωΧΊ“λ–‘ΫαΚœ Smad224 , Smad7 ΒΡ SARA (Smadanchor for receptor activation) κΡ Χε ,”Ο”Ύ“÷÷ΤΉΣΜ·…ζ≥Λ“ρΉ”(TGF) ΫιΒΦΒΡ–≈Κ≈ΉΣΒΦΚΆ…œΤΛœΗΑϊœρΦδ÷ œΗΑϊΉΣΜ·(epithelial2to2mesenchymal transitionby , EMT) ΒΡ…ζάμΙΐ≥Χ.TGF ΦΛΜνΆΨΨΕ «”κΧεΡΎΧΊ“λ–‘≈δΧεΫαΚœΚσ ,ΦΛΜν ήΧεΦΛΟΗ ,“ΐΤπ Smad2 ΚΆ Smad3 ΒΡΝΉΥαΜ·[18 ].―–ΨΩΖΔœ÷ , Β―ι÷–κΡ Χε÷ΜΡή”κΝΉΥαΜ·ΒΡSmad224 ΧΊ“λ–‘ΫαΚœ ,”κΈόΜν–‘ Smad224 ΫαΚœΚή…Ό ,’βΩ…Ρή «”…”Ύ Smad Ψ≠ΝΉΥαΜ·Κ󱩬Ε≥ω Smad ΟΣΕ® ήΧεΡή”κ…Η―Γ≥ωΒΡ SARA κΡ ΧεΆ®ΙΐΫαΚœΡΘ–ρœύΜΞ Ε±πΓΔΫαΚœ ,¥”Εχ“÷÷ΤΕΰΨέΜ· ,ΉηΑ≠ΝΥ TGF ΫιΒΦΒΡΜυ“ρ±μ¥ο. Smad7 «“Μ÷÷ΗΚ–‘ΒςΫΎ“ρΉ”[19 ],SARA κΡ Χε”κ Smad7 ΒΡΫαΚœ”κ«ΑΫι…ήΒΡ”–Υυ≤ΜΆ§ , Υϋ≤Μ ήSmad7 ΝΉΥαΜ·Υ°ΤΫΒΡ”Αœλ ,“ρ¥Υ SARA κΡ ΧεΩ…“‘”Ο”ΎΦλ≤β–≈Κ≈ΉΣΒΦΆΨΨΕ÷– Smad ΒΡΜν–‘Υ°ΤΫ.
κΡ Χε”κœ÷”–ΒΡΖ÷Ή”…ζΈο―ßΦλ≤βΦΦ θœύ±» ,”≈ Τ‘Ύ”ΎΩ…“‘÷±Ϋ”ΦχΕ®’φΚΥœΗΑϊ‘ΊΧεΥυ±μ¥οΒΡΑ–±ξΒΑΑΉ ,Έό–η¥ΩΜ· ,ΈΣΓΑ¬ΏΦ≠“≈¥Ϊ―ßΓ±(logic of genetics) ΒΡ―–ΨΩΖΔ’ΙΒλΕ®ΝΥΦα ΒΒΡΜυ¥Γ. κΡ Χε «“Μ÷÷ΡΎ‘¥–‘ΒΡΕύκΡΤ§ΕΈ ,Ά®Ιΐ”κΧΊ“λ–‘Α–ΒΑΑΉΧΊ“λΙΠΡήΈΜΒψΫαΚœΒςΩΊΜυ“ρ±μ¥ο¥”ΕχΡΘΡβΓΑΗ…»≈Μυ“ρΓ±(perturbogens)ΒΡΉς”Ο[20 ]. de Chassey Β»[8 ] …Η―Γ≥ωΝΥ“Μ÷÷Ω…“‘“÷÷Τ÷ΉΝωœΗΑϊ‘ω÷≥ΒΡ R5G42 κΡ Χε ,Ζ÷ΈωΤδΜζ÷ΤΩ…Ρή «Ά®ΙΐκΡ ΧεΧΊ“λ–‘ Ε±πΉΣ¬Φ“ρΉ”ΒΡΙ≤ΒςΩΊΒΑΑΉ ,Εΰ’ΏΫαΚœΚσ–Έ≥…“ΜΕ®ΒΡΩ’ΦδΈΜΉη , ÷±Ϋ”ΜρΦδΫ”ΒΡ“÷÷ΤΜυ“ρ±μ¥ο ,¥”Εχ“÷÷ΤΝΥ÷ΉΝωœΗΑϊΒΡ‘ω÷≥ΓΔΖ÷Μ·. AbedΒ»[21 ] ―–÷ΤΒΡκΡ Χε F4 ,’κΕ‘ΤδΑ–ΒΑΑΉ Fur (ferricuptake regulator) ΨΏ”–Οςœ‘ΒΡ“÷÷ΤΉς”Ο ,≤ΔΆ®Ιΐ”κΜυ“ρ«Ο≥ΐΒ»ΦΦ θ±»Ϋœ ,±μΟςκΡ Χε‘ΎΕύΙΠΡήΒΑΑΉΒΡ―Γ‘ώ–‘“÷÷ΤΖΫΟφΨΏ”–ΧΊ βΒΡΉς”Ο. κΡ Χε≤ΜΒΪΨΏ”–“÷÷ΤΉς”Ο ,Ε‘Ρ≥–©Α–ΒΑΑΉΕχ―‘ ,κΡ ΧεΜΙ «“Μ÷÷”––ßΒΡΦΛΜνΦΝ.
