
ЃлеЊ вЊЃн ПЙОњвЉЮяРФгУЕМжТЯИОњФЭвЉаЮЪЦВЛЖЯМгОчЃЌЯИОњФЭвЉадвбГЩЮЊШевцбЯжиЕФЙЋЙВЮРЩњЮЪЬтЁЃБОЮФНщЩмВЛЭЌПЙОњвЉЮяШчКЮЗЂЛгПЙОњзїгУЃЌвдМАЯИОњЭЈЙ§ИФБфЯИАћФЄЭЈЭИадЕШЗНЪНВњЩњФЭвЉадЕФЛњжЦЁЃЮЊНтОіЯИОњФЭвЉадЮЪЬтЃЌбЇепУЧЬсГіСЫПЙОњвЉЮяСЊКЯгІгУЕФаЭЌПЙОњВпТдЃЌвдЦкЭЈЙ§ЖрАаЕуЁЂЖрЭООЖЕФзїгУЗНЪНЃЌНЕЕЭЯИОњВњЩњФЭвЉЕФИХТЪЃЌЭЌЪБЬсИпПЙОњаЇСІЁЃБОЮФДгаЭЌПЙОњЛњжЦГіЗЂЃЌЖдВЛЭЌПЙОњвЉЮяСЊКЯгІгУЕФаЭЌПЙОњзїгУНјаазлЪіЃЌВЂжиЕуНщЩмПЙОњыФгыДЋЭГПЙОњвЉЮяСЊКЯгІгУЕФаЭЌаЇЙћЃЌвдМАдкВЛЭЌСьгђгІгУЕФбаОПНјеЙЃЌжМдкНјвЛВНЭЦЖЏПЙОњыФЕШаТаЭПЙОњвЉЮягІгУВпТдДДаТЃЌвжжЦЯИОњФЭвЉадЕФЗЂеЙЁЃ
ПЙОњвЉЮяЪЧвЛРрФмЙЛвжжЦЯИОњЁЂецОњЕШВЁдЮЂЩњЮяЩњГЄЗБжГЛђжБНгЩБУ№ВЁдЮЂЩњЮяЕФЛЏбЇЮяжЪЃЌШчПЙЩњЫиЁЂПЙОњыФЕШЁЃзд1928ФъгЂЙњПЦбЇМвИЅРГУїЗЂЯжЕквЛИіПЙЩњЫиЁЊЧрУЙЫивдРДЃЌПЙЩњЫиБЛЙуЗКгІгУгкМВВЁЕФЗРжЮжаЃЌЭьОШСЫЮоЪ§ЕФЩњУќЁЃЦ№ГѕЃЌПЙЩњЫиЪЧвЛжжБЛгУзїжЮСЦЯИОњИаШОЕФвЉЮяЃЌШЛЖјНќФъРДПЙОњвЉЮяЕФГЌБъЪЙгУЁЂЮоНкжЦРФгУЃЌЕМжТЯИОњФЭвЉадЮЪЬтШевцбЯжиЁЃММЪѕЕќДњЛКТ§жТЪЙаТвЉбаЗЂРЇФбЃЌЯИОњВњЩњФЭвЉЕФЫйЖШдЖГЌЙ§аТПЙОњвЉЮябаЗЂЫйЖШЃЌПЙОњвЉ Юя гШ Цф ЪЧ ПЙ Щњ Ыи Сь гђ ЕФ зЈ Рћ Щъ Чы Ъ§ СП дк2011ФъКѓГЪЯжЯТНЕЕФЧїЪЦЃЌЪРНчЗЖЮЇФкМДНЋУцСйжижЂЯИОњИаШОЮовЉПЩгУЕФОГЕиЁЃвђДЫЃЌКЯРэЪЙгУЯжгаПЙОњвЉЮяЃЌЬсЩ§ПЙОњвЉЮяЕФРћгУаЇТЪЃЌвжжЦЯИОњФЭвЉадЕФЗЂеЙЦШдкУМНоЁЃБОЮФОЭВЛЭЌПЙОњвЉЮяЕФПЙОњЛњжЦЃЌвдМАЯИОњВњЩњФЭвЉадЕФЬиЕуЃЌжиЕуНщЩмМЋОпПЙЩњЫиЬцДњЧБжЪЕФAMPsгыДЋЭГПЙОњвЉЮяСЊКЯПЙОњЕФВпТдЁЃИУВпТдзїЮЊвЛжжаТЕФжЮСЦФЭвЉЯИОњИаШОКЭМѕЩйЯИОњФЭвЉадВњЩњЕФгааЇЗНЗЈЃЌЗЂЛгаЭЌдіаЇЕФПЙОњзїгУЃЌНЕЕЭЕЅИіПЙОњвЉЮяЕФгУСПЃЌДДаТAMPsгІгУВпТдЃЌЮЊНтОіДЋЭГПЙОњвЉЮяФЭвЉадЮЪЬтЬсЙЉаТЗНЯђЁЃ
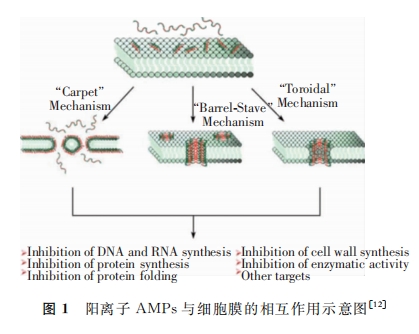
ЃБ ПЙОњвЉЮяЕФПЙОњЛњжЦ
ПЙОњвЉЮяЕФПЙОњЛњжЦжївЊгавдЯТЃДжжЃКвжжЦЯИОњЯИАћБкЕФаЮГЩЃЌЦЦЛЕЯИАћ ФЄНсЙЙЃЌИЩШХЯИОњФкЃФЃЮЃС ЕФИДжЦЁЂзЊТММАЗвыЃЌЦЦЛЕЕААзжЪКЯГЩЁЃжЕЕУзЂвтЕФЪЧЃЌПЙОњвЉЮяВЛНіПЩвдЭЈЙ§ЦфжаФГвЛжжЛњжЦЗЂЛгПЙОњзїгУЃЌвВПЩвдЭЈЙ§ЖржжЛњжЦЯрНсКЯЕФЗНЪНЗЂЛгПЙОњаЇСІЁЃ
