
’Σ“Σ ΘΚ“‘Μυ÷ Ϋπ τΒΑΑΉΟΗ-14(MMP-14)¥ΏΜ·ΫαΙΙ”ρΈΣΑ–±ξΘ§Ά®Ιΐ …ΨζΧεΥφΜζ °ΕΰκΡΩβ…Η―ΓΚΆΖ÷Ή”ΡΘΡβΓΔœΗΑϊΟβ“Ώ”ΪΙβΓΔΫπ τάκΉ”«ΉΚΆ≤ψΈω“‘ΦΑΧεΆβœΗΑϊΉς”Ο≤βΕ®Β»ΦΦ θΘ§Ϋχ––ΝΥΥΪΑ–œρ MMP-14 ΚΆΫπ τάκΉ”–ΓΖ÷Ή”ΫαΚœΕύκΡΒΡ…Η―Γ”κ―–ΨΩ. Ψ≠ 4 ¬÷…Η―ΓΘ§ …ΨζΧεΒΟΒΫ”––ßΗΜΦ·≤ΔΜώΒΟ 13 Χθ≤ΜΆ§ΒΡΕύκΡ–ρΝ–. –ρΝ–Ζ÷Έωœ‘ ΨΘ§Ω…ΡήΒΡ“Μ÷¬–ρΝ–”–:AHQLHΓΔHHXHΓΔEI / L PLL / I . Ζ÷Ή”ΡΘΡβ”κΕ‘Ϋ”Ϋχ“Μ≤Ϋ»Ζ»œ“Μ÷¬–ρΝ– AHQLHΓΔHHTHΓΔLPLL ”κ MMP-14 ¥ΏΜ·ΫαΙΙ”ρΒΡΑ±ΜυΥα 120-125 «χ”ρΝΦΚΟΖ÷Ή”Ε‘Ϋ”≤ΔΨΏ”–“ΜΕ®ΒΡΉ®“Μ–‘Θ§ΕύΧθ MMP-14 ΫαΚœκΡ≤ΜΫωΑ–œρ MMP-14Θ§Ά§ ±ΫαΚœΫπ τάκΉ”. œΗΑϊ…ζΈο―ß―–ΨΩ»Ζ»œΘ§Υυ≤βΕ®ΒΡΫαΚœκΡ …ΨζΧεΕ‘ MMP-14 ”’ΒΦ±μ¥οΒΡ MG63 œΗΑϊΨΏ”–ΝΦΚΟΒΡΫαΚœΉς”ΟΘ§Ϋ“ ΨΫαΚœκΡΕ‘MMP-14 ΒΡΑ–œρΫαΚœΧΊ–‘Θ§≤Δ«“Κœ≥…ΒΡ AHQLHΓΔLPLL “Μ÷¬–ρΝ–ΕύκΡΕ‘ MG63 œΗΑϊΜνΝΠΨΏ”–“ΜΕ®ΒΡ“÷÷ΤΡήΝΠ. ’β–©–¬ΒΡΚΆΨΏ”–“ΜΕ® MMP-14 Ή®“Μ–‘ΒΡ“Μ÷¬–ρΝ–Ω…Άϊ”Ο”ΎΑ–œρ MMP-14 ΩΙ÷ΉΝω“©ΈοΒΡ―–ΖΔΚΆάϊ”Ο.
Μυ÷ Ϋπ τΒΑΑΉΟΗ( MMPs) «“ΜάύΫαΙΙœύΙΊΒΡZn2+ΚΆCa2+“άάΒΒΡΡΎκΡΟΗΘ§ τ”ΎΖ÷ΫβœΗΑϊΆβΜυ÷ ΉιΖ÷( extracellular matrixΘ§ECM ) ΒΡΒΑΑΉΥ°ΫβΟΗ≥§Φ“ΉεΘέ1Θί. Ή‘ 1962 Ρξ Ή¥Έ”…GrossΚΆLapiere ±®ΒάΧΊ“λ–‘ΫΚ‘≠ΫΒΫβΟΗΫΚ‘≠ΟΗ“‘ά¥Θέ2]Θ§÷ΝΫώ÷Ν…Ό“―ΖΔœ÷ 23 ÷÷»ΥMMPs. ―–ΨΩΫαΙϊœ‘ ΨΘ§MMPs ‘Ύ–μΕύ…ζάμΚΆ≤ΓάμΙΐ≥Χ»γ≈ΏΧΞΖΔ”ΐΓΔ–ΈΧ§Ϋ®≥…ΓΔΉι÷·–όΗ¥ΚΆ÷ΊΫ®ΓΔ―ΣΙήΖΔ…ζΓΔ―Ή÷ΔΖ¥”ΠΘ§ΧΊ±π «‘Ύ÷ΉΝω«÷œ°”κΉΣ“Τ÷–ΖΔΜ”Ή≈÷Ί“ΣΒΡΉς”ΟΘέ3Θ§4Θί. MMP-14(“≤≥ΤΈΣ MT1-MMP)ΉςΈΣΒΎ 1ΗωΦχ±πΒΡΡΛ–ΆΜυ÷ Ϋπ τΒΑΑΉΟΗΘ§÷±Ϋ”ΜρΦδΫ”ΫΒΫβœΗΑϊΆβΜυ÷ ÷–ΒΡΕύ÷÷ΉιΖ÷Θ§±Μ»œΈΣ «MMPsΦ“Ήε÷–”κ÷ΉΝωΙΊœΒΉνΟή«–ΒΡΟΗΘέ5Θί.
…ζΈοΦΦ θΒΡΖ…ΥΌΖΔ’ΙΈΣΗΏΆ®ΝΩΩλΥΌ…Η―Γ“©Α–ΫαΚœΖ÷Ή”ΧαΙ©ΝΥ–¬ΒΡΥΦ¬ΖΚΆΆΨΨΕΘ§ …ΨζΧε’Ι ΨΦΦ θΨΆ «ΫϋΡξά¥ΖΔ’Ι―ΗΥΌ«“ΙψΖΚ”Π”ΟΒΡ ÷ΕΈ÷°“Μ. Υϋ «ΫΪ±ύ¬κΆβ‘¥ΕύκΡ(ΒΑΑΉ) ΒΡΜυ“ρΜρDNAΤ§ΕΈ”κ …ΨζΧεΒΡΆβΩ«ΒΑΑΉ»ΎΚœΕχ ’Ι Ψ ‘Ύ … ΨζΧε±μΟφ. »τ≤ε»κΒΡDNAΤ§ΕΈΈΣΥφΜζΙ―ΚΥή’ΥαΘ§‘ρΙΙΫ®ΜώΒΟ …ΨζΧεΥφΜζκΡΩβ. ΗΟΦΦ θΒΡΉν¥σ”≈Βψ «ΫΪ±μ–Ά”κΜυ“ρ–Ά÷±Ϋ”ΝΣœΒ‘Ύ“ΜΤπΘ§άϊ”Ο≈δΧε”κΑ– ήΧεΒΡΧΊ“λ–‘«ΉΚΆΝΠΩ……Η―Γ≥ωΑ–Ζ÷Ή”ΗΏ–ßΫαΚœκΡ. ΡΩ«Α“―ΩΣ’ΙΕύ÷÷ MMPs œύΙΊΒΑΑΉΒΡ …ΨζΧεΈΡΩβ…Η―ΓΙΛΉςΘ§’βΖΫΟφΙζΡΎΆβΨυ”–œύΙΊ―–ΨΩ±® ΒάΘέ6 ΓΪ 10Θί. ΨΓ Ιή »γ ¥ΥΘ§Α– œρ MMP-14 ΒΡ …ζΈοΈΡΩβ…Η ―Γ …Ό ”– ―– ΨΩΘ§Εχ …ν »κ ΩΣ ’Ι ΥΪ Α– œρ MMP-14ΚΆΫπ τάκΉ”ΒΡΫαΚœκΡΖ÷Ή”…Η―Γ”κ―–ΨΩ‘ρΈ¥Φϊ±®Βά.
±Ψ―–ΨΩ“‘»ΥΙΛΚœ≥…ΒΡ MMP-14 ¥ΏΜ·ΫαΙΙ”ρΈΣΑ–±ξΘ§άϊ”Ο…ΧΤΖΜ· …ΨζΧεΥφΜζ °ΕΰκΡΩβΕ‘ΤδΫχ––…Η―ΓΘ§Ά®ΙΐΕύ¬÷…Η―ΓΗΜΦ·ΜώΒΟ”κΑ–±ξΧΊ“λ–‘ΫαΚœΒΡΕύκΡΘ§»ΖΕ®“Μ÷¬–ρ Ν–Θ§≤ΔΕ‘ΤδΫχ––Ά§‘¥–ρΝ–±»Ε‘ΓΔΖ÷Ή”ΡΘΡβΓΔΫπ τάκΉ”«ΉΚΆ≤ψΈωΓΔΧεΆβœΗΑϊ≤βΕ®Β»Ζ÷ΈωΘ§»ΖΕ®ΫαΚœκΡΕ‘ MMP-14 ΚΆΫπ τάκΉ”ΒΡΥΪΑ–œρ–‘“‘ΦΑœΗΑϊΜνΝΠΒΡ“÷÷ΤΉς ”ΟΘ§ΈΣ Μυ ”Ύ MMP-14 ΒΡ ΩΙ ÷Ή Νω œ» ΒΦ “©Έο…Η―ΓΚΆ―–ΖΔΧαΙ©«ΑΤΎΙΛΉςΜυ¥Γ.
1 ≤ΡΝœ”κΖΫΖ®
1. 1 ≤ΡΝœ
…ΨζΧεΥφΜζ ° Εΰ κΡ Ωβ(1. 5 ΓΝ 1013pfu /mL) ΚΆ Υό÷ςΨζ ER2738( F'lacIq ΠΛ( lacZ)M15 proA + B + /zzf ©s©sTn10( TetR ) fhuA2 supE thi ΠΛ( lac-proAB) ΠΛ ( hsdMSmcrB)5 ( r kΘ≠ mkΘ≠ )McrBC Θ≠ )ΙΚΉ‘ New England Biolabs(Cat. 8110S) ;»Υ≥…Ι«»βΝω MG63 œΗΑϊΘ§ΫΧ”ΐ≤Ω…ζΈο≤ΡΝœ÷ΊΒψ Β ―ι “ «ϋ ς –¬ ΫΧ Ύ Μί ‘υ;Hitrap ς ÷§ ΙΚ Ή‘Pharmacia Biotech(Cat. 17-0575-01) ;Ni-NTA ς÷§ΙΚΉ‘ Qiagen( Superflow 30430) ;MMP-14 ΒΑΑΉ¥ΏΜ·ΫαΙΙ”ρΙΚΉ‘±±Ψ©”―“ξ÷–ΝΣ…ζΈοΩΤΦΦ”–œόΙΪΥΨ;.≥ΐ±ξΟςΆβΘ§ΤδΥϋ ‘ΦΝΨυΙΚ”Ύ≥…ΕΦ ‘ΦΝΙΪΥΨΘ§Μ·―ß ‘ΦΝΨυΈΣΖ÷Έω¥Ω“‘…œ.
1. 2 MMP-14 Α–ΒΑΑΉΒΡ …ΨζΧεΥφΜζ °ΕΰκΡΩβ…Η―Γ
“‘ MMP-14 ΒΑΑΉ¥ΏΜ·ΫαΙΙ”ρ(100 ΠΧg /mL) ΈΣΑ–±ξΘ§BSA ΈΣ“θ–‘Ε‘’’Θ§…Η―ΓΓΔά©‘ωΓΔΒΈΕ»≤βΕ®ΓΔ …ΨζΧεΒΞΝ¥ DNA( ssDNA) ΒΡ÷Τ±ΗΚΆ≤β–ρΒ»ΖΫΖ®≤Έ’’ Ph.D. -12 TM …ΨζΧε’Ι ΨκΡΩβ ÷≤α. ΟΩΩΉΦ” »κ 150 ΠΧL ―υΤΖΑϋ±Μ 96 ΩΉ ΑεΘ§‘ω Σ »ί Τς ÷–4 Γφ «α ΈΔ ’π Β¥ Ζθ ”ΐ Ιΐ“Ι;ΒΙ»ΞΑϋ±Μ“ΚΘ§4 Γφ BSA Ζβ±’’ώ “Γ Ζθ ”ΐ 1 h;≥ΐ »ΞΖβ ±’ “ΚΘ§TBS ΜΚ ≥ε “Κ ( 50 mmol /L Tris-HClΘ§150mmol /L NaCl) Ωλ ΥΌ œ¥ ΩΉ 6 ¥Έ;ΟΩ ΩΉ Φ” »κ 5. 0 ΓΝ 1010pfu …ΨζΧεΘ§ “Έ¬«αΈΔ’πΒ¥ΫαΚœ 30 ΓΪ 60 min;Τζ»ΞΈ¥ΫαΚœ …ΨζΧεΘ§0. 1% TBST( TBS + 0. 1% Tween-20)«εœ¥ 5 ¥ΈΘ§200 ΠΧL Η Α±Υα-HClΘ§pH2. 2 œ¥Ά―ΧΊ“λ–‘ ΫαΚœ …ΨζΧεΘ§÷ΊΗ¥ 2 ¥Έ. ’Φ·œ¥Ά―“ΚΘ§Tris-HCl( pH9. 1)÷–ΚΆ. ±ΘΝτ 1 /5 ―υΤΖΘ§Τδ”ύ”Ο”ΎΒΈΕ»≤βΕ®ΚΆœ¥Ά―Έοά©‘ωΘ§ά©‘ωΈο”Ο”Ύœ¬¬÷…Η―ΓΘ§Ι≤Ϋχ–– 4 ¬÷…Η―Γ. ΦΤΥψ P /N ÷Β( P:Α–ΒΑΑΉΫαΚœ …ΨζΧεœ¥Ά―ΈοΒΈΕ»Θ§N:BSA Ε‘’’ΒΑΑΉΫαΚœ …ΨζΧεœ¥Ά―ΈοΒΈΕ») .
1. 3 …ΨζΧεDNA≤β–ρ
Χτ―Γ …ΨζΧεΈΡΩβ…Η―ΓΒΡΒΎ 3 ΚΆ 4 ¬÷ …ΨζΧεΒΞΩΥ¬ΓΘ§÷Τ±Η ssDNAΘ§”…±±Ψ©»ΐ≤©‘Ε÷Ψ ΙΪ ΥΨ ≤β –ρΘ§ΥΪ Ά― ―θABI PRISM 3730 XL DNA Ή‘Ε·≤β–ρ“«≤βΕ®. ≤β–ρ“ΐΈοΈΣ-96 gIII:5Γδ-HOCCCTCATAGTTAGCGTAACG-3Γδ.Ι≤≤β–ρ 17 Ηω …ΨζΧεΒΞΩΥ¬Γ.
