Ής’ΏΘΚ««―©, Άθ“ε≈τ, ”ΎΚΘΡΰ, Β». …ζΈοΙΛ≥Χ―ß±®, 2019, 35(8): 1391-1400Θ§≤ΈΩΦΈΡœΉΩ…‘ΡΕΝΤΎΩ·‘≠ΈΡ
’Σ“Σ
ΫϋΡξά¥Α©÷ΔΒΡΖΔ…ζ¬ ΚΆΥάΆω¬ ≥ œ÷÷πΫΞ…œ…ΐΒΡ«ς ΤΘ§ «Άΰ–≤»Υάύ…ζΟϋΒΡ÷ς“ΣΦ≤≤Γ÷°“ΜΓΘΩΙΑ©κΡ(Anticancer peptidesΘ§ACPs)Φ¥ΨΏ”–ΩΙ÷ΉΝωΜν–‘ΒΡ…ζΈοΜν–‘κΡΘ§ΤδΙψΖΚ¥φ‘Ύ”ΎΕύ÷÷…ζΈοΧεΡΎΘ§Αϋά®≤Η»ιΕ·ΈοΓΔΝΫΤήάύΕ·ΈοΓΔάΞ≥φΓΔ÷≤ΈοΚΆΈΔ…ζΈοΒ»ΓΘΩΙΑ©κΡ‘Ύ÷ΈΝΤ÷ΉΝωΖΫΟφΨΏ”–÷ΎΕύ”≈ ΤΘ§»γΖ÷Ή”ΝΩΒΆΓΔΫαΙΙΦρΒΞΓΔΗΏΩΙΑ©Μν–‘ΓΔΗΏ―Γ‘ώ–‘ΓΔΫœ…ΌΒΡΗ±Ής”ΟΓΔΕύ÷÷Ηχ“©ΖΫ ΫΓΔ≤Μ“Ή“ΐΤπΕύ÷ΊΡΆ“©–‘Β»ΓΘΈΡ÷–ΫαΚœ±ΨΩΈΧβΉιœύΙΊΙΛΉςΘ§ΙιΡ…ΝΥΡΩ«ΑΥυΖΔœ÷ΒΡΩΙΑ©κΡΒΡΉς”ΟΜζ÷ΤΘ§“‘ΤΎΈΣ–¬–ΆκΡάύΩΙ÷ΉΝω“©ΈοΒΡ―–ΖΔΧαΙ©“ΜΕ®ΒΡΖΫœρΓΘ
Α©÷Δ“≤≥ΤΈΣΕώ–‘÷ΉΝωΘ§ «“Μ÷÷”…ΒςΩΊœΗΑϊΖ÷Ν―‘ω÷≥Μζ÷Τ ß≥ΘΕχ“ΐΤπΒΡΦ≤≤ΓΓΘΨίΙζΦ Α©÷Δ―–ΨΩΜζΙΙΙάΦΤΘ§‘Ύ»Ϊ άΫγΖΕΈßΡΎΘ§2018Ρξ–¬‘ωΑ©÷ΔΜΦ’Ώ‘ΦΈΣ1 810ΆρΘ§ΥάΆω»Υ ΐ‘Φ960Άρ[1]ΓΘΨίΙάΦΤ‘Ύ2025Ρξ»Ϊ«ρΟΩΡξΒΡ–¬‘ωΑ©÷Δ≤ΓάΐΫΪ≥§Ιΐ2 000Άρ[2]ΓΘΡΩ«ΑΘ§ ÷ θ÷ΈΝΤΓΔΖ≈…δ÷ΈΝΤ(Ζ≈ΝΤ)ΚΆΜ·―ß“©Έο÷ΈΝΤ(Μ·ΝΤ) «÷ς“ΣΒΡ¥ΪΆ≥Α©÷Δ÷ΈΝΤΖΫΖ®ΓΘ»ΜΕχ¥ΪΆ≥ΒΡΩΙΑ©άύ“©ΈοΥδ»ΜΩΙΑ©–߬ ΫœΗΏΘ§ΒΪ «“≤¥φ‘Ύ“Μ–©»±ΒψΘ§»γΒΆ―Γ‘ώ–‘ΓΔΗ±Ής”ΟΟςœ‘ΓΔΟβ“Ώ“÷÷ΤΓΔ…ώΨ≠ΚΆ≥ΠΈΗΥπ…ΥΒ»[3-4]ΘΜΗϋ÷Ί“ΣΒΡ «Θ§’β–©ΩΙΑ©άύ“©ΈοΒΡΝΣΚœ Ι”ΟΦΪ“Ή“ΐΤπ÷ΉΝωΒΡΕύ“©ΡΆ“©–‘(MDR)[5]ΓΘ“ρ¥ΥΘ§÷ΉΝωΒΡ“©Έο÷ΈΝΤΤΎ¥ΐΉ≈–¬ΒΡΆΜΤΤΓΘΩΙΨζκΡ(AMPs) «“ΜάύΧλ»Μ≤ζ…ζΒΡœ»ΧλΟβ“ΏΒΡ÷Ί“ΣΖά”υΈο÷ Θ§ΩΙΨζκΡΙΠΡήΕύ―υΘ§Τδ÷–ΫΪΨΏ”–ΩΙ÷ΉΝωΜν–‘ΒΡΩΙΨζκΡΫ–ΉωΩΙΑ©κΡ(ACPs)ΓΘΕχΩΙΑ©κΡΒΡΧΊ βΉς”ΟΜζ÷ΤΘ§ ΙΤδ≥…ΈΣΫϋΡξά¥…ζΈο“©Έο―–ΨΩ÷–ΒΡ“ΜΗω»»ΒψΘ§“≤ΈΣ–¬–ΆΩΙΑ©“©ΈοΒΡ―–ΨΩΧαΙ©ΝΥ–¬ΖΫœρΓΘ
1 ΩΙΑ©κΡΒΡΜυ±ΨΧΊ’ς
ΡΩ«ΑΖΔœ÷ΒΡΩΙΑ©κΡΩ…Ζ÷ΈΣΝΫάύΘΚ“Μάύ «Ε‘œΗΨζΚΆΑ©œΗΑϊ”–…±…ΥΉς”ΟΘ§ΒΪ «Ε‘’ΐ≥ΘœΗΑϊΟΜ”–ΕΨ–‘Θ§»γcecropinsΚΆmagaininsΓΘΝμ“Μάύ «Ε‘œΗΨζΓΔΑ©œΗΑϊΚΆ’ΐ≥ΘœΗΑϊΨυ”–ΤΤΜΒΉς”ΟΘ§»γάΞ≥φdefensinsΚΆtachyplesin ΔρΒ»[6]ΓΘ“ρ¥ΥΒΎ“ΜάύΩΙΑ©κΡΨΏ”–ΚήΚΟΒΡ―–ΨΩΦέ÷ΒΚΆ”Π”Ο«ΑΨΑΓΘΩΙΑ©κΡΒΡ≥ΛΕ»ΚΆ–ρΝ–Εύ―υΘ§ΒΪ¥σ≤ΩΖ÷ΩΙΑ©κΡΕΦ”–ΝΫΗωΙ≤Ά§ΧΊ’ςΘΚ―τάκΉ”–‘ΚΆΝΫ«Ή–‘ΓΘΩΙΑ©κΡΆ®≥Θ”…5®C40ΗωΑ±ΜυΥαΉι≥…Θ§Τδ÷–ΨΪΑ±ΥαΓΔάΒΑ±ΥαΚΆΉιΑ±ΥαΒΡ¥φ‘Ύ ΙΤδ±μœ÷≥ωΫœ«ΩΒΡ―τάκΉ”ΧΊ–‘Θ§±μΟφΨΜΒγΚ…ΖΕΈßΈΣ+2®C+9[7-8]ΓΘΩΙΑ©κΡΦ»ΨΏ”–«ΉΥ°–‘”÷ΨΏ”–«Ή÷§–‘Θ§÷ς“Σ «“ρΈΣΤδΫαΙΙ…œΨΏ”–«ΉΥ°–‘ΚΆ ηΥ°–‘ΒΡ≤ύΝ¥ΫαΙΙΘ§ΕχΨχ¥σΕύ ΐΒΡΩΙΑ©κΡΚ§”–ΠΝ-¬ί–ΐΜρΠ¬-’έΒΰΫαΙΙΘ§’β÷÷ΝΫ«Ή–‘≤ύΝ¥‘ΎΨΏ”–ΠΝ-¬ί–ΐΫαΙΙΒΡΩΙΑ©κΡΝ¥…œΖ÷±π≈≈≤Φ‘Ύ¬ί–ΐΒΡΝΫ≤ύΘ§Μρ’ΏΦ·÷–”ΎΝΫΕΥΘ§“ρ¥ΥΩ…–Έ≥…«ΉΥ°ΟφΚΆ ηΥ°ΟφΜρ’ΏΟςœ‘ΒΡ«ΉΥ°ΕΥΚΆ ηΥ°ΕΥ[9]ΓΘΒ±ΩΙΑ©κΡ”κΑ©œΗΑϊΡΛœύΜΞΉς”Ο ±Θ§ ηΥ°«χ”ρ”κΑϊΡΛ÷§÷ ΫαΚœΘ§¥χ’ΐΒγΚ…ΒΡ«ΉΥ°«χ”ρ”κ¥χ”–ΗΚΒγΚ…ΒΡΑ©œΗΑϊΡΛ±μΟφΆ®ΙΐΨ≤ΒγΈϋΗΫΕχ”––ßΫαΚœΘ§’βΈΣΩΙΑ©κΡΡήΙΜ―Γ‘ώ–‘Ής”Ο”ΎΑ©œΗΑϊΒλΕ®ΝΥΜυ¥Γ[10]ΓΘ
2 ΩΙΑ©κΡΒΡΉς”ΟΜζ÷Τ
2.1 ΩΙΑ©κΡΒΡΡΛΝ―ΫβΜζ÷Τ
ΩΙΑ©κΡΡήΙΜΑ–œρΉς”Ο”ΎΑ©œΗΑϊΘ§Εχ≤ΜΥπ…Υ’ΐ≥ΘΉι÷·œΗΑϊΒΡ÷ς“Σ‘≠“ρ‘Ύ”ΎΑ©œΗΑϊΡΛ±μΟφ“Μ–©“θάκΉ”≥…Ζ÷ΒΡΧΊ“λ–‘±μ¥οΘ§Αϋά®ΝΉ÷§θΘΥΩΑ±ΥαΓΔO-Χ«ΜυΜ·’≥ΒΑΑΉΓΔΆΌ“ΚΥα…ώΨ≠ΫΎή’÷§ΚΆΗΈΥΊΒ»[11]ΓΘ¥σΝΩ―–ΨΩ±μΟςΘ§ΩΙΑ©κΡΒΡΉς”ΟΜζ÷ΤΩ…“‘Ζ÷ΈΣΡΛΝ―ΫβΜζ÷ΤΚΆΖ«ΡΛΝ―ΫβΜζ÷Τ(ΆΦ 1ΚΆΆΦ 2)ΓΘΤδ÷–ΩΙΑ©κΡΒΡΡΛΝ―ΫβΜζ÷Τ÷ς“Σ”–“‘œ¬3÷÷Θ§Ζ÷±π «ΓΑΆΑΑεΡΘ–ΆΓ±ΓΔΓΑ’±ΧΚΡΘ–ΆΓ±ΚΆΓΑΜΖ–ΈΩΉΡΘ–ΆΓ±(ΆΦ 1)[12-13]ΓΘ
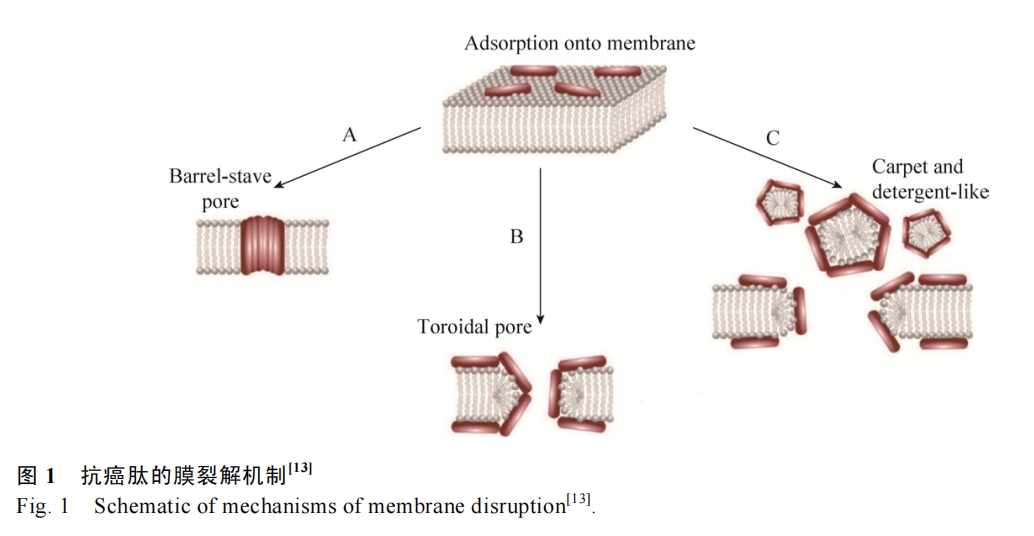
2.1.1 ΆΑΑεΡΘ–Ά
1977ΡξEhrensteinΒ» Ή¥ΈΧα≥ωΝΥΩΙΨζκΡΒΡΆΑΑεΡΘ–Ά(ΆΦ 1)[14]ΓΘΩΙΑ©κΡ‘ΎΑ©œΗΑϊΡΛ±μΟφΆ®Ιΐ ηΥ°Ής”ΟΙ―ΨέΧεΜ·Θ§Τδ÷–ΩΙΑ©κΡΒΡ ηΥ°ΟφœρΆβ≥·œρœΗΑϊΡΛΒΡθΘΜυΝ¥Θ§Εχ«ΉΥ°Οφ–Έ≥…ΩΉΜρ≤έΘ§Ήν÷’‘Ύ≤ΜΕœΒΡΨέΦ·Ιΐ≥Χ÷––Έ≥…¥©ΆΗ÷ ΡΛάκΉ”ΩΉΒάΘ§ΫχΕχΒΦ÷¬Α©œΗΑϊΡΎ»ίΈοΝς≥ωΘ§ ß»Ξ¥σΝΩάκΉ”ΚΆΡήΝΩΘ§ΑϊΡΎ…χΆΗ―ΙΗΡ±δΘ§ΫχΕχΆΏΫβœΗΑϊΡΛΓΘάμ¬έ…œ’β÷÷ΩΉΒά÷Ν…Ό”…3ΗωΩΙΑ©κΡΖ÷Ή”Ήι≥…Θ§≤Δ«““Σ«σΥϋΟ«”–“ΜΕ®ΒΡΕΰΦΕΫαΙΙΘ§±»»γΝΫ«Ή–‘ΒΡΠΝ-¬ί–ΐΓΔΠ¬-’έΒΰΜρΆ§ ±Κ§”–ΠΝ-¬ί–ΐΚΆΠ¬-’έΒΰ[15]ΓΘΚήΕύΩΙΑ©κΡΨυ±Μ÷ΛΟς «Ά®ΙΐΆΑΑεΡΘ–Άά¥ΖΔΜ”ΤδΩΙΑ©Ής”ΟΒΡΓΘ»γ‘Ύ1994ΡξΘ§SuiΒ»÷ΛΟςΝΥΖ÷άκΉ‘“βΖδApis melliferaΒΡMelittinΩ…“‘Ά®ΙΐΆΑΑεΡΘ–ΆΤΤΜΒΑ©œΗΑϊΡΛΒΡΆξ’ϊ–‘[16]ΓΘMelittin”÷ΟϊΖδΕΨκΡΘ§ «ΖδΕΨΒΡ÷ς“Σ≥…Ζ÷Θ§”…26ΗωΑ±ΜυΥα≤–ΜυΉι≥…Θ§ΤδΙΠΡήΕύ―υΘ§»γ”–ΩΙ―ΉΓΔ’ρΆ¥ΓΔΩΙΨζΓΔΩΙHIVΦΑΩΙ÷ΉΝωΒ»Εύ÷÷“©άμΜν–‘[17]ΓΘΖδΕΨκΡΨΏ”–ΙψΤΉΒΡΩΙ÷ΉΝωΜν–‘Θ§Αϋά®»ΥΗΈœΗΑϊΑ©ΓΔΑΉ―Σ≤ΓΓΔ»ιœΌΑ©Β»Θ§ΆΑΑεΡΘ–Ά «ΤδΕύ÷÷ΩΙ÷ΉΝωΜζ÷Τ÷°“Μ[18-19]ΓΘά¥Ή‘≈Ίœχ≤ίΆήLitoria raniformisΒΡΩΙΑ©κΡAurein1.2Ε‘ΑΉ―Σ≤ΓΓΔΖΈΑ©ΓΔΫα≥ΠΑ©Β»Ψυ”–…±…ΥΉς”ΟΘ§Τδ÷ς“ΣΜζ÷Τ“≤ «Ά®ΙΐΆΑΑεΡΘ–Άά¥ΤΤΜΒ÷ΉΝωœΗΑϊ[20]ΓΘ
2.1.2 ’±ΧΚΡΘ–Ά
1992ΡξPounyΒ»Χα≥ωΝΥΩΙΨζκΡΒΡΓΑ’±ΧΚΓ±ΡΘ–Ά[21]ΓΘ―τάκΉ”ΩΙΑ©κΡ“≤Ω…“‘Ά®ΙΐΨ≤ΒγΉς”ΟΫαΚœΒΫ¥χΗΚάκΉ”ΒΡΑ©œΗΑϊΡΛ…œΘ§“‘άύΥΤ’±ΧΚΒΡΫαΙΙΤΫ––≈≈Ν–Θ§Β±ΩΙΑ©κΡ¥οΒΫΝΌΫγ≈®Ε» ±Θ§œΗΑϊΡΛΡήΝΩΕώΜ·Θ§Έ»Ε®–‘ΫΒΒΆΕχ≥ωœ÷œ‘÷χΒΡΆδ«ζ¥”ΕχΤΤΝ―(ΆΦ 1)ΓΘ«χ±π”ΎΆΑΑεΡΘ–ΆΘ§’±ΧΚΡΘ–Ά≤Μ–η“ΣΩΙΑ©κΡΨΏ”–ΧΊ βΫαΙΙΘ§≤Δ«“≤Μ–Έ≥…ΩγΡΛΆ®Βά[15]ΓΘCecropinsάύΩΙΑ©κΡΘ§”÷ΟϊΧλ≤œΥΊΘ§ «ΒΎ“ΜΗω±ΜΖΔœ÷ΒΡΕ·ΈοΩΙΨζκΡΘ§‘ΎάΞ≥φΚΆ≤Η»ιάύΕ·Έο÷–Ψυ”–ΖΔœ÷Θ§ΤδΕ‘ΑΉ―Σ≤ΓΓΔΑρκΉΑ©Β»Ψυ”–«Ω…±…ΥΉς”ΟΘ§Ω…Ά®Ιΐ’±ΧΚΡΘ–ΆΖΔΜ”Ής”Ο[22]ΓΘChuangΒ»±®ΒάΝΥ»ΥΧεΈ®“Μ“Μ÷÷CathelicidinάύΩΙΨζκΡLL-37Θ§“≤Ω…“‘Ά®Ιΐ’±ΧΚΡΘ–Ά―Γ‘ώ–‘ΒΊΝ―Ϋβ¬―≥≤Α©[23]ΓΘΝμΆβΘ§Magainins (ά¥Ή‘Ζ«÷όΉΠσΗΘ§Xenopus laevis)[24]ΓΔCitropin 1.1 (ά¥Ή‘”ξ±θΆήΘ§Litoria citropa)[25]ΓΔGaegurins (ά¥Ή‘÷εΤΛΆήΘ§Rana rugosa)[26]Β»Εύ÷÷ΩΙΑ©κΡΘ§ΨυΩ…Ά®Ιΐ’±ΧΚΡΘ–Άά¥ΖΔΜ”ΤδΩΙΑ©Ής”ΟΓΘ
2.1.3 ΜΖ–ΈΩΉΡΘ–Ά
‘ΎΜΖ–ΈΩΉΡΘ–Ά÷–Θ§ΩΙΑ©κΡΒΡ ηΥ°«χ”κΑ©œΗΑϊΡΛ…œΒΡ ηΥ°«χœύΜΞ“ΤΕ·ΕχΒΦ÷¬ΑϊΡΛΤΤΝ―»± ßΘ§Ήν÷’–Έ≥…ΩγΡΛΩΉΒά(ΆΦ 1)ΓΘΤδ”κΆΑΑεΡΘ–ΆΉν÷ς“ΣΒΡ«χ±π‘Ύ”ΎΩΙΑ©κΡ Φ÷’”κΝΉ÷§ΒΡΆΖ≤ΩΫαΚœΕχ“ΜΤπΙΙ≥…ΩγΡΛΆ®ΒάΓΘ1997ΡξΘ§MatsuzakiΒ»±®ΒάΝΥMagainin-2Ω…“‘ΓΑΜΖ–ΈΩΉΓ±ΡΘ–ΆΖΔΜ”ΩΙΨζΉς”Ο[27]ΓΘMagaininsΘ§”÷ΟϊΉΠσΗΥΊΘ§Ζ÷άκΉ‘Ζ«÷όΉΠσΗΒΡΤΛΖτΘ§ «Ϋœ‘γΖΔœ÷ΒΡΝΫΤήΕ·ΈοΩΙΨζκΡΘ§ΤδΨΏ”–ΙψΤΉΒΡΩΙΨζΩΙΑ©Μν–‘Θ§Τδ÷–Ω…“‘Ά®ΙΐΜΖ–ΈΩΉΡΘ–ΆΤΤΜΒ»ΥΒΡΙ§Ψ±Α©œΗΑϊHeLaΒΡœΗΑϊΡΛ[28]ΓΘ
≥ΐΝΥ…œ ω3÷÷Μζ÷ΤΆβΘ§ΜΙ”–“Μ÷÷ΤΤΡΛΜζ÷ΤΫ–ΉωShai-Huang-Matsuzaki (SHM)ΡΘ–ΆΘ§±Μ»œΈΣ «’±ΧΚΡΘ–ΆΚΆΜΖ–ΈΩΉΡΘ–ΆΒΡΫαΚœ[29]ΓΘ’β–©ΡΘ–ΆΥδ»ΜΕΦ «ΩΙΑ©κΡ”κΑ©œΗΑϊΡΛΒΡœύΜΞΉς”ΟΒΦ÷¬œΗΑϊΡΛΝ―ΫβΘ§ΒΪ «ΤδΡΎ‘ΎΒΡΖ÷Ή”Μζ÷Τ”–Υυ≤ΜΆ§ΓΘΒΪ «Θ§¥σΝΩ±®Βά÷ΛΟςΘ§–μΕύΩΙΑ©κΡΩ…“‘Ά®Ιΐ≤ΜΆ§ΒΡΉς”ΟΖΫ Ϋ”ΠΕ‘≤ΜΆ§ΒΡΑ©œΗΑϊΘ§“‘MagaininάύΩΙΑ©κΡΈΣάΐΘ§ΤδΩΙΑ©Ής”ΟΖΫ ΫΦ»”–ΜΖ–ΈΩΉ”÷”–’±ΧΚΡΘ–ΆΘ§…θ÷ΝΜΙ”–Ζ«ΡΛΝ―ΫβΜζ÷Τ[24, 28, 30]ΓΘ
2.2 ΩΙΑ©κΡΒΡΖ«ΡΛΝ―ΫβΜζ÷Τ
ΩΙΑ©κΡΒΡΉς”ΟΜζ÷Τ≥ΐΝΥΗΡ±δΑ©œΗΑϊΡΛΆ®ΆΗ–‘“‘ΆβΘ§ΜΙΩ…“‘”κΑ©œΗΑϊΒΡΡΎ‘¥Α–±ξœύΜΞΉς”ΟΘ§ΫχΕχ”’ΒΦΑ©œΗΑϊΒΡΥάΆω(ΆΦ 2)ΓΘ
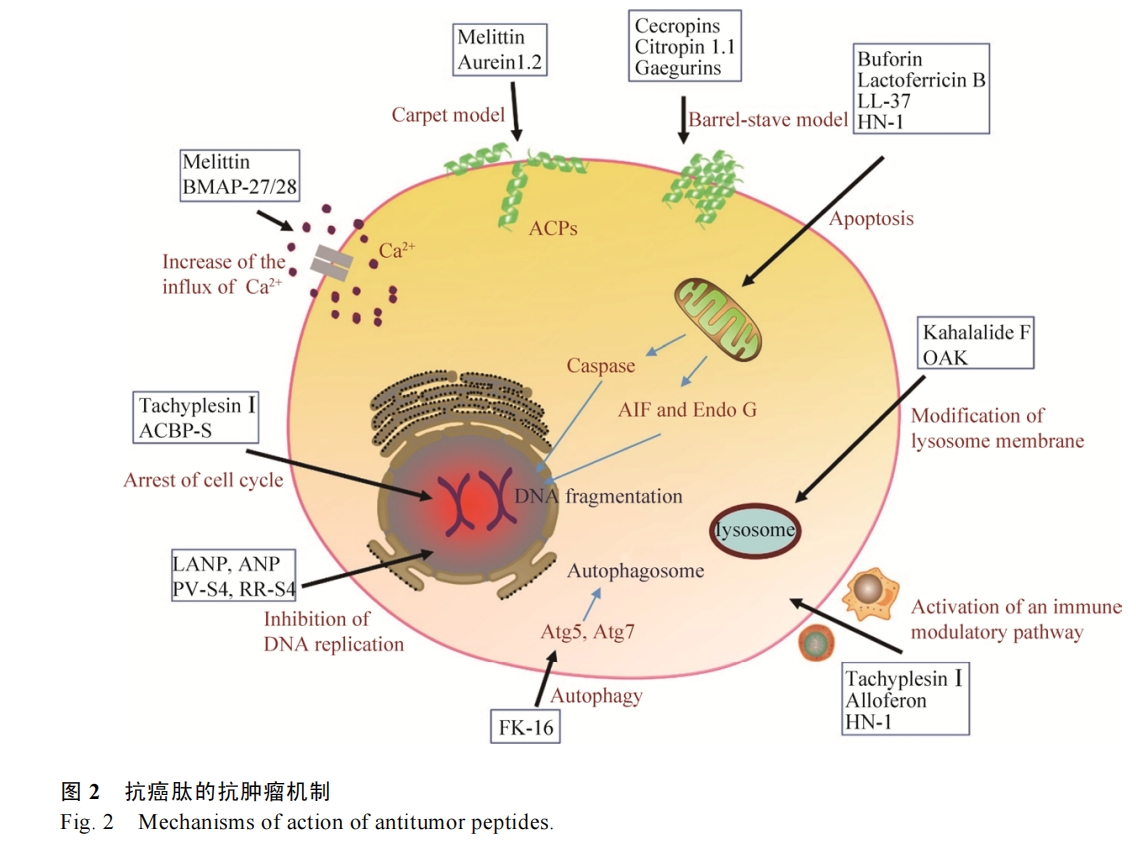
2.2.1 ”’ΒΦΒρΆωΆΨΨΕ
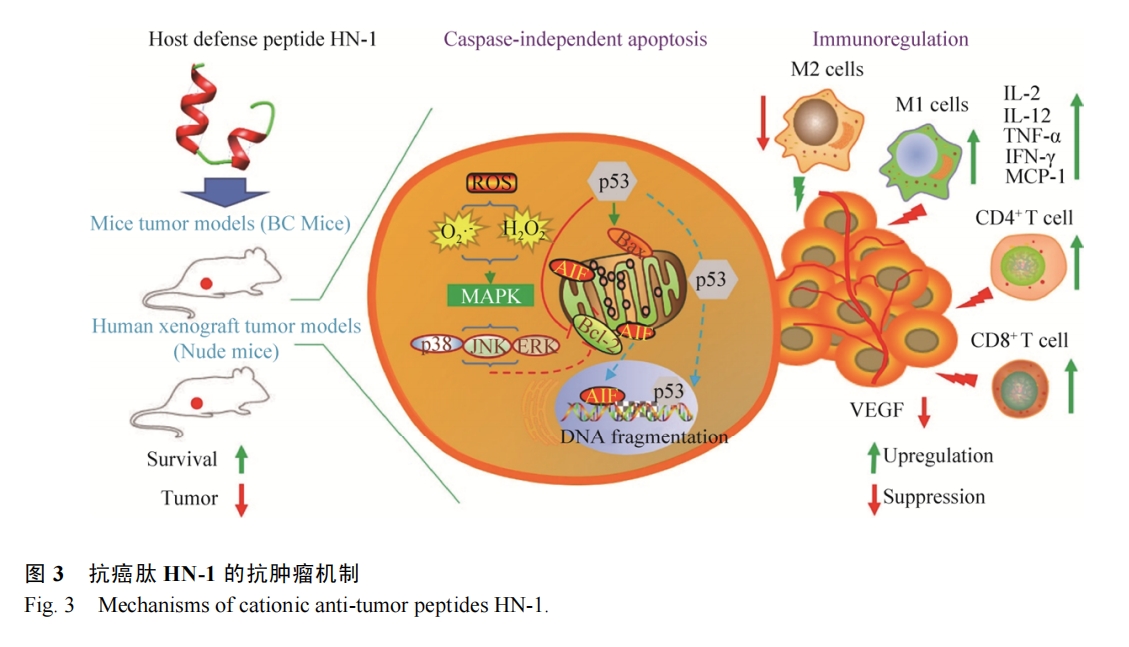
¥σΝΩΒΡ―–ΨΩ÷Λ ΒΩΙΑ©κΡΩ…Ά®ΙΐΦΛΜνΒρΆωΆ®¬Ζά¥÷¥––ΙΠΡήΘ§ΒρΆωœΗΑϊ±μœ÷≥ω“ΜœΒΝ––ΈΧ§ΚΆ…ζΜ·ΧΊ’ςΒΡ±δΜ·Θ§»γΑϊ÷ ÷εΥθΓΔΝΉ÷§θΘΥΩΑ±ΥαΆβΖ≠ΓΔ»Ψ…Ϊ÷ ΡΐΨέΓΔDNAΤ§ΕΈΜ·ΓΔΚΥΡΛΚΥ» ΤΤΥιΒ»[31]ΓΘ“ΜΑψά¥ΥΒΒρΆωΖ÷ΈΣΡΎ‘¥–‘ΒρΆωΦ¥œΏΝΘΧεΆΨΨΕΚΆΆβ‘¥–‘ΒρΆωΦ¥ΥάΆω ήΧεΆΨΨΕΘ§Τδ÷–ΑκκΉΧλΕ§ΟΗ(Caspase)-9ΚΆ-8Ζ÷±π «Εΰ’Ώ±ξ÷Ψ–‘÷–ΦδΦΛΜνΈοΘ§caspase-3Θ§6Θ§7 «Εΰ’ΏΙ≤Ά§ΒΡΒρΆω÷¥––’Ώ[32]ΓΘLeeΒ»ΖΔœ÷ά¥Ή‘÷–ΜΣ¥σσΗρήBufo bufo gargarizansΒΡΩΙΑ©κΡBuforinΔρbΕ‘Εύ¥ο62÷ξΑ©œΗΑϊΕΦ”–«Ω…±…ΥΉς”ΟΘ§ΤδΩ…ΤΤΜΒœΏΝΘΧεΡΛΘ§ ΆΖ≈œΗΑϊ…ΪΥΊCΘ§ΫχΕχΦΛΜνcaspaseΦΕΝΣΘ§”’ΒΦ“ΜœΒΝ–ΒΑΑΉΥ°ΫβΖ¥”ΠΒΦ÷¬œΗΑϊΆΏΫβ(ΡΎ‘¥–‘ΒρΆω)[33]ΓΘChenΒ»ΫΪtachyplesinΔώΒΡCΕΥΝ§Ϋ”“ΜΕΈ¥χ”–RGDΒΡΕΧκΡΘ§ΖΔœ÷ΜώΒΟΒΡRGD-tachyplesinΔώ‘ΎΧεΡΎΆβΨυΩ…“÷÷Τ÷ΉΝω…ζ≥ΛΘ§≤Δ«“ΦΛΜνcaspase-3,6,7,8,9“‘ΦΑfas≈δΧεœύΙΊΥάΆω”ρ±μ¥οΘ§“≤ΨΆ «ΥΒΤδΩ…Ά§ ±ΦΛΜνœΏΝΘΧεΆΨΨΕΚΆΥάΆω ήΧεΆΨΨΕ[34]ΓΘ≤ΜΫω»γ¥ΥΘ§ΫϋΡξά¥œύΦΧ≥ωœ÷ΚήΕύ±®Βά÷ΛΟςΩΙΑ©κΡΜΙΡήΙΜΦΛΜν≤Μ“άάΒ”ΎcaspaseΒΡΒρΆωΆ®¬ΖΓΘ’β÷÷ΒρΆωΆ®¬Ζ≤Μ–η“ΣcaspaseΒΡΦΛΜνΘ§Εχ «¥Ό Ι¥φ‘Ύ”ΎœΏΝΘΧεΡΎΆβΡΛΦδœΕΒΡœΗΑϊΒρΆω”’ΒΦ“ρΉ”(AIF)ΚΆΚΥΥαΡΎ«–ΟΗG (EndoG)ΚΥΉΣ“ΤΘ§ΫχΕχ“ΐΤπDNAΝ―ΫβΚΆ»Ψ…Ϊ÷ ΡΐΦ·Β»[35]ΓΘRenΒ»ΖΔœ÷LL-37 «Ά®ΙΐΦΛΜνcaspaseΖ«“άάΒ–‘ΒΡΒρΆωΆ®¬Ζ“÷÷Τ»ΥΫα≥ΠΑ©œΗΑϊΒΡ…ζ≥ΛΓΘLL-37”––ßΦΛΜνœΗΑϊ÷–ΒΡ“÷Α©Μυ“ρp53ΒΡ±μ¥οΘ§ΫχΕχ”’ΒΦΕύ÷÷ΉΣ¬ΦΑ–±ξΒΡ±μ¥οΑϋά®Bcl-2Φ“ΉεΒΡ¥ΌΒρΆωΒΑΑΉ»γBAXΓΔBakΚΆPumaΘ§’β–©“ρΉ”Ρή¥ΌΫχœΏΝΘΧε…ζάμΜζΡήΒΡΗΡ±δΘ§ΫχΕχ ΆΖ≈AIFΚΆEndoGΫχ»κœΗΑϊΚΥ÷¥––ΒρΆωΙΠΡή[36]ΓΘ±Ψ Β―ι “«ΑΤΎΙΛΉς¥”ΚΘΡœΆΡΆήAmolops hainanensis÷–Χα»ΓΒΫΝΥ“Μ÷÷ΩΙΨζκΡHN-1ΨΏ”–ΙψΤΉΒΡΩΙΨζΜν–‘[37]Θ§Κσ–χ Β―ιΖΔœ÷ΤδΕ‘Α©œΗΑϊΨΏ”–―Γ‘ώ–‘…±…ΥΉς”ΟΘ§«“”––ßΦΛΜνΝΥcaspaseΖ«“άάΒ–‘ΒΡΒρΆωΆ®¬Ζ“÷÷Τ»Υ»ιœΌΑ©œΗΑϊMCF-7ΒΡ…ζ≥Λ(ΆΦ 3)ΓΘ
2.2.2 Ήη÷ΙœΗΑϊ÷ήΤΎ”ΎG0ΓΔG1ΜρSΤΎ
œΗΑϊ÷ήΤΎ «œΗΑϊ…ζΟϋΜνΕ·ΒΡΜυ±ΨΙΐ≥ΧΘ§Τδ“άάΒ”ΎΗςΦΕΒςΩΊ“ρΉ”ΒΡΨΪ»ΖΒςΩΊΓΘ¥σΝΩ±®Βά÷Η≥ωΩΙΑ©κΡΩ…Ήη÷ΆΑ©œΗΑϊ”Ύ≤ΜΆ§ ±ΤΎΘ§¥”Εχ“÷÷ΤΑ©œΗΑϊΒΡ‘ω÷≥ΓΘLiΒ»[38]ΖΔœ÷TachyplesinΔώ Ι»ΥΗΈΑ©œΗΑϊSMMC-7721œΗΑϊΉη÷Ά‘ΎG0/G1ΤΎΘ§ Β―ι÷–TachyplesinΔώœ¬ΒςΆΜ±δp53ΓΔœΗΑϊ÷ήΤΎΒΑΑΉD1ΚΆCDK4ΒΡΒΑΑΉΥ°ΤΫΘ§ΫΒΒΆc-MycΒΡmRNAΥ°ΤΫΘ§≤Δ«“¥ΌΫχp16ΚΆp21WAF1/CIP1ΒΡ±μ¥οΘ§Ω…ΦϊTachyplesinΔώΆ®ΙΐΕ‘’β–©œΗΑϊ÷ήΤΎœύΙΊΜυ“ρ±μ¥οΒΡΒςΫΎΘ§ΫχΕχ“÷÷ΤSMMC-7721ΒΡ‘ω÷≥ΓΘZhaoΒ»άϊ”Ο‘≠ΚΥ±μ¥οΜώΒΟΩΙΑ©κΡAGAPΒΡ÷ΊΉιΧεrAGAPΘ§ΖΔœ÷rAGAPΡήΙΜ“÷÷Τ»ΥΫΚ÷ ΝωœΗΑϊSHG-44ΒΡ‘ω÷≥ΚΆ«®“ΤΘ§ΤδΜζάμ «Ά®Ιΐ“÷÷ΤG1œΗΑϊ÷ήΤΎΒςΩΊΒΑΑΉCDK2ΓΔCDK6ΚΆp-RBΒΡ±μ¥οΘ§ ΙSHG-44œΗΑϊ÷ήΤΎ±ΜΉη÷Ά‘ΎG1ΫΉΕΈΘ§ΫχΕχœ‘÷χ“÷÷ΤΤδ‘ω÷≥[39]ΓΘΫΪΚζΧ“κΡWP1”κΡ…ΟΉΈχΫαΚœΡήΙΜΉη÷ΆMCF-7œΗΑϊ”ΎSΤΎΘ§ΫχΕχ“÷÷ΤΤδ‘ω÷≥[40]ΓΘ
2.2.3 ΤΤΜΒ»ήΟΗΧε
Α©œΗΑϊ÷–»ήΟΗΧεΒΡΆ®ΆΗ–‘Ά®≥ΘΜαΖΔ…ζΗΡ±δΘ§≤Δ«“Κœ≥…Ζ÷ΟΎ¥σΝΩΒΡΉι÷·ΒΑΑΉΟΗΘ§ΥϋΟ«”κ÷ΉΝωΒΡ…ζ≥ΛΓΔ«÷œ°ΚΆΉΣ“ΤœΔœΔœύΙΊ[41]ΓΘΨί±®Βά“Μ–©ΩΙΑ©κΡΩ…“‘ΤΤΜΒ»ήΟΗΧεΡΛΘ§ ΆΖ≈»ήΟΗΧεΡΎ»ίΈοΘ§ΒΦ÷¬œΗΑϊΡΎΜΖΨ≥ΥαΜ·Θ§÷±÷ΝΑ©œΗΑϊΥάΆω[42]ΓΘ»γά¥Ή‘“Μ÷÷ΚΘ≤ΈElysia rufescensΒΡΩΙΑ©κΡKahalalide F (KF)Θ§Ω…“‘Ά®ΙΐΤΤΜΒΑ©œΗΑϊ»ήΟΗΧεΫαΙΙά¥…±ΥάΑ©œΗΑϊΘ§Τδ“÷÷ΤΒΡœΗΑϊ÷ξΑϋά®Ϋα≥ΠΑ©ΓΔ»ιœΌΑ©ΓΔΖ«–ΓœΗΑϊΖΈΑ©ΓΔ«ΑΝ–œΌΑ©ΓΔΚΎ…ΪΥΊΝωΒ»[43]ΓΘΥό÷ςΖά”υκΡΡΘΡβΈοOAKΕ‘–Γ σ«ΑΝ–œΌœΌΑ©œΗΑϊTRAMP-C2ΦΑΤδΕ·ΈοΡΘ–ΆΨυ”–ΝΦΚΟΒΡ“÷÷ΤΉς”ΟΘ§≤Δ«“Ω…“‘ΩΥΖΰΕύ“©ΡΆ“©–‘ΓΘHeld-KuznetsovΒ»ΖΔœ÷Θ§OAKΒΡΉς”ΟΜζ÷Τ «Ά®ΙΐΤΤΜΒœΏΝΘΧεΚΆ»ήΟΗΧεΖΔΜ”Ής”Ο[44]ΓΘ
