
ժҪ ���Ի��ʽ�������ø-14(MMP-14)���ṹ��Ϊ�б꣬ͨ���ɾ������ʮ���Ŀ�ɸѡ�ͷ���ģ�⡢ϸ������ӫ�⡢���������Ͳ����Լ�����ϸ�����òⶨ�ȼ�����������˫���� MMP-14 �ͽ�������С���ӽ�϶��ĵ�ɸѡ���о�. �� 4 ��ɸѡ���ɾ���õ���Ч��������� 13 ����ͬ�Ķ�������. ���з�����ʾ�����ܵ�һ��������:AHQLH��HHXH��EI / L PLL / I . ����ģ����Խӽ�һ��ȷ��һ������ AHQLH��HHTH��LPLL �� MMP-14 ���ṹ��İ����� 120-125 �������÷��ӶԽӲ�����һ����רһ�ԣ����� MMP-14 ����IJ������� MMP-14��ͬʱ��Ͻ�������. ϸ������ѧ�о�ȷ�ϣ����ⶨ�Ľ�����ɾ���� MMP-14 �յ������ MG63 ϸ���������õĽ�����ã���ʾ����Ķ�MMP-14 �İ��������ԣ����Һϳɵ� AHQLH��LPLL һ�����ж��Ķ� MG63 ϸ����������һ������������. ��Щ�µĺ;���һ�� MMP-14 רһ�Ե�һ�����п������ڰ��� MMP-14 ������ҩ����з�������.
���ʽ�������ø( MMPs)��һ��ṹ��ص�Zn2+��Ca2+����������ø�����ڷֽ�ϸ����������( extracellular matrix��ECM ) �ĵ���ˮ��ø�������1��. �� 1962 ���״���Gross��Lapiere ���������Խ�ԭ����ø��ԭø������2]�����������ѷ��� 23 ����MMPs. �о������ʾ��MMPs �����������Ͳ�����������̥��������̬���ɡ���֯�����ؽ���Ѫ�ܷ�������֢��Ӧ���ر�����������Ϯ��ת���з�������Ҫ�����ã�3��4��. MMP-14(Ҳ��Ϊ MT1-MMP)��Ϊ�� 1�������Ĥ�ͻ��ʽ�������ø��ֱ�ӻ��ӽ���ϸ��������еĶ�����֣�����Ϊ��MMPs��������������ϵ�����е�ø��5��.
���\���ķ��ٷ�չΪ��ͨ������ɸѡҩ�н�Ϸ����ṩ���µ�˼·��;�����ɾ���չʾ�������ǽ�������չѸ���ҹ㷺Ӧ�õ��ֶ�֮һ. ���ǽ�������Դ����(����) �Ļ����DNAƬ�����ɾ������ǵ����ں϶� չ ʾ �� �� �������. �������DNAƬ��Ϊ����Ѻ����ᣬ������ɾ�������Ŀ�. �ü���������ŵ��ǽ������������ֱ����ϵ��һ������������������������������ɸѡ���з��Ӹ�Ч�����. Ŀǰ�ѿ�չ���� MMPs ��ص����ɾ����Ŀ�ɸѡ�������ⷽ��������������о��� ����6 �� 10��. �� �� �� �ˣ��� �� MMP-14 �� �����Ŀ�ɸ ѡ �� �� �� ������ �� �� �� չ ˫ �� �� MMP-14�ͽ������ӵĽ���ķ���ɸѡ���о���δ������.
���о����˹��ϳɵ� MMP-14 ���ṹ��Ϊ�б꣬������Ʒ���ɾ������ʮ���Ŀ�������ɸѡ��ͨ������ɸѡ���������б������Խ�ϵĶ��ģ�ȷ��һ���� �У����������ͬԴ���бȶԡ�����ģ�⡢���������Ͳ���������ϸ���ⶨ�ȷ�����ȷ������Ķ� MMP-14 �ͽ������ӵ�˫�������Լ�ϸ�������������� �ã�Ϊ �� �� MMP-14 �� �� �� �� �� �� ҩ��ɸѡ���з��ṩǰ�ڹ�������.
1 �����뷽��
1. 1 ����
�ɾ������ ʮ �� �� ��(1. 5 �� 1013pfu /mL) �� ������ ER2738( F'lacIq ��( lacZ)M15 proA + B + /zzf �s�sTn10( TetR ) fhuA2 supE thi ��( lac-proAB) �� ( hsdMSmcrB)5 ( r k�� mk�� )McrBC �� )���� New England Biolabs(Cat. 8110S) ;�˳ɹ����� MG63 ϸ������������������ص�ʵ �� �� �� �� �� �� �� �� ��;Hitrap �� ֬ �� ��Pharmacia Biotech(Cat. 17-0575-01) ;Ni-NTA ��֬���� Qiagen( Superflow 30430) ;MMP-14 ���״��ṹ���Ա���������������Ƽ�����˾;.�������⣬�����Լ������ڳɶ��Լ���˾����ѧ�Լ���Ϊ����������.
1. 2 MMP-14 �е����ɾ������ʮ���Ŀ�ɸѡ
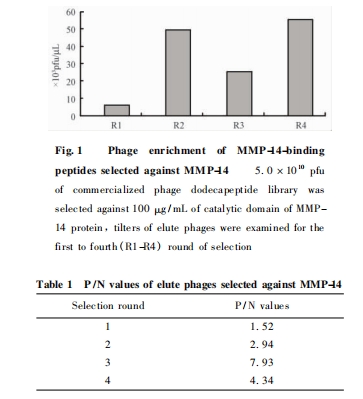
�� MMP-14 ���״��ṹ��(100 ��g /mL) Ϊ�б꣬BSA Ϊ���Զ��գ�ɸѡ���������ζȲⶨ���ɾ��嵥�� DNA( ssDNA) ���Ʊ��Ͳ���ȷ������� Ph.D. -12 TM�ɾ���չʾ�Ŀ��ֲ�. ÿ�� �� 150 ��L ��Ʒ���� 96 �� �壬�� ʪ �� �� ��4 �� �� �� �� �� �� ��ҹ;��ȥ����Һ��4 �� BSA ����� ҡ �� �� 1 h;�� ȥ�� �� Һ��TBS �� �� Һ ( 50 mmol /L Tris-HCl��150mmol /L NaCl) �� �� ϴ �� 6 ��;ÿ �� �� �� 5. 0 �� 1010pfu �ɾ��壬��������� 30 �� 60 min;��ȥδ����ɾ��壬0. 1% TBST( TBS + 0. 1% Tween-20)��ϴ 5 �Σ�200 ��L �ʰ���-HCl��pH2. 2 ϴ�������� ����ɾ��壬�ظ� 2 ��. �ռ�ϴ��Һ��Tris-HCl( pH9. 1)�к�. ���� 1 /5 ��Ʒ���������ڵζȲⶨ��ϴ������������������������ɸѡ�������� 4 ��ɸѡ. ���� P /N ֵ( P:�е�����ɾ���ϴ����ζȣ�N:BSA ���յ�����ɾ���ϴ����ζ�) .
1. 3 �ɾ���DNA����
��ѡ�ɾ����Ŀ�ɸѡ�ĵ� 3 �� 4 ���ɾ��嵥��¡���Ʊ� ssDNA���ɱ�������Զ־ �� ˾ �� ��˫ �� ��ABI PRISM 3730 XL DNA �Զ������Dzⶨ. ��������Ϊ-96 gIII:5��-HOCCCTCATAGTTAGCGTAACG-3��.������ 17 ���ɾ��嵥��¡.
1. 4 ����¡�ɾ����������ɸ�ⶨ
ɸѡ��õĺ���������е���¡�ɾ��������������� 1. 2 ������������ MMP-14 �� �� �� ��ɸ�������� P /N ֵ�����ڲⶨ������ɾ���� MMP-14 ��������P/N ֵԽ�߱�ʾ����Խǿ.
1. 5 ��������з�����һ�����л��
���� Blastp��Clustal X 1. 81 ��������Ϣ�������� MMP-14 ����� �� �� �� �� �� �� �� �� �� �� �� �����Ի�ÿ��ܵ�һ������.
1. 6 һ�������� MMPs �ķ��ӶԽ�
Ϊ���ڷ���ˮƽԤ��һ�����ж� MMP-14 �İ����ԣ��� �� Molegro Virtual Docker ( MVD) �� SwissPDB Viewer �� �� �� �� �� һ �� �� �� �� MMP-14 ��MMPs �����Ա�ķ���ģ����Խӷ���. MVD �����µ�һ����ƽ̨�Խ����������ṩ�˶Խ�����Ҫ�����й��ܣ����ҶԽӽ����Ϊȷ����������ʹ��.�أ�����ͨ����ѯ PDB ���ݿ��� MMP-14 ����ά �� �� 1bqqM��һ �� �� �� �� �� �� �� ģ �� �� ��Swiss-PDB Viewer �� ����Ȼ �� �� �� �� �� MMP-14 ��һ��������ά�ṹ���� �� �� �� MVD �� �� �� �� �У����������Ԥ�⡢�Խӹ������� �� �� �裬�� �� Լ 10�ֵ����ҵ������� �� �� �� �� ģ ʽ �� �� �� �� �� �� ���ܡ����������ðе��Լ���� ���õ��������.ͬ���������� �� һ �� �� �� �� MMPs �� �� �� �� �� Ա�ķ���ģ��Խ�. ͨ �� �� ѯ PDB �� �� �� �� �� �� 13�� MMPs( MMP1 �� 3��MMP7 �� 14��MMP16��MMP 20�� MMP 23 ) �� �� ֪ �� ά �� ������ Ӧ ģ �� �� �� Ϊ1SU3A��1RTG��1C3IB��2DDY��3DPE��1L6J��1Q3A��1HV5��2W08��1EUBA��1bqqM��1RM8A��2JSD ��2K72.
1. 7 ����¡������ɾ���Խ������ӵ��Ͳⶨ
Ni2+NTA��֬ȥNi2 + ��Hitrap��֬�� Zn2+�����������Ͳⶨ�ȴ����������ף�9�ݽ���.
1. 7. 1 NTA��֬ȥNi2+���� ��ȡ�������֬����װ��2֧���Ĺ��У�TBS ϴ�ӣ�����1֧���Ĺ��м��� 0. 5 mol /L EDTA ( pH8. 0 ) ���� �� 1 �Σ��� ��0. 1%TBSTϴ3 �Σ�������TBS�����ȡ��� װ Ϊ100 ��L /�ܣ�4 �� ����. EDTA ����δ��������֬�ֱ�����Ϊ M �� �� M + ��M ��ʾ����Ԫ��.
1. 7. 2 Hitrap ��֬��Zn2+���� ������NTA ��֬�������ƣ���ͬ���� Hitrap ��֬δ���ؽ�������. ��ȡ����� Hitrap ��֬�������㹻�� Zn2 + (0. 3 mol /LZnSO4) �����ģ�����TBS��0. 1%TBSTϴ�ӣ���װ��4 �� ����.
1. 7. 3 ����¡������ɾ����Zn2+�Ͳⶨ �ֱ��Լ��غ�δ���� Zn2 + �� Hitrap ��֬Ϊ�������Դ�����ȡ M + ��M �� ��֬Ep �ܸ�1֧������ BSA �������1h�������壬���� 5 �� 1010pfu �����ĵ���¡�ɾ��������� 1 h�������壬0. 1% TBST ϴ�ӣ�0. 2 mol /L �ʰ���-HCl ϴ�� 2 �Σ��ϲ� 2 ��ϴ��Һ���ⶨ�ɾ���ζȣ����� P /N ֵ( ��������������֬�ɾ���ϴ�ѵζ� /δ���ؽ�������������֬���ɾ���ϴ�ѵζ�) . Ni2 + �� ֬�������ⶨ������ͬ Zn2 + ��֬�ⶨ.
1. 7. 4 ����¡������ɾ����Ni2+�Ͳⶨ ����ͬ1. 7. 3���Լ��غ�δ����Ni2+��NTA��֬Ϊ�������Դ���.
1. 8 ����¡������ɾ�����ϸ����ϵ�����ӫ����
MG-63ϸ�����ա�������������� ( �� 10%FBS �� DMEM��2 mmol /L�Ȱ�������100 U /mL�� ù�غ� 100 ��g /mL��ù����37 �� ��5% CO2 �� FORMACO2 ������������) ����6�װ����Ʊ� MMP-14 �յ������δ�յ������MG-63ϸ����Ȼ����������ĵ���¡�ɾ��壬��������ϴ�ӣ�ϸ���̶��Ȳ��������M13����( Amersham Biosciences ��˾) ������ϡ�ϴ�ӵȲ��裬����FITC���������� IgG�����ͷ�Ƭ��Olympusӫ�������¹۲��ɾ�����ϸ��������ӵĽ��ˮƽ��������ӫ����.

1. 9 �ϳɶ��Ķ�MG-63 ϸ������������MTT�ⶨ
MG-63 ϸ �� �� �ա������������������ �� 5 ��104 �� /mL ϸ����Һ���� 96 �� �� ��ÿ�� 100 ��L��5% CO2 ������������ 24 h ���ں����μ��벻ͬŨ�ȵĺϳɶ���(0. 1��1��10 �� 100 ��g /mL)��������72h ��ÿ���� 20 ��L MTT(5 mg /mL) ����������4h���������壬�ټ� �� DMSO 150 ��L����10min��ʹ�����ܽ⣬ø���Dzⶨ 490 nm ������������ȣ�����ϸ������������.
������ = (1-��ҩ���ƽ��Aֵ /�������ƽ�� A ֵ) ��100% ���ظ�3��.
2 ���
2. 1 ����MMP-14���ɾ�������Ŀ�ɸѡ
��MMP-14���״��ṹ��Ϊ�б꣬����4���ɾ������12�Ŀ�ɸѡ���ɾ���õ�������ϴ���ɾ���ĵζȽ���� Fig. 1���ɼ��ɾ�������ɸѡ���������Ӷ��õ����������ɾ���ζȴӵ�1�ֵ�3. 8��103pfu /��L���� ��4�ֵ�5. 3 �� 104pfu /��L;ͬʱ��1��3���У���P /N ֵҲ����ɸѡ���������Ӷ���ߣ��� Table 1.
2. 2 ���� MMP-14 ��������е�ȷ����������ɸ�ⶨ
�ֱ���ȡɸѡ��3��4�ָ�14��3���� 17 ���ɾ��嵥��¡����������ȡ�� ��¡�ɾ���ssDNA��1% ��֬��������Ӿȷ�ϣ����������õ� 13 ����ͬDNA���У�ȷ�������������н���� Table 2.���� C3-5��C3-9 Ϊ�ظ�����. ��BSAΪ���գ��ⶨ��������ĵ���¡�ɾ���� MMP-14 ���״��ṹ�������. �����ʾ����������ɾ���� MMP-14��������һ�������������п�¡C3-52 ������ߣ����ΪC3-62�� C3-9(C3-5) ��C3-52 ����2��Ala��C3-62 ����4��Leu����Ala��Leu�� �� MMP-14�����һ����Ҫ��ɳɷ֣����Ƿ�˵���� 2 ���������о��� MMP-14 �� �� �� �����һ�����о�֤��.���⣬�ж������У������ǵ� 4 �ֵ������к��нϸ߱�����His�����Ǽ�����ʵ���ҵ��о���ȷ�ϸߺ�����His ���� Zn2+�Ƚ������Ӿ���ǿ��������11�ݣ��� Zn2 + ��ǡ�� MMPs ���ṹ����Ҫ�Ľ������ӣ����� MMPs ��Ĺؼ�.
2. 3 MMP-14 ��������е�ͬԴ�ȶԼ�һ������ȷ��
��ɸѡ�õ��� MMP-14 ����������� NCBI ���� GenBank Blast ������δ�����������������ͬԴ����. ���ж������о������������Clustal X1. 81�������бȶԣ�������ִ��ڿ��ܵ�һ������AHQLH��HHXH��L / IE PLL / I(X �������ⰱ����) ����Fig. 2��ר�������:200910161829X��12��.

2. 4 һ�����ж����� MMP-14 �ķ���ģ��Խ�
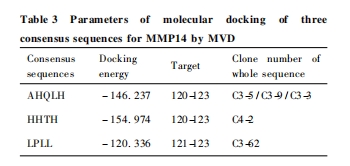
ΪԤ���ȷ��ɸѡ�õ���һ�����ж�MMP-14�İ����ԣ�ѡȡһ������ �� ������3������AHQLH��HHTH��LPLL �� �� ���� MMP-14 �� �� �� ����ģ��Խ�. MMP-14 ���ṹ��1bqqM(����������λ��:114��227) ����Ѫ���� �� �� �� 3c7xA( �� ��������λ��:316 �� 511 ) �Ľ���Ϊȷ���зֶԽӶ��Ĵ����˼���Ŀɿ���. �ֱ��� MMP-14 �� ���ṹ�����Ѫ���ؽṹ��ķ��ӶԽӽ����ʾ��3��һ�����о��ܺܺõضԽ��� MMP-14 ���ṹ��� aa. 120��125 �� ������ĶԽ�λ��ΪHis-Asn-Glu�������� MMP-14 �� Zn2 + �������;����Ԥ����ͬ������һ���� �� �� MMP-14 ����Ѫ���ؽṹ������ �� �� �� �� �� ��. �� �� �� �� �� AHQLH��HHTH��LPLL �� MMP-14 �ķ��ӶԽ��������仯�� Fig. 3(�� 347 ҳ��ҳ)�� Table 3.
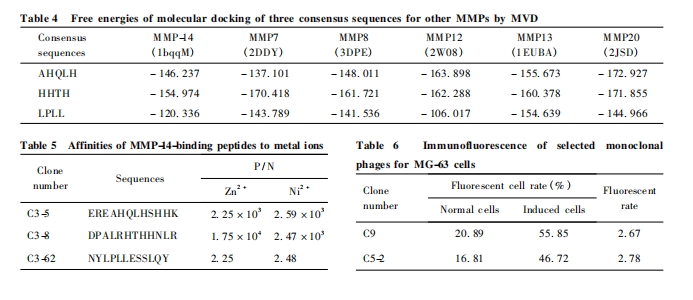
Ϊ��һ��Ԥ���ȷ��һ�����ж�MMP-14��רһ�ԣ��������Ƶķ����� AHQLH��HHTH��LPLL �� 3��һ�����ж� PDB ���ݿ����ѽ�����ά�ṹ�� 14�� MMPs ( MMP1-3��MMP7-14��MMP16��MMP 20��MMP 23)�����˷��ӶԽ�. �������:һ�����п���MMP7-8��MMP12-13 �� MMP20 ��5��MMPs �� ���Ա�Ĵ��ṹ��Խӣ���������8�� MMPs �����Ա��Խ���Ч. ���������5��MMPs �Ļ����ṹ�й�:MMP7 �� �� ������Ľṹ������4��MMPs ���ű��Ľṹ���13��14��. 3 ��һ�����жԽӺ������仯�ȼ� Fig. 4 �� Table 4.

2. 5 ����¡�ɾ���Խ������ӵ�������
��Ȼ���ӶԽӽ����ʾ3 ��һ�������ܹ���MMP-14 �� Zn2 + �������ܺöԽӣ�����Ԥ��õ���¡�ɾ�����������ĶԽ������ӿ��ܾ���һ��������. ��3�� һ �� �� �е���¡�ɾ����������Zn2 + ��Ni2 + �Ľ����Ͳ��������ⶨ����� Table5. �� �� �� �� Ԥ �� �ģ��� �� �� �� �� �� �� �� Zn2 + ��Ni2 + ����һ������������¡ C3-5��C3-8�����źܸߵ����������һ�����ж� MMP-14 �ĶԽӽ��������Ϊ����ȷ����һ�����о��� MMP-14 �� �� ��Zn2 + �Ƚ�����˫����������.

2. 6 ����¡������ɾ���� MMP-14 ��ϸ��ˮƽ�İ�����
SABC-FITC ����ӫ���ⵥ��¡������ɾ���C3-5��C3-52 �� MMP-14 �յ��ͷ��յ������MG-63ϸ���Ľ��ˮƽ������� Fig. 5. �ɼ��������ȣ�2���ɾ�����MMP-14�յ�����MG-63 ϸ �� �� ӫ ��ǿ�����Ը��ڶ���ϸ�������յ�ϸ����ӫ�������Ը���δ�յ�ϸ������ Table 6.