1. 4 ΒΞΩΥ¬Γ …ΨζΧεΒΡ«ΉΚΆΝΠΖ¥…Η≤βΕ®
…Η―ΓΜώΒΟΒΡΚ§ΫαΚœκΡ–ρΝ–ΒΞΩΥ¬Γ …ΨζΧεά©‘ωΚσΘ§Α¥’’άύΥΤ 1. 2 Υυ ωΖΫΖ®Ϋχ–– MMP-14 Α– ΒΑ ΑΉ Ζ¥…ΗΘ§ΦΤΥψΤδ P /N ÷ΒΘ§”Ο”Ύ≤βΕ®ΫαΚœκΡ …ΨζΧεΕ‘ MMP-14 ΒΡ«ΉΚΆΝΠΘ§P/N ÷Β‘ΫΗΏ±μ Ψ«ΉΚΆΝΠ‘Ϋ«Ω.
1. 5 ΫαΚœκΡ–ρΝ–Ζ÷Έω”κ“Μ÷¬–ρΝ–ΜώΒΟ
άϊ”Ο BlastpΓΔClustal X 1. 81 Β»…ζΈο–≈œΔ»μΦΰΫχ–– MMP-14 ΫαΚœκΡ –ρ Ν– ΒΡ ±» Ε‘ ΦΑ Εύ ÷Ί –ρ Ν– Ζ÷ ΈωΘ§“‘ΜώΒΟΩ…ΡήΒΡ“Μ÷¬–ρΝ–.
1. 6 “Μ÷¬–ρΝ–”κ MMPs ΒΡΖ÷Ή”Ε‘Ϋ”
ΈΣΝΥ‘ΎΖ÷Ή”Υ°ΤΫ‘Λ≤β“Μ÷¬–ρΝ–Ε‘ MMP-14 ΒΡΑ–œρ–‘Θ§άϊ ”Ο Molegro Virtual Docker ( MVD) ΚΆ SwissPDB Viewer »μ Φΰ Ϋχ –– ΝΥ “Μ ÷¬ –ρ Ν– ”κ MMP-14 ΦΑMMPs Φ“Ήε≥…‘±ΒΡΖ÷Ή”ΡΘΡβ”κΕ‘Ϋ”Ζ÷Έω. MVD «Ήν–¬ΒΡ“ΜΗωΩγΤΫΧ®Ε‘Ϋ”»μΦΰΘ§ΥϋΧαΙ©ΝΥΕ‘Ϋ”Υυ–η“ΣΒΡΥυ”–ΙΠΡήΘ§≤Δ«“Ε‘Ϋ”ΫαΙϊΫœΈΣΉΦ»ΖΘ§ΫγΟφ»ί“Ή Ι”Ο.ΦρΒΞΒΊΘ§ Ήœ»Ά®Ιΐ≤ι―· PDB ΐΨίΩβΜώΒΟ MMP-14 ΒΡ»ΐΈ§ Ϋα ΙΙ 1bqqMΘ§“Μ ÷¬ –ρ Ν– ΕΧ κΡ Ϋα ΙΙ ΡΘ Ρβ ≤… ”ΟSwiss-PDB Viewer »μ ΦΰΘ§»Μ Κσ ΫΪ Μώ ΒΟ ΒΡ MMP-14 ”κ“Μ÷¬–ρΝ–»ΐΈ§ΫαΙΙ“ά¥Έ ΒΦ »κ ΒΫ MVD Ε‘ Ϋ” »μ Φΰ ÷–Θ§Ψ≠ΫαΚœ«χ”ρ‘Λ≤βΓΔΕ‘Ϋ”Ιΐ≥Χ…η÷Ο Β» ≤Ϋ ÷ηΘ§Ψ≠ Ιΐ ‘Φ 10¬÷Βϋ¥ζ’“ΒΫΡήΝΩΉν ”≈ ΒΡ Ε‘ Ϋ” ΡΘ Ϋ ≤Δ ΒΟ ΒΫ Ε‘ Ϋ” Ή‘ ”…ΡήΓΔ«ΉΚΆΝΠΓΔΉς”ΟΑ–Βψ“‘ΦΑ«βΦϋ Ής”ΟΒ»œύΙΊ ΐΨί.Ά§―υΖΫΖ®Ϋχ–– ΝΥ “Μ ÷¬ –ρ Ν– ”κ MMPs Φ“ Ήε Τδ Υϋ ≥… ‘±ΒΡΖ÷Ή”ΡΘΡβΕ‘Ϋ”. Ά® Ιΐ ≤ι ―· PDB ΐ Ψί Ωβ Ι≤ Μώ ΒΟ 13÷÷ MMPs( MMP1 ΓΪ 3ΓΔMMP7 ΓΪ 14ΓΔMMP16ΓΔMMP 20ΚΆ MMP 23 ) ΒΡ “― ÷Σ »ΐ Έ§ Ϋα ΙΙΘ§Ε‘ ”Π ΡΘ Αε Ζ÷ ±π ΈΣ1SU3AΓΔ1RTGΓΔ1C3IBΓΔ2DDYΓΔ3DPEΓΔ1L6JΓΔ1Q3AΓΔ1HV5ΓΔ2W08ΓΔ1EUBAΓΔ1bqqMΓΔ1RM8AΓΔ2JSD ΚΆ2K72.
1. 7 ΒΞΩΥ¬ΓΫαΚœκΡ …ΨζΧεΕ‘Ϋπ τάκΉ”ΒΡ«ΉΚΆ≤βΕ®
Ni2+NTA ς÷§»ΞNi2 + ΓΔHitrap ς÷§Φ” Zn2+ΓΔΫπ τάκΉ”«ΉΚΆ≤βΕ®Β»¥Πάμ≤Έ’’ΈΡœΉΘέ9ΘίΫχ––.
1. 7. 1 NTA ς÷§»ΞNi2+¥Πάμ Έϋ»ΓΒ»ΧεΜΐ ς÷§Θ§Ζ÷ΉΑ”Ύ2÷ßάκ–ΡΙή÷–Θ§TBS œ¥Β”Θ§Τδ÷–1÷ßάκ–ΡΙή÷–Φ”»κ 0. 5 mol /L EDTA ( pH8. 0 ) Θ§÷Ί Η¥ 1 ¥ΈΘ§‘Ό ”Ο0. 1%TBSTœ¥3 ¥ΈΘ§ΉνΚσΦ”»κTBSΘ§Μλ‘»ΓΔΖ÷ ΉΑ ΈΣ100 ΠΧL /ΙήΘ§4 Γφ ±Θ¥φ. EDTA ¥ΠΚΆΈ¥¥ΠάμΒΡ ς÷§Ζ÷±πΟϋΟϊΈΣ M Θ≠ ΚΆ M + Θ§M ±μ ΨΫπ τ‘ΣΥΊ.
1. 7. 2 Hitrap ς÷§Φ”Zn2+¥Πάμ ΖΫΖ®”κNTA ς÷§¥ΠάμœύΥΤΘ§≤ΜΆ§ΒΡ « Hitrap ς÷§Έ¥Φ”‘ΊΫπ τάκΉ”. Έϋ»ΓΒ»ΧεΜΐ Hitrap ς÷§Θ§Φ”‘ΊΉψΙΜΝΩ Zn2 + (0. 3 mol /LZnSO4) Θ§άκ–ΡΘ§“ά¥ΈTBSΘ§0. 1%TBSTœ¥Β”Θ§Ζ÷ΉΑΚΆ4 Γφ ±Θ¥φ.