2.2.4 ‘ωΦ”ΗΤάκΉ”ΡΎΝς
Α©œΗΑϊ÷–Ca2+Έ»Χ§ΜαΖΔ…ζΗΡ±δΘ§’β–©ΗΡ±δ”κ÷ΉΝωΒΡΖΔ…ζΓΔ‘ω÷≥ΓΔ¥ζ–ΜΚΆ―ΣΙήΒΡ…ζ≥…”–ΙΊ[45]ΓΘΩΙΑ©κΡΩ…ΗΡ±δœΗΑϊΡΛΆ®ΆΗ–‘ΕχΫχ»κœΗΑϊΘ§≤Δ«“Ω…‘ωΦ”œΗΑϊΡΎCa2+ΡΎΝςΘ§ΥφΚσΆ®ΙΐΨ≤ΒγΈϋΗΫΉς”Ο”ΎœΏΝΘΧεΘ§‘ΎCa2+ΒΡ–≠Ά§œ¬Ής”Ο”ΎœΏΝΘΧεΆ®ΆΗ–‘ΉΣΜΜΩΉ(PTP)Θ§ΒΦ÷¬ΡΎ»ίΈοΆβΝς“ΐΤπΑ©œΗΑϊΥάΆωΓΘΖδΕΨκΡMelittinΩ…“‘Ά®Ιΐ‘ω«ΩCa2+ΒΡΝς»κΚΆΆΑΑεΡΘ–ΆΒ»Μζ÷Τά¥…±Υά»ΥΒΡΗΈΑ©œΗΑϊ[46]ΓΘΝμΆβΘ§RissoΒ»“≤ΖΔœ÷ά¥Ή‘Φ“≈ΘBos taurusΒΡBMAP-27/28Ε‘ΑΉ―Σ≤ΓΓΔΝήΑΆΑ©Β»ΕΦ”–ΚήΚΟΒΡ“÷÷ΤΉς”ΟΘ§ΤδΜζ÷ΤΨΆ «ΗΡ±δœΗΑϊΡΛΒΡΆ®ΆΗ–‘ΚΆΧαΗΏœΗΑϊ÷–Ca2+ΡΎΝςΘ§≤ΔΑιΥφΉ≈DNAΒΡΤ§ΕΈΜ·Θ§ΫχΕχ”’ΒΦΑ©œΗΑϊΒΡΥάΆω[47]ΓΘ
2.2.5 “÷÷ΤDNAΚœ≥…
”’ΒΦΑ–œΗΑϊDNAΤ§ΕΈΜ· «Εύ÷÷ΩΙΑ©κΡΒΡΉς”Ο–ßΙϊΘ§ΒΪ≤Μ“ΜΕ® «ΩΙΑ©κΡ÷±Ϋ”Ής”Ο”ΎDNAΘ§±»»γΒρΆωΆ®¬Ζ“≤Μα“ΐΤπDNAΕœΝ―ΓΘΫϋΡξά¥―–ΨΩΖΔœ÷”––©ΩΙΑ©κΡΩ…“‘÷±Ϋ””κΑ©œΗΑϊ»Ψ…ΪΧεDNAΜρœύΙΊΟΗœύΜΞΉς”ΟΘ§ΫχΕχΗ…»≈Μρ“÷÷ΤΑ©œΗΑϊΒΡDNAΚœ≥…ΓΘGowerΒ»[48]ΖΔœ÷4÷÷άϊΡρΡΤκΡ(LANPΓΔANPΓΔBNPΚΆCNP)Ε‘»ΥΒΡΫα≥ΠΑ©œΗΑϊ”–“÷÷ΤΉς”ΟΘ§ΤδΩΙ÷ΉΝωΜζ÷ΤΨΆ «Ά®Ιΐ“÷÷ΤΜΖΝΉΥαΡώή’(Cyclic GMP)ΫιΒΦΒΡΑ©œΗΑϊDNAΚœ≥…ά¥Ήη÷ΙΑ©œΗΑϊ‘ω÷≥ΒΡΓΘHariton-GazalΒ»[49]Ά®Ιΐ Β―ιΖΔœ÷Ψ≠ΙΐΦ”ΙΛΗΡ‘λΚσΒΡΩΙΑ©κΡPV-S4ΚΆRR-S4Ω…“‘ΫαΚœΒΫHelaœΗΑϊΚΥ»Ψ…ΪΧε…œΘ§ ΙΤδDNA≥ωœ÷ΕœΝ―Θ§ΫχΕχ”’ΒΦ÷ΉΝωœΗΑϊΥάΆωΓΘΝμΆβΘ§Ζ÷άκΉ‘Elysia rufescensΒΡΩΙΑ©κΡKahalalide F (KF)ΓΔά¥Ή‘ΒμΖέΚΥ–Γ«ρ‘εΒΡCPAPΒ»ΨυΡήΆ®ΙΐΉη÷ΙDNAΗ¥÷Τά¥…±Υά÷ΉΝωœΗΑϊ[18]ΓΘ
2.2.6 ¥Ό ΙΑ©œΗΑϊΉ‘ …
œΗΑϊΉ‘ …“≤≥ΤΔρ–Ά≥Χ–ρ–‘œΗΑϊΥάΆωΘ§ «œΗΑϊ÷–ΗΏΕ»±Θ ΊΒΡΉ‘––ΫΒΫβΙΐ≥ΧΘ§Ή‘ … ΙΦΔΕωΜρ»±ΖΠ…ζ≥Λ“ρΉ”ΒΡœΗΑϊΒΟ“‘‘ί ±≥…ΜνΘ§ΕχΡ«–©≥÷–χ≤ΜΡήΜώΒΟ”Σ―χΒΡœΗΑϊΫΪœϊΜ·Υυ”–Ω…ΜώΒΟΒΡΜυ÷ Θ§Ήν÷’ΒΦ÷¬Ή‘ …œύΙΊ–‘œΗΑϊΥάΆωΓΘΫϋΡξά¥ΖΔœ÷Τδ“≤ «ΩΙΑ©κΡΒΡ÷ΉΝω“÷÷ΤΜζ÷Τ÷°“Μ[50-51]ΓΘRenΒ»[52]±®ΒάΝΥLL-37ΒΡΤ§ΕΈFK-16 (ΒΎ17®C32ΗωΑ±ΜυΥα)Ω…”’ΒΦΫα≥ΠΑ©œΗΑϊHCT116ΒρΆωΚΆΉ‘ …Θ§ΒΪ «Ε‘’ΐ≥ΘΫα≥Π…œΤΛœΗΑϊNCM460ΕΨ–‘Κή–ΓΓΘFK-16ΧαΗΏΝΥΫα≥ΠΑ©œΗΑϊ÷–Ή‘ …œύΙΊΒΑΑΉLC3-Δώ/ΔρΓΔAtg5ΚΆAtg7ΒΡ±μ¥οΝΩΘ§Ά§ ±‘ΎΙ≤ΨέΫΙΚΆΒγΉ”œ‘ΈΔΨΒœ¬Ιέ≤λΒΫLC3―τ–‘ΒΡΉ‘ …ΧεΒΡ–Έ≥…ΓΘΒ±«Ο≥ΐΒτAtg5ΚΆAtg7ΚσΘ§¥σ¥σΫΒΒΆΝΥFK-16Ε‘Ϋα≥ΠΑ©œΗΑϊΒΡ…±…ΥΉς”ΟΘ§ΥΒΟς”’ΒΦΑ©œΗΑϊΉ‘ … «FK-16ΖΔΜ”Ής”ΟΒΡ÷Ί“ΣΜζ÷ΤΓΘ