ИљОнЯИОњжжРрВЛЭЌЯИОњЕФЯИАћБкГЩЗжжївЊАќРЈыФОлЬЧЁЂжЌЖрЬЧМАСзБкЫсЕШЃЌЪЧаэЖрПЙОњвЉЮязїгУЕФАаЕуЁЃПЙОњвЉЮяФмЙЛЕМжТЯИАћБкГЩЗжЩњЮяКЯГЩЛњжЦЗЂЩњЙЪеЯЃЌГіЯжКЯГЩКЭНЕНтЕФЮоаЇбЛЗЃЌДгЖјгеЕМЯИАћЕїЭіЁЃЯИАћБкФкВрЕФЯИАћФЄЪЧБЃЛЄЯИАћФкЛЗОГЕФживЊЦСеЯЁЃШЫУЧЦеБщШЯЮЊЃЌAMPsжївЊЭЈЙ§ЦЦЛЕЯИАћФЄНсЙЙЃЌЪЙЯИАћФЄЩЯаЮГЩПзЕРЃЌжТЪЙЯИАћШмНтЃЌДгЖјЩБУ№ВЁдЮЂЩњЮяЁЃФПЧАвбЗЂЯжЕФAMPsЦЦФЄЛњжЦжївЊАќРЈЁАЭААхЁБЁАЬКЪНЁБЁАЛЗПзЁБЕШФЃЪНЃЌШчбєРызгAMPsЪз ЯШ гы ГЪ ИК Еч ад ЕФ ЯИ Аћ ФЄ Нс КЯЃЌЫц КѓAMPsЭЈЙ§ВЛЭЌФЃЪНЭИЛЏЯИОњЯИАћФЄЃЌгАЯьЯИОњЯИАћФкЕФЩњУќЛюЖЏЃЌМћЭМЃБЁЃ
дкDNAИД жЦ Й§ ГЬ жаЃЌDNAЭи ЦЫ вь ЙЙ УИ Дп ЛЏDNAСДЕФЖЯСбгыНсКЯЃЌгАЯьDNAСДЕФЭиЦЫзДЬЌЁЃа§зЊУИКЭЭиЦЫвьЙЙУИЃЩЃжзїЮЊDNAИДжЦЙ§ГЬжазюживЊЕФСНИіЭиЦЫвьЙЙУИЃЌЪЧПЙОњвЉЮяГЃМћАаЕуЁЃФПЧАDNAОлКЯУИвВЪЧбаОПНЯЮЊЙуЗКЕФвЉЮяЧБдкАаЕуЁЃДЫЭтЃЌПЙОњвЉЮяПЩвдЭЈЙ§гыЃвЃЮЃС ОлКЯУИНсКЯЃЌИЩШХЯИОњDNAзЊ ТМ МА ЃвЃЮЃС бг ЩьЃЌЦ№ЕНЩБОњзїгУЁЃВПЗжПЙОњвЉЮяЛЙФмЙЛЭЈЙ§ИЩШХЕААзжЪКЯГЩЦ№ЪМНзЖЮЁЂыФСДЕФ бгЩьЁЂАБЛљЫсДњаЛЕШЗЂЛгПЙ ОњЙІФмЁЃФГаЉИЛКЌИЌАБЫсЕФ ЃСЃЭЃаЃѓПЩвдБЛЯИОњЕФЬиЪтзЊдЫдиЬхОжї ЖЏ дЫЪфЕНЯИ АћФкЬиЖЈАаЕуЃЌгы70SКЫЬЧЬхЯрЛЅзїгУНјЖјЦЦЛЕЕААзжЪКЯГЩЃЌДяЕНЩБУ№ЯИОњЕФФПЕФЁЃ
ЃВ ЯИОњФЭвЉадЕФВњЩњ
ФЭвЉЕФЗЂЩњЪЧЯИОњдкГжајПЙОњвЉЮязїгУЯТЃЌИаЪмздЩэЫљДІЕФВЛСМЛЗОГЬѕМўЖјВњЩњЕФЗРгљЛњжЦЁЃЯИОњЭЈЙ§ЯИАћФЄЭЈЭИадИФБфЃЌжїЖЏЭтХХБУЯЕЭГЙ§БэДяЃЌзїгУАаЕуНсЙЙИФБфЃЌПЙОњвЉЮяЕФЦЦЛЕгыаоЪЮЃЌвдМАаЏДјЭтдДадФЭвЉЛљвђЕШЗНЪНЖдПЙОњвЉЮяВњЩњФЭвЉадЃЌШчЭМЃВЫљЪО
ЯИАћФЄЕФЭЈЭИадвдМАПЙОњвЉЮяНјШыЯИАћЕФФмСІЪЧОіЖЈвЉЮяСЦаЇЕФЙиМќадвђЫиЃЌвЛаЉЯИОњЯИАћФЄЭЈЕРЛђЯИАћБкПзЕРЕФИФБфЃЌЪЙЕУПЙОњвЉЮяВЛвзНјШыОњЬхЃЌНјЖјВњЩњФЭвЉадЁЃвЉЮяЭтХХБУдкЯИОњжаЦеБщДцдкЃЌВЛНіФмМѕЩйПЙОњвЉЮядкЯИОњЯИАћФкЕФЛ§РлЃЌЛЙЮЊЯИОњЬсЙЉСЫзуЙЛЕФЪБМфЪЪгІВЛЭЌвЉЮяЃЌНјШыОњЬхЕФвЉЮядкЭтХХБУзїгУЯТбИЫйХХГіЃЌОњЬхЯИАћФквЉЮяХЈЖШНЕЕЭЃЌФЭвЉадЫцжЎВњЩњЁЃвЉЮяАаЕуНсЙЙЕФИФБфдђЛсНЕЕЭвЉЮягыАаЕуЕФЧзКЭСІЃЌНјЖјЕМжТПЙОњвЉЮяЛюадМѕШѕЛђЪЇЛюЃЌЪЧЯИОњВњЩњФЭвЉадЕФСэвЛживЊЛњжЦЁЃДЫЭтЃЌПЙОњвЉЮядкЗЂЛгзїгУЕФЙ§ГЬжаЦфБОЩэвВПЩФмБЛНЕНтЛђЗЂЩњНсЙЙИФБфЃЌИУЙ§ГЬЭљЭљгаУИЕФВЮгыЁЃвЛЗНУцЃЌЕААзУИЕФЫЎНтзїгУЕМжТЖдУИУєИаЕФПЙОњвЉЮяЕФЙиМќЗДгІжааФБЛЦЦЛЕЛђЗЂЩњВЛвзФцзЊЕФНсЙЙжиХХЃЛСэвЛЗНУцЃЌПЙОњвЉЮяЕФЙВМлаоЪЮзшАвЉЮягыЦфАаЕуЕФЯрЛЅзїгУЃЌДгЖјЕМжТФЭвЉЕФВњЩњЁЃ
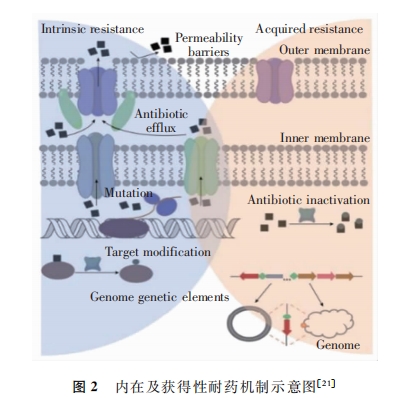
ШчЙћНівРППЩЯЪіФЭвЉЛњжЦЃЌЯИОњЕФЖржиФЭвЉВЛвздкЖЬЪБМфФкБЉЗЂЁЃДЫЭтЃЌЯИОњЛЙЭЈЙ§ХМСЊЁЂзЊЛЏЛђзЊЕМЛёЕУаЏДјФЭвЉЛљвђЕФ ЃФЃЮЃС дЊ МўЃЌвд ЫЎ ЦН Лљ вђ зЊ вЦ ЕФЗНЪНДгЦфЫћЯИОњжаЛёЕУЭтдДадФЭвЉЛљвђЃЌећКЯШыздЩэЛљвђзщЖјВњЩњMDRЁЃ
ЃГ СЊКЯгУвЉаЭЌПЙОњ
гЩгкЯИОњФЭвЉадЕФГжајЗЂеЙЃЌжТЪЙКмЖрбЯжиЕФСйДВЯИОњИаШОЪЙгУЯжгаПЙОњвЉЮяКмФбДяЕНРэЯыЕФжЮСЦаЇЙћЃЌЖјаТвЉбаЗЂЛКТ§ЃЌГЩБОНЯИпЃЌЭЦЙуРЇФбЃЌЧвЭЌбљДцдкв§ЗЂФЭвЉадЕФЗчЯеЁЃвђДЫЃЌЛљгкВЛЭЌПЙОњвЉЮяЖдЯИОњПЙОњЛњжЦВЛЭЌЖјНјааСЊКЯгУвЉЃЌгаЭћГЩЮЊЖдПЙЯИОњФЭвЉадЕФаТВпТдЁЃДЫВпТдФмЙЛНЋЯжгаПЙОњвЉЮяНјаадйРћгУЃЌЪЙПЙОњаЇЙћзюДѓЛЏЃЌБмУтаТвЉбаЗЂЕФОоЖюЗбгУЁЃгыЕЅвЛПЙОњвЉЮяЯрБШЃЌПЙОњвЉЮяСЊгУОпгаЖрАаЕуЕФПЙОњзїгУЗНЪНЃЌФмЙЛгааЇЩБУ№ФЭвЉОњжъЃЌВЂвжжЦЯИОњФЭвЉадЕФЗЂеЙЁЃ
3.1 аЭЌПЙОњЛњжЦ ФПЧАЃЌбЇепУЧжївЊМЏжагквдЯТЫФжжаЭЌПЙОњЛњжЦЕФбаОПЃКдіЧППЙОњвЉЮяЕФЩјЭИадЃЌвжжЦЙВЭЌЕФЩњЛЏЭООЖЃЌвжжЦЯИОњФкФГаЉБЃЛЄадУИЕФВњЩњЃЌвдМАЯИАћБкЛюадМСЕФзщКЯЁЃЦфжаЃЌЖддіЧППЙОњвЉЮяЕФЩјЭИадБЈЕРзюЙуЗКЃЌДЫЛњжЦжївЊЪЧЛљгкЦфжавЛИіПЙОњвЉЮяЖдЯИОњЯИАћФЄЭЈЭИадМАЙІФмЕФгАЯьЃЌЪЙСэвЛИіПЙОњвЉЮяНјШыЯИАћЕФаЇТЪЩ§ИпЛђвжжЦЯИАћФЄЩЯЭтХХБУЖдПЙОњвЉЮяЕФЭтХХзїгУЃЌНјЖјЬсИпЯИАћФкПЙОњвЉЮяЕФХЈЖШЃЌЬсЩ§ПЙОњаЇЙћЁЃAMPsгыПЙОњвЉЮяЕФСЊгУЖрВЩгУДЫЛњжЦЗЂЛгаЭЌПЙОњзїгУЁЃAMPsПЩвдИФБфЯИОњЯИАћФЄЕФЭЈЭИадЃЌДйНјПЙОњвЉЮяНјШыЯИОњЯИАћЃЌзїгУгкЯИАћФкАаЕуЃЌМћЭМЃГЁЃГ§ДЫжЎЭтЃЌСНжжВЛЭЌЕФПЙОњвЉЮяФмЙЛЭЈЙ§ЙВЭЌвжжЦФГвЛЩњЛЏЭООЖЃЌШчЕААзжЪКЯГЩЕШЖјдіЧППЙОњзїгУЁЃзюНќбаОПБэУїЃЌОпгаПЙбзКЭПЙбѕЛЏзїгУЕФЫЋЧтЧрняЫигыЭЗцппЛаСФмЙЛаЭЌвжжЦЕААзжЪЩњГЩЁЃаэЖрЯИОњЛЙФмЙЛВњЩњВЛЭЌжжРрЕФБЃЛЄадУИШчІТЊВФкѕЃАЗУИЃЌФмЙЛЫЎНтЧрУЙЫиЕШзїгУгкЯИОњЯИАћБкЕФІТЊВФкѕЃАЗРрПЙЩњЫиЃЌЕМжТДЫРрПЙЩњЫизїгУаЇЙћВЛМбЁЃІТЊВФкѕЃАЗУИвжжЦМСПЩвжжЦФЭвЉОњФкІТ-ФкѕЃАЗУИВњЩњЃЌЙЪСйДВЩЯГЃНЋІТ-ФкѕЃАЗУИвжжЦМС Шчrelebactamгы ІТ-ФкѕЃАЗРрПЙЩњЫиСЊгУЃЌвдгааЇБЃЛЄПЙЩњЫиВЛБЛЫЎНтЃЌНјЖјЪЙПЙЩњЫиПЙОњзїгУЗЂЛгжСзюДѓЁЃзїгУгкЯИОњЯИАћБкЕФЛЏКЯЮяГ§ІТ-ФкѕЃАЗРрПЙЩњЫиЭтЃЌЛЙгаЬЧыФРрЁЂСзУЙЫиЕШЁЃ2022ФъбаОПЗЂЯжЃЌЭЌЮЊЯИАћБкЛюадМСЕФбЧАЗХрФЯгыСзУЙЫиСЊгУФмЙЛЭЈЙ§ИЩШХЯИАћБкДњаЛВњЩњаЭЌзїгУЁЃ
Г§ЕЅвЛЯИОњздЩэФмЙЛЖдПЙОњвЉЮяВњЩњФЭвЉадЭтЃЌЯИОњ№ЄИНгкЮяЬхБэУцЃЌЭЈЙ§ЗжУкЖрЬЧЕШЮяжЪЪЙБЫДЫОлМЏдквЛЦ№ВЂНјвЛВНдіжГЃЌзюжеаЮГЩжТУмЕФОпгаЦСеЯзїгУЕФЩњЮяФЄЃЌЕжПЙПЙОњвЉЮяЕФзїгУЃЌЪЙЕУЯИОњЕФФЭвЉадДѓЗљЖШЬсИпЁЃвђДЫЃЌгабЇепНЋЖдЯИОњЩњФЄОпгаИпЖШЦЦЛЕзїгУЕФПЙОњвЉЮягыЦфЫћВЛОпгаЩњЮяФЄЦЦЛЕзїгУЛђЦЦЛЕзїгУНЯаЁЕФПЙОњвЉЮяСЊгУЃЌвдЦкФмЙЛЗЂЛгаЭЌПЙОњЛюадЁЃвбгабаОПЗЂЯжЃЌЃСЃЭЃаЃѓФмЙЛЦЦЛЕЯИОњЩњЮяФЄЃЌЕБ ЃСЃЭЃаЃѓгыПЙОњвЉЮяСЊКЯЪЙгУЪБЃЌЃСЃЭЃаЃѓЕФПЙЩњЮяФЄЬиадФмЙЛЬсИпПЙОњвЉЮяЕФПЙОњаЇЙћЁЃ
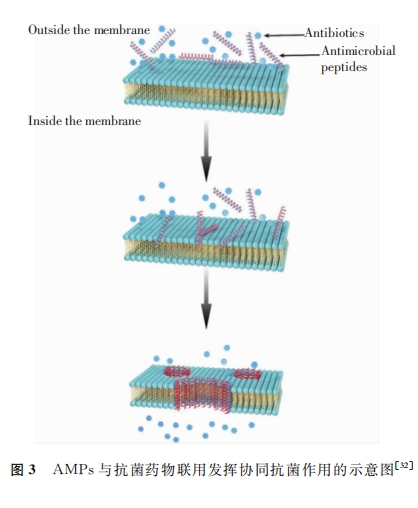
3.2 ДЋЭГПЙОњвЉЮяЕФСЊКЯгІгУ ДЋЭГЗНЗЈЕФЕЅвЛПЙОњвЉЮяжЮСЦЯИОњИаШОКмШнвзв§Ц№ФЭвЉЃЌЖјПЙОњвЉЮяСЊКЯгІгУЃЌМДЭЌЪБЪЙгУСНжжЛђСНжжвдЩЯЕФПЙОњвЉЮяЃЌЭЈЙ§зїгУгкЖрИіАаЕуЃЌДгЖјЬсИпПЙОњаЇЙћЃЌбгЛКФЭвЉадЕФЗЂеЙЃЌЪЧФПЧАЖдЯжгаДЋЭГПЙОњвЉЮядйРћгУЕФгааЇЗНЪНЁЃ
ПЙОњвЉЮяЕФаЭЌПЙОњЛюадПЩЭЈЙ§ЦхХЬЗЈВтЖЈСЊКЯгУвЉЪБПЙОњвЉЮяЕФзюЕЭвжОњХЈЖШМЦЫуВПЗжвжОњХЈЖШжИ Ъ§РДЦРМлЃЌЕБFIC indexЁм0.5ЪБЃЌХаЖЈЮЊаЭЌзїгУЃЛЕБ0.5ЃМFIC indexЃМЃДЪБЃЌХаЖЈЮЊЮоЯрЛЅзїгУЃЛЕБFIC indexЁнЃДЪБЃЌХаЖЈЮЊозПЙзїгУЁЃБЪепЧАЦкбаОПЗЂЯжЃЌНЋЭзВМУЙЫигыЬиЪтЙйФмЭХСЌНгЃЌЩшМЦЛёЕУЕФСНЧзадЭзВМУЙЫибмЩњЮяПЩвдЯджјЬсИпРћИЃЦНЛђЖўМзАЗЫФЛЗЫиЖдЖржиФЭвЉЭТЬМйЕЅАћОњЕФПЙОњЛюадЃЌБэЯжГіМЋЧПЕФаЭЌПЙОњзїгУЃЌЦфFIC