3. 2 κΡ ΧεΒΡΈ»Ε®–‘ΚΆ¥©ΆΗ–‘
κΡ Χε‘ΎΧεΡΎΩ…’έΒΰ–Έ≥…Έ»Ε®ΒΡ»ΐΦΕΙΙœσ ,”κ“ΜΑψ”ΈάκΒΡΑ±ΜυΥαœύ±»ΫΒΒΆΝΥ»»ΝΠ―ßΒΡλΊ÷Β ,”–άϊ”Ύ±Θ≥÷ΫœΗΏΒΡ…ζΈο―ßΜν–‘. ΥφΉ≈Μυ“ρΉι―ßΚΆΒΑΑΉ÷ Ήι―ßΝλ”ρΒΡ―–ΨΩΫχ’Ι ,»ΥΙΛ±μ¥οΒΑΑΉΒΡΈ»Ε®–‘ΚΆ≤ζ¬ ΕΦ‘Ύ≤ΜΕœΧαΗΏ. œ÷“―”–Εύ÷÷–ΓΤ§ΕΈ ,“Ή±μ¥οΒΡ÷ßΦήΒΑΑΉ±ΜΙψΖΚ”Ο”ΎκΡ ΧεΝλ”ρ[22 ]. ΥϋΟ«ΗςΉ‘ΒΡ…ζΈο―ßΧΊ–‘”–άϊ”ΎκΡ Χε”Π”ΟΒΡΫχ“Μ≤ΫΖΔ’Ι ,»γ‘Ύ±μ¥οΙΐ≥Χ÷–ΡήΆ®ΙΐΩ’Φδ’έΒΰ–Έ≥…±ΘΜΛΙΠΡή–ß”Π”ρ ,”κ–ß”ΠΖ÷Ή”ΫαΚœΚσ ,Ά®Ιΐ±δΙΙ–ß”Π±©¬ΕΜν–‘ΙΠΡή–ß”Π”ρ ,¥οΒΫΕ‘κΡ ΧεΒΡ±ΘΜΛΉς”Ο. ΡΩ«Α Ι”ΟΫœΙψΖΚΒΡ «Νρ―θΜΙΒΑΑΉ A(thioredoxinA ,TrxA) ,άϊ”Ο TrxA ΉςΈΣœό÷Τ–‘κΡ ΧεΒΡ÷ßΦή”–άϊ”ΎΧαΗΏκΡ Χε”κ…Η―ΓΑ–Έο÷ ΫαΚœΒΡ«ΉΚΆΝΠ. Ψ≠―–ΨΩ÷ΛΟς ,Colas Β»[23 ] …Η―Γ≥ωΒΡ“Μ÷÷”Ο”Ύ“÷÷Τ“άάΒœΗΑϊ÷ήΤΎ–‘ΦΛΟΗ 2 (cdk2) ΒΡ TrxA2κΡ ΧεΗ¥ΚœΈο ,ΤδΑκ“÷÷Τ≈®Ε» ( IC50) ±»Τ’Ά®”ΈάκΉ¥Χ§œ¬“ΣΒΆ 50ΓΪ1 000±Ε. Ηϋ÷Ί“ΣΒΡ « ,κΡ Χε»ΎΚœ”Ύ Trx ÷ßΦήΒΑΑΉ…œ”–άϊ”ΎΤδ‘Ύ≤Η»ιΕ·ΈοœΗΑϊ÷–ΒΡ±μ¥ο. CohenΒ»[24 ]ΙΙΫ®ΝΥάϊ”ΟΙ«»βΝωœΗΑϊ (Saos22) ±μ¥οΒΡ TrxA2κΡ ΧεΩ…“‘”––ßΒΊ“÷œΗΑϊ‘ω÷≥ΚΆΖ÷Μ·. ≤Δ“‘¥ΥΈΣΜυ¥Γ―–ΖΔ≥ωΩ…”κ Mdm2 Μυ“ρ±ύ¬κΒΡ–Ω÷ΗΒΑΑΉΧΊ“λ–‘ΫαΚœΒΡ TrxA2κΡ Χε ,”Ο”ΎΜνΜ· p53 ΟτΗ–ΒΡ±®ΒάΜυ“ρΨΏ”–ΝΦΚΟΒΡ–ßΙϊ[25 ]. Cui Β»[26 ]…Η―Γ≥ωΒΡ“‘ TrxA ΉςΈΣ÷ßΦήΒΑΑΉ≤Δ‘Ύ xFoxH1b ΚΆ Lef1 ÷–ΫœΚΟ±μ¥οΒΡκΡ Χε ,Ω…”κ Smads ΧΊ“λ–‘ΫαΚœ≤Δ―Γ‘ώ–‘ΒΡ“÷÷Τ TGF ΟτΗ–ΒΡ±®ΗφΜυ“ρ.