1. 7. 3 ΒΞΩΥ¬ΓΫαΚœκΡ …ΨζΧεΒΡZn2+«ΉΚΆ≤βΕ® Ζ÷±π“‘Φ”‘ΊΚΆΈ¥Φ”‘Ί Zn2 + ΒΡ Hitrap ς÷§ΈΣ―τΓΔ“θ–‘¥ΠάμΘ§»Γ M + ΓΔM Θ≠ ς÷§Ep ΙήΗς1÷ßΘ§Φ”»κ BSA Ζβ±’≈ύ―χ1hΘ§Τζ…œ«εΘ§Φ”»κ 5 ΓΝ 1010pfu ά©‘ωΒΡΒΞΩΥ¬Γ …ΨζΧεΫαΚœ≈ύ”ΐ 1 hΘ§Τζ…œ«εΘ§0. 1% TBST œ¥Β”Θ§0. 2 mol /L Η Α±Υα-HCl œ¥Ά― 2 ¥ΈΘ§Κœ≤Δ 2 ¥Έœ¥Ά―“ΚΘ§≤βΕ® …ΨζΧεΒΈΕ»Θ§ΦΤΥψ P /N ÷Β( Ϋπ τάκΉ”ρϋΚœ ς÷§ …ΨζΧεœ¥Ά―ΒΈΕ» /Έ¥Φ”‘ΊΫπ τάκΉ”ρϋΚœ ς÷§ΒΡ …ΨζΧεœ¥Ά―ΒΈΕ») . Ni2 + ς ÷§ΒΡ«ΉΚΆΝΠ≤βΕ®ΖΫΖ®άύΆ§ Zn2 + ς÷§≤βΕ®.
1. 7. 4 ΒΞΩΥ¬ΓΫαΚœκΡ …ΨζΧεΒΡNi2+«ΉΚΆ≤βΕ® ΖΫΖ®Ά§1. 7. 3Θ§“‘Φ”‘ΊΚΆΈ¥Φ”‘ΊNi2+ΒΡNTA ς÷§ΈΣ―τΓΔ“θ–‘¥Πάμ.
1. 8 ΒΞΩΥ¬ΓΫαΚœκΡ …ΨζΧε”κœΗΑϊΫαΚœΒΡΟβ“Ώ”ΪΙβΦλ≤β
MG-63œΗΑϊΗ¥Υ’ΓΔ≈ύ―χΑ¥≥ΘΙφ≤ΌΉς ( Κ§ 10%FBS ΒΡ DMEMΘ§2 mmol /LΙ»Α±θΘΑΖΘ§100 U /mL«ύ ΟΙΥΊΚΆ 100 ΠΧg /mLΝ¥ΟΙΥΊ”Ύ37 Γφ Θ§5% CO2 ΒΡ FORMACO2 ≈ύ―χœδ÷–≈ύ―χ) Θ§‘Ύ6ΩΉΑε÷–÷Τ±Η MMP-14 ”’ΒΦ±μ¥οΚΆΈ¥”’ΒΦ±μ¥οΒΡMG-63œΗΑϊΘ§»ΜΚσΦ”»κά©‘ωΒΡΒΞΩΥ¬Γ …ΨζΧεΘ§Ψ≠Ζθ”ΐΓΔœ¥Β”Θ§œΗΑϊΙΧΕ®Β»≤Ϋ÷ηΚσΘ§Φ”»κ σΩΙM13ΒΞΩΙ( Amersham Biosciences ΙΪΥΨ) Θ§Ψ≠ΫαΚœΓΔœ¥Β”Β»≤Ϋ÷ηΘ§Φ”»κFITC±ξΦ«―ρΩΙ σΕΰΩΙ IgGΘ§Η ”ΆΖβΤ§Θ§Olympus”ΪΙβœ‘ΈΔΨΒœ¬Ιέ≤λ …ΨζΧε”κœΗΑϊ±μΟφΖ÷Ή”ΒΡΫαΚœΥ°ΤΫΘ§≤ΔΦΤΥψ”ΪΙβ¬ .

1. 9 Κœ≥…ΕύκΡΕ‘MG-63 œΗΑϊΜνΝΠΒΡΧεΆβMTT≤βΕ®
MG-63 œΗ Αϊ Η¥ Υ’ΓΔ≈ύ―χΑ¥≥ΘΙφ≤ΌΉςΘ§≈δ ÷Τ 5 ΓΝ104 Ηω /mL œΗΑϊ–ϋ“ΚΘ§”Ύ 96 ΩΉ Αε ÷–ΟΩΩΉΦ” 100 ΠΧLΘ§5% CO2 ≈ύ―χœδ÷–≈ύ―χ 24 h Χυ±ΎΚσΘ§“ά¥ΈΦ”»κ≤ΜΆ§≈®Ε»ΒΡΚœ≥…ΕύκΡ(0. 1Θ§1Θ§10 ΚΆ 100 ΠΧg /mL)ΦΧ–χ≈ύ―χ72h ΚσΘ§ΟΩΩΉΦ”»κ 20 ΠΧL MTT(5 mg /mL) Θ§ΦΧ–χ≈ύ―χ4hΚσΈϋΨΓ…œ«εΘ§‘ΌΦ” »κ DMSO 150 ΠΧLΘ§’ώΒ¥10minΘ§ ΙΤδ≥δΖ÷»ήΫβΘ§ΟΗ±ξ“«≤βΕ® 490 nm ≤®≥Λ¥ΠΒΡΈϋΙβΕ»Θ§ΦΤΥψœΗΑϊ…ζ≥Λ“÷÷Τ¬ .
“÷÷Τ¬ = (1-Φ”“©ΉιΒΡΤΫΨυA÷Β /Ε‘’’ΉιΒΡΤΫΨυ A ÷Β) ΓΝ100% Θ§÷ΊΗ¥3¥Έ.
2 ΫαΙϊ
2. 1 Α–œρMMP-14ΒΡ …ΨζΧεΥφΜζκΡΩβ…Η―Γ
“‘MMP-14ΒΑΑΉ¥ΏΜ·ΫαΙΙ”ρΈΣΑ–±ξΘ§Ψ≠Ιΐ4¬÷ …ΨζΧεΥφΜζ12κΡΩβ…Η―ΓΘ§ …ΨζΧεΒΟΒΫΗΜΦ·Θ§œ¥Ά― …ΨζΧεΒΡΒΈΕ»ΫαΙϊΦϊ Fig. 1Θ§Ω…Φϊ …ΨζΧεΥφΉ≈…Η―Γ¬÷ ΐΒΡ‘ωΦ”Εχ÷πΫΞΒΟΒΫΗΜΦ·Θ§Τδ …ΨζΧεΒΈΕ»¥”ΒΎ1¬÷ΒΡ3. 8ΓΝ103pfu /ΠΧL…ΐ÷Ν ΒΎ4¬÷ΒΡ5. 3 ΓΝ 104pfu /ΠΧL;Ά§ ±‘Ύ1ΓΪ3¬÷÷–Θ§ΤδP /N ÷Β“≤ΥφΉ≈…Η―Γ¬÷ ΐΒΡ‘ωΦ”ΕχΧαΗΏΘ§Φϊ Table 1.