indexДяЕН0.039ЁЃСНепЕФаЭЌЛњжЦба ОПЗЂ ЯжЃЌСН ЧзадЭзВМУЙЫибмЩњЮяФмЙЛгАЯьЯИАћФЄЩЯКєЮќСДЕФЕчзгДЋЕнНјЖјНЕЕЭ ЃСЃдЃа ЕФЩњГЩЃЌвжжЦЯИАћФЄЩЯПЙОњвЉЮяЭтХХБУЕФЙІФмЃЌНјЖјЗЂЛгаЭЌПЙОњзїгУЁЃЯИОњЯИАћФЄЩЯПЙОњвЉЮяЭтХХБУЕФЙ§БэДяЪЧКмЖрЯИОњВњЩњФЭвЉадЕФжївЊЛњжЦжЎвЛЃЌФПЧАбаОПЕФПЙОњвЉЮяЭтХХБУвжжЦМСШчЃБЃЃЈЃБЃнСМзЛљЃЉппрКЕШФмЙЛЛжИДФГаЉИяРМвѕадОњЖдПЙОњвЉЮяЕФУєИаадЃЌЕЋЖдгкЭЈЙ§ЯИАћБкЦСеЯзїгУВњЩњФкдДФЭвЉадЕФЯИОњгааЇадДѓДѓНЕЕЭЁЃвђДЫЃЌдіЧПЯИАћФЄЭЈЭИадЭЌЪБНЕЕЭПЙОњвЉЮяЭтХХБУЙІФмЪЧНтОіДЫЮЪЬтЕФРэЯыВпТдЁЃБОПЮЬтзщНЋЭзВМУЙЫизїЮЊдиЬхЃЌгыЃБЃЃЈЃБЃнСМзЛљЃЉппрКЕШВЛЭЌЭтХХБУвжжЦМСЯрСЌЃЌЛёЕУСЫвЛЯЕСаЛЏКЯЮяЃЌИУЛЏКЯЮягыДЋЭГПЙОњвЉЮяСЊКЯгІгУЃЌЯджјЬсИпСЫЗњрХЕЭЊРрПЙОњвЉЮяЛЗБћЩГаЧЁЂФЊЮїЩГаЧЖдФЭвЉадЭТЬМйЕЅАћОњЕФПЙОњЛюад.
Н№ЛЦЩЋЦЯЬбЧђОњЪЧвЛжжИяРМбєадЧђаЮЯИОњЃЌФмЙЛв§Ц№ОжВПИаШОЩѕжСЮЃМАЩњУќЃЌЭђЙХУЙЫиКЭЛЗБћЩГаЧЕШГЃБЛгУгкжЮСЦН№ЛЦЩЋЦЯЬбЧђОњИаШОЁЃЮЊБмУтЕЅЖРЪЙгУетаЉПЙОњвЉЮядкжЮСЦЙ§ГЬжаПЩФмв§ЗЂЕФФЭвЉадЮЪЬтЃЌKambleЕШЬсГіСЊКЯЪЙгУВЛЭЌПЙОњвЉЮяЃЌНЋДяЭаУЙЫиКЭЭђЙХУЙЫиетРреыЖдЯИАћНсЙЙЕФПЙЩњЫигыСэвЛжжзїгУгк ЃФЃЮЃС ИДжЦЛђЕААзжЪКЯГЩЙ§ГЬЕФПЙОњвЉЮяШчЛЗБћЩГаЧЛђЭзВМУЙЫиСНСНзщКЯЃЛЫљгазщКЯОљЯдЪОГіаЭЌаЇгІЃЌВЂвдМССПвРРЕЕФЗНЪНвжжЦКЭЦЦЛЕЩњЮяФЄЁЃгЩДЫПЩМћЃЌетаЉДЋЭГПЙОњвЉЮяСЊКЯЪЙгУБШЕЅЖРЪЙгУСНепжаШЮКЮвЛжжвЉЮяИќЮЊгааЇЃЌВЂгаЭћЛКНтЯИОњФЭвЉЮЪЬтЁЃ
3.3 AMPsгыДЋЭГПЙОњвЉЮяСЊКЯгІгУ MDRОњЕФГіЯжвдМАЫцКѓЪЙгУДЋЭГПЙОњвЉЮяжЮСЦЕФЪЇАмЃЌжњЭЦСЫаТаЭПЙОњвЉЮяЕФбаЗЂЁЃAMPsзїЮЊЩњЮяЬхЬьШЛУтвпЯЕЭГЕФзщГЩВПЗжЃЌМјгкЦфЙуЦзЕФПЙОњЛюадЁЂЮоВаСєЁЂЮоЮлШОЕШЬиЕуЃЌБЛШЯЮЊОпгаПЊЗЂЮЊаТаЭПЙОњвЉЮяЬцДњПЙЩњЫиЕФЧБжЪЁЃИќживЊЕФЪЧЃЌЦеБщШЯЮЊAMPsЭЈЙ§ЦЦЛЕЯИОњЕФЯИАћФЄНсЙЙЭъећадЗЂЛгПЙОњзїгУЃЌДЫжжвжОњЗНЪНВЛвзв§ЗЂФЭвЉадЃЌЭЌЪБвВЮЊСЊКЯгУвЉбаОПЬсЙЉСЫРэТлЛљДЁЁЃФПЧАЃЌЙигкAMPsгыДЋЭГПЙОњвЉЮяСЊКЯЪЙгУЕФбаОПвбгажюЖрБЈЕРЃЌЧвЦеБщШЯЮЊЖўепЕФаЭЌПЙОњЛњжЦЪЧAMPsЖдЯИОњЯИАћФЄЕФЦЦЛЕзїгУЃЌжТЪЙЯИАћФкПЙОњвЉЮяХЈЖШЩ§ИпЁЃБОПЮЬтЧАЦкбаОПЗЂЯжЃЌОпгаЯИОњЯИАћФЄЦЦЛЕЛюадЕФAMPsгыЖржжПЙОњвЉЮяСЊгУФмЙЛЬсИпПЙОњЛюадЃЌЧвЖдСйДВЗжРыЕФФЭвЉОњжъвВОпгаНЯКУЕФПЙОњЛюадЁЃШчИЛКЌИЌАБЫсЕФжЌыФ ЃУЃБЃВ-ЃаЃвЃадкгыЖўМзАЗЫФЛЗЫиЛђРћИЃЦНСЊКЯгІгУЪБЃЌПЩЯджјЬсИпЖўепЖд ЃЭЃФЃвКЭЙуЦзФЭвЉЭТЬМйЕЅАћОњЕФПЙОњЛюадЃЌжЌыФЃУЃБЃВ-ЃаЃвЃаОпгаНјвЛВНПЊЗЂЮЊПЙОњвЉЮязєМСЕФЧБжЪЁЃДЫЭтЃЌгабаОПЗЂЯжЃЌAMPsгыЭЌбљзїгУгкЯИОњЯИАћФЄЕФПЙОњвЉЮяСЊКЯгІгУЪБвВПЩвдВњЩњаЭЌПЙОњаЇЙћЁЃFiЕШбаОПЗЂЯжЃЌAMPsNisinгыПЙЩњЫиЖр№ЄОњЫи ЃХ СЊКЯгІгУЪБЃЌAMPsNisinДѓЗљНЕЕЭЖр№ЄОњЫи ЃХ вжжЦЯИОњЩњЮяФЄЫљашЕФвЉЮяХЈЖШЃЌЬсИпПЙЩњЫиЕФПЙОњаЇТЪКЭЯИОњЖдПЙЩњЫиЕФУєИаадЁЃJaЕШбаОПБэУїЃЌAMPsNisinКЭЖр№ЄОњЫи ЃХСЊКЯгІгУЖдЙуЦзФЭвЉаЭБЋТќВЛЖЏИЫОњКЭЖр№ЄОњЫи ЃХ ФЭвЉадЭТЬМйЕЅАћОњОљБэЯжГіаЭЌПЙОњЛюадЁЃ
ВПЗжAMPsДцдквЛЖЈЯИАћЖОадЧввзБЛЕААзУИЫЎНтЃЌвВЪЧжЦдМЦфдкСйДВжагІгУЕФгжвЛживЊдвђЁЃФПЧАЃЌЮЊСЫзюДѓЯоЖШЕиЬсИпAMPsЖдВЛЭЌЯИАћЕФбЁдёадЃЌвЛаЉбаОПЪдЭМЭЈЙ§ађСааоЪЮЁЂыФСДдгКЯКЭАБЛљЫсЬцДњЕШЗНЗЈЖдAMPsНјаааоЪЮЛђИФдьЁЃAamAP1-LysineЪЧвдаЋЖОжаЗЂЯжЕФAMPsAamAP1зїЮЊЛљДЁЩшМЦЛёЕУЕФвЛжжКЯГЩыФбмЩњЮяЃЌЦфгызѓбѕЗњЩГаЧЁЂТШУЙЫиЁЂРћИЃЦНЁЂКьУЙЫиСЊКЯгІгУЪБЃЌЖдН№ЛЦЩЋЦЯЬбЧђОњКЭЭТЬМйЕЅАћОњБэЯжГіВЛЭЌГЬЖШЕФаЭЌПЙОњзїгУЃЌЧвдквЛаЉзщКЯжаAamAP1-LysineЕФзюаЁвжОњХЈЖШЩѕжСДгЃГІЬЃэЃяЃьЃЏЃЬНЕЕЭжСЃАЃЎЃАЃБІЬЃэЃяЃьЃЏLЁЃAlmaaytahЕШЃлЃЕЃБЃнЖд AamAP1-LysineгыРћИЃЦНСЊгУЕФПЙЩњЮяФЄЛюадбаОПжаЃЌВтЕУFICindexЕЭжСЃАЃЎЃАЃЕЃИЃЌБэЯжГіЧПаЭЌзїгУЁЃЃФЃѕЃсЃю ЕШба ОП вВ ЗЂ ЯжЃЌРД дД гк Ап Тэ гу ЕФAMPsбмЩњыФ ЃаЃєЃЕ-ЃБЃугыПЙЩњЫиСЊгУКѓДйНјПЙЩњЫиНјШыЯИОњЯИАћЃЌДгЖјЖд ЃЭЃФЃв ОњВњЩњаЭЌПЙОњаЇгІвдМАПЙЩњЮяФЄаЇгІЁЃ
3.4 AMPsгыЦфЫћЛЏКЯЮяЕФаЭЌПЙОњзїгУ Г§ДЋЭГПЙОњвЉЮяЭтЃЌФГаЉ AMPsгыЦфЫћЛЏКЯЮявВПЩВњЩњаЭЌПЙОњзїгУЁЃШчЗфдД AMPsMelittinЪЧвЛжжбаОПБШНЯЭИГЙЕФЫЎШмадКУЁЂЯЕЭГЖОадИпЕФAMPsЁЃMelittinФмЙЛЦЦЛЕЯИОњЯИАћФЄЃЌОпгаЧППЙОњЛюадЃлЃЕЃГЃнЁЃЫящЊЕШЃлЃЕЃДЃнбаОПЗЂЯжЃЌMelittinгыФћУЪЫсСЊКЯгІгУЪБЖдДѓГІАЃЯЃОњЁЂГІбзЩГУХОњМАН№ЛЦЩЋЦЯЬбЧђОњОљЯдЪОГіаЭЌПЙОњаЇгІЃЌЕБгыввЫсСЊКЯгІгУЪБЖдГІбзЩГУХОњБэЯжЮЊаЭЌПЙОњзїгУЁЃЮЂЩњЮядДAMPsЪЧгЩЮЂЩњЮяЗжУкЕФгУРДБЃЛЄздЩэВЂЩБУ№ЛђвжжЦЦфЫћЮЂЩњЮяЕФаЁЗжзгЖрыФЃЌЭЈГЃОпгаЙуЦзПЙОњЛюадЃЌШчРћгУВрцпЖЬбПцпИЫОњВњЩњЕФAMPsBrevilaterinЁЃФўбЧЮЌЕШбаОПжЄЪЕЃЌAMPsгыФћУЪЫсСНепаЭЌвжжЦДѓГІАЃЯЃОњЕФЩњГЄЃЌЦЦЛЕЯИОњЯИАћФЄЕФЭъећадЃЌЕМжТЯИАћБфаЮЁЂЃФЃЮЃС НЕНтМАЯИАћФкШнЮяаЙТЉЁЃФћУЪЫсФмЙЛгааЇвжжЦгаЛњЫсЕФНЕНтЃЌбгЛКЪГЮяИЏАмЃЌЭЌЪБAMPsгыФћУЪЫсСЊКЯгІгУНЕЕЭСЫЕЅИіПЙОњвЉЮяЕФЪЙгУСПЃЌгаЭћдкЪГЦЗБЃЯЪжаЭЦЙугІгУЁЃ
ЃД AMPsгыДЋЭГПЙОњвЉЮяСЊКЯгІгУНјеЙ
ЃД.1 ЪГЦЗЗРИЏ AMPsгУзїЗРИЏМСЃЌОпгаШШЮШЖЈадКУЁЂАВШЋЮоЖОЁЂПЙОњадЧПЕШгХЕуЃЌвђДЫдкЪГЦЗЕФЗРИЏКЭБЃЯЪжаОпгаЙуРЋЕФгІгУЧАОАЁЃ AMPsNisinдкШЫЬхФкВЛЛсГЄЦкЛ§РлЃЌЖОадаЁЃЌзїЮЊЪГЦЗЗРИЏМСОпгаКмИпЕФАВШЋадЃЌЧвдкФГаЉЕиЧјвбОБЛХњзМдкЪГЦЗжќВижаЪЙгУЁЃMirhoseiniЕШбаОПЗЂЯжЃЌNisinгыбѕЛЏУОФЩУзПХСЃОпгааЭЌПЙОњаЇгІЃЌгІгУгкХЃФЬЕФЩБОњУ№ЛюжаЃЌФмЙЛНЕЕЭХЃФЬжаДѓГІАЃЯЃОњКЭН№ЛЦЩЋЦЯЬбЧђОњЕФОњШКЪ§СПЁЃNisinгыЯуЧлЗгЕФСЊгУЭЌбљФмвжжЦХЃФЬжаЯИОњЗБжГЃЌгаЭћГЩЮЊФЬжЦЦЗЕФБЃЯЪВпТдЁЃДЫЭтЃЌЃЬЃщЕШЃлЃЖЃБЃнНЋІТ-ЛЗК§ОЋгыNisinСЊКЯгІгУЃЌгааЇЬсИпСЫNisinдквШЕААзУИжаЕФЮШЖЈадЃЌбгЛКNisinЕФНЕНтЃЌЯджјвжжЦЕЭЪьжэШтжаЯИОњЕФЩњГЄЗБжГЃЌЪЙжэШтБЃГждгагЊбјМлжЕКЭЖРЬиЕФЗчЮЖЁЃ
4.2 вНвЉЮРЩњ ФПЧАЃЌгЩMDRОњв§Ц№ЕФвНдКЛёЕУадИаШОбЯжиЮЃКІЙЋЙВЮРЩњАВШЋЃЌЦШЧаашвЊбаЗЂаТЕФПЙОњВпТдЙЅПЫЯИОњФЭвЉадЁЃAMPsОпгаЖржжЩњЮябЇЙІФмЃЌдкЯШЬьУтвпЯЕЭГжаЗЂЛгживЊзїгУЁЃдкAMPsСйДВгІгУВпТдДДаТбаОПжаЃЌSaloutiЕШЗЂЯжЃЌЕБAMPsMBP-1гывјФЩУзСЃзгСЊКЯЪЙгУЪБЃЌЯджјНЕЕЭН№ЛЦЩЋЦЯЬбЧђОњЕФЖЈжВТЪЃЌЬсИпЩЫПкгњКЯТЪЁЃПЙОњвЉЮяСЊКЯгІгУдкЬхФкЕФаЭЌПЙОњЛюадвВЕУЕНжЄЪЕЁЃеХмАдЊЕШбаОПЗЂЯжЃЌAMPs LL-37КЭѕЃАЗУИСЊКЯЪЙгУвжжЦИаШОЗрГІЧђОњаЁЪѓЬхФкдрЦїжаЯИОњЕФдіжГЃЌМѕЧсдрЦїВЁРэЫ№ЩЫЁЃЬНЫїЦфаЭЌЛњжЦЗЂЯжЃЌѕЃАЗУИЯШЦЦЛЕЯИОњЯИАћБкЃЌгаРћгкAMPs LL-37НгДЅЕНЯИОњЯИАћФЄЃЌLL-37ВЛНіФмЦЦЛЕЯИАћФЄНсЙЙв§ОњЬхЯИАћФкШнЮяЭтаЙЃЌЛЙФмЙЛгыЯИАћФкЕФЕААзжЪЁЂКЫЫсЯрНсКЯЃЌгАЯьЯИАћЕФЩњУќЛюЖЏЃЌСНЛњжЦЯрИЈЯрГЩгааЇЬсИпЖдЗрГІЧђОњЕФЩБЩЫаЇТЪЁЃгыДЫЭЌЪБЃЌИУЗНЗЈНЕЕЭѕЃАЗУИЕФЪЙгУСПЃЌвжжЦЯИОњФЭвЉадЕФВњЩњЁЃзюНќбаОПЗЂЯжЁЃLL-37гыЭђЙХУЙЫиЁЂАЂЦцУЙЫиЁЂЖр№ЄОњЫиЕШПЙЩњЫиСЊКЯЪЙгУгаЭћГЩЮЊжЮСЦЭТЬМйЕЅАћОњИаШОЕФаТВпТдЁЃДЫЭтЃЌAMPs LP-23ЁЂDP-23КЭSPOгыЧьДѓУЙЫиСЊгУЖдСйДВЗжРыЕФФЭЧьДѓУЙЫиЕФДѓГІАЃЯЃОњБэЯжГіаЭЌПЙОњзїгУЃЌНјвЛВНжЄЪЕСЫAMPsгыПЙОњвЉЮяСЊгУдкжЮСЦФЭвЉОњжъв§Ц№ЕФЯИОњИаШОЗНУцОпгаКмДѓЧБСІЁЃ
4.3 ЖЏЮябјжГ дкЖЏЮябјжГжаЃЌAMPsПЩгУгкИФЩЦЖЏЮяНЁПЕзДПіЃЌДйНјЖЏЮяЩњГЄЃЌЬсИпЩњГЄадФмЁЃЭѕРђЕШбаОПжЄЪЕЃЌЬьВЯЫиКЭІТ-ИЪТЖОлЬЧУИЖд№Ц№ШЕФЩњГЄгааЭЌДйНјзїгУЃЌвЛжжКЯРэЕФНтЪЭЪЧІТ-ИЪТЖОлЬЧУИПЩНЕНтИЪТЖОлЬЧГЩЮЊИЪТЖЙбЬЧЃЌЖјИЪТЖЙбЬЧКЭAMPsОљПЩДЬМЄЛњЬхУтвпЯЕЭГЯИАћЃЌМѕЩйГІЕРгаКІОњЃЌЬсИпШедіжиЃЌНЕЕЭСЯШтБШЁЃдкШтМІШеСИжаСЊКЯЬэМгЬьВЯЫиКЭКЯЩњЫиИФЩЦСЫШтМІаЁГІ№ЄФЄНсЙЙЃЌДйНјаЁГІ№ЄФЄУтвпЯИАћдіжГЁЃСэвЛИіБЛЙуЗКбаОПЕФжэдДAMPs PR-39гыАЂФЊЮїСжЁЂЭСУЙЫиЁЂСђЫсСДУЙЫиСЊКЯгІгУЃЌеыЖдаѓЧнГЃМћВЁдОњБэЯжГіУїЯдаЭЌПЙОњзїгУЁЃ
ЃЕ аЁНсгыеЙЭћ
ЯИОњГігкздЮвБЃЛЄЕФдвђЖдЯжгаПЙОњвЉЮяВњЩњФЭвЉЃЌЧвЫцзХЯИОњФЭвЉЕФВЛЖЯЗЂеЙЖдШЫРрНЁПЕЕФЭўаВШевцМгОчЁЃБОЮФзлЪіЛљгкВЛЭЌПЙОњЛњжЦЕФПЙОњвЉЮяСЊКЯгІгУЪЧдкВЛПЊЗЂаТвЉЕФЧАЬсЯТЃЌзюОМУИпаЇЕФвжжЦЯИОњФЭвЉадЃЌжЮСЦСйДВЯИОњИаШОЕФВпТджЎвЛЁЃдкжюЖрПЙОњвЉЮяСЊКЯгІгУбаОПжаЃЌAMPsгыДЋЭГПЙОњвЉЮяаЭЌПЙОњГЩЮЊСЫбаОПШШЕуЃЌЧвгаЭћЙЅПЫЯИОњФЭвЉадЁЃЕЋПЙОњвЉЮяСЊКЯгІгУЖдгкВЛЭЌЯИОњЕФзюМбзщКЯЁЂСйДВПЙОњаЇЙћЃЌвдМАЪЧЗёШдДцдкв§ЗЂФЭвЉадЕФЗчЯеЕШЮЪЬтШдашЩюШыбаОПЃЌвдЦкПЙОњвЉЮяаЭЌПЙОњВпТддкЪГЦЗЁЂвНСЦЁЂаѓФСЕШСьгђЕУЕНИќЙуЗКЕФгІгУЁЃ
Утд№ЩљУїЃКБОЮФЮЊаавЕНЛСїбЇЯАЃЌАцШЈЙщдзїепМАддгжОЫљгаЃЌШчгаЧжШЈЃЌПЩСЊЯЕЩОГ§ЁЃЮФеТБъзЂгазїепМАЮФеТГіДІЃЌШчашдФЖСдЮФМАВЮПМЮФЯзЃЌПЩдФЖСддгжО