ΨΏ”–œΗΑϊ¥©ΆΗ–‘ΒΡΑ±ΜυΥακΡΕΈ“ΜΑψΈΣΦφ–‘Α±ΜυΥα ,ΥϋΟ«Ά®Ιΐ”κΉΣ‘Υ‘ΊΧεΙ≤ΦέΝ§Ϋ”ΫιΒΦΤδ‘ΎΑϊΡΎΒΡΉΣ‘Υ. Matsushita Β»[27 ]…ηΦΤΝΥ“Μ÷÷–¬–ΆΒςΩΊΖ÷ΟΎΒΡ…ζΈοΩΣΙΊ. œ»‘ΎΕύκΡΤ§ΕΈΒΡ N2Ρ©ΕΥ…œΝ§Ϋ” 1 Ηω±Θ≥÷œΗΑϊ¥©ΆΗ–‘ΒΡ–ρΝ– ,Εχ‘Ύ C Ρ©ΕΥΙΧΕ®“ΜΗω“÷÷ΤΖ÷ΟΎΙΠΡήΒΡ–ρΝ– ,Ά®ΙΐΦΛΜν≤ΜΆ§–ρΝ–ΒΡΉΣ¬ΦΜν–‘ ,ΜνΜ·Μρ“÷÷ΤΖ÷ΟΎΙΠΡή. ≤Δ“‘¥ΥΈΣΜυ¥Γ―–÷Τ≥ωΝΥœΗΑϊ¥©ΆΗ–‘κΡ Χε(cell2penetrating aptamer peptides CPAPs) . Εύ÷÷ά¥‘¥ΒΡ CPAPs Ω…”κ≤ΜΆ§…ζΈο―ßΜν–‘ΒΡ–ό ΈΜυΆ≈Ι≤ΦέΝ§Ϋ” ,ΫιΒΦΤδ‘ΎΑϊΡΎΉΣ‘Υ≤ΔΧΊ“λ–‘ Ε±πΑ–Έο÷ ,ΖΔΜ”ΉςΈΣΑ–œρΖ÷Ή”ΒΡ…ζΈο―ßΉς”Ο[28 ]. Green Β»[29 ] άϊ”ΟCPAPs ΙΙΫ®ΝΥ―–ΨΩΒΑΑΉΉΣΒΦΙΠΡή”ρΒΡ…ζΈο―ßΡΘ–Ά.Trehin Β»[30 ]ΫΪ CPAPs ”Ο”ΎΑ–œρΕ®ΈΜ“©Έο‘ΎΧεΡΎΉΣ‘ΥΓΔ¥ζ–ΜΒΡ―–ΨΩΜώΒΟΝΥΝΦΚΟΒΡ≥…–ß.