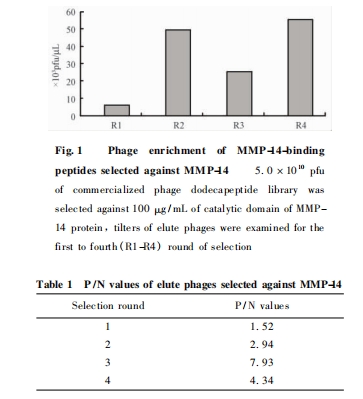
2. 2 Α–œρ MMP-14 ΫαΚœκΡ–ρΝ–ΒΡ»ΖΕ®ΦΑ«ΉΚΆΝΠΖ¥…Η≤βΕ®
Ζ÷±πΧτ»Γ…Η―ΓΒΎ3ΚΆ4¬÷Ης14”κ3ΗωΙ≤ 17 Ηω …ΨζΧεΒΞΩΥ¬ΓΘ§ά©‘ωΚσΧα»ΓΒΞ ΩΥ¬Γ …ΨζΧεssDNAΘ§1% «μ÷§Χ«ΡΐΫΚΒγ”Ψ»Ζ»œΘ§ΥΆ―υ≤β–ρΘ§Ι≤ΒΟΒΫ 13 Χθ≤ΜΆ§DNA–ρΝ–Θ§»ΖΕ®Τδ±ύ¬κΒΡκΡ–ρΝ–ΫαΙϊ»γ Table 2.Τδ÷– C3-5ΓΔC3-9 ΈΣ÷ΊΗ¥–ρΝ–. “‘BSAΈΣΕ‘’’Θ§≤βΕ®ΗςΚ§ΫαΚœκΡΒΞΩΥ¬Γ …ΨζΧεΕ‘ MMP-14 ΒΑΑΉ¥ΏΜ·ΫαΙΙ”ρΒΡ«ΉΚΆΝΠ. ΫαΙϊœ‘ ΨΘ§ΗςΫαΚœκΡ …ΨζΧεΕ‘ MMP-14ΒΑΑΉΨυΨΏ”–“ΜΕ®ΒΡ«ΉΚΆΝΠΘ§Τδ÷–ΩΥ¬ΓC3-52 «ΉΚΆΝΠΉνΗΏΘ§Τδ¥ΈΈΣC3-62ΚΆ C3-9(C3-5) Θ§C3-52 Κ§”–2ΗωAlaΘ§C3-62 Κ§”–4ΗωLeuΘ§ΕχAlaΓΔLeu’ΐ « MMP-14ΒΉΈοΒΡ“ΜΗω÷Ί“ΣΉι≥…≥…Ζ÷Θ§’β «ΖώΥΒΟς’β 2 ΧθΕύκΡ–ρΝ–ΨΏ”– MMP-14 ΧΊ “λ –‘ ΜΙ–ηΫχ“Μ≤ΫΒΡ―–ΨΩ÷ΛΟς.¥ΥΆβΘ§”–ΕύΧθ–ρΝ–Θ§”»Τδ «ΒΎ 4 ¬÷ΒΡ–ρΝ–÷–Κ§”–ΫœΗΏ±»άΐΒΡHisΘ§Έ“Ο«ΦΑΤδΥϋ Β―ι “ΒΡ―–ΨΩ“―»Ζ»œΗΏΚ§ΝΩΒΡHis Ε‘”Ύ Zn2+Β»Ϋπ τάκΉ”ΨΏ”–«ΩΒΡ«ΉΚΆΝΠΘέ11ΘίΘ§Εχ Zn2 + ‘ρ«Γ « MMPs ¥ΏΜ·ΫαΙΙ”ρ÷Ί“ΣΒΡΫπ τάκΉ”Θ§Ηϋ « MMPs ΜνΜ·ΒΡΙΊΦϋ.
2. 3 MMP-14 ΫαΚœκΡ–ρΝ–ΒΡΆ§‘¥±»Ε‘ΦΑ“Μ÷¬–ρΝ–»ΖΕ®
Ε‘…Η―ΓΒΟΒΫΒΡ MMP-14 ΫαΚœκΡ–ρΝ–‘Ύ NCBI Ϋχ–– GenBank Blast Υ―ΥςΘ§Έ¥ΖΔœ÷ΤδΥϊ…ζΈοΧε¥φ‘ΎΆ§‘¥–ρΝ–. Υυ”–ΕύκΡ–ρΝ–Ψ≠Ψ≠―ιΖ÷άύΚσΫχ––Clustal X1. 81Εύ÷Ί–ρΝ–±»Ε‘Θ§ΫαΙϊΖΔœ÷¥φ‘ΎΩ…ΡήΒΡ“Μ÷¬–ρΝ–AHQLHΓΔHHXHΓΔL / IE PLL / I(X ¥ζ±μ»Έ“βΑ±ΜυΥα) Θ§ΦϊFig. 2Θ§Ή®άϊ…ξ«κΚ≈:200910161829XΘέ12Θί.

2. 4 “Μ÷¬–ρΝ–ΕύκΡ”κ MMP-14 ΒΡΖ÷Ή”ΡΘΡβΕ‘Ϋ”
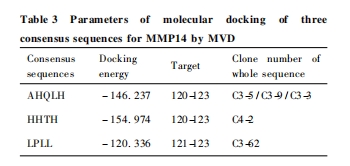
ΈΣ‘Λ≤βΚΆ»Ζ»œ…Η―ΓΒΟΒΫΒΡ“Μ÷¬–ρΝ–Ε‘MMP-14ΒΡΑ–œρ–‘Θ§―ûÓΜ÷¬–ρΝ– ÷– ¥ζ±μ–‘3Χθ–ρΝ–AHQLHΓΔHHTHΓΔLPLL Ϋχ –– Τδ”κ MMP-14 ΒΑ ΑΉ ΒΡ Ζ÷Ή”ΡΘΡβΕ‘Ϋ”. MMP-14 ¥ΏΜ·ΫαΙΙ«χ1bqqM(Α±ΜυΥαΥυ‘ΎΈΜ÷Ο:114ΓΪ227) ΚΆάύ―ΣΚλΥΊ Ϋα ΙΙ ”ρ 3c7xA( Α± ΜυΥαΥυ‘ΎΈΜ÷Ο:316 ΓΪ 511 ) ΒΡΫβΈωΈΣΉΦ»ΖΫχ––Ζ÷Ε‘Ϋ”ΕύκΡ¥χά¥ΝΥΦΪ¥σΒΡΩ…ΩΩ–‘. Ζ÷±π”κ MMP-14 ΒΡ ¥ΏΜ·ΫαΙΙ”ρΚΆάύ―ΣΚλΥΊΫαΙΙ”ρΒΡΖ÷Ή”Ε‘Ϋ”ΫαΙϊœ‘ ΨΘ§3Χθ“Μ÷¬–ρΝ–ΨυΡήΚήΚΟΒΊΕ‘Ϋ””Ύ MMP-14 ¥ΏΜ·ΫαΙΙ”ρΒΡ aa. 120ΓΪ125 «χ ”ρΘ§ΤδΚΥ–ΡΕ‘Ϋ”ΈΜΒψΈΣHis-Asn-GluΘ§’ΐ¥Π”Ύ MMP-14 ΒΡ Zn2 + ΫαΚœ«χ”ρ;Εχ”κ‘ΛΤΎœύΆ§Θ§Υυ”–“Μ÷¬–ρ Ν– ”κ MMP-14 ΒΡάύ―ΣΚλΥΊΫαΙΙ”ρ«χ”ρΕΦ ≤Μ Ρή Ε‘ Ϋ” ≥… ΙΠ. ¥ζ ±μ –‘ –ρ Ν– AHQLHΓΔHHTHΓΔLPLL ”κ MMP-14 ΒΡΖ÷Ή”Ε‘Ϋ””κΡήΝΩ±δΜ·Φϊ Fig. 3(Φϊ 347 “≥≤ “≥)ΚΆ Table 3.