2.2.7 ΦΛΜν÷ΉΝωΟβ“Ώ
Οβ“ΏœΒΆ≥‘ΎΜζΧεΩΊ÷ΤΚΆ«ε≥ΐ÷ΉΝωΖΫΟφΤπΒΫΝΥ÷ΝΙΊ÷Ί“ΣΒΡΉς”ΟΘ§ΒΪ»‘Ρ―“‘Ήη÷Ι÷ΉΝωΒΡΖΔ…ζΚΆΖΔ’ΙΓΘ’β”κΗ¥‘”ΒΡ÷ΉΝωΈΔΜΖΨ≥Οή≤ΜΩ…Ζ÷Θ§“ΜΖΫΟφΘ§÷ΉΝωœΗΑϊΩ…Ζ÷ΟΎ¥ΌΫχ÷ΉΝω…ζ≥ΛΓΔΉΣ“ΤΒΡœΗΑϊ“ρΉ”Θ§»γΉΣΜ·…ζ≥Λ“ρΉ”-Π¬ΓΔ―ΣΙήΡΎΤΛ…ζ≥ΛΒ»[53]ΘΜΝμ“ΜΖΫΟφΘ§Εώ–‘÷ΉΝωΩ…“‘Ά®ΙΐΕύ÷ΊΜζ÷ΤΟβ“Ώ”Π¥πΘ§¥”ΕχΧ”“ίΟβ“ΏœΒΆ≥ΒΡΙΞΜςΉς”ΟΘ§»γ Ι÷ΉΝωΫΰ»σΒΡCD8+ CTLsΚΆCD4+ Th1œΗΑϊ¥Π”Ύ“Μ÷÷ΙΠΡήΚΡΫΏΜρΈόΡήΉ¥Χ§Θ§ΈόΖ®Ε‘÷ΉΝωΫχ––Οβ“ΏΦύ ”ΚΆ«ε≥ΐ[54]ΓΘ“ρ¥Υ”’ΒΦΟβ“ΏΜνΜ·ΓΔ¥ρΤΤΟβ“ΏΡΆ ήΒ»“―≥…ΈΣ»»Ο≈ΒΡΟβ“Ώ÷ΈΝΤΖΫœρΘ§»γΟβ“ΏΦλ≤ιΒψ“÷÷ΤΦΝ[55]ΓΘ”–―–ΨΩ÷ΛΟς”––©ΩΙΑ©κΡΩ…“‘ΒςΫΎΜζΧεΒΡΟβ“Ώ”Π¥πΖΔΜ”ΤδΩΙ÷ΉΝωΉς”ΟΓΘChenΒ»[56]±®ΒάTachyplesinΔώΡήΙΜ¥Ό ΙœΗΑϊ±μΟφΒΡΆΗΟς÷ ΆιΚΆ―Σ«ε÷–≤ΙΧεΆΨΨΕΒΡΙΊΦϋ≥…Ζ÷C1q≤ΙΧεœύΜΞΉς”ΟΘ§≤Δ«“ΦΛΜνΤδœ¬”ΈΒΡC3ΚΆC4ΒΡΝ―ΫβΚΆ≥ΝΜΐΘ§“‘ΦΑC5b-9ΒΡ–Έ≥…Θ§ΦΛΜνΒδ–Ά≤ΙΧεΆΨΨΕ¥”ΕχΤΤΜΒΑ©œΗΑϊΒΡΆξ’ϊ–‘ΓΘChernyshΒ»±®ΒάΝΥά¥Ή‘ΚλΆΖάω”§Calliphora vicinaΒΡΩΙΨζκΡalloferonΩ…Ά®ΙΐΦΛΜνΟβ“Ώ”Π¥π“÷÷Τ÷ΉΝω…ζ≥ΛΓΘΆ®ΙΐΧεΆβΫΪ–Γ σΝήΑΆœΗΑϊΜρ»ΥΒΡ―Σ“ΚΒΞΚΥœΗΑϊΫχ–– ‘―ιΘ§ΖΔœ÷alloferonΩ…ΦΛΜνΉ‘»Μ…±…Υ(NK)œΗΑϊΚΆΗ…»≈ΥΊ(IFN)ΒΡ±μ¥ο[57]ΓΘHuangΒ»[58]ΖΔœ÷¥”±»ΡΩ”ψ±ΣςΘPardachirus marmoratusΖ÷άκΒΡΩΙΑ©κΡGE33Ω…“‘ΉςΈΣ“ΏΟγΉτΦΝΧα…ΐΟπΜνΑρκΉΑ©œΗΑϊ(MBT-2)ΒΡΟβ“Ώ‘≠–‘Θ§‘Ύ–Γ σΧεΡΎœ‘÷χΧαΗΏΝΥCTLœΗΑϊΚΆNKœΗΑϊ ΐΝΩ“‘ΦΑΧΊ“λ–‘ΩΙΧεΥ°ΤΫΒ»Θ§÷ΛΟςΝΥΩΙΑ©κΡΒΡΟβ“ΏΒςΫΎ«±ΡήΓΘ±ΨΩΈΧβΉιΖΔœ÷ΒΡΩΙΑ©κΡHN-1‘ΎΕ·ΈοΧεΡΎΦΛΜνΝΥCD4+ TœΗΑϊΚΆΨό …œΗΑϊ‘Ύ÷ΉΝω÷–ΒΡΫΰ»σΘ§≤Δ«“ΧαΗΏΝΥ÷ΉΝωœύΙΊœΗΑϊ“ρΉ”‘Ύ―Σ«ε÷–ΒΡΥ°ΤΫ(ΆΦ 3)ΓΘ
2.2.8 “÷÷Τ÷ΉΝω―ΣΙή–¬…ζ
–¬…ζ―ΣΙή–Έ≥…”κ÷ΉΝω«÷œ°ΚΆΉΣ“ΤœΔœΔœύΙΊΘ§ΤδΈΣ÷ΉΝωΉι÷·ΧαΙ©―θΤχΚΆ”Σ―χΘ§¥ΌΫχ÷ΉΝωœΗΑϊ―ΗΥΌ‘ω÷≥Θ§Ά§ ±ΈΣ÷ΉΝωΒΡ‘ΕΕΥΉΣ“ΤΧαΙ©ΉΣ‘Υ[59]ΓΘ“ρ¥ΥΑ–œρ÷ΉΝω–¬…ζ―ΣΙή…ζ≥…ΜρœύΙΊ“ρΉ”ΒΡΩΙΑ©άύ“©Έο―–ΖΔΨΏ”–÷Ί¥σ“β“εΓΘMaderΒ»[60]Ά®Ιΐ Β―ιΖΔœ÷Lactoferricin B‘ΎΧεΆβΆ®ΙΐΉη÷ΙœΗΑϊ…ζ≥Λ“ρΉ”(bFGF)ΚΆ―ΣΙήΡΎΤΛ…ζ≥Λ“ρΉ”(VEGF165)”κ ήΧεΫαΚœΕχ“÷÷Τ»ΥΤξΨ≤¬ωΡΎΤΛœΗΑϊ(HUVECs)‘ω÷≥Θ§≤Δ«“‘ΎC57BL/6–Γ σΧεΡΎ“÷÷ΤΕΰ’Ώ”’ΒΦΒΡ―ΣΙή…ζ≥…ΓΘHouΒ»[61]ΫΪ»ΥΙΛ…ηΦΤΩΙΨζκΡ”κΑ–œρΗχ“©–ρΝ–isoDGRœύΝ§Ϋ”Θ§ΖΔœ÷ΤδΡήΙΜ”κΠΝvΠ¬3+ (÷ΉΝωœΗΑϊΙΐ±μ¥ο)ΫαΚœ¥”Εχ“÷÷Τ―ΣΙήΒΡ…ζ≥…Θ§≥δΖ÷÷ΛΟςΝΥΩΙΑ©κΡΨΏ”–“÷÷Τ―ΣΙή–¬…ζΒΡ«±ΡήΓΘ
3 ΉήΫα”κ’ΙΆϊ
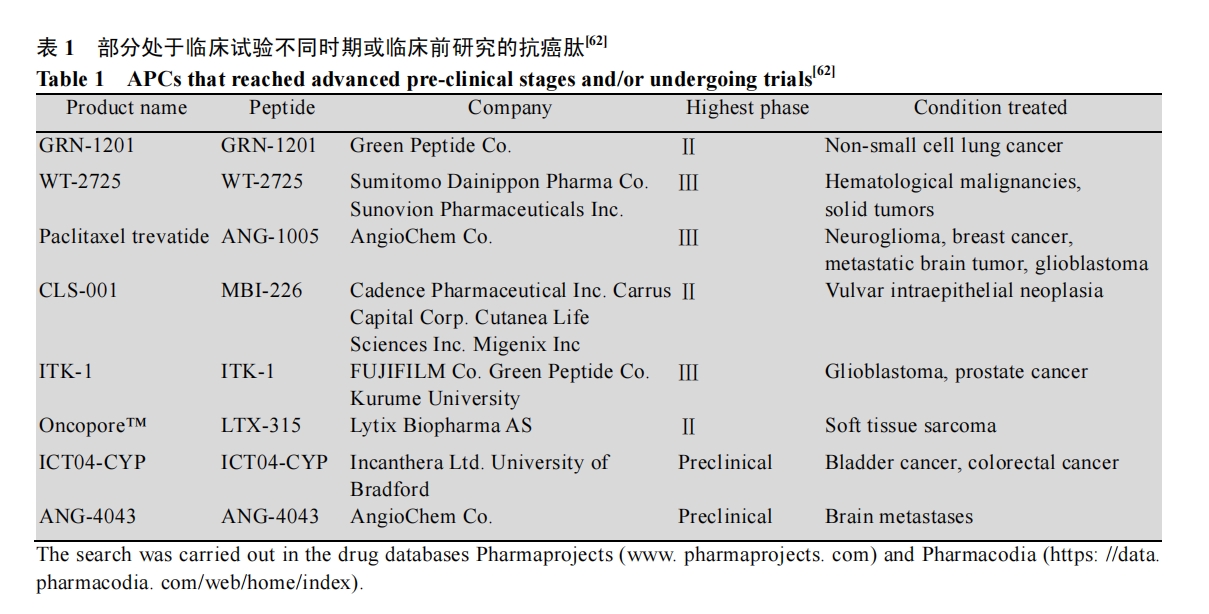
‘ΎΙΐ»ΞΒΡ20Ρξ÷–Θ§Χλ»ΜΜν–‘Ζ÷Ή”ΒΡΕύ÷÷÷ΈΝΤ«±ΡήΒΡ≥÷–χΖΔœ÷“ΐΖΔΝΥΩΤ―ßΫγΒΡΙψΖΚΉΔ“βΓΘΩΙΑ©κΡ”…”ΎΤδΧΊ βΒΡ―τάκΉ”ΚΆΝΫ«Ή–‘ΒΡΫαΙΙΧΊ’ς“‘ΦΑΤδ÷ΎΕύΒΡΩΙΑ©Μζ÷ΤΘ§ ΙΤδΖΔΜ”ΝΥΝΦΚΟΒΡΩΙΑ©Ής”ΟΜρ’Ώ‘ω«ΩΜ·ΝΤ“©ΈοΒΡ–ßΙϊΘ§“ρΕχ”–Άϊ”Ο”ΎΧαΗΏΜ·ΝΤ“©ΈοΒΡΟτΗ––‘Θ§Ά§ ±Φθ…ΌΕ‘’ΐ≥ΘΉι÷·ΒΡΕΨΗ±Ής”ΟΓΘΡΩ«Α“―”–¥σΝΩ≤ΜΆ§÷ΈΝΤΡΩΒΡκΡάύ“©ΈοΫχ»κΝΌ¥≤Μρ≈ζΉΦ…œ –Θ§»γ±μ 1Υυ ΨΘ§Β±«Α“―”–≤ΩΖ÷¥Π”ΎΝΌ¥≤ ‘―ιΫΉΕΈΒΡΩΙΑ©κΡΘ§≤Δ»ΓΒΟΝΥ“ΜΕ®ΒΡ–ßΙϊ[62]ΓΘΒΪ «ΩΙΑ©κΡΒΡ”Π”ΟΚΆ―–ΖΔ»‘ΟφΝΌ“Μ–©Χτ’ΫΘ§»γΚœ≥…≥…±ΨΫœΗΏΓΔ“Ή±ΜΒΑΑΉΟΗΥ°ΫβΓΔ“ΉΨέΚœΓΔΑκΥΞΤΎΫœΕΧΒ»[63]ΓΘ“ρΕχΒ±«ΑΩΙΑ©κΡΒΡ…ηΦΤ“≤Φ·÷–”ΎΫΊΕΧ–ρΝ–Β»Θ§“‘ΫΒΒΆ≥…±Ψ[64]ΘΜΫβΨωΒΑΑΉΟΗΒΡΥ°ΫβΈ ΧβΘ§Ω…“‘Ά®ΙΐΫΪΧλ»ΜΑ±ΜυΥαΧφΜΜ≥…Ζ«Χλ»ΜΑ±ΜυΥαΘ§±»»γ…ηΦΤD-Ε‘”≥ΧεκΡΓΔΠ¬2,2Α±ΜυΥαΧφΜΜΓΔκΡΙ«Η…ΜΖΜ·ΓΔend-capping»γc-θΘΑΖΜ·ΓΔΧ«άύcoatingΒ»[65-67]ΘΜΧαΗΏΑκΥΞΤΎΩ…“‘ΫΪΩΙΑ©κΡΨέ““Εΰ¥Φ–ό Έ(PEGylation)Θ§ΫαΚœΒΫ―Σ«εΑΉΒΑΑΉΜρΩΙΧεΤ§ΕΈΒ»[68]ΓΘ”…¥ΥΩ…ΦϊΥδ»ΜΩΙΑ©κΡ‘Ύ÷ΉΝω÷ΈΝΤΖΫΟφ”–ΚήΚΟΝΌ¥≤”Π”ΟΦέ÷ΒΘ§ΒΪ «»‘–ηΩΥΖΰ’β–©»±œίΚΆΧτ’ΫΘ§≤≈Ρή‘Ύ÷ΉΝω“©Έο÷ΈΝΤΝλ”ρ”–“Μœ·÷°ΒΊΓΘ“ρ¥ΥΘ§Ϋχ“Μ≤Ϋ»ΖΕ®ΚΆΖΔœ÷ΗϋΕύΩΙΑ©κΡΡΘΑεΚΆΩΙ÷ΉΝωΜζ÷Τ“‘ΦΑΩΥΖΰκΡάύ“©Έο»±ΒψΒΡ–¬ΖΫΖ®Ε‘ΩΙ÷ΉΝωΝΌ¥≤÷ΈΝΤ“©ΦΝΒΡΖΔ’ΙΨΏ”–÷Ί¥σ“β“εΓΘ
Οβ‘π…υΟςΘΚ±ΨΈΡΈΣ––“ΒΫΜΝς―ßœΑΘ§ΈΡ’¬±ξΉΔΝΥ‘≠Ής’ΏΦΑ≥ω¥ΠΘ§Αφ»®Ιι‘≠Ής’ΏΥυ”–Θ§»γ”–«÷»®Θ§Ω…ΝΣœΒ…Ψ≥ΐΓΘ