4 κΡ ΧεΒΡ”Π”Ο
κΡ ΧεΆ®Ιΐ”κ≈δΜυΒΡΧΊ“λ–‘ Ε±π ,ΈΣΗΏ÷Ο–≈œόΥ°ΤΫΑ–±ξ»Ζ»œΧαΙ©ΝΥ”–άϊΒΡΙΛΨΏ. Ζ÷ΈωκΡ Χε”κΡΩΒΡΒΑΑΉ÷°ΦδΒΡΨΪœΗΒςΫΎΩ…‘ΎΚρ―ΓΒΑΑΉ»Κ÷–…Η―Γ≥ω“β“εΒΑΑΉ ,ΈΣ–¬“©―–ΖΔ÷–ΒΡ“©ΈοΑ–œρΈΜΒψ»ΖΕ®ΧαΙ©ΝΥ
“ΜΗω–¬ΒΡΦΦ θΤΫΧ®. œ÷“―”–“Μ÷÷κΡ Χε”ΓΦ«ΦΦ θ(AptaPrintTM)[3 ]Ω…”Ο”ΎΩλΥΌΑ–œρΕ®ΈΜ (Fig. 4) . Ήœ»Ά®ΙΐΖ÷ΈωΑ–ΒΑΑΉΒΡ…ζΈο―ßΜν–‘ ,Ε®ΈΜΧΊ’ς–‘ΙΠΡή”ρΜρ «ΙΙΫ®Ά§‘¥–‘ΡΘ–Ά ,»γ :ΟΗΒΡΜν–‘ΈΜΒψΓΔΫιΒΦΒΑΑΉœύΜΞΉς”ΟΒΡΫαΚœΈΜΒψ ,Μρ «–Έ≥…ΧΊ βΩ’ΦδΒΡΓΑΩΎ¥ϋΓ±ΫαΙΙ”ρ ;Ϋ”Ή≈άϊ”ΟΑ±ΜυΥα÷ΟΜΜΫιΒΦ’β–©ΙΠΡήΧΊ“λ–‘ΈΜΒψΒΡΕ®œρΆΜ±δ ,Ψ≠ΫΆΡΗΥΪ‘”ΫΜœΒΆ≥’Ι ΨΆΜ±δ“λ÷÷ ,≤ΔΩ…Ά®ΙΐΉ‘…μΒΡ―τ–‘―Γ‘ώΦχ±π”–“β“εΒΡΫαΙΙΙΠΡή”ρ. ’β÷÷ΦΦ θΒΡΆΜΤΤ‘Ύ”Ύ AptaPrintTM Ω…“άΩΩΡΩΒΡΈΜΒψΜρ «ΝΎΫϋ≤ΩΈΜΒΡΗΏΖ÷±φ¬ ΫβΨω”…”ΎΑ±ΜυΥα÷ΟΜΜΫιΒΦΒΡΫαΙΙ–ό Έ ,Εχ≤ζ…ζ”’ΒΦΉς”ΟΜρ «”ΑœλΝΥΒΑΑΉ÷ ΙΧ”–ΒΡ’έΒΰ. Νμ“ΜΗωΆΜΤΤΨΆ «άϊ”Ο»μΦΰΩ…“‘ΨΪ»ΖΒΊΙΙΫ®Ά§‘¥–‘Έο÷ ΒΡΫαΙΙΡΘ–Ά. κΡ ΧεΩ…“‘ΫαΚœ”Ύ≤ΜΆ§Έο÷ ΒΡΖ¥”Π±μΟφ ,ΧΊ“λ–‘ Ε±πΒΑΑΉΒΡ≤ΜΆ§ΙΠΡή”ρ.Υδ»ΜκΡ Χε «–ΓΖ÷Ή”ΒΑΑΉ ,ΒΪΥϋ≤Μ «“Μ÷÷ΤΫΟφΫαΙΙ ,Εχ «“‘“Μ÷÷ΕύΧ§–‘ΒΡΩ’ΦδΫαΙΙ¥φ‘ΎΒΡ (Αϋά®Ης÷÷’έΒΰΓΔ¬ί–ΐΓΔΩΎ¥ϋΒ») .

5 ’ΙΆϊ
”κΖ¥“εΚΥΥαΦΦ θœύ±» ,κΡ ΧεΦΦ θΜΙœ‘ΒΟœύΒ±Ρξ«α ,ΒΪΥϋΟ«’κΕ‘Α–Έο÷ ΒΡΕύ‘Σ…ζΈο―ßΜν–‘(Φ»Ω…“‘“÷÷Τ ,“≤Ω…“‘ΦΛΜν) “―“ΐΤπΝΥ―ß’ΏΒΡΙψΖΚΙΊΉΔ. άϊ”ΟκΡ ΧεΕ‘≤ΜΆ§±μ–ΆΒΡ…Η―Γ“‘ΦΑΕ‘≤ΜΆ§±μΟφΖ÷Ή”≤ΜΆ§ΙΠΡήΫαΙΙ”ρΒΡΦχ±π ,”–άϊ”ΎΆΊΩμΕ‘ΒΑΑΉ÷ Ήι―ßΒΡ―–ΨΩ ,Φ”Ωλ–¬“©―–ΖΔΒΡΫχ≥Χ.
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷Ψ