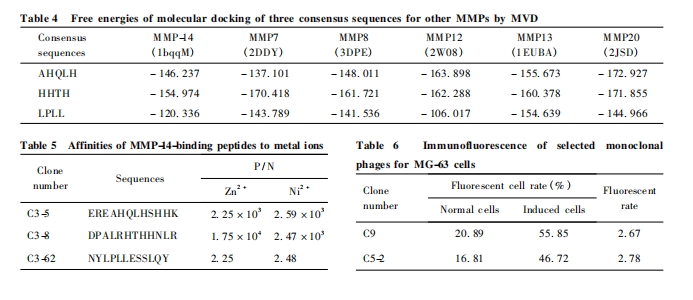
ΈΣΫχ“Μ≤Ϋ‘Λ≤βΚΆ»ΖΕ®“Μ÷¬–ρΝ–Ε‘MMP-14ΒΡΉ®“Μ–‘Θ§≤…”ΟάύΥΤΒΡΖΫΖ®ΨΆ AHQLHΓΔHHTHΓΔLPLL Β» 3Χθ“Μ÷¬–ρΝ–Ε‘ PDB ΐΨίΩβ÷–“―ΫβΈω»ΐΈ§ΫαΙΙΒΡ 14÷÷ MMPs ( MMP1-3Θ§MMP7-14Θ§MMP16Θ§MMP 20Θ§MMP 23)Ϋχ––ΝΥΖ÷Ή”Ε‘Ϋ”. ΫαΙϊΖΔœ÷:“Μ÷¬–ρΝ–Ω…”κMMP7-8Θ§MMP12-13 ΚΆ MMP20 Β»5÷÷MMPs Φ“ Ήε≥…‘±ΒΡ¥ΏΜ·ΫαΙΙ”ρΕ‘Ϋ”Θ§Εχ”κΤδΥϋ8÷÷ MMPs Φ“Ήε≥…‘±‘ρΕ‘Ϋ”Έό–ß. ’βΩ…Ρή”κ’β5÷÷MMPs ΒΡΜυ±ΨΫαΙΙ”–ΙΊ:MMP7 ”– Ή≈ ΉνΜυ±ΨΒΡΫαΙΙ”ρΘ§ΕχΤδΥϋ4÷÷MMPs ”–Ή≈±ξΉΦΒΡΫαΙΙ”ρΘέ13Θ§14Θί. 3 ÷÷“Μ÷¬–ρΝ–Ε‘Ϋ”ΚΆΡήΝΩ±δΜ·Β»Φϊ Fig. 4 ΚΆ Table 4.

2. 5 ΒΞΩΥ¬Γ …ΨζΧεΕ‘Ϋπ τάκΉ”ΒΡ«ΉΚΆΡήΝΠ
Φ»»ΜΖ÷Ή”Ε‘Ϋ”ΫαΙϊœ‘ Ψ3 ÷÷“Μ÷¬–ρΝ–ΡήΙΜ”κMMP-14 ΒΡ Zn2 + ΫαΚœ«χ”ρΚήΚΟΕ‘Ϋ”Θ§Έ“Ο«‘Λ≤βΗΟΒΞΩΥ¬Γ …ΨζΧεΥυΚ§ΫαΚœκΡΕ‘Ϋπ τάκΉ”Ω…ΡήΨΏ”–“ΜΕ®ΒΡ«ΉΚΆΝΠ. Κ§3÷÷ “Μ ÷¬ –ρ Ν–ΒΞΩΥ¬Γ …ΨζΧεά©‘ωΈοΕ‘Zn2 + ΓΔNi2 + ΒΡΫπ τ«ΉΚΆ≤ψΈω«ΉΚΆΝΠ≤βΕ®ΫαΙϊΦϊ Table5. ’ΐ »γ Έ“ Ο« ‘Λ ΤΎ ΒΡΘ§Υυ Φλ ≤β ΒΡ … Ψζ Χε Ε‘ Zn2 + ΚΆNi2 + Ψυ”–“ΜΕ®ΒΡ«ΉΚΆΝΠΘ§ΩΥ¬Γ C3-5Θ§C3-8Θ§”–Ή≈ΚήΗΏΒΡ«ΉΚΆΝΠΘ§ΫαΚœ“Μ÷¬–ρΝ–Ε‘ MMP-14 ΒΡΕ‘Ϋ”ΫαΙϊΘ§Ω…»œΈΣΈ“Ο«»ΖΕ®ΒΡ“Μ÷¬–ρΝ–ΨΏ”– MMP-14 ΒΑ ΑΉ ΚΆZn2 + Β»Ϋπ τΒΡΥΪΑ–œρΫαΚœΡήΝΠ.

2. 6 ΒΞΩΥ¬ΓΫαΚœκΡ …ΨζΧεΕ‘ MMP-14 ‘ΎœΗΑϊΥ°ΤΫΒΡΑ–œρ–‘
SABC-FITC Οβ“Ώ”ΪΙβΦλ≤βΒΞΩΥ¬ΓΫαΚœκΡ …ΨζΧεC3-5ΓΔC3-52 ”κ MMP-14 ”’ΒΦΚΆΖ«”’ΒΦ±μ¥οΒΡMG-63œΗΑϊΒΡΫαΚœΥ°ΤΫΘ§ΫαΙϊΦϊ Fig. 5. Ω…Φϊ”κΕ‘’’œύ±»Θ§2÷÷ …ΨζΧε”κMMP-14”’ΒΦ±μ¥οMG-63 œΗ Αϊ ΒΡ ”Ϊ Ιβ«ΩΕ»Οςœ‘ΗΏ”ΎΕ‘’’œΗΑϊΘ§Τδ”’ΒΦœΗΑϊΒΡ”ΪΙβ¬ Οςœ‘ΗΏ”ΎΈ¥”’ΒΦœΗΑϊΘ§Φϊ Table 6.

2. 7 “Μ÷¬–ρΝ–ΕύκΡΕ‘ MG-63 œΗΑϊΜνΝΠΒΡ“÷÷ΤΉς”Ο
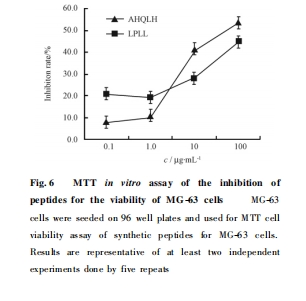
≤…”ΟMTTΖΫΖ®Φλ≤β Κœ ≥… ΒΡ “Μ ÷¬–ρΝ–ΕύκΡ Ε‘MG-63 œΗΑϊΜνΝΠΒΡ“÷÷ΤΉς”ΟΘ§Ι≤Κœ≥…ΚΆ≤βΕ®ΝΥ2 ΧθΕύκΡ–ρΝ– AHQLH ΚΆ LPLLΘ§ΫαΙϊΦϊ Fig. 6. Ω…ΦϊΥφΉ≈Κœ≥…ΕύκΡ≈®Ε»ΒΡ‘ωΦ”Θ§ΤδΕ‘œΗΑϊΒΡ“÷÷ΤΉς”Ο÷πΫΞ‘ω«ΩΘ§≥ œ÷≥ωΟςœ‘ΒΡΦΝΝΩ“άάΒΙΊœΒ. Τδ÷– AHQLH ‘Ύ≈®Ε»ΈΣ 10 ΠΧg /mL œ¬ Ε‘œΗΑϊΒΡ“÷÷Τ¬ Φ¥ Ω…¥οΒΫ41. 6% .
3 Χ÷¬έ
Ϋϋ 20 Ρξά¥Θ§Μυ”Ύ MMPs ΒΡΕύ÷÷ΩΙ÷ΉΝω“÷÷ΤΦΝœ»ΚσΫχ»κΝΌ¥≤ ‘―ιΘ§ΒΪ¥σΕύ“ρ“©ΈοΗ±Ής”Ο¥σ /―Γ‘ώ–‘≤νΕχ‘ΎΝΌ¥≤ ‘―ιΒΡΔρΓΔΔσΤΎ±ΜΖώΨω. Τδ÷–ΒΡ“ΜΗω÷Ί“Σ‘≠“ρ «“ρΈΣ’β–©“÷÷ΤΦΝΨΏ”–ΙψΤΉΒΡΧΊ“λ–‘Θ§≤ΜΫω“÷÷ΤΝΥΑ–ΟΗΘ§“≤“÷÷ΤΝΥ“Μ–©”–“φΟΗΘέ15Θί. ΤυΫώΈΣ÷ΙΘ§÷Μ”– 1ΗωΜυ”Ύ MMPs ΒΡ Doxycycline Ά®Ιΐ FDA »œ÷Λ.