2. 7 һ�����ж��Ķ� MG-63 ϸ����������������
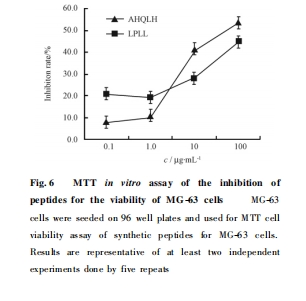
����MTT������� �� �� �� һ �����ж��� ��MG-63 ϸ���������������ã����ϳɺͲⶨ��2 ���������� AHQLH �� LPLL������� Fig. 6. �ɼ����źϳɶ���Ũ�ȵ����ӣ����ϸ����������������ǿ�����ֳ����Եļ���������ϵ. ���� AHQLH ��Ũ��Ϊ 10 ��g /mL �� ��ϸ���������ʼ� �ɴﵽ41. 6% .
3 ����
�� 20 ���������� MMPs �Ķ��ֿ��������Ƽ��Ⱥ�����ٴ����飬�������ҩ�︱���ô� /ѡ���Բ�����ٴ�����Ģ��ڱ����. ���е�һ����Ҫԭ������Ϊ��Щ���Ƽ����й��������ԣ����������˰�ø��Ҳ������һЩ����ø��15��. ����Ϊֹ��ֻ�� 1������ MMPs �� Doxycycline ͨ�� FDA ��֤.
���� MMP-14 ����Ϊ��������������ϵ�����е�һ�� MMPs������ѡ�� MMP-14 Ϊ�б�����ɾ�������Ŀ��ɸѡ���о�������ͨ���ɾ������ʮ���Ŀ�ɸѡ�ͷ���ģ�⡢ϸ�� in vitro ���о�ȷ����Ч��רһ�Ե� MMP-14 ���Ƽ����Ӷ����ڰ��� MMP-14�ȵ�ҩ����з�. �� MMP-14 ��������Ϊ�б���ɾ�������Ŀ�ɸѡ�����ǵõ����ɾ���ĸ����������� P /N ֵ�ϵ�( < 8) ���ҵ�4�ֵ��ڵ�3�֣����ڵ�5�� P /N ֵҲδ�����(����δ����) ��Ϊ������ѡȡ�˵�3��4 �ֵĸ����ɾ�����в����ǵ�4 �ֵĶ����Ѻ����His��������ǰ�ڵ��о��Ѿ�����His �Ĵ��ڶ� �� �� Zn2 + ���ڵĶ��ֽ�������ǿ��������11��16�ݣ���Ȼ����� His ������������������Ϊ MMPs ��רһ�����Ƽ�. Ϊ�ˣ����ǶԵ� 3 �ָ������ɾ�������˴����IJ���. ������������������˶�����һ�����еĶ��ģ������������� Ala��Leu �Ȱ����� �� MMP-14 ���ﵰ�������ԣ�Ҳ ��ʾ��ɸѡ����Ŀɿ��ԣ������������˷ܵ��ǣ�ͨ��������ɸ���õ��˶�����P/N ֵ �Ľ���ģ�����ߵ����ɾ��������ﵽ 4024. 39��Զ���ڸ��ֵĸ�������( P /N = 7. 93)
Ϊ�˽�һ��Ԥ���ȷ�����õ���һ��������MMP-14 �Ƿ����������Լ�ȷ���� MMP-14 ������õĽṹ�����Dz��ü��������ģ����ԽӺ�ϸ������ӫ�����ַ�����������ط���. �������ṹ����Ѫ���ؽṹ�����ڵ� MMP-14 ���Ķ�������Լ����� MMPs �����Ա��ά�ṹ����ȫ������Ϊ���ǽ��о�ȷ�ķ��ӶԽ��ṩ�˾�ķ��������. ���Ǵӳ���ȷ����һ��������ѡȡ��3 ������(AHQLH��HHTH��LPLL) ���������� MMP-14 �ļ��������ģ����Խӣ��Խӽ��һ�����һ��ȷ���� 3 ��һ��������MMP-14��Ч��ϣ�����ȷ��������ӶԽ�λ����MMP-14 �� �� �� �� �� ��His-Asn-Glu λ�㣬��HX�ԽӵIJ��� ��������һ�Ƕ�֤����ɸѡ��õ�һ�����ж�����MMP-14 ���ṹ���רһ �Խ���� �ã�ͬ��Ҳ֤����������MMP-14�Ĵ��ṹ��Ϊ�б�����Ŀ�ɸѡ��ȷ��. ����������ǣ���3��һ�������� 14 ���ѽ�����ά�ṹ�� MMPs �е� 5 ��MMPs Ҳ�ܹ���Ч���ӶԽӣ���ʾ�������һ�����Ż��� 3 ��һ��������������MMP-14 ��רһ��. ��һ������ϸ��ˮƽ������ӫ����Ҳȷ�Ͻ�����ɾ�����MMP-14 ����Ч���. MG-63 ϸ����һ����Ҫ�������ڴƼ��ص��� ���¿��Թ�������MMP-14 ( ʵ �� �� �� δ����) ��ͨ���յ��ͷ��յ����� MG-63ϸ��������ӫ���⣬������ϸ��ˮƽȷ���˵���¡������ɾ�����ͨ���������ϸ�������յ������ MMP-14 �Ľ�϶��������õ�. ���⣬���ӶԽӽ��ͬʱ����ʾ������ɸѡ��ò��ֶ��Ŀ��ܻ���������Ӿ��а��������ã�Ϊ�ˣ����ǽ��еĽ������������ⶨȷ���˸������ã�ȷ�ϲ���һ�����о��� MMP-14 �� ��������˫���������. ���Ƿdz���Ҫ�ķ��֣���Ϊ�����з��İ��� MMPs �����Ƽ���� �� �����Ʒ��ӽṹ�Ļ�����ż�������������ϼ����γɣ�1�ݣ�һ�����еĽ����������������Ա�������������ϼ���ʹ�ö������ڽ��Ͷ������÷��淢����Ҫ����.
�����Ϲ����Ļ����ϣ����Ǿ�ɸѡһ�����н����˳�����ϸ������ �� �� �ⶨ�����Ƕ��His �� �ĶԽ������ӹ��������ԣ���Ȼ������Ϊ�����ȵ�����ҩ��. Ϊ�ˣ����Ǻϳ��� AHQLH��LPLL�������ģ�����������ϸ�������о�֤�����ⶨ���������Ķ� MG-63 ϸ������һ����ϸ�������������ã�����������Ϊ�ȵ�ҩ��Ũ���Դ��ڽϴ��࣬��һ���Ķ����Ż���������ڽ�չ��.
4 ��Ҫ����
���о���MMP-14 ���״��ṹ��Ϊ�б꣬ͨ���ɾ������ʮ���Ŀ�ɸѡ�ͷ���ģ�⡢ϸ������ӫ�⡢���������Ͳ����Լ�����ϸ�����òⶨ�ȼ���������˫���� MMP-14 С���ӽ�϶��ĵ�ɸѡ���о�. ����ȷ����˫����MMP-14 �� Zn2 + �Ƚ������ӵ�AHQLH��HHXH��EI / L PLL / I �ȶ���һ �����У�ȷ����AHQLH��HHTH��LPLL �� 3 ��һ��������MMP-14 ����Ч�ԽӼ����ṹ����λ���Լ�һ���� MMP-14רһ ��. ����ϸ���ⶨ����ȷ��AHQLH��LPLL �� MG63 ����һ����ϸ��������������. ������Ϊ���뿪չMMP-14 ���Ƽ�����������õ��о����Լ�˫ ���� MMP-14�ͽ������ӵĿ�����С���Ӷ������Ƽ����з��ṩ����Ҫ��ǰ�ڹ�������.
��������������Ϊ��ҵ����ѧϰ����Ȩ��ԭ����ԭ��־���У�������Ȩ������ϵɾ�������±�ע���������³����������Ķ�ԭ�ļ��ο����ף����Ķ�ԭ��־