ΩΦ¬« MMP-14 ±Μ»œΈΣ «”κ÷ΉΝωΖΔ…ζΙΊœΒΉνΟή«–ΒΡ“Μ÷÷ MMPsΘ§Έ“Ο«―Γ‘ώ MMP-14 ΈΣΑ–±ξΫχ–– …ΨζΧεΥφΜζκΡΩβΒΡ…Η―Γ”κ―–ΨΩΘ§ΤΎΆϊΆ®Ιΐ …ΨζΧεΥφΜζ °ΕΰκΡΩβ…Η―ΓΚΆΖ÷Ή”ΡΘΡβΓΔœΗΑϊ in vitro Β»―–ΨΩ»ΖΕ®ΗΏ–ßΚΆΉ®“Μ–‘ΒΡ MMP-14 “÷÷ΤΦΝΘ§¥”Εχ”Ο”ΎΑ–œρ MMP-14œ»ΒΦ“©ΈοΒΡ―–ΖΔ. “‘ MMP-14 ¥ΏΜ·«χΒΑΑΉΈΣΑ–±ξΒΡ …ΨζΧεΥφΜζκΡΩβ…Η―ΓΘ§Έ“Ο«ΒΟΒΫΝΥ …ΨζΧεΒΡΗΜΦ·Θ§ΒΪΉήΧε P /N ÷ΒΫœΒΆ( < 8) Θ§«“ΒΎ4¬÷ΒΆ”ΎΒΎ3¬÷Θ§Εχ‘ΎΒΎ5¬÷ P /N ÷Β“≤Έ¥”–ΧαΗΏ(Ή ΝœΈ¥ΖΔ±μ) Θ§ΈΣ¥ΥΈ“Ο«―Γ»ΓΝΥΒΎ3ΓΔ4 ¬÷ΒΡΗΜΦ· …ΨζΧεΫχ––≤β–ρΘ§ΩΦ¬«ΒΎ4 ¬÷ΒΡΕύκΡ“―Κ§ΕύΗωHisΘ§ΕχΈ“Ο««ΑΤΎΒΡ―–ΨΩ“―Ψ≠ΟςΘ§His ΒΡ¥φ‘ΎΕ‘ Αϋ ά® Zn2 + ‘ΎΡΎΒΡΕύ÷÷Ϋπ τάκΉ”«ΩΒΡ«ΉΚΆΝΠΘέ11Θ§16ΘίΘ§œ‘»ΜΚ§ΕύΗω His ΒΡ–ρΝ–ΧΊ’ς≤Μ “ΥΉςΈΣ MMPs ΒΡΉ®“Μ–‘“÷÷ΤΦΝ. ΈΣ¥ΥΘ§Έ“Ο«Ε‘ΒΎ 3 ¬÷ΗΜΦ·ΒΡ …ΨζΧεΫχ––ΝΥ¥σΝΩΒΡ≤β–ρ. »γΈ“Ο«ΥυΤΎΆϊΘ§ΜώΒΟΝΥΕύΧθΚ§“Μ÷¬–ρΝ–ΒΡΕύκΡΘ§ΕύκΡ–ρΝ–ΥυΚ§ AlaΓΔLeu Β»Α±ΜυΥα ”κ MMP-14 ΒΉΈοΒΑΑΉΒΡœύΥΤ–‘Θ§“≤ Ϋ“ Ψ±Ψ…Η―ΓΫαΙϊΒΡΩ…ΩΩ–‘Θ§Ηϋ«“ΝνΈ“Ο«–ΥΖήΒΡ «Θ§Ά®Ιΐ«ΉΚΆΝΠΖ¥…ΗΘ§ΒΟΒΫΝΥΕύΧθΗΏP/N ÷Β ΒΡΫαΚœκΡΘ§ΤδΉνΗΏΒΞΧθ …ΨζΧε«ΉΚΆΝΠ¥οΒΫ 4024. 39Θ§‘ΕΗΏ”ΎΗΟ¬÷ΒΡΗΜΦ·±Ε ΐ( P /N = 7. 93)
ΈΣΝΥΫχ“Μ≤Ϋ‘Λ≤βΚΆ»Ζ»œΥυΒΟΒΫΒΡ“Μ÷¬–ρΝ–”κMMP-14 «Ζώ¥φ‘ΎœύΜΞΉς”Ο“‘ΦΑ»ΖΕ®‘Ύ MMP-14 …œœύΜΞΉς”ΟΒΡΫαΙΙ”ρΘ§Έ“Ο«≤…”ΟΦΤΥψΜζΖ÷Ή”ΡΘΡβ”κΕ‘Ϋ”ΚΆœΗΑϊΟβ“Ώ”ΪΙβΝΫ÷÷ΖΫΖ®Ϋχ––ΝΥœύΙΊΖ÷Έω. Αϋά®¥ΏΜ·ΫαΙΙ”ρΓΔάύ―ΣΚλΥΊΫαΙΙ”ρ‘ΎΡΎΒΡ MMP-14 ΒΑΑΉΒΡΕύΗω«χ”ρ“‘ΦΑΕύ÷÷ MMPs Φ“Ήε≥…‘±»ΐΈ§ΫαΙΙΒΡΆξ»ΪΫβΈωΘ§ΈΣΈ“Ο«Ϋχ––ΨΪ»ΖΒΡΖ÷Ή”Ε‘Ϋ”ΧαΙ©ΝΥΨό¥σΒΡΖΫ±ψ”κΜυ¥Γ. Έ“Ο«¥”≥θ≤Ϋ»ΖΕ®ΒΡ“Μ÷¬–ρΝ–÷–―Γ»ΓΝΥ3 ΧθΕΧκΡ(AHQLHΘ§HHTHΘ§LPLL) Ϋχ––ΝΥΤδ”κ MMP-14 ΒΡΦΤΥψΜζΖ÷Ή”ΡΘΡβ”κΕ‘Ϋ”Θ§Ε‘Ϋ”ΫαΙϊ“ΜΖΫΟφΫχ“Μ≤Ϋ»Ζ»œΝΥ 3 Χθ“Μ÷¬–ρΝ–”κMMP-14”––ßΫαΚœΘ§Εχ«“»ΖΕ®ΝΥΤδΖ÷Ή”Ε‘Ϋ”ΈΜΒψ‘ΎMMP-14 ¥Ώ Μ· Ϋα ΙΙ ”ρ ΒΡHis-Asn-Glu ΈΜΒψΘ§”κHXΕ‘Ϋ”ΒΡ≤Μ≥… ΙΠΘ§¥”Νμ“ΜΫ«Ε»÷ΛΟςΝΥ…Η―ΓΜώΒΟΒΡ“Μ÷¬–ρΝ–ΕύκΡ”κMMP-14 ¥ΏΜ·ΫαΙΙ”ρΒΡΉ®“Μ –‘ΫαΚœΉς ”ΟΘ§Ά§―υ“≤÷ΛΟςΝΥΈ“Ο«“‘MMP-14ΒΡ¥ΏΜ·ΫαΙΙ«χΈΣΑ–±ξΥφΜζκΡΩβ…Η―ΓΒΡΉΦ»Ζ–‘. ≤ΜΨΓ»Υ“βΒΡ «Θ§ΗΟ3Χθ“Μ÷¬–ρΝ–”κ 14 ÷÷“―ΫβΈω»ΐΈ§ΫαΙΙΒΡ MMPs ÷–ΒΡ 5 ÷÷MMPs “≤ΡήΙΜ”––ßΖ÷Ή”Ε‘Ϋ”Θ§Χα ΨΈ“Ο«–ηΫχ“Μ≤ΫΒΡ”≈Μ·ΗΟ 3 Χθ“Μ÷¬–ρΝ–“‘ΧαΗΏΤδΕ‘MMP-14 ΒΡΉ®“Μ–‘. Ϋχ“Μ≤ΫΒΡ‘ΎœΗΑϊΥ°ΤΫΒΡΟβ“Ώ”ΪΙβΦλ≤β“≤»Ζ»œΫαΚœκΡ …ΨζΧε”κMMP-14 ΒΡ”––ßΫαΚœ. MG-63 œΗΑϊΒΡ“ΜΗω÷Ί“ΣΧΊ’ς «‘Ύ¥ΤΦΛΥΊΒΡ”’ ΒΦœ¬Ω…“‘ΙΐΝΩ±μ¥οMMP-14 ( Β ―ι ΐ Ψί Έ¥ΖΔ±μ) Θ§Ά®Ιΐ”’ΒΦΚΆΖ«”’ΒΦ±μ¥ο MG-63œΗΑϊΒΡΟβ“Ώ”ΪΙβΦλ≤βΘ§Έ“Ο«‘ΎœΗΑϊΥ°ΤΫ»Ζ»œΝΥΒΞΩΥ¬ΓΫαΚœκΡ …ΨζΧε «Ά®ΙΐΫαΚœκΡ”κœΗΑϊ±μΟφ”’ΒΦ±μ¥οΒΡ MMP-14 ΒΡΫαΚœΕχΖΔΜ”Ής”ΟΒΡ. ΝμΆβΘ§Ζ÷Ή”Ε‘Ϋ”ΫαΙϊΆ§ ±ΜΙΫ“ ΨΘ§Έ“Ο«…Η―ΓΜώΒΟ≤ΩΖ÷ΕύκΡΩ…ΡήΜΙ”κΫπ τάκΉ”ΨΏ”–Α–œρΫαΚœΉς”ΟΘ§ΈΣ¥ΥΘ§Έ“Ο«Ϋχ––ΒΡΫπ τάκΉ”«ΉΚΆΝΠ≤βΕ®»Ζ»œΝΥΗΟ÷÷Ής”ΟΘ§»Ζ»œ≤ΩΖ÷“Μ÷¬–ρΝ–ΨΏ”– MMP-14 ΚΆ Ϋπ τάκΉ”ΥΪΑ–œρΒΡΉς”Ο. ’β «Ζ«≥Θ÷Ί“ΣΒΡΖΔœ÷Θ§“ρΈΣœ÷––―–ΖΔΒΡΑ–œρ MMPs ΒΡ“÷÷ΤΦΝ¥σΕύ « ‘Ύ“÷÷ΤΖ÷Ή”ΫαΙΙΒΡΜυ¥Γ…œ≈ΦΝΣΫπ τάκΉ”ρϋΚœΦΝΕχ–Έ≥…Θέ1ΘίΘ§“Μ÷¬–ρΝ–ΒΡΫπ τάκΉ”«ΉΚΆΧΊ’ςΩ…“‘±ήΟβΫπ τάκΉ”ρϋΚœΦΝΒΡ Ι”ΟΕχΩ…Άϊ‘ΎΫΒΒΆΕΨΗ±Ής”ΟΖΫΟφΖΔΜ”÷Ί“ΣΉς”Ο.
‘Ύ“‘…œΙΛΉςΒΡΜυ¥Γ…œΘ§Έ“Ο«ΨΆ…Η―Γ“Μ÷¬–ρΝ–Ϋχ––ΝΥ≥θ≤ΫΒΡœΗΑϊΜνΝΠ “÷ ÷Τ ≤βΕ®Θ§ΩΦ¬«ΕύΨέHis ΕΧ κΡΕ‘Ϋπ τάκΉ”ΙψΤΉΒΡ«ΉΚΆΧΊ–‘Θ§œ‘»Μ≤Μ“ΥΉςΈΣΑ–œρœ»ΒΦΕύκΡ“©Έο. ΈΣ¥ΥΘ§Έ“Ο«Κœ≥…ΝΥ AHQLHΘ§LPLLΝΫΧθΕΧκΡΘ§≥θ≤ΫΒΡΧεΆβœΗΑϊ“÷÷Τ―–ΨΩ÷ΛΟςΥυ≤βΕ®ΒΡΝΫΧθΕΧκΡΕ‘ MG-63 œΗΑϊΨΏ”–“ΜΕ®ΒΡœΗΑϊΜνΝΠ“÷÷ΤΉς”ΟΘ§ΒΪ”κΤΎΆϊΉςΈΣœ»ΒΦ“©Έο≈®Ε»»‘¥φ‘ΎΫœ¥σ≤νΨύΘ§Ϋχ“Μ≤ΫΒΡΕΧκΡ”≈Μ·”κΗΡ‘λ»‘‘ΎΫχ’Ι÷–.
4 ÷ς“ΣΫα¬έ
±Ψ―–ΨΩ“‘MMP-14 ΒΑΑΉ¥ΏΜ·ΫαΙΙ”ρΈΣΑ–±ξΘ§Ά®Ιΐ …ΨζΧεΥφΜζ °ΕΰκΡΩβ…Η―ΓΚΆΖ÷Ή”ΡΘΡβΓΔœΗΑϊΟβ“Ώ”ΪΙβΓΔΫπ τάκΉ”«ΉΚΆ≤ψΈω“‘ΦΑΧεΆβœΗΑϊΉς”Ο≤βΕ®Β»ΦΦ θΫχ––ΝΥΥΪΑ–œρ MMP-14 –ΓΖ÷Ή”ΫαΚœΕύκΡΒΡ…Η―Γ”κ―–ΨΩ. ≥θ≤Ϋ»ΖΕ®ΝΥΥΪΑ–œρMMP-14 ΚΆ Zn2 + Β»Ϋπ τάκΉ”ΒΡAHQLHΓΔHHXHΓΔEI / L PLL / I Β»ΕύΧθ“Μ ÷¬–ρΝ–Θ§»ΖΕ®ΝΥAHQLHΓΔHHTHΓΔLPLL Β» 3 Χθ“Μ÷¬–ρΝ–”κMMP-14 ΒΡ”––ßΕ‘Ϋ”ΦΑ¥ΏΜ·ΫαΙΙ”ρΫαΚœΈΜΒψ“‘ΦΑ“ΜΕ®ΒΡ MMP-14Ή®“Μ –‘. ΧεΆβœΗΑϊ≤βΕ®≥θ≤Ϋ»Ζ»œAHQLHΓΔLPLL Ε‘ MG63 ΨΏ”–“ΜΕ®ΒΡœΗΑϊΜνΝΠ“÷÷ΤΉς”Ο. ±ΨΙΛΉςΈΣ…ν»κΩΣ’ΙMMP-14 “÷÷ΤΦΝ”κ÷ΉΝωœύΜΞΉς”ΟΒΡ―–ΨΩΘ§“‘ΦΑΥΪ Α–œρ MMP-14ΚΆΫπ τάκΉ”ΒΡΩΙ÷ΉΝω–ΓΖ÷Ή”ΕύκΡ“÷÷ΤΦΝΒΡ―–ΖΔΧαΙ©ΝΥ÷Ί“ΣΒΡ«ΑΤΎΙΛΉςΜυ¥Γ.
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§Αφ»®Ιι‘≠Ής’ΏΦΑ‘≠‘”÷ΨΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘΈΡ’¬±ξΉΔ”–Ής’ΏΦΑΈΡ’¬≥ω¥ΠΘ§»γ–η‘ΡΕΝ‘≠ΈΡΦΑ≤ΈΩΦΈΡœΉΘ§Ω…‘ΡΕΝ‘≠‘”÷Ψ